short chain fatty acids accumulation and microbial community succession during ultrasonic-pretreated...
TRANSCRIPT

lable at ScienceDirect
International Biodeterioration & Biodegradation 94 (2014) 128e133
Contents lists avai
International Biodeterioration & Biodegradation
journal homepage: www.elsevier .com/locate/ ibiod
Short chain fatty acids accumulation and microbial communitysuccession during ultrasonic-pretreated sludge anaerobicfermentation process: Effect of alkaline adjustment
Yali Liu, Xin Li*, Xiaorong Kang, Yixing Yuan**, Maoan DuSchool of Municipal and Environmental Engineering, Harbin Institute of Technology, Harbin 150090, China
a r t i c l e i n f o
Article history:Received 2 December 2013Received in revised form5 July 2014Accepted 10 July 2014Available online
Keywords:Waste activated sludgeMicrobial communitySCFAsAlkaline adjustmentProtease
* Corresponding author. Tel.: þ86 18646594718.** Corresponding author. Tel.: þ86 13904631327; fa
E-mail addresses: [email protected] (Y. Liu),[email protected] (X. Kang), [email protected] (M. Du).
http://dx.doi.org/10.1016/j.ibiod.2014.07.0040964-8305/© 2014 Elsevier Ltd. All rights reserved.
a b s t r a c t
Short chain fatty acids (SCFAs) are value-added products from waste activated sludge (WAS) anaerobicfermentation. The effects of alkaline adjustment, in the range of pH 7e10, on SCFAs accumulation andmicrobial community of ultrasonic-pretreated WAS at 20 ± 2 �C were investigated in this study.Experimental results indicated that optimum conditions for SCFAs accumulation were pH 9 andfermentation time of 7 days. The changes of protease activity showed that on the initial phase, ultrasonicpretreatment and alkaline adjustment rather than biotic effect was the leading cause of SCFAsimprovement at high pHs. DGGE analysis revealed that alkaline adjustment had a strong impact onmicrobial community. Some bacteria belonged to Firmicutes, Proteobacteria and Bacteroidetes could adaptto high alkaline environments and contribute to proteins and carbohydrates degradation and SCFAsproduction in the later phase of anaerobic fermentation.
© 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Short chain fatty acids (SCFAs) can be used as raw materials toproduce polyhydroxyalkanoates (Jia et al., 2013), preferred sub-strate to generate electricity (Gonzalez del Campo et al., 2012), andfavorable internal carbon source to remove phosphorus and ni-trogen in wastewater (Zhu and Chen, 2011). Recently, SCFAs havebeen considered to be the value-added products of waste activatedsludge (WAS), which can be produced in anaerobic fermentationprocess (Cokgor et al., 2009).
Three steps, hydrolysis, acidogenesis and methanogenesis areinvolved in anaerobic fermentation process, in which acidificationis recognized as a promising and feasible way for the production ofSCFAs fromWAS (Liu et al., 2012). It is proposed that the high SCFAsproduction is due to two aspects: more soluble substances aregenerated and methanogens are inhibited. As we know, initial hy-drolysis of solid sludge to soluble substances is the limiting step ofanaerobic fermentation (Chen et al., 2007). Noticeably, severalpretreatment methods have been developed to enhance the sludge
x: þ86 [email protected] (X. Li),(Y. Yuan), dmaharbin@163.
hydrolysis, such as ultrasound (Luste et al., 2011), alkaline (Kimet al., 2013), Fenton reagent (Erden and Filibeli, 2010), and biolog-ical enhancement (Oh et al., 2013). Among these methods, ultra-sonic pretreatment is one of the most promising technologies foraccelerating sludge hydrolysis and shortening subsequent acidifi-cation time (Park et al., 2013). The optimal ultrasonic parametersfor sludge disintegration were studied by some researchers (Kimet al., 2010; Yan et al., 2010). Simultaneously, acetate as the finalproduct of acidification stage is split intomethane (CH4) and carbondioxide (CO2) by methanogens. As one of the most important pa-rameters for fermentation, pH can obviously affect the SCFAs pro-duction. Some studies have reported that SCFAs production wassignificantly improved under alkaline conditions (such as pH 10.0)than under acid conditions (Wu et al., 2009; Yan et al., 2010).
