signal transduction mutants of arabidopsis uncouple...
TRANSCRIPT

Cell, Vol. 74, 787-799, September IO, 1993, Copyright 0 1993 by Cell Press
Signal Transduction Mutants of Arabidopsis Uncouple Nuclear CAB and RBCS Gene Expression from Chloroplast Development Ronald E. Susek; Frederick M. Ausubel,t and Joanne Chory’ *Plant Biology Laboratory The Salk Institute for Biological Studies San Diego, California 92186-5800 tDepartment of Genetics Harvard Medical School and Department of Molecular Biology Massachusetts General Hospital Boston, Massachusetts 02114
Summary
Chloroplast development requires coordinate nuclear and chloroplast gene expression. A putative signal from the chloroplast couples the transcription of cer- tain nuclear genes encoding photosynthesis-related proteins with chloroplast function. We have identified at least three Arabidopsis nuclear genes (GUNl, GUNP, and GUN3) necessary for coupling the expres- sion of some nuclear genes to the functional state of thechloroplast. Homozygousrecessivegun mutations allow nuclear gene expression in the absence of chlo- roplast development and furthermore may interfere with the switch from dark-grown to light-grown devel- opment. Other reports suggest this intracellular cross- talk also involves mitochondrial interactions. The GUN genes thus define steps in one specific branch of a complex interorganellar regulatory network.
Introduction
The cells of multicellular plants and animals differentiate into a wide variety of specialized cell types with distinct structures and functions. In higher plants, plastids (chloro- plasts are one type of differentiated plastid) assume char- acteristic structures that are consistent with their organel- lar functions (Kirk and Tilney-Basset, 1978; Whittaker and Danks, 1978). In the photosynthetic mesophyll Cells of a leaf, proplastidsdevelop intochloroplasts. In contrast, pro- plastids differentiate into starch-containing plastids (amy- loplasts) in root cortical cells. In dark-grown (etiolated) plants, the mesophyll cell plastids develop into etioplasts, characterized by their central array of paracrystalline tu- bules (i.e., prolamellar bodies). When dark-grown plants are transferred to the light, the etioplasts differentiate into chloroplasts. Thus, both environmental signals, such as light, and intrinsic signals determine the developmental fate of the undifferentiated proplastid.
There is a complex interplay of regulatory signals be- tween the nucleus and cytoplasmic organelles during de- velopment. The nuclear genome encodes both structural and regulatory proteins required for organellar develop- ment. Mutations of either class of nuclear gene can inter- fere with organelle function or biogenesis (RBdei, 1973; Taylor et al., 1987; Tzagoloff and Dieckmann, 1990). For
example, mutations of the nuclear DE T and COP genes of Arabidopsis thaliana cause dark-grown plants to develop leaves and chloroplasts and to express light-regulated genes inappropriately, as if they were grown in the light (Chory et al., 1989a; Chory and Peto, 1990; Chory, 1991; Deng and Quail, 1992). A second class of Arabidopsis genes, the HY genes, plays a critical role in light percep- tion. hy mutants show reduced light responses and incom- plete chloroplast development, with reduced chlorophyll accumulation and less extensive grana thylakoid mem- brane stacking (Chory et al., 1989b). Thus, although a plastid contains itsown genome, the nucleus plays a major role in determining the developmental fate of plastids (Kirk and Tilney-Basset, 1978; Whittaker and Danks, 1978).
Furthermore, organelles signal as well as respond to the nucleus. In yeast, for example, molecular oxygen and heme, synthesized in mitochondria, regulate the transcrip- tion of nuclear genes that encode mitochondrial proteins (Forsburg and Guarente, 1989). Similarly, as undifferenti- ated plastids develop into photosynthetically competent chloroplasts, plants coordinately express many nuclear and plastid genes encoding structural components of the photosynthetic apparatus (Kendrick and Kronenber, 1986). Genetic or environmental conditions that prevent chloroplast development specifically block the expression of several nuclear genes, such as CAB and RBCS (encod- ing the chlorophyll a/b-binding protein of the photosystem II light harvesting complex and the small subunit of ribu- lose bisphosphate carboxylase, respectively), that encode chloroplast proteins associated with photosynthetic func- tion (Harpster et al., 1984; Mayfield and Taylor, 1984). For example, in bright light, carotenoid-deficient plants suffer severe free radical photooxidation. This results in acoordi- nate loss of both chloroplast function and CAB transcrip- tion in the nucleus (Oelmtiller, 1989; Susek and Chory, 1992). The accumulation of nuclear-encoded transcripts for nitrate reductase and hydroxypyruvate reductase is also sensitive to photobleaching, even though they en- code nonchloroplast proteins with functions related to pho- tosynthesis (OelmDller and Briggs, 1990; Schwartz et al., 1991). In other studies, nitrogen starvation or the inhibition of plastid transcription or translation from the time of ger- mination blocks CAB messenger RNA (mRNA) accumula- tion (Oelmiiller et al., 1986; Taylor et al., 1987; Plumley and Schmidt, 1989; Taylor, 1989; Mathews and Durbin, 1990; Rapp and Mullet, 1991). In contrast, less severe chloroplast lesions, such asachlorophyll biosyntheticdefi- ciency or a chloroplast translational block in green plants, do not block CAB expression (Harpster et al., 1984; Oel- miiller et al., 1986).
Thus, it has been hypothesized that signals from both mitochondria and chloroplasts regulate nuclear gene tran- scription (Forsburg and Guarente, 1989; Oelmtiller, 1989; Taylor, 1989). In plants, this proposed signal transduction pathway coordinates the expression of a subset of nuclear genes with chloroplast genes during chloroplast biogene- sis, maintenance, and senescence (Taylor et al., 1986;

Cell 788
Taylor, 1989). To date, the molecular nature of the signal and its mechanism of transduction to the nuclear transcrip- tional machinery are unknown. One possibility is that this signal transduction pathway in plants resembles the path- ways in yeast in which mitochondrial function and heme modulate nuclear gene expression (Forsburg and Guar- ente, 1989; Susek and Chory, 1992; Liao and Butow, 1993).
If a signal transduction pathway between the chloroplast and nucleus exists, then it should be possible to generate mutations that disrupt this pathway and uncouple nuclear gene expression from plastid function. We designed a mu- tant screen to identify rare mutant plants that express transgenic CAB reporter gene fusions following growth under conditions that prevent chloroplast development. We used the Arabidopsis CAB3 promoter because its ex- pression pattern has been extensively characterized. CAB expression is light dependent, is restricted to green, chlo- roplast-containing cells, is subject to circadian control, and is initiated early in chloroplast development (Martineau and Taylor, 1986; Gilmartin et al., 1990; Millar and Kay, 1991). The CA6 promoters contain redundant control ele- ments with identical or tightly linked light-responsive and tissue-specific regulatory elements that interact with a host of transcription factors (Schindler and Cashmore, 1990). Full regulation of the Arabidopsis CAB genes requires only 209 nt of promoter sequence, although maximal levels of expression require additional upstream sequences (Mitra et al., 1989; An, 1987; Ha and An, 1988). Of the nuclear genes that respond to the state of the chloroplast in higher plants, only the CAB genes show chloroplast-dependent expression in all plants examined (Taylor, 1989; Susek and Chory, 1992). Furthermore, much of the effect of the chloroplasts on CAB expression is transcriptional (Bat- schauer et al., 1986; Simpson et al., 1986; Burgess and Taylor, 1987).
We used a transgenic line of Arabidopsis that carries CAB3 reporter genes to identify mutations in at least three genes that play a role in coupling nuclear CAB expression to chloroplast function. We designate these genes GUN7, GUNP, and GUN3 (for genomes uncoupled) to indicate the mutant phenotype of uncoupled nuclear and plastid gene expression. These mutants support the existence of a sig- nal transduction pathway between the chloroplast and nu- cleus. In addition, gun7 mutants are defective in the switch from dark growth to light growth, suggesting that GUN7 also modulates the coordinate changes in the nucleus and chloroplast during early stages of photosynthetic growth. With this novel class of mutants, we can begin to charac- terize this elusive interorganellar signal transduction pathway.
