settlement and metamorphosis of red abalone (haliotis rufescens
TRANSCRIPT

Settlement and Metamorphosis of Red Abalone
(Haliotis rufescens) Larvae: A Critical Examination
of Mucus, Diatoms, and ~-Aminobutyric Acid
(GABA) as Inductive Substrates
A Thesis
Presented to
The Faculty of the Department of Biology
San Jose State University
In Partial Fulfillment
of the Requirements for the Degree
Master of Arts
By
Marc Slattery
December, 1987

iii
Abstract
Settlement and metamorphosis of red abalone,
~~~~~~ larvae in the presence of three inductive cues
(mucus, diatoms, and j'-aminobutyric acid) was tested
without the use of antibiotics. Larval settlement differed
between substrates. Mucus from juvenile abalones yi
significantly higher settlement. Settlement varied
during the year and was highest between August and mid
September. Metamorphosis and survival (to the development
of the first respiratory pore) was variable among the
substrates. At 11 weeks, approximately 50 %, 20 %, and 0 %
of the larvae had survived on mucous, diatom, and GABA
substrates, respectively. In all treatments an initial high
rate of mortality and stunting of some larvae suggested the
abalone were feeding inefficiently.

iv
Acknowledgements
I would like to express my sincerest gratitude to the
many people who provided support and encouragement through
out the course of my research and manuscript preparation.
I am indebted to the members of my committee: Dr. James w.
Nybakken, Dr. Michael S. Foster, and Dr. Gregor M. cailliet,
who provided invaluable experience, time, and patience in
critical review of my thesis. Dr. Daniel E. Morse's
enlightening comments are also appreciated. My deepest
gratitude is extended to Mr. Earl E. Ebert, of the
Fish and Game Marine Culture Lab, for sharing his insights
and ideas on abalone culture.
Thanks to Dr. Phil Law and Allen Grover for their
technical assistance. Special thanks to the staff of the
Marine Culture Lab for their continued cooperation. The
help of Lynn McMasters, and the faculty, staff, and students
of Moss Landing Marine Labs is gratefully acknowledged.
My family and friends have been a source of constant
encouragement for which I am extremely grateful. Thanks
to Dr. Roy s. Houston who taught me to look at "la pintura
grande." Finally, this thesis is dedicated in loving memory
to Kim Peppard.

Abstract
Acknowledgements
List of Tables
Table of contents
List of Figures
Introduction
Materials and Methods
Results
Discussion
References
Tables
Figures
Appendix A.
Appendix B.
v
Page
iii
iv
vi
vii
1
7
12
14
21
27
30
32
33

1.
2.
3 •
List of Tables
ANOVA Summary Table showing the effects of water treatment (FSW and FSW + GABA), substrate (clean plastic, diatoms, 24 hour mucus, and 72 hour mucus), period, and interactions (between treatment, substrate, and period) on larval settlement.
Tukey's Studentized Range Groupings for mean larval settlement with respect to substrate (a), and period (b).
ANOVA Summary Table showing the effects of substrate (diatoms, diatoms + GABA, and diatoms + mucus), period, and interactions (between substrate and period) on larval survival.
vi
27
28
29

1.
2.
vii
List of Figures
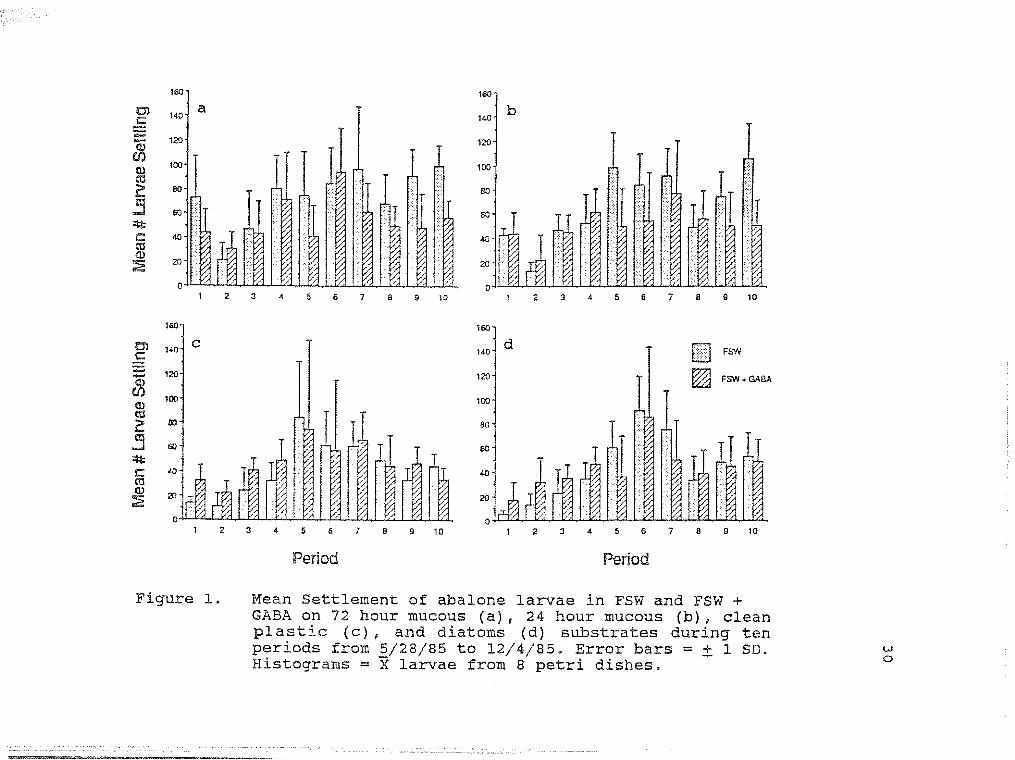
Mean Settlement of abalone larvae in 30 FSW and FSW + GABA on 72 hour mucous (a), 24 hour mucous (b), clean plast (c), and diatoms (d) substrates during ten periods from 5/28/85 to 12/4/85.