Polymerase chain reaction-denaturing gradient gel electropho-resis (PCR-DGGE), a culture-independent molecular biologicalmethod, offers a valuable tool for the characteristics of the micro-bial community structure in sludge fermentation process. Thespecies of microbes involved in fermentation and their response tofermentation reaction have been well documented (Shin et al.,2010; Kim et al., 2011a). The purpose of these studies was toanalyze the relationship between microbial community andfermentation efficiency, including hydrogen production and CH4yield. Therefore, grasping the effect of alkaline adjustment on mi-crobial community structure is benefit for SCFAs accumulation

Y. Liu et al. / International Biodeterioration & Biodegradation 94 (2014) 128e133 129
during ultrasonic-pretreated sludge anaerobic fermentationprocess.
The purpose of this study was to investigate the optimal pH andfermentation time for SCFAs accumulation in ultrasonic-pretreatedsludge fermentation process. Of interest, the composition of SCFAswas examined, and the mechanisms of SCFAs improvement in thiscase were discussed. Simultaneously, the impacts of alkalineadjustment on the microbial community structure were analyzedby PCR-DGGE.
2. Materials and methods
2.1. Experimental materials
The WAS used in this study was collected from the municipalwastewater treatment plant of Taiping located in Harbin, China. Thesludge was settled at 4�C for 2 h with supernatant discharge, andthe main characteristics of WAS are summarized in Table 1. Theinoculated sludge was collected from an upflow anaerobic sludgeblanket (UASB) reactor in our laboratory.
Sludge was pretreated by an ultrasonicator (JY98-IIIN, XinzhiInc., China) at a frequency of 20 kHz, with ultrasonic density 1.0 W/mL and ultrasonic time 10 min (Yan et al., 2011). The ultrasonic-pretreated sludge was stored at 4 �C for the fermentation tests.
2.2. Batch fermentation tests
The effect of alkaline adjustment on anaerobic fermentation ofultrasonic-pretreated sludge was performed in five stirred tankreactors. 700 mL ultrasonic-pretreated sludge and 100 mL inocu-lated sludge were added into 1 L reactor. The pH values in reactorswere respectively adjusted and maintained at 7, 8, 9 and 10 using1.0 M hydrochloric acid (HCl) or 1.0 M sodium hydroxide (NaOH)solutions, which was automatically added into reactors accordingto the pH changes. Simultaneously, parallel reactor in the same runwas also operated with the same sludge without any pH adjust-ment. All reactors were flushed with nitrogen gas (N2) for 2 minbefore seal. During the fermentation process, all reactors wereincubated at 20 ± 2 �C and continuously stirred at a rate of 70 rpmto sustain homogenous mixing. The sludge samples were collectedto monitor its main chemical characteristics.
2.3. Analytical methods
All analyses on conventional parameters were measured ac-cording to Standard Methods (APHA, 1976) except for proteins,carbohydrates, SCFAs and protease activity. The sludge samplesfrom reactors were first centrifuged at 10, 000 rpm for 10 min, andthen filtered through 0.45 mm membrane filters. The filtrate wasimmediately analyzed for soluble proteins, soluble carbohydrates,
Table 1Characteristics of the WAS.a
Parameters Value
pH 6.8TSS (total suspended solid) 10,876.8VSS (volatile suspended solid) 8107.4TCOD (total chemical oxygen demand) 11,343.6SCOD (soluble chemical oxygen demand) 73.4Total proteins (as COD) 6339.1Soluble proteins (as COD) 26.3Total carbohydrates (as COD) 1042.7Soluble carbohydrates (as COD) 7.5
a All values are expressed in mg/L except pH. Values are the averages of de-terminations taken over in duplicates.
and SCFAs. Soluble proteins were determined by the Lowry-Folinmethod with bovin serum albumin (BSA) as the standard (Lowryet al., 1951), while soluble carbohydrates were analyzed by theanthrone sulfuric method with glucose as the standard (Morris,1948). The total proteins content was estimated from the corre-sponding TKN concentration by subtracting the inorganic nitrogenconcentration and dividing the difference by 0.16, then multiplyingthe result by 1.5 (Miron et al., 2000). Protease activity was quan-titatively analyzed according to a standard procedurewith casein asa substrate and L-tyrosine as standard (Nabarlatz et al., 2010). Thetotal SCFAs, including acetic, propionic, n-butyric, iso-butyric, n-valeric and iso-valeric acids, were determined based on gas chro-matography (Agilent 6890 N) with a flame ionization detector (FID)and DB-WAXETR column (30 m � 0.53 mm � 1.0 mm) (Chen et al.,2007).
2.4. DNA extraction
Total DNA was extracted from each bioreactor biomass on thefifteenth day using the E.Z.N.A Soil DNA Kit (Omega Bio-Tek. Inc.).Prior to extraction, samples of suspended biomass were concen-trated by centrifuge at 10, 000 rpm for 5 min. Each extraction wasperformed in duplicate and the extracted DNA was eluted in Tris-HCl buffer (pH 8.0) and stored at �20�C.