Results
Construction of a Transgenic Line to Identify Mutants We constructed a transgenic line of Arabidopsis in which the full-length CAB3 promoter controls the expression of two different reporter genes (Figure 1). These reporter genes provide rapid and reproducible assays for CAB3
promoter activity. The double transgene approach should help us distinguish cis-acting mutations in the transgenic construct from trans-acting signal transduction mutations. We pursued only those candidate signal transduction mu- tant plants that inappropriately expressed both transgenes. The transgenic construct, pOCA107 (Figure l), contained a 950 bp CAB3 promoter (An, 1987), fused at -9 (with respect to the CAB translation start site) both to the hph gene (encoding hygromycin phosphotransferase), which confers resistance to the antibiotic hygromycin (Gritz and Davies, 1983), and to the uidA gene (encoding /3-glu- curonidase [GUS]), which converts commercially avail- able substrates into colored or f luorescent products (Jef- ferson et al., 1986). We transformed the pOCA107 construct into the A. thaliana Columbia ecotype and se- lected transformants by their resistance to kanamycin (the npN/gene expressed from the constitutive 35s cauliflower mosaic virus promoter). We chose to focus on one trans- formed line, pOCA107-2, with a single pOCA107 insertion site that maps to chromosome II, 3.0 CM from HY7 (L. Altschmied and J. C., unpublished data). As expected, the transgenes in pOCAl07-2 show light-dependent, tis- sue-specific, and chloroplast-dependent expression (Ta- ble l), as well as the proper transcript 5’ end (data not shown).
Isolation of gun Mutants We screened for mutants of the pOCA107-2 line that ex- pressed both of the CA63 promoter-driven transgenes in the absence of chloroplast development. Figure 2 summa- rizes the screen. We treated pOCA107-2 transgenic seeds with the chemical mutagen ethyl methanesulfonate and allowed plants (Ml) to self-fertilize (Chory et al., 1989a, 1989b). To block chloroplast development in M2 seed- lings, we treated plants with the herbicide Norflurazon (4-chloro-5-(methylamino)-2-[3-(trif luoromethyl)phenyl]-3- (2H)-pyridazinone), a noncompetit ive inhibitor of the carot- enoid biosynthetic enzyme phytoene desaturase (Chamo- vitz et al., 1991). In the absence of carotenoids, chlorophyll absorbs light to form excited electronic triplet states that react with molecular oxygen to yield free radicals. Thus, without carotenoids, plastids may initiate chloroplast bio-
pocp.107
Figure 1. Diagram of the Transgenic Construct Used to Select Mu- tants The chimeric gene construction, pOCA107. was inserted into an Agro- bacterium tumefaciens Ti plasmid-derived binary vector and then transformed into A. thaliana. DNA transfer initiates at the right border (RB) of the vector and proceeds to the lefi border (LB). Transformed plants were selected by their resistance to the drug kanamyctn. The structural gene for kanamycin resistance (nptll) is driven by the strong, constitutive cauliflower mosaic virus promoter (35s). The 3’ poly(A) site from the Agrobacterium Ti plasmid nopalme synthase gene (nos) aids transcript processing. CA63 is the promoter from the light- inducible Arabidopsis CAB3 gene, encoding the photosystem II light harvesting complex. It drives the expression of hph (for hygromycin resistance) and of uidA (for GUS). Arrows indicate direction of tran- scription.

U$?upling Nuclear and Chloroplast Gene Expression
Table 1. CA63-GUS Transgene Expression in pOCA107-2 Screen For Mutants
Tissue GUS Activity Ratio
Light 43,000 120 Dark 370
Leaves 32,000 71 Roots 450
Minus Norflurazon 33,000 550 Pius Norflurazon 60
The &A gene encodes GUS. GUS activity is reported in picomoles of 4-methylumbelliferone per minute per mill igram of protein. The pat- terns of transgene and endogenous CAB gene expression are identi- cal. Of particular note is that IO-day-old green seedlings but not Norflur- azon-photobleached seedlings express CABB-MA.
genesis but are rapidly photooxidized or photobleached (Oelmtiller, 1989).
We first screened Norf lurazon-photobleached M2 plants for CABShph transgene expression by growing them for 7-10 days in the presence of 20 ug/ml hygromycin. Puta- tive mutants were large when grown on hygromycin com- pared with the wild-type pOCAl07-2 and nontransgenic lines (Figure 3). For a positive control, we included pHG1, a line of Arabidopsis carrying a transgenic construct simi- lar to pOCAlO7 but in which PAL7 (phenyalanine ammonia lyase) promoters (not affected by Norflurazon treatment) replace the CAB3 promoters (S. Ohl, B. Kraft, J. C., and C. J. Lamb, unpublished data; see also Experimental Pro- cedures) (Figure 3). Retarded root development in the mu- tants relative to the pHG1 control suggested that the CABS-hph transgene expression was confined to the leaves, the normal pattern of CAB expression.
We then performed a secondary screen to assay for the expression of the second CAB3 promoter in Norflurazon- grown plants. A cotyledon was removed from each puta- tive mutant and assayed for CAB34dA gene expression using a histochemical stain for GUS activity (Jefferson et al., 1986). Cotyledons from the putative mutants, but not from the wild-type pOCAl07-2 plants, stained blue (Figure 48). In the absence of Norflurazon, both mutant and wild- type cotyledons stained equally blue (Figure 4A).
We screened 100,000 M2 plants and found 227 photo- bleached plants that appeared hygromycin resistant and that showed GUS histochemical staining. Of these, 138 recovered and were fertile. We assayed their M3 and M4 generation progeny and found 12 independent mutants with heritable mutant phenotypes. The results presented below show that nine of these mutants accumulated en- dogenous CAB mRNA following growth on Norflurazon. We call these mutants gun (for genomes uncoupled). These mutations appear to be specific for this interorganel- lar signal transduction pathway because other aspects of CAB promoter regulation are unaffected in the mutants (see below).
We have begun a genetic characterization of the gun mutants to assess the complexity of the putative signaling pathway coupling nuclear and chloroplast gene expres- sion. Each gun mutant represents an independent muta- tional event, since each originated from a different M2
Harvest 62 seeds
0 Screen M2 seedhngs on
Norflurazon + Hygromycm B A
Screen for hygromycm r&tant (i e large) plants (CAB-hph tramg ne expreswon)
6 Screen for GUS actwty
(CAB-uidA tram ene expression) 8 Recover and propagate putatwe mutants
0 Assay progeny for gun phenotype
Figure 2. Steps in Mutant Screen
EMS, ethyl methanesulfonate.
Figure 3. CABS-hph Transgene Expression in Control and gun7 Mu- tant Plants Wild-type Columbia (Col-0), transgenic wild type (pOCA107-2) a posi- tive control for growth on hygromycin (pHGl), and gun7 mutants (gonl-1, gunl-2, and gun73) were germinated and grown for 7 days in the presence of Norflurazon with or without hygromycin B. (a) In the absence of hygromycin B, all photobleached plants are the same size. (b) In the presence of hygromycin B, the gun7 mutants and PHG7 appear large owing to their hygromycin resistance. In contrast, the hygromycin-sensitive pOCAl07-2 and Columbia plants are small. Scale bar, 1 mm.
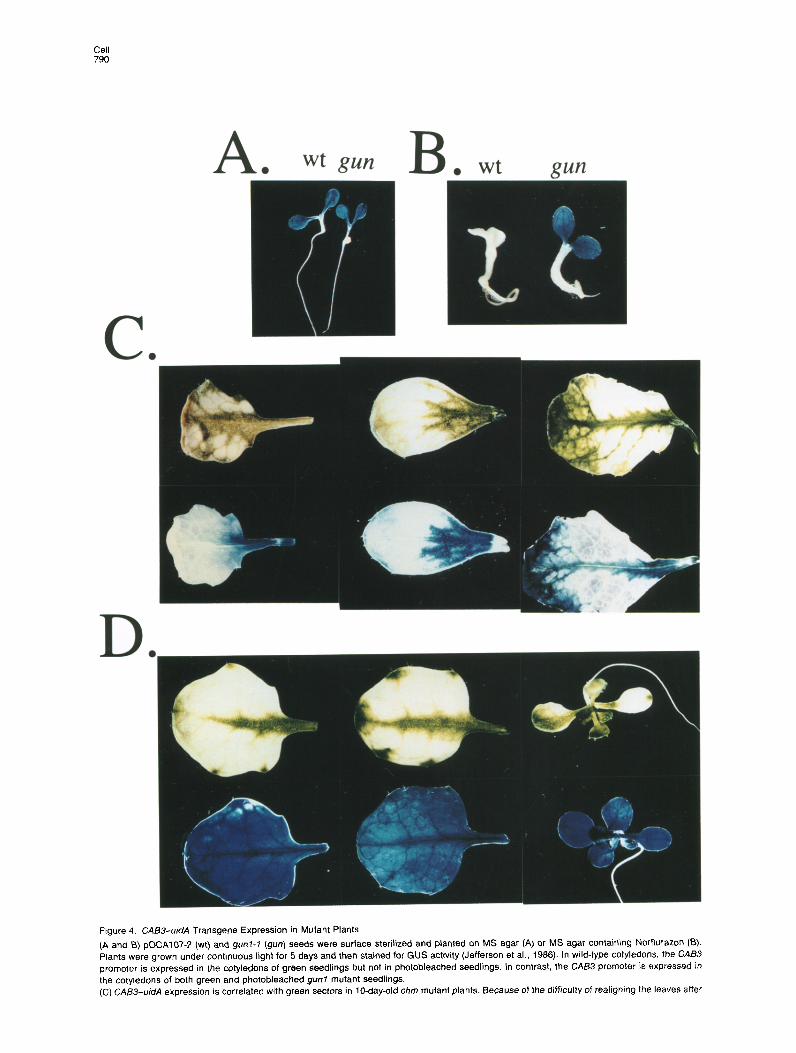
Cell 790
D
Frgure 4. (A and B) Plants we, promoter the cotyk (C) CABS-
CABS-&A Transgene Expression in Mutant Plants pOCAI07-2 (wt) and gunf-7 (gun) seeds were surface sterilized and planted on MS agar (A) or MS agar containmg Norflurazon (B).
re grown under continuous light for 5 days and then stained for GUS activity (Jefferson et al., 1986). In wild-type cotyledons, the CAB3 is expressed in the cotyledons of green seedlings but not in photobleached seedlings. In contrast, the CAB3 promoter is expressed in dons of both green and photobleached gun7 mutant seedlings, -uidA expression is correlated with green sectors in lo-day-old chm mutant plants. Because of the difficulty of realigning the leaves after

Uncoupling Nuclear and Chloroplast Gene Expression 791
Table 2. Genetic Analysis of gun Mutants Table 3. Genetic Complementation of gun Mutants
Cross
Percent Gener- Wild Percent ation Total Type Mutant x2
Wild type x gunl-l Fl F2
pOCA107-2 x gunI-l F2
gunl-1 x pOCAl07-2 Fl
Wild type x guni- Fl F2
pOCAl07-2 x gunl-3 Fl F2
pOCAl07-2 Y gunl-4 Fl F2
pOCAl07-2 x gun2-7 Fl F2
pOCA107-2 x gunO-10 Fl F2
pOCAl07-2 x gunO-34 Fl F2
pOCAl07-2 x gun@6 F2
pOCA107-2 x gun@38 F2
4 100 0 234 86 14
116 74 26
5 100 0
15 100 0 20 95 5
7 100 0 96 61 19
17 94 6 277 72 28
16 94 6 219 76 24
a 100 0 63 67 33
6 100 0 113 67 33
351 71 29
131 78 22
3.3
0.046
2.5
2.0
1.5
0.075
2.4
3.6
3.1
0.57
Each cross is indicated as female parent crossed with male parent. The gun mutants were crossed with the wild-type Columbia ecotype (wild type) or the wild-type transgenic line pOCA107-2. Fl and F2 plants were analyzed for the gun phenotype (GUS activity following growth on Norflurazon). For a recessive mutant crossed with pOCAl07-2, we expect a wild-type phenotype in the Fl generation and I:3 segregation of mutant phenotype in the F2 generation. For mutants crossed to wild-type Columbia, both the mutation and transgene segregate, such that we predict 3:16 segregation of the mutant phenotype in the F2 generation. ~‘valuesindicate theobserved deviation from the predicted segregation ratios. A value of <3.8 is acceptable (p > 0.05).
seed pool. The mutants were backcrossed and the mutant phenotype assayed in the Fl and F2 progeny. The segre- gation ratios of mutant to wild-type phenotypes suggest that each of the nine mutant phenotypes results from a monogenic recessive nuclear mutation (Table 2). We have not completed the analysis for a tenth isolate, gunO-4.
Complementation analysis indicates that four of the re- cessive mutations are allelic. We designate this gene as GUN7 and the four independent mutant alleles as gunl-7, gunl-2, gun73, and gunl-4 (Table 3). The mutation gun2-7 complements the gun7 mutations, suggesting that these mutations affect different genes. The low fertility of several mutant lines prevented us from completing the complementation analysis. We designate these remaining mutants as gun0. Two of these, gun0-70 and gonO-34, are neither allelic to one another nor to gunl. Therefore, the
Percent Wild Percent
cross Generation Total Type Mutant
gunl-1 x gunl-2 Fl 7 0 100 gunl-2 x gunl-l Fi ia 0 100 gunl-3 x gunl-7 Fl 2 0 100 gunl-4 x gunl-l Fl 1 0 100 gunl-1 x gunl-4 Fl 15 0 100 gunl-1 x gun2-1 Fl 27 100 0 gunO-10 x gunl-3 Fl a 100 0 gunl-4 x gun@34 Fl 4 100 0 gunO-10 x gun@34 Fl 28 100 0
The results of complementation crosses are presented as for Table 2.
nine recessive mutations represent lesions in three to six different GUN genes.
gun1 Mutations Affect the Accumulation of Both CAB and RBCS mRNAs Signal transduction mutants should express both the CA63 promoter-driven transgenes, as well as the endoge- nous CA6 genes, despite photobleaching. To test this pre- diction, we performed Northern hybridization analysis on total RNA extracted from wild-type plants and from mutant seedlings grown on Norflurazon (Figure 5). Wild-type plants, grown under continuous white light without Norflur- azon, accumulated high levels of the nuclear-encoded CAB and RBCStranscripts and of the chloroplast-encoded PSA-AS (which encodes the P700 apoproteins of pho- tosystem I) and RSCL (which encodes the large subunit of ribulose bisphosphate carboxylase) transcripts (Figure 5, lane wt-Nf). None of these transcripts accumulated in photobleached wild-type plants (Figure 5, lane wt).
In contrast with wild-type plants, each of the gun mutants aberrantly accumulated endogenous CAB mRNA when photobleached (Figure 5). Most of the mutants also accu- mulated nuclear-encoded RBCS mRNA. However, in the photobleached mutants, the levels of CA6 and RBCS mRNAs were approximately lo-fold lower than in the green controls (Figure 5, lane wt-Nf). This was confirmed when we examined the levels of GUS that accumulated as a result of CA63 promoter expression. For instance, in the gunl-7 allele, the expression of the CA63-uicfA transgene was approximately 14-fold lower in photo- bleached gun7 seedlings than in gun7 seedlings not treated with the herbicide (Table 4). This was in contrast with the pOCA107-2 control, for which CABB-uidA expres- sion was 116-to 500-fold lower in photobleached seedlings (Tables 1 and 4).
staining for GUS activity and destaining to remove chlorophyll, we show three independent leaves. The indivrdual leaves were photographed before (top) and after (bottom) histochemical staining for GUS activity. (D) CA83 is expressed in both white and green sectors in the chm, gunl-i double mutant. As in (C), the individual leaves or whole plants were photographed before (top) and after (bottom) staining for GUS activity. The gun1 mutation clearly restores GUS activity in the unpigmented sectors in lo-day-old seedlings.

Cdl 792
.___ Table 4. Effect of Chloroplast Inhibitors on CABS-uidA Expression in gunl-1
Tissue Process GUS Activity Ratio .___
pOCAl07-2 Green (no Norflurazon) 7,410 z? 300 Photobleached (Norflurazon) 637 25 116 t
cab >> gonl-1 Green (no Norflurazon) 7,530 * 150 Photobleached (Norflurazon) 552 ? 37.5 13.6
pOCA107-2 No chloramphenicol 10,900 f 150 39 Chloramphenicol 280 r 120
gunl-1 No chloramphenicol 15.800 k 5,80’0 Chloramphenicol 5,210 * 3,390
3.0 .-__
GUS activity is reported as in Table 1. Seeds were germinated on MS agar with or without Norflurazon for 7 days before harvesting. The
psaA-B,j -
Figure 5. Transcript Accumulation in Photobleached Wild-Type and Mutant Plants
Wild-type (wt) and gun mutant plants were photobleached by growth on Norflurazon (Nf) unless otherwise noted. RNA was extracted, ana- lyzed by Northern analysis, and hybridized with radiolabeled probes from nuclear (cab and rbcS ribosomal RNA [rRNA]) and chloroplast (rbcL and psaA-B) genes.