2.5. PCR and DGGE
The V3 region was amplified by PCR using the forward primer338F (50- ACTCCTACGGGAGGCAGCAG-30) and the reverse primer534R (50- ATTACCGCGGCTGCTGG-30) with a GC clamp (50-CGCCCGCCGCGCCCCGCGCCCGGCCCGCCGCCCCCGCCCC-30) at the 50
end. The detailed steps of PCR amplification followed literature(Shin et al., 2010). DGGE was performed with the DCode™ Uni-versal Mutation Detection system (Biorad Laboratories, Hercules,CA, USA). In this experiment, denaturing gradient ranged from 40%to 60% and electrophoresis was carried out at a constant voltage of140 V for 7 h. After electrophoresis, the gel was stained withethidium bromide and scanned under UV transillumination (Kimet al., 2011b).
2.6. Cloning of 16S rRNA fragments
Bands of interest were excised from the gel for furthersequencing and phylogenetic analyses. Each gel fragment wascrushed, and the DNA was resolved in 50 mL of sterile deionizedwater at 4 �C. PCR was performed with the primer 338F and 534R.The PCR fragments were purified using E.Z.N.A Cycle Pure Kit(Omega Bio-Tek. Inc.) and cloned in Escherichia.coli DH5a (Takara,Dalian, China) using PMD19-T Vector (Takara, Dalian, China) ac-cording to the manufacturer's instruction.
2.7. DNA sequencing and analysis
16S rRNA gene fragment clones were sequenced. A search forthe closest reference microorganisms in the GenBank databaseusing the partial 16S rRNA sequences was carried out with theBLAST program (http://www.ncbi.nlm.nih.gov/blast), and RDPclassifier (http://rdp.cme.msu.edu/classifier/classifier.jsp).
3. Results and discussions
3.1. WAS hydrolysis
In this study, proteins and carbohydrates were major compo-nents of WAS, accounting for 55.9% and 9.2%. The change of
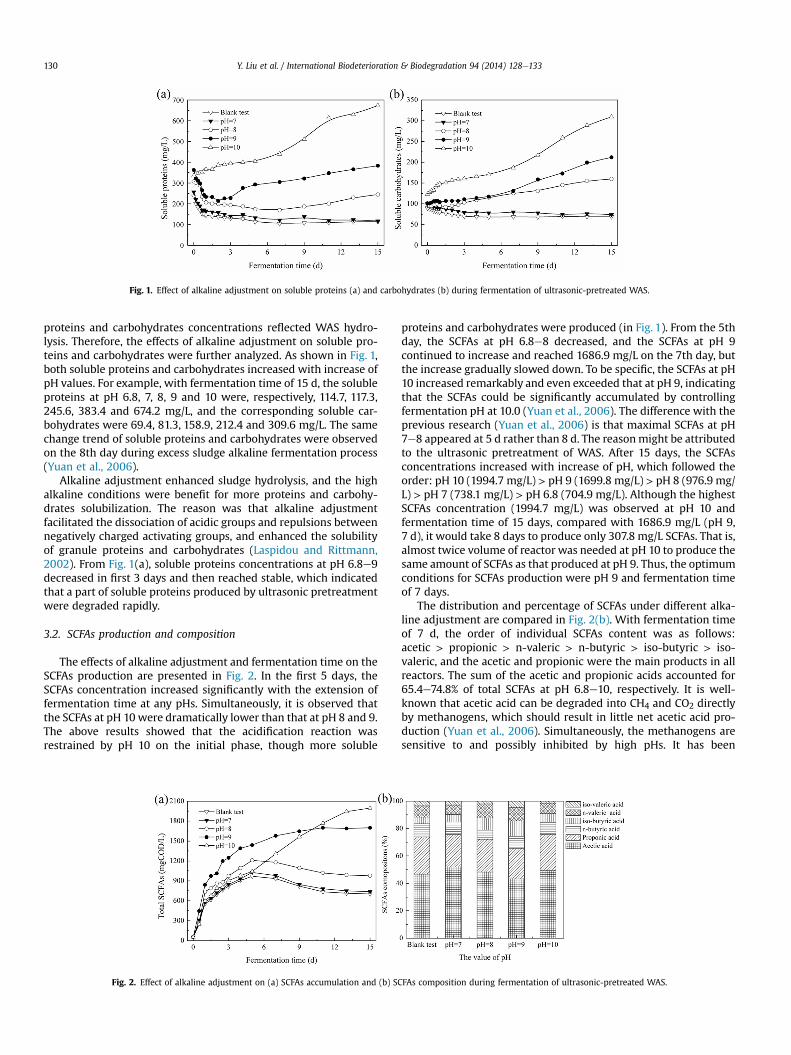
Fig. 1. Effect of alkaline adjustment on soluble proteins (a) and carbohydrates (b) during fermentation of ultrasonic-pretreated WAS.