Norflurazon-mediated photobleaching effectively blocked chloroplast development as determined by several crite- ria. First, none of the mutants accumulated carotenoids or chlorophylls under the conditions used for the screen (data not shown). Second, the chloroplast-encoded PSA- AB transcripts did not accumulate in either wild type or the mutants, confirming that the plastids were affected by Norflurazon (Figure 5). Third, electron microscopic analy- sis revealed that Norflurazon treatment blocked chloro- plast structural development in both wild-type Arabidopsis and in the gunl-7 mutant (Figure 6). Curiously, a second plastid transcript, RBCL, accumulated in most of the pho- tobleached mutants, with the exception of gunl-7 (see Figure 5). This may be due to the long half-life of RBCL transcripts (Klaff and Gruissem, 1991; Rapp et al., 1992). In addition, photobleached gunl-7 showed particularly strong GUS histochemical staining and CAB mRNA accu- mulation, indicating that it may be asevere allele of GUN7. For these reasons, we initially focused our attention on characterizing gunl-7 and the other gun7 mutants.
gun7 Mutations Affect Chloroplast Dependence but Not Other Aspects of CAB Regulation CAB transcription is regulated by endogenous develop- mental pathways, as well as by light. Yet these aspects
higher levels of GUS activity reported in Table 1 are from lo-day-old seedlings. For chloramphenicol treatment, seeds were sown on sterile filter paper layered on MS agar and then transferred to MS agar con- taining 150-200 pg/ml chloramphenicol after 45-48 hr of continuous white light. Plants were harvested after an additional week of growth.
of CABexpression appear unaltered in the mutants. Analy- ses of endogenous CAB mRNA accumulation demon- strated that CAB expression was light dependent in seed- lings, repressed in mature l ight-grown plants transferred to the dark, and under circadian control in both wild-type and gunl-7 mutant plants (Figure 7). Furthermore, none of the gun mutations perturb the normal pattern of t issue- specific CAB gene expression, since CABS-uidA transgene expression is restricted to the mesophyll layers of leaves in wild-type and gun mutant plants (see Figure 48; data not shown). Thus, the gun7 mutations specifically affect signaling between the chloroplast and the nucleus and do not appear to alter other aspects of CAB promoter regu- lation.
Alternatives to Herbicide-Mediated Chlorosis The primary effect of Norflurazon is to inhibit the enzyme phytoene desaturase, resulting in photooxidative damage to chloroplasts in bright light. We were concerned that possible secondary effects of Norflurazon or partial herbi- cide resistance in the mutants could complicate the inter- pretation of our results. Therefore, we sought alternatives for inhibiting chloroplast development.
Treatment with chloramphenicol, an inhibitor of organel- lar translation, prevented normal chloroplast development and inhibited nuclear CAB34dA expression by 39-fold in pOCAlO7-2 plants (Table 4). In contrast, chloramphenicol treatment inhibited CAB3 expression by only 3-fold in gunl-7 (Table 4). GUS activity is thus 19 times higher in the gunl-7 mutant than in pOCAl07-2 following growth on chloramphenicol. This difference is consistent with our observations from Norflurazon treatments (6-9 times higher in gunl-7 than in pOCA107-2) (Table 4).
We also made use of a genetic lesion affecting chloro- plast development. Chloroplast mutator chm is a recessive nuclear mutation of Arabidopsis with a variegated pheno- type. A single leaf may contain both normally pigmented sectors and albino sectors lacking chloroplast develop- ment (Redei, 1973). Initially, we crossed the pOCAl07-2

Uncoupling Nuclear and Chloroplast Gene Expression 793
a b d
Figure 6. Effect of Norflurazon-Mediated Photobleaching on Plastid Ultrastructure (a-d) Wild-type and gunl-i mutant seedlings were grown in the presence or absence of Norflurazon for 7 days. Cotyledon cross sections (1 pm thick) were analyzed by light microscopy. In the absence of Norflurazon, the photosynthetic mesophyll cells of both wild-type (a) and gunl-7 (b) plants are ringed with darkly stained chloroplasts. When grown on Norflurazon, plastids are not readily visible in either wild-type (c) or gunl-I (d) mesophyll cells. Scale bar, 50 Wm. (e-h) Ultrathin sections were analyzed by electron microscopy. Mutant chloroplasts (f) are as developed as wild-type chloroplasts (e) in green plants. In photobleached wild-type(g) and mutant(h) plants, plastids are present; however, they are small and fail to develop either the lens shape or thylakoid membranes characteristic of chloroplasts. Structures outside of the plastids (P), such as mitochondria (M), appear to be normal. Scale bar, 1 pm.

C&II 794
A.
B.
Cab *
Figure 7. CAB mRNA Accumulation rn Dark-Grown or Dark-Adapted Wild-Type and gunl-7 Plants
(A) RNA was extracted from 7-day-old pOCAlO7-2 (wt) and guni- (gunl) seedlings grown in either the dark or the light. The Northern blot was hybridized with an Arabidopsis CAB probe and then with a nuclear rRNA probe (data not shown) to confirm equal loading. A total of 3 ug of total RNA was loaded per lane. (B) Plants were grown on MS agar without sucrose and with 16 hr of light per day (Light). We then transferred plants to the dark (Dark) for the indicated number of hours before harvesting the plants and extracting RNA. Northern analysis was as described in (A).
transgenic line with chm and observed a pattern of CABS- uidA expression that resembled the pattern of chlorophyll pigmentation (see Figure 4C). Thus, the chm mutation ap- pears to be a suitable genetic substitute for Norflurazon photobleaching in blocking both chloroplast development and CAB expression. We next crossed the gunl-l muta- tion into the chm background and recovered chm, gunl-l double mutants (see Figure 4D). In striking contrast with the chm plants, chm, gunl-l double mutant plants clearly stained for GUS activity in both albino and green sectors (see Figure 4D).
We attempted to quantify the differences in CAB3-uidA expression in the green and white sectors of chm leaves in the presence or absence of the gunl-l mutation. We dissected by hand the green and white sectors from chm and the chm, gunl-l double mutant and assayed them for GUS activity using a fluorescent substrate. In six inde- pendent experiments, white sectors from the chm, gunl-l plants showed a 2.5 to 3.9-fold increase in specific GUS activity (average 700 U) over the activity measured in white sectors from the chm single mutant (average 240 U). CAB3-uidA expression in green sectors was high and comparable between the two lines (12,000-14,000 U). Though these results are less spectacular than those ob- tained with chloramphenicol and Norflurazon treatments, they may be explained by the fact that we had to use fully expanded, mature leaves for dissection studies (owing to the very small size of the sectors). We know from other studies using Norflurazon treatments that the highest lev- els of CA/33-uidA expression in gun1 alleles were ob-
served in young seedlings (5-7 days postgermination), with declining levels seen as the leaves matured and be- gan to senesce (data not shown). The histochemical stain- ing results presented in Figures 4C and 4D were from leaves taken from young plants and therefore appear more striking than the specific GUS activities that we quantified from green and albino sectors of mature plants.
Thus, herbicide treatment, chloramphenicol treatment, and the chm mutation all block normal chloroplast devel- opment. They do so by three very different primary mecha- nisms. Yet in each case CAB expression is severely re- duced in wild-type plants, whereas it is partially restored by the gunl-l mutation (by 3-to 20-fold, depending on the treatment).
gun1 Mutants May Be Defective in the Transition from Dark to Light Growth Mutations that disrupt important intracellular signaling pathways might cause severe physiological or morpholog- ical abnormalities. Nevertheless, most gun mutants look remarkably normal under most growth conditions” In par- ticular, gunl-l has a nearly wild-type phenotype when grown under conditions of variable day length, light quality, or intensity or when grown in the dark, on soil, or on plates with or without exogenous sucrose (data not shown).