Y. Liu et al. / International Biodeterioration & Biodegradation 94 (2014) 128e133130
proteins and carbohydrates concentrations reflected WAS hydro-lysis. Therefore, the effects of alkaline adjustment on soluble pro-teins and carbohydrates were further analyzed. As shown in Fig. 1,both soluble proteins and carbohydrates increased with increase ofpH values. For example, with fermentation time of 15 d, the solubleproteins at pH 6.8, 7, 8, 9 and 10 were, respectively, 114.7, 117.3,245.6, 383.4 and 674.2 mg/L, and the corresponding soluble car-bohydrates were 69.4, 81.3, 158.9, 212.4 and 309.6 mg/L. The samechange trend of soluble proteins and carbohydrates were observedon the 8th day during excess sludge alkaline fermentation process(Yuan et al., 2006).
Alkaline adjustment enhanced sludge hydrolysis, and the highalkaline conditions were benefit for more proteins and carbohy-drates solubilization. The reason was that alkaline adjustmentfacilitated the dissociation of acidic groups and repulsions betweennegatively charged activating groups, and enhanced the solubilityof granule proteins and carbohydrates (Laspidou and Rittmann,2002). From Fig. 1(a), soluble proteins concentrations at pH 6.8e9decreased in first 3 days and then reached stable, which indicatedthat a part of soluble proteins produced by ultrasonic pretreatmentwere degraded rapidly.
3.2. SCFAs production and composition
The effects of alkaline adjustment and fermentation time on theSCFAs production are presented in Fig. 2. In the first 5 days, theSCFAs concentration increased significantly with the extension offermentation time at any pHs. Simultaneously, it is observed thatthe SCFAs at pH 10 were dramatically lower than that at pH 8 and 9.The above results showed that the acidification reaction wasrestrained by pH 10 on the initial phase, though more soluble
Fig. 2. Effect of alkaline adjustment on (a) SCFAs accumulation and (b) SC
proteins and carbohydrates were produced (in Fig. 1). From the 5thday, the SCFAs at pH 6.8e8 decreased, and the SCFAs at pH 9continued to increase and reached 1686.9 mg/L on the 7th day, butthe increase gradually slowed down. To be specific, the SCFAs at pH10 increased remarkably and even exceeded that at pH 9, indicatingthat the SCFAs could be significantly accumulated by controllingfermentation pH at 10.0 (Yuan et al., 2006). The difference with theprevious research (Yuan et al., 2006) is that maximal SCFAs at pH7e8 appeared at 5 d rather than 8 d. The reasonmight be attributedto the ultrasonic pretreatment of WAS. After 15 days, the SCFAsconcentrations increased with increase of pH, which followed theorder: pH 10 (1994.7 mg/L) > pH 9 (1699.8 mg/L) > pH 8 (976.9 mg/L) > pH 7 (738.1 mg/L) > pH 6.8 (704.9 mg/L). Although the highestSCFAs concentration (1994.7 mg/L) was observed at pH 10 andfermentation time of 15 days, compared with 1686.9 mg/L (pH 9,7 d), it would take 8 days to produce only 307.8 mg/L SCFAs. That is,almost twice volume of reactor was needed at pH 10 to produce thesame amount of SCFAs as that produced at pH 9. Thus, the optimumconditions for SCFAs production were pH 9 and fermentation timeof 7 days.
The distribution and percentage of SCFAs under different alka-line adjustment are compared in Fig. 2(b). With fermentation timeof 7 d, the order of individual SCFAs content was as follows:acetic > propionic > n-valeric > n-butyric > iso-butyric > iso-valeric, and the acetic and propionic were the main products in allreactors. The sum of the acetic and propionic acids accounted for65.4e74.8% of total SCFAs at pH 6.8e10, respectively. It is well-known that acetic acid can be degraded into CH4 and CO2 directlyby methanogens, which should result in little net acetic acid pro-duction (Yuan et al., 2006). Simultaneously, the methanogens aresensitive to and possibly inhibited by high pHs. It has been
FAs composition during fermentation of ultrasonic-pretreated WAS.