We did observe a subtle phenotypic difference between gunl-l and wild-type seedlings during the transition from dark-grown (etiolated) to l ight-grown development, a pro- cess called de-etiolation. We found that dark-grown seed- lings of gunl-l lost the ability to de-etiolate after shorter periods in the dark than the wild type (Table 5). When dark-grown seedlings were transferred to white light after various times in the dark, a smaller proportion of mutant seedlings than wild-type seedlings de-etiolated. Whereas wild-type seedlings grown on sucrose lost their ability to de-etiolate after 9-10 days of growth in the dark, gunl-l seedlings lost their ability to de-etiolate by 7-9 days of dark growth. This effect on viability was striking; only 11% of gunl-l seedlings turned greened after 9 days of growth in the dark, while 95% of the wild-type seedlings were still able to turn green. Moreover, when dark-grown gun1
.- Table 5. Ability of gun1 to De-etiolate
Days of Total Percent Percent Dark Growth Strain Number Green Dead
3 Wild type 160 100 0 gun1-1 60 98 2
7 Wild type 64 100 0 gunl-1 39 49 51
9 Wild type 110 95 5 gull-1 72 11 89
10 Wild type 46 8 92 gunl-1 66 0 100
Wild-type pOCA107-2 and gun7 mutant plants were grown rn the dark for 3, 7, 9, or 10 days and then transferred to continuous white light for at least 7 days before we scored plants with long hypocotyls as green (i.e., cotyledons expanded and pigmented) or dead (i e., fully etiolated).

Uncoupling Nuclear and Chloroplast Gene Expression 795
mutant plants were transferred to the light, they accumu- lated chlorophyll and developed chloroplasts more slowly and less efficiently than dark-grown wild-type plants (data not shown). The effect on de-etiolation in gunl-1 seedlings was verified using several different seed batches (data not shown). In addition, we assayed this phenotype in two additional gun1 mutants, gunl-2 and gunl-4, and ob- served that they were also defective in greening, though the de-etiolation defect in these two alleles was not as severe as for gunl-1 (data not shown). This greening de- fect did not appear to be a result of aberrant growth in the dark. That is, dark-grown gun1 mutants resembled normal etiolated wild-type plants; they failed to accumulate CAB mRNA or chlorophyll, had long hypocotyls and folded coty- ledons, and contained etioplasts (data not shown).
Discussion
Genetic Evidence for a Pathway That Couples Nuclear CAB and RBCS Expression with Chloroplast Function Previously published molecular, physiological, and ge- netic studies suggest that the transcription of several nu- clear genes encoding photosynthesis-related proteins is dependent upon chloroplast development (Taylor, 1989; Barkan, 1993). Here, we provide genetic evidence for the existence of such a signal transduction pathway. We have identified nine nuclear gun mutations that allow the partial expression of the nuclear genes CAB and RBCS in the absence of chloroplast development. Complementation analysis indicates that at least three to six GUN genes function in this signaling pathway(s).
By a number of criteria, the gun mutants carry genetic lesions that are specific for a pathway that couples the expression of certain nuclear genes with chloroplast devel- opment. First, the gun mutants were not Norflurazon resis- tant. Norflurazon effectively blocked carotenoid and chlo- rophyll pigment accumulation (data not shown) and chloroplast development in both wild-type and mutant seedlings (Figure 6). Plastids proliferated in both photo- bleached strains; however, they were small and irregularly shaped and lacked internal thylakoid membranes. Al- though the gun mutants lost the chloroplast dependence of CA6 expression, they maintained normal patterns of tissue-specific, cell-type-specific, l ight-dependent, and dark-adapted CA6 expression (Figures 46 and 7). gunl-1 also showed a wild-type pattern of CA6 mRNA accumula- tion following growth under a variety of other controlled environmental conditions, including variable hours of day- light, light quality, light intensity, and supply of exogenous sucrose (data not shown).
Though our original screen relied on the herbicide Nor- flurazon to block normal chlorcplast development, we have subsequently verified the mutant phenotypes using an inhibitor of organellar translation. Chloramphenicol treatment blocks chloroplast development and simultane- ously reduces nuclear CABS-uidA expression by 39-fold in wild-type plants, but only 3-fold in gunl-7 mutants. Fi- nally, a set of genetic experiments using the chm varie- gated mutant of Arabidopsis also verified that the gunl-1
mutation uncoupled nuclear CA6 expression from chloro- plast function. Albino sectors of chm plants have severely impaired chloroplast development, and nuclear CABB- uidA is expressed at very low levels in these white sectors (Figure 4C). However, CABS-uidA levels are, in general, greater than 3-fold higher in the white sectors of chm, gun1 double mutants versus the chm single mutant. Thus, we have shown that three separate mechanisms of blocking chloroplast function repress nuclear CA6 expression in wild-type plants. In each case, the gun1 mutation partially restores CA6 promoter activity (by 3-to 20-fold, depending on the treatment). Thus, we have multiple lines of evidence in support of the hypothesis that the gun1 mutation uncou- ples nuclear CA6 and RBCS expression from chloroplast function.
The requirement of nuclear GUN genes for coupling transcription of nuclear genes such as CA6 and RBCS to the functional state of the chloroplast suggests that infor- mation flows from chloroplasts to the nucleus. Although the CA6 and RBCS promoters share common sequences that bind some of the same transcription factors, it is not yet clear which of these are responsible for responding to the functional state of the chloroplast (Datta and Cash- more, 1989; Schindler and Cashmore, 1990). Northern analysis and CABB-uidA expression studies confirm aber- rant CA6 expression in photobleached gun mutants, yet the mutations do not restore CA6 expression to its maxi- mum in green plants (Table 4). This suggests that this signal transduction pathway from plastids to nucleus is just one factor contributing to the final CA6 and RBCS gene expression responses.
The gun1 mutants appear quite normal at later stages of development and under normal growth conditions were indistinguishable from wild type. However, we observed a deficiency in the greening process during de-etiolation in the gun1 mutants. Although each of the gun mutants develops normally in the dark, three independent gun7 mutant alleles lost the ability to de-etiolate after less time in the dark than the wild type. Moreover, in preliminary experiments designed to examine early greening events, gun1 accumulated chlorophyll and carotenoids and devel- oped chloroplasts more slowly than the wild type (FL E. S. and J. C., unpublished data). This suggests that coupled nuclear and chloroplast gene expression may be espe- cially important during early events in chloroplast differen- tiation. This is consistent with our results that show CA63 expression in gun7 is highest in very young seedlings (5-7 days old). Further work will be required to determine whether these data reflect a requirement for GUN7 during de-etiolation. Our ongoing molecular and plastid ultra- structural analyses should further detail these ObseNa- tions.
It was unexpected that chloroplast-encoded RBCL tran- scripts would accumulate in most of the photobleached gun mutants. Clearly, the mutants that accumulate RBCL mRNA are not resistant to photobleaching since they lack detectable chlorophyll and carotenoid pigments and fail to accumulate PSA-A6 transcripts. It is possible that wild- type plants transiently express RBCL early during chloro- plast biogenesis, despite photobleaching. Thus, the pho-

Cell 796
Figure 8. Possible Models to Explain gun Mutations
(A) gun mutations disrupt a linear, unidirectional signaling pathway from plastid to nucleus. Given the recessive nature of the gun muta- tions, the simplest model is that a negative regulatory signal from damaged plastids would act to repress nuclear CA6 transcription. gun mutations could affect any of the transduction or perception steps. Our data do not exclude the possibility that a positive regulatory signal from intact chloroplasts could function to activate nuclear CA6 and RBCS transcription. In this model and the following ones, the sche- matic plant cell (large box) contains a nucleus. The bent, closed arrow in the nucleus represents transcription from CA8 and WCS genes. The open arrowheads depict the signal transduction pathways. (B) gun mutations affect the synthesis of the chloroplast signal. In this model, a normal plastid metabolite (6) serves as the primary signal molecule in a simple signal transduction pathway. As in (A), this could be either a positive or negative regulatory signal. A-B4 represents a hypothetical plastid metabolic pathway. Agun mutation would affect metabolite synthesis or accumulation. If the normal case is a positive signal from functional chloroplasts that activates CAB and RBCS ex- pression, a gun lesion might prevent the conversion of metabolite B to C, thereby elevating the levels of B even after photobleaching. If, instead, metabolite B functions to repress CAB and RBCSexpression after plastid damage, a gun lesion blocking the conversion of metabo- lite A to B would minimize accumulation of the negative regulator, thereby allowing CA8 and RBCS transcription. (C) gun mutation disrupts a bidirectional communication pathway in- volving a cytoplasmic informational processor (stippled cube). A hypo- thetical cytoplasmic component both monitors and responds to the physiological status of the plastid and nucleus. Mutations in this infor- mational processor or in any of the signal transduction steps could lead to a gun phenotype.
tobleached mutants might accumulate RBCL transcripts aberrantly because of elevated or prolonged REEL tran- scription or because RBCL transcripts are stabilized. This explanation is feasible since photobleaching requires chlorophyll function, and thus the initiation of chloroplast biogenesis, the rates of synthesis, and the stability of chlo- roplast transcripts can vary greatly among genes or among developmental stages (Deng and Gruissem, 1987; Kuchka
et al., 1989; Ngernprasirtsiri et al., 1990; Klaff and Gruis- sem, 1991; Rapp and Mullet, 1991; Rapp et al., 1992). An alternative explanation for our RBCL results is that gun mutants maintain a low level of chloroplast transcription, despite photobleaching, that may be sufficient for RBCL transcript accumulation. Regardless of the explanation, our major focus was gun 7-7, a particularly strong gun allele that does not show the unexpected RBCL transcript accu- mulation.
Possible Roles of GUN Genes The GUN genes appear to coordinate nuclear CAB and RBCS transcription with chloroplast function; neverthe- less, the identity of the primary signal molecule and the method of its transduction are unknown. We propose three models for this interorganellar signal transduction pathway.
In the simplest model (Figure 8A), chloroplasts or dam- aged chloroplasts produce either a positive or negative regulatory signal, respectively, that is transduced to the nucleus, where it affects CABand RBCStranscription. The nuclear GUN genes could function in signal transduction or response in such a simple unidirectional pathway. Given the recessive nature of the mutations, it appears likely that a pathway functions to repress CAB and RBCS expression. We do not have data to address whether a positive signaling pathway also exists.
Alternatively, thegun mutations may affect the synthesis of the plastid signal itself (Figure 88). The signal could be a normal plastid metabolic pathway intermediate or product. For instance, metabolite B could be a positively acting signal. Photobleaching might diminish the meta- bolic flux through this pathway and thus reduce the levels of the positive signal. A gun lesion could block the conver- sion of B to C and thereby restore high levels of metabolite 6 and CAB gene expression. Alternatively, metabolite B could be a negative regulatory signal that accumulates to high levels only in photobleached plants. A gun mutation that blocked the conversion of metabolite A to B could thus prevent synthesis of the negative regulator, leading to derepression of CAB gene expression. Although herbi- cide-treated plastids are impaired functionally, they likely retain some metabolic activities (e.g.? lipid and porphyrin biosynthesis) because the plants recover when rescued from the herbicide. Yeast provide a precedent for an orga- nellar metabolite functioning to modulate nuclear gene transcription. In yeast, mitochondrial heme appears to modulate the activity of nuclear transcription factors that specifically regulate nuclear genes encoding mitochon- drial proteins (Forsburg and Guarente, 1989).
The final model involves a multidirectional flow of infor- mation through a cytoplasmic component that integrates and responds to a host of physiological cues (Figure 8C). Such a cytoplasmic regulator could couple nuclear and chloroplast gene transcription. gun mutants might affect the cytoplasmic regulator or the signal transduction steps, From these three models, we can envision a nuclear, cyto- plasmic, or chloroplastic site of localization for the GUN gene products.

Uncoupling Nuclear and Chloroplast Gene Expression 797
Complex Intracellular Regulatory Network among Organelles There is growing evidence for a complex intracellular regu- latory network controlling organellar development. Other nuclear genes, such as DET, HY, and COP (Chory et al., 1989a; Chory and Peto, 1990; Chory, 1991; Deng and Quail, 1992), that act to couple leaf development and chlo- roplast biogenesis to the light environment are additional components of this complex regulatory network control- ling chloroplast biogenesis. Genetic studies provide evi- dence for mitochondrial involvement in chloroplast devel- opment as well. The variegated pigmentation mutant used in this study was named chm (chloroplast mutator) to sug- gest that this recessive nuclear mutation caused muta- tions of the chloroplast genome, resulting in cytoplasmi- tally inherited variegation (Redei, 1973). A recent study demonstrates that chm instead generates mitochondrial genome deletions and rearrangements, apparently with- out altering the plastid genome (Martinez-Zapater et al., 1992). The resulting mitochondrial deficiencies block chlo- roplast biogenesis. Similarly, the maize nonchromosomal stripe (ncs) mitochondrial mutations confirm a role for mito- chondria in chloroplast development (Lauer et al., 1990; Roussell et al., 1991). One likely explanation is that mito- chondria supply the energy requirements for chloroplast differentiation (Leaver and Forde, 1980; Wellburn, 1984; Owen and Wellburn, 1987). In addition, chloroplasts sup- ply lipids and heme to the developing mitochondria (Stumpf, 1980; Howe and Smith, 1991). In yeast, the nu- clear RTG7 and RTG2 genes control the interorganellar communication among mitochondria, nucleus, and peroxi- somes (Liao and &tow, 1993).
We previously suggested that a chloraphyll precursor, heme, or other porphyrin could act as the signal between chloroplast and nucleus (Susek and Chory, 1992). This same signal could play a larger role in the regulatory net- work by also helping to coordinate chloroplast and mito- chondrial functions. There is precedent for porphyrins as- suming regulatory functions. For example, chlorophyll precursors specifically inhibit CAB mRNA accumulation in Chlamydomonas (Johanningmeier and Howell, 1984). Heme is a feedback regulator of porphyrin biosynthesis in plants (Johanningmeier and Howell, 1984; Goodwin and Mercer, 1985). In yeast, heme also regulates the import and processing of nuclear-encoded mitochondrial pro- teins as well as the transcription, accumulation, and trans- lation of their mRNAs (Johanningmeier and Howell, 1984; Forsburg and Guarente, 1989).
Summary Chloroplast biogenesis involves complex cross-talk among the chloroplast, nucleus, and mitochondrion. The coordinated efforts of these organelles afford plants an amazing potential for development and adaptation to changing environmental conditions. We have screened for mutants displaying aberrant CA63 promoter-driven transgene expression and identified a class of mutations that affects the branch of the network that couples nuclear
CAB and RBCS gene expression to chloroplast function. The gun mutants will serve as useful tools for further dis- section of this intricate intracellular signal transduction pathway.
Experimental Procedures
Construction of the pOCA107-2 Transgenic Line of Arabidopsis We created the pOCAl07 transgenic construct by modification of pOCA18 (Olszewski et al., 1988). Initially. the nopaline synthase pro- moter of pOCA18 was replaced by the cauliflower mosaic virus 35s promoter (Herrera-Estrella et al., 1983; Ow et al., 1986). We next cre- ated two transcriptional fusions (-9 to ATG) between the A. thalrana CAB3 (also called CAB Al3180 [Leutwiler et al., 19861) promoter and either the uidA (GUS [Jefferson et al., 19FJ6]) or hph (Gritz and Davies, 1983) coding sequences. We added nos terminator sequences, then inserted the CA53 reporter gene fusions between the right border and the 35S-nptll-110s fusion. This yielded pOCA107. We introduced pOCA107 into A. thaliana ecotype Columbia using a modified version of the standard Agrobacterium-mediated root transformation protocol (Valvekens et al., 1988; Chory and Peto, 1990) generating more than 30 independent transgenic lines. We picked one line, pOCAl07-2, for which the segregation of kanamycin resistance suggested a single pOCA107 insertion site. The pOCA107-2 transgenic insertion maps to chromosome II, 3 CM from hyl (L. Altschmied and J. C., unpublished data).
Plant Material, Growth Conditions, and Genetic Methods Seeds (50,000) of the pOCAl07-2 transgenic line were mutagenized with ethyl methanesulfonate, as previously described (Chory et al., 1989a). These Ml seeds were sown on soil and allowed to self- pollinate. M2 seeds were harvested in 200 pools. We surface stenlrzed 100,000 M2 seeds and then plated them on Murashige-Skoog (MS) agar plates containing 2% sucrose, 85 vitamins, 5 uM Norflurazon (provided by Sandoz Chemical Company), and 20 vg/ml hygromycin B (Boehringer Mannheim). Plates were chilled at 4OC overnight and then maintained at 20°C-220C under a mixture of fluorescent and incandescent lights that provided continuous high fluence light (2 x lO’6 quantals/cm2). We measured light intensities with a QSL-100 quantum scalar irradiance meter (Biospherical Instruments, Incorpo- rated). After 7-10 days, we monitored CA83-hph transgene expres- sion by visual inspection for large seedlings. We assayed putative hygromycin-resistant seedlings for CABS-uidA transgene expression by snipping off a cotyledon and staining it for GUS activity (Jefferson et al., 1986). The remainder of the seedling was transferred to a fresh plate lacking Norflurazon and allowed to recover and self-propagate.