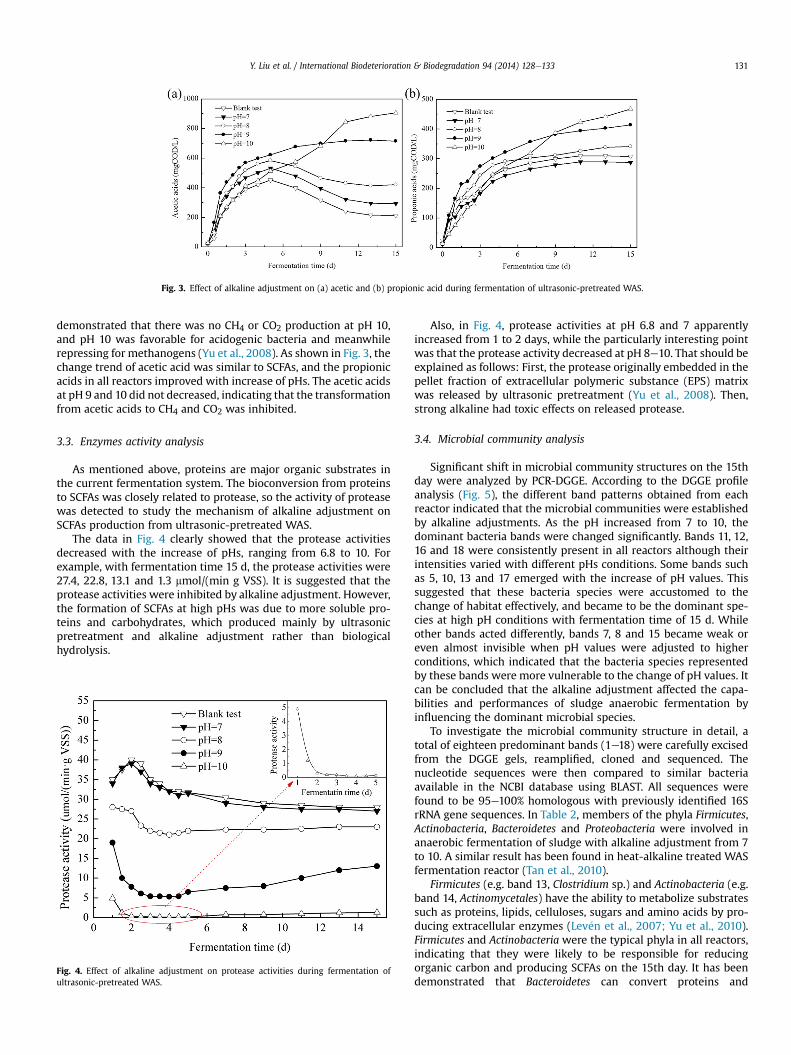
Fig. 3. Effect of alkaline adjustment on (a) acetic and (b) propionic acid during fermentation of ultrasonic-pretreated WAS.
Y. Liu et al. / International Biodeterioration & Biodegradation 94 (2014) 128e133 131
demonstrated that there was no CH4 or CO2 production at pH 10,and pH 10 was favorable for acidogenic bacteria and meanwhilerepressing for methanogens (Yu et al., 2008). As shown in Fig. 3, thechange trend of acetic acid was similar to SCFAs, and the propionicacids in all reactors improved with increase of pHs. The acetic acidsat pH 9 and 10 did not decreased, indicating that the transformationfrom acetic acids to CH4 and CO2 was inhibited.
3.3. Enzymes activity analysis
As mentioned above, proteins are major organic substrates inthe current fermentation system. The bioconversion from proteinsto SCFAs was closely related to protease, so the activity of proteasewas detected to study the mechanism of alkaline adjustment onSCFAs production from ultrasonic-pretreated WAS.
The data in Fig. 4 clearly showed that the protease activitiesdecreased with the increase of pHs, ranging from 6.8 to 10. Forexample, with fermentation time 15 d, the protease activities were27.4, 22.8, 13.1 and 1.3 mmol/(min g VSS). It is suggested that theprotease activities were inhibited by alkaline adjustment. However,the formation of SCFAs at high pHs was due to more soluble pro-teins and carbohydrates, which produced mainly by ultrasonicpretreatment and alkaline adjustment rather than biologicalhydrolysis.
Fig. 4. Effect of alkaline adjustment on protease activities during fermentation ofultrasonic-pretreated WAS.
Also, in Fig. 4, protease activities at pH 6.8 and 7 apparentlyincreased from 1 to 2 days, while the particularly interesting pointwas that the protease activity decreased at pH 8e10. That should beexplained as follows: First, the protease originally embedded in thepellet fraction of extracellular polymeric substance (EPS) matrixwas released by ultrasonic pretreatment (Yu et al., 2008). Then,strong alkaline had toxic effects on released protease.