All our mutant screens included two negative controls: non- transgenic wild-type and wild-type pOCA107-2 plants. As a positive control, we included pHG1 plants carrying a transgenic construct simi- lar to pOCAlO7, but in which the Arabidopsis PAL1 promoter replaces the CA63 promoters (S. Ohl, B. Kraft, J. C., and C. J. Lamb, unpub- lished data). PAL7 expression occurs in roots, hypocotyls, and cotyle- dons and is insensitive to photobleaching. We initially used these three controls to optimize the screening conditions for ease of screening and maximal plant recovery and fertility.
In subsequent screens we incorporated two modifications. First, we shortened the period of Norflurazon treatment to 5 days to achieve an improved rate of recovery and fertility. Second, we added a formal- dehyde fixation step to prevent recovery from photobleaching during the overnight staining procedure for GUS activity (Jefferson et al., 1986). With these changes we now observe far fewer false positive plants.
In experiments to assess RNA accumulation in the dark or the transi- tion from etiolated to light-grown development, seeds were plated as described above and then given a 12 hr light treatment before they were transferred to the dark. In the greening experiments, dark-grown plants were moved to continuous white light for the indicated periods. RNA from dark-adapted plants was extracted from plants grown on

Cell 790
MS agar lacking sucrose for 14 days with 16 hr of light per day and then were transferred to the dark for the indicated times.
GUS Activity Assays GUS activity was assayed in seedlings grown on plates as described above. For histochemical staining, tissue was harvested, fixed, and incubated overnight at 37’C in 1 mglml X-GLUC (Jefferson et al., 1986). For quantitative GUS assays, harvested tissues were frozen, ground in extraction buffer, and assayed as previously described (Jef- ferson et al , 1986).
Light and Electron Microscopy of Plastid Development Seedlings were grown on MS plates, with or without Norflurazon, for 7 days or as described above. Cotyledons were harvested and fixed for 2-3 hr in 5% gluteraldehyde in 0.1 M sodium phosphate buffer (pH 7.2) with gentle vacuum infiltration. The tissue was buffer rinsed, postfixed in buffered 2% osmium tetraoxide, buffer rinsed again, dehy- drated in ethanol, and then embedded in Spurr’s resin. Sections (1 vrn thick) were cut, stained with toluidine blue, and examined by light microscopy. Thin sections of - 0.1 pm werestained with uranyl acetate and lead citrate or with bismuth subnitrate and were examined on a Zeiss EM-IOB or Philips 300 electron microscope.
Northern Analysis Plant tissue was harvested, and the RNA was extracted and analyzed on Northern blots as previously described (Chory et al., 1991). The hybridization probes used were previously described (Chory et al., 1989a).
Pigment Analysis Chlorophyll and carotenoid pigments were extracted in 80% acetone from tissue frozen and ground to a fine powder in liquid nitrogen. Pigment determinations were made spectrophotometrically using the equations of Lichtenthaler (1987).
Acknowledgments
Correspondence should be addressed to J. C. We thank the members of the Chory and Lamb laboratories and Hsou-min Li and Lothar Alt- schmied in particular for their comments and suggestions throughout the course of this work. We also thank Lothar Altschmied for the muta- genized pOCAlO7-2 seeds and Casey Peto and AbbyAnn Sisk for their expert assistance with the electron microscopy. We thank Stephen Worland, Chris Lamb, and Steven Rodermel for their comments on the manuscript. Also, we are grateful to the editor and to a reviewer who made several suggestions that improved our manuscript. This work was supported by grants from the Department of Energy, the Samuel Roberts Noble Foundation, and the Biomedical Research Sup- port Grant Program (grant number RR05595) at the Salk Institute to J. C. R. E. S. received postdoctoral training funds from the Charles and Ruth Billingsley Foundation and the National Institutes of Health. Early stages of this work were supported by a grant to Massachusetts General Hospital from Hoechst Aktiengesellschaft.
Received December 10, 1992; revised June 25, 1993
References
An, G. (1987). Integrated regulation of the photosynthetic gene family from Arabidopsis fhaliana in transformed tobacco cells. Mol. Gen. Genet. 270, 210-216.
Barkan, A. (1993). Nuclear mutants of maize with defects in chloroplast polysome assembly have altered chloroplast RNA metabolism. Plant Cell 5. 389-402.
Batschauer, A., Miisinger, E., Kreuz, K., D6rr, I., and Apel, K. (1986). The implication of a plastid-derived factor in the transcriptional control of nuclear genes encoding the light-harvesting chlorophyll a/b protein. Eur. J. Biochem. 754, 625-634. Burgess, D. G., and Taylor, W. C. (1987). Chloroplast photooxidation affects the accumulation of cytosolic mRNAs encoding chloroplast proteins In maize. Pianta Med. 770, 520-527. Chamovitz. D.. Pecker, I., and Hirschberg, J. (1991). The molecular
basis Of resistance to the herbicide Norflurazon. Plant Mol. Biol. 76, 967-974.
Chory, J. (1991). Light signals in leaf and chloroplast development: photoreceptorsand downstream responses in search of a transduction pathway. New Biol. 3, 538-548.
Chory, J., and Peto, C. A. (1990). Mutations in the DET7 gene affect cell-type-specific expression of light regulated genes and chloroplast development in Arabidopsis. Proc. Natl. Acad. Sci. USA 87, 8776- 8780.
Chory, J., Peto, C., Feinbaum, R., Pratt, L., and Ausubel, F. (1989a). Arabldopsis thaliana mutant that develops as a Ilght-grown plant In the absence of light. Cell 58, 991-999.
Chory, J., Peto, C. A., Ashbaugh, M., Saganich, R.. Pratt, L., and Ausubel, F. (1989b). Different roles for phytochrome in etlolated and green plants deduced from characterization of Afabidopsis thaliana mutants. Plant Cell 7, 867-880.
Chory, J., Nagpal, P., and Peto, C. A. (1991). Phenotypic and genetlc analysis of def2, a new mutant that affects light regulated seedling development in Arabidopsis. Plant Cell 3, 445-459.
Datta, N.,andCashmore,A. R. (1989). Bindingofapeanuclear protein to promoters of certain photoregulated genes is modulated by phos- phorylation. Plant Cell 7, 1069-1077.
Deng, X.-W., and Gruissem, W. (1987). Control of plastid gene expres- sion during development: the limited role of transcriptional regulation. Cell 49, 379-387.
Deng, X.-W., and Quail, P. (1992). Genetic and phenotypic character- ization of cop1 mutants of Afabidopsis thaliana. Plant J. 2. 83-95.
Forsburg, S. L., and Guarente, L. (1989). Commumcatlon between mitochondria and the nucleus in regulation of cytochrome genes in the yeast Saccharomyces cerevisiae. Annu. Rev. Cell BioV. 5. 153- 180.
Gilmartin, P. M., Sarokin, L., Memelink, J., and Chua, N.-H. (1990). Molecular light switches for plant genes. Plant Cell 2. 369-378. Goodwin, T. W., and Mercer, E. I. (1985). Introduction to Plant Bio- chemistry (New York: Pergamon).
Gritz, L., and Davies, J. (1983). Plasmid-encoded hygromycm B rests- tance: the sequence of hygromycin B phosphotransferase gene and its expression in Escherichia co/i and Saccharomyces cerevisiae. Gene 25, 179-l 88.
Ha, S. B., and An, G. (1988). Identification of upstream regulatory elements involved in the developmental expression of the Arabidopsis thaliana cab7 gene. Proc. Natl. Acad. Sci. USA 85, 8017-8021.
Harpster, M. H., Mayfield, S. P., and Taylor, W. C. (1984). Effects of pigment-deficient mutants on the accumulation of photosynthetic proteins in maize. Plant Mol. Biol. 3, 59-71.
Herrera-Estrella, L., DeBlock, M., Messens, E., Hernalsteens, J., van Montagu, M., and Schell, J. (1983). Chimeric genes as dominant se- lectable markers in plant cells. EMBO J. 2, 987-995. Howe, C. J., and Smith, A. G. (1991). Plants without chlorophyll Nature 349, 109.