3.4. Microbial community analysis
Significant shift in microbial community structures on the 15thday were analyzed by PCR-DGGE. According to the DGGE profileanalysis (Fig. 5), the different band patterns obtained from eachreactor indicated that the microbial communities were establishedby alkaline adjustments. As the pH increased from 7 to 10, thedominant bacteria bands were changed significantly. Bands 11, 12,16 and 18 were consistently present in all reactors although theirintensities varied with different pHs conditions. Some bands suchas 5, 10, 13 and 17 emerged with the increase of pH values. Thissuggested that these bacteria species were accustomed to thechange of habitat effectively, and became to be the dominant spe-cies at high pH conditions with fermentation time of 15 d. Whileother bands acted differently, bands 7, 8 and 15 became weak oreven almost invisible when pH values were adjusted to higherconditions, which indicated that the bacteria species representedby these bands were more vulnerable to the change of pH values. Itcan be concluded that the alkaline adjustment affected the capa-bilities and performances of sludge anaerobic fermentation byinfluencing the dominant microbial species.
To investigate the microbial community structure in detail, atotal of eighteen predominant bands (1e18) were carefully excisedfrom the DGGE gels, reamplified, cloned and sequenced. Thenucleotide sequences were then compared to similar bacteriaavailable in the NCBI database using BLAST. All sequences werefound to be 95e100% homologous with previously identified 16SrRNA gene sequences. In Table 2, members of the phyla Firmicutes,Actinobacteria, Bacteroidetes and Proteobacteria were involved inanaerobic fermentation of sludge with alkaline adjustment from 7to 10. A similar result has been found in heat-alkaline treated WASfermentation reactor (Tan et al., 2010).
Firmicutes (e.g. band 13, Clostridium sp.) and Actinobacteria (e.g.band 14, Actinomycetales) have the ability to metabolize substratessuch as proteins, lipids, celluloses, sugars and amino acids by pro-ducing extracellular enzymes (Lev�en et al., 2007; Yu et al., 2010).Firmicutes and Actinobacteria were the typical phyla in all reactors,indicating that they were likely to be responsible for reducingorganic carbon and producing SCFAs on the 15th day. It has beendemonstrated that Bacteroidetes can convert proteins and

Fig. 5. Denaturing gradient gel electrophoresis (DGGE) profiles of the V3 region of the16S rRNA gene amplified with primers 338F-GC and 534R from DNA samples.
Y. Liu et al. / International Biodeterioration & Biodegradation 94 (2014) 128e133132
carbohydrates to propionate and acetate as its primary products insludge anaerobic fermentation (Ueki et al., 2006). Therefore, Bac-teroidetes might be involved in the SCFAs production with solubleproteins and carbohydrates as the main substrates. Proteobacteriawas major phyla in hydrolysis and acidification process ofultrasonic-pretreated WAS (Kang et al., 2011). In this study, Pro-teobacteria was prevalent in anaerobic reactors at high pHs, indi-cating that it could adapt well to the alkaline conditions andparticipate in anaerobic fermentation process. As shown in Table 2,
Table 2The matched species of 16S rRNA sequences in GenBank.
Bands Closest relative (accession no.) Phyla Identity
1 Erythrobacter citreus(HQ908700.1)
Alphaproteobacteria 100%
2 TM7 phylum sp.(AF385568.1) TM 7 98%3 Uncultured Chloroflexi bacterium
(EF613976.1)Chloroflexi 100%
4 Soehngenia sp.(HQ133002.1) Firmicutes 98%5 Acinetobacter xiamenensis
(FJ477061.1)Gammaproteobacteria 99%
6 Dysgonomonas sp. (AB547446.1) Bacteroidetes 100%7 Rhodocyclaceae bacterium
(AB174824.1)Betaproteobacteria 97%
8 Enterococcus aquimarinus (GQ337015.1) Firmicutes 99%9 Uncultured Actinobacteria bacterium
(CU926454.1)Actinobacteria 100%
10 Uncultured alpha proteobacterium(FJ485092.1)
Alphaproteobacteria 100%
11 Petrimonas sulfuriphila (AY570690.1) Bacteroidetes 95%12 Acetoanaerobium noterae (GU562448.1) Firmicutes 98%13 Clostridium sp.(GU370098.1) Firmicutes 100%14 Uncultured Actinomycetales bacterium
(FJ946539.1)Actinobacteria 98%
15 Uncultured Bacteroides sp.(FM204979.1) Bacteroidetes 100%16 Uncultured Firmicutes bacterium
(CU922223.1)Firmicutes 97%
17 Uncultured Firmicutes bacterium(CU926242.1)
Firmicutes 98%
18 Alcaligenaceae bacterium (HQ703917.1) Betaproteobacteria 100%
band 1 closely related to Erythrobacter citreus was only observed atpH 8. It is hypothesized that E. citreus might be involved in thedecrease of acetic acid, because some members of Proteobacteriahas been reported to degrade organic acids through denitrificationin the fermentation process (Nelson et al., 2011).
4. Conclusions
The experiment demonstrated that alkaline adjustment canaccelerate the sludge hydrolysis, restrain acetic acid consumption,and promote SCFAs accumulation. The concluding remarks of thisstudy are outlined as follows:
� Alkaline adjustment enhanced hydrolysis of proteins and car-bohydrates and the production of SCFAs. The optimum condi-tions for SCFAs productionwere pH 9 and fermentation time of 7days.
� Protease activity was inhibited by alkaline adjustment in therange of pH 8e10, indicating the SCFAs production was affectednot only by enzyme activity, but also was related to sludgedisintegrated degree.
� Firmicutes, Proteobacteria and Bacteroidetes participated inanaerobic fermentation of ultrasonic-pretreated WAS andcontributed to proteins and carbohydrates biodegradation andSCFAs production, simultaneously, some bacteria could surviveunder the high alkaline environments.
Acknowledgments
This work was financially supported by the National HighTechnology Research and Development Program of China (863Program) (NO. 2012AA063503-02). Special thanks to those peoplein Harbin Institute of Technology University who provided help tothis paper.
References
APHA, 1976. Standard Methods for the Examination of Water and Wastewater, 14thed. American Public Health Association, Washington, DC.
Chen, Y., Jiang, S., Yuan, H., Zhou, Q., Gu, G., 2007. Hydrolysis and acidification ofwaste activated sludge at different pHs. Water Res. 41, 683e689.
Cokgor, E.U., Oktay, S., Tas, D.O., Zengin, G.E., Orhon, D., 2009. Influence of pH andtemperature on soluble substrate generation with primary sludge fermentation.Bioresour. Technol. 100, 380e386.
Erden, G., Filibeli, A., 2010. Improving anaerobic biodegradability of biologicalsludges by Fenton pre-treatment: effects on single stage and two-stageanaerobic digestion. Desalination 251, 58e63.
Gonzalez del Campo, A., Ca~nizares, P., Lobato, J., Rodrigo, M.A., Fernandez, F.J., 2012.Electricity production by integration of acidogenic fermentation of fruit juicewastewater and fuel cells. Int. J. Hydrog. Energy 37, 9028e9037.
Jia, Q., Wang, H., Wang, X., 2013. Dynamic synthesis of polyhydroxyalkanoates bybacterial consortium from simulated excess sludge fermentation liquid. Bio-resour. Technol. 140, 328e336.
Kang, X.R., Zhang, G.M., Chen, L., Dong, W.Y., Tian, W.D., 2011. Effect of initial pHadjustment on hydrolysis and acidification of sludge by ultrasonic pretreat-ment. Ind. Eng. Chem. Res. 50, 12372e12378.
Kim, D.H., Jeong, E., Oh, S.E., Shin, H.S., 2010. Combined (alkaline þ ultrasonic)pretreatment effect on sewage sludge disintegration. Water Res. 44,3093e3100.
Kim, D.H., Kim, S.H., Jung, K.W., Kim, M.S., Shin, H.S., 2011a. Effect of initial pH in-dependent of operational pH on hydrogen fermentation of food waste. Bio-resour. Technol. 102, 8646e8652.
Kim, J., Shin, S.G., Han, G., O'Flaherty, V., Lee, C., Hwang, S., 2011b. Common keyacidogen populations in anaerobic reactors treating different wastewaters:molecular identification and quantitative monitoring. Water Res. 45,2539e2549.
Kim, J., Yu, Y., Lee, C., 2013. Thermo-alkaline pretreatment of waste activated sludgeat low-temperatures: effects on sludge disintegration, methane production, andmethanogen community structure. Bioresour. Technol. 144, 194e201.
Laspidou, C.S., Rittmann, B.E., 2002. A unified theory for extracellular polymericsubstances, soluble microbial products, and active and inert biomass. WaterRes. 36, 2711e2720.

Y. Liu et al. / International Biodeterioration & Biodegradation 94 (2014) 128e133 133
Lev�en, L., Eriksson, A.R., Schnürer, A., 2007. Effect of process temperature on bac-terial and archaeal communities in two methanogenic bioreactors treatingorganic household waste. FEMS Microbiol. Ecol. 59, 683e693.