Jefferson, R. A., Burgess, S. M., and Hirsh, D. (1986). !%Glucuronldase from Escherichia co/i as a gene-fusion marker. Proc. Natl. Acad. Sci. USA 83, 8447-8451.
Johanningmeier, U., and Howell, S. (1984). Regulation of light- harvesting chlorophyll-binding protein mRNA accumulation in Chlamy- domonas reinhardi. J. Biol. Chem. 259, 13541-13549
Kendrick, R. E., and Kronenber, G. H. M. (1986). Photomorphogenesis in Plants (Bordrecht, The Netherlands: Mantinus Nijhoff).
Kirk, J. T. 0.. and Tilney-Basset, R. A. E (1978). The Plastlds. Their Chemistry, Structure, Growth and Inheritance (Amsterdam’ Elsevier). Klaff, P., and Gruissem, W. (1991). Changes In chloroplast mRNA stability during leaf development. Plant Cell 3, 517-529.
Kuchka, M. R., Goldschmidt-Clermont, M., van Dillewijn, J., and Ro- chaix. J.-D. (1989). Mutation at the Chlamydomonas nuclear NACP locus specifically affects stability of the chloroplast psbD transcript encoding polypeptide D2 of PS II. Cell 58, 889-876. Lauer. M., Knudsen, C., Newton, K., Gabay-Laughnan, S., and Laugh- nan, J. (1990). A partially deleted mitochondnal cytochrome oxidase

Uncoupling Nuclear and Chloroplast Gene Expression 799
gene in the NCSG abnormal growth mutant of maize. New Biol. 2, 179-186.
Leaver, C., and Forde, B. (1980). Genome Organization and Expres- sion in Plants (New York: Plenum Press).
Leutwiler, L., Meyerowitz, E., and Tobin. E. (1986). Structure and ex- pression of three light-harvesting chlorophyll a/b-binding protein genes in Arabidopsis tbaliana. Nucl. Acids Res. 14. 4051-4064.
Liao, X., and Butow, R. A. (1993). RTG7 and RTGP: two yeast genes required for a novel path of communication from mitochondria to the nucleus. Cell 72, 61-71.
Lichtenthaler, H. K. (1987). Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth. Enzymol. 748, 350-382.
Martineau, B., and Taylor, W. C. (1986). Cell-specific gene expression and cellular differentiation in developing maize leaves. Plant Physiol. 78, 399-404.
Martinez-Zapater, J. M., Gil, P., Capel, J., and Somerville, C. R. (1992). Mutations at the Arabidopsis CHM locus promote rearrangements of the mitochondrial genome. Plant Cell 4, 889-899.
Mathews, D. E., and Durbin, R. D. (1990). Tagetitoxin inhibits RNA synthesis directed by RNA polymerases from chloroplasts and Esche- richia co/i. J. Biol. Chem. 265, 493-498.
Mayfield, S. P., and Taylor, W. C. (1984). Carotenoid-deficient maize seedlings fail to accumulate light-harvesting chlorophyll a/b binding protein (LHCP) mRNA. Eur. J. Biochem. 144, 79-84.
Millar, A. J., and Kay, S. A. (1991). Circadian control of cab gene transcription and mRNA accumulation in Arabidopsis. Plant Cell 3, 541-550.
Mitra, A., Choi, H. K., and An, G. (1989). Structural and functional analyses of Arabidopsis thaliana chlorophyll a/b-binding protein (cab) promoters. Plant Mol. Biol. 72, 169-179.
Ngernprasirtsiri, J., Kobayashi, H., and Akazawa, T. (1990). Expres- sion of photosynthetic genes is distinctly different between chloro- plasts and amyloplasts in the liquid-cultured cells of sycamore (Acer pseudoplatanus L.). Cell Struct. Funct. 15, 273-283.
Oelmiiller, R. (1989). Photooxidative destruction of chloroplasts and Its effects on nuclear gene expression and extraplastidic enzyme levels. Photochem. Photobiol. 49, 229-239.
Oelm0ller, R., and Briggs, W. R. (1990). Intact plastids are required for nitrate- and light-induced accumulation of nitrate reductase activity and mRNA in squash cotyledons. Plant Physiol. 92, 434-439.
Oelmdller, R., Levitan, I., Bergfeld, R., Rajasekhar, V. K., and Mohr, H. (1986). Expression of nuclear genes as affected by treatments act- ing on the plastids. Planta Med. 768, 482-492.
Olszewski, N., Martin, F., and Ausubel, F. (1988). Specialized binary vector for plant transformation: expression of the Arabidopsis thaliana AHASgene in Nicotiana tabacum. Nucl. Acids Res. 16, 1076510782.
Ow. D., Wood, K., DeLuca, M., DeWet, J., Helinski, D., and Howell, S. (1988). Transient and stable expression of the firefly luciferase gene rn plant cells and transgenic plants. Science 234, 856-859. Owen, J., and Wellburn, A. (1987). Respiratory participation by mito- chondria during early plastidogenesis in barley seedlings. In Plant Mitochondna: Structural, Functional and Physiological Aspects, J. Owen and A. Wellburn, eds. (New York: Plenum Press), pp. 211-214. Plumley, F. G., and Schmidt, G. W. (1989). Nitrogen-dependent regula- tion of photosynthetic gene expression. Proc. Natl. Acad. Sci. USA 86, 2678-2682.
Rapp, J., and Mullet, J. (1991). Chloroplast transcription is required to express the nuclear genes RBCS and CAB: plastid DNA copy num- ber is regulated independently. Plant Mol. Biol. 17, 813-823. Rapp, J.. Baumgartner, B., and Mullet, J. (1992). Quantitative analysrs of transcription and RNA levels of fifteen barley chloroplast genes: transcription rates and mRNA levels vary over 300-fold; predicted mRNA stabilities vary 30-fold. J. Biol. Chem. 267, 21404-21411.
Redei, G. (1973). Extra-chromosomal mutability determined by a nu- clear gene locus in Arabidopsis. Mutat. Res. 78, 149-162.
Roussell, D., Thompson, D., Pallardy, S., Miles, D., and Newton, K. (1991). Chloroplast structure and function is altered in NCS2 maize mitochondrial mutant. Plant Physiol. 96, 232-238.
Schindler, U., and Cashmore, A. R. (1990). Photoregulated gene ex- pression may involve ubiquitous DNA binding proteins. EMBO J. 9, 3415-3427.
Schwartz, B. W., Daniel, S. G., and Becker, W. M. (1991). Chloroplast photobleaching reduces expression of the peroxisomal NADH- dependent hydroxypyruvate reductase in developing cucumber cotyle- dons. Plant Physiol. (Suppl.) 96, 120.
Simpson, J., van Montagu, M., and Herrera-Estrella, L. (1986). Photo- synthesis-associated gene families: differences in response to trssue- specific and environmental factors. Science 233, 34-38
Stumpf, P. (1980). The Biochemistry of Plants: Lipids Structure and Function (New York: Academic Press).
Susek, R., and Chory, J. (1992). A tale of two genomes: role of a chloroplast signal in coordinating nuclear and plastid genome expres- sion. Aust. J. Plant Physiol. 19, 387-399.
Taylor, W. C. (1989). Regulatory interactions between nuclear and plastid genomes. Annu. Rev. Plant Physiol. Plant Mol. Brol. 40, 211- 233.
Taylor, W. C., Burgess, D. G., and Mayfield, S. P. (1986). The use of carotenoid deficiencies to study nuclear-chloroplast regulatory tnter- actions. Curr. Topics Plant Biochem. Physiol. 5, 117-127.
Taylor, W. C., Barkan, A., and Martienssen, R. A. (1987). Use of nu- clear mutants in the analysis of chloroplast development. Dev. Genet. 8, 305-320.
Tzagoloff, A., and Dieckmann, C. (1990). Pet genes of Saccharomyces cerevisiae. Microbial. Rev. 54, 211-225.
Valvekens, D., van Montagu, M., and van Lrjsebetterns, M. (1988). Agrobacterium tumefaciens-mediated transformation of Arabidospsis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. SCI. USA 85, 5536-5540.
Wellburn, A. (1984). Chloroplast Biogenesis (Amsterdam: Elsevier).
Whittaker, P. A., and Danks, S. M. (1978). Mrtochondna: Structure, Function and Assembly (New York: Longman).