Liu, H., Wang, J., Liu, X., Fu, B., Chen, J., Yu, H.Q., 2012. Acidogenic fermentation ofproteinaceous sewage sludge: effect of pH. Water Res 46, 799e807.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem. 193, 265e275.
Luste, S., Vilhunen, S., Luostarinen, S., 2011. Effect of ultrasound and addition ofbacterial product on hydrolysis of by-products from the meat-processing in-dustry. Int. Biodeterior. Biodegrad. 65, 318e325.
Miron, Y., Zeeman, G., van Lier, J.B., Lettinga, G., 2000. The role of sludge retentiontime in the hydrolysis and acidification of lipids, carbohydrates and proteinsduring digestion of primary sludge in CSTR systems. Water Res. 34, 1705e1713.
Morris, D.L., 1948. Quantitative determination of carbohydrates with Dreywood'santhrone reagent. Science 107, 254e255.
Nabarlatz, D., Vondrysova, J., Jenicek, P., Stüber, F., Font, J., Fortuny, A., Fabregat, A.,Bengoa, C., 2010. Hydrolytic enzymes in activated sludge: extraction of proteaseand lipase by stirring and ultrasonication. Ultrason. Sonochem. 17, 923e931.
Nelson, M.C., Morrison, M., Yu, Z., 2011. A meta-analysis of the microbial diversityobserved in anaerobic digesters. Bioresour. Technol. 102, 3730e3739.
Oh, M.H., Lee, S.M., Hong, S.H., Choi, H.N., Lee, E.Y., 2013. Monitoring of Lactobacillussp. inoculated in the reactor to evaluate the solubilization efficiency of primarysludge. Int. Biodeterior. Biodegrad. 85, 603e607.
Park, K.Y., Kweon, J., Chantrasakdakul, P., Lee, K., Cha, H.Y., 2013. Anaerobic digestionof microalgal biomass with ultrasonic disintegration. Int. Biodeterior. Bio-degrad. 85, 598e602.
Shin, S.G., Han, G., Lim, J., Lee, C., Hwang, S., 2010. A comprehensive microbialinsight into two-stage anaerobic digestion of food waste-recycling wastewater.Water Res. 44, 4838e4849.
Tan, R., Miyanaga, K., Toyama, K., Uy, D., Tanji, Y., 2010. Changes in composition andmicrobial communities in excess sludge after heat-alkaline treatment andacclimation. Biochem. Eng. J. 52, 151e159.
Ueki, A., Akasaka, H., Suzuki, D., Ueki, K., 2006. Paludibacter propionicigenes gen.nov., sp. nov., a novel strictly anaerobic, Gram-negative, propionate-producingbacterium isolated from plant residue in irrigated rice-field soil in Japan. Int. J.Syst. Evol. Microbiol. 56, 39e44.
Wu, H., Yang, D., Zhou, Q., Song, Z., 2009. The effect of pH on anaerobic fermen-tation of primary sludge at room temperature. J. Hazard. Mater. 172, 196e201.
Yan, Y., Chen, H., He, Q., Tan, X., Dai, X., Zhou, Q., 2011. Effect of ultrasonic time onwaste activated sludge hydrolysis and volatile fatty acids accumulation underalkaline condition. Fresen. Environ. Bull. 20, 2861e2868.
Yan, Y., Feng, L., Zhang, C., Wisniewski, C., Zhou, Q., 2010. Ultrasonic enhancement ofwaste activated sludge hydrolysis and volatile fatty acids accumulation at pH10.0. Water Res. 44, 3329e3336.
Yu, G.H., He, P.J., Shao, L.M., He, P.P., 2008. Toward understanding the mechanism ofimproving the production of volatile fatty acids from activated sludge at pH10.0. Water Res. 42, 4637e4644.
Yu, Z., Morrison, M., Schanbacher, F.L., 2010. Production and utilization of methanebiogas as renewable fuel. In: Biomass to Biofuels: Strategies for Global In-dustries. Wiley, Chichester, UK, pp. 403e413.
Yuan, H., Chen, Y., Zhang, H., Jiang, S., Zhou, Q., Gu, G., 2006. Improved bio-production of short-chain fatty acids (SCFAs) from excess sludge under alkalineconditions. Environ. Sci. Technol. 40, 2025e2029.
Zhu, X., Chen, Y., 2011. Reduction of N2O and NO generation in anaerobic-aerobic(low dissolved oxygen) biological wastewater treatment process by usingsludge alkaline fermentation liquid. Environ. Sci. Technol. 45, 2137e2143.