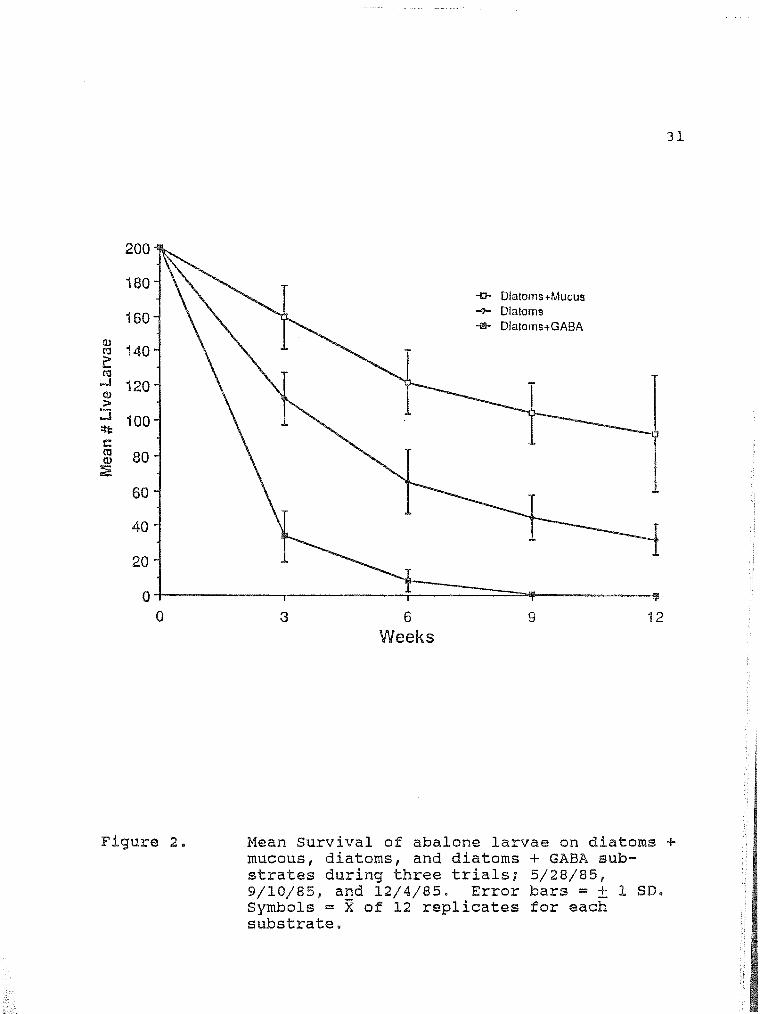
Mean Survival of abalone larvae on diatoms 31 + mucous, diatoms, and diatoms + GABA substrates during three trials; 5/28/85, 9/10/85, and 12/4/85.

1
Introduction
A critical stage in the life history of marine inverte
brate larvae occurs during the termination of the planktonic
or dispersive stage. The transformation from larva to
juvenile involves two distinct processes: settlement and
metamorphosis (Chia, 1978; Crisp, 1974; Hadfield, 1984).
Settlement has been described as a behavioral change
typically characterized by the active searching for and
orientation to certain environmental factors (Crisp, 1974;
Hadfield, 1984). Metamorphosis is a non-reversible state
that involves anatomical and physiological changes in the
organism (Bonar, 1976; Scheltema, 1974).
The specific factors involved in larval settlement and
metamorphosis are quite variable (see for example Burke,
1983). However, the classic model appears to be a simple
stimulusjresponse system, possibly under some neuronal
control (Chia, 1978; Hadfield, 1978). Larvae grow until
they reach a time of "competence" that enables them to
respond to certain environmental cues (Burke, 1983). This
typically corresponds to development of sensory structures
by the organisms (Bonar, 1976; Crisp, 1974; Morita, 1972;
Morse et al., 1979a).

Settlement is affected by several factors including
age, diet, and physio-chemical characteristics of the
available substrate (Hadfield, 1984). In the presence
of the requisite stimulus the larva responds with species
specific tissue morphogenesis (Bonar, 1976). Metamorphos
2
can be delayed if the correct environmental cues are not
encountered (Burke, 1983; crisp, 1974). It is the nature of
the cues that triggers settlement and metamorphosis in the
larva (Baloun and Morse, 1984).
The red abalone, Haliotis rufescens, is highly prized
as food for humans, much research has been done on its
commercial culture potential (Ault, 1982; Kan-no, 1975;
Kikuchi and Uki. 1974; Leighton, 1972; Leighton et al.,
1981; Morse et al., 1977 and 1979a; Seki and Kan-no, 1977
and 198la). According to Ault (1982), Horse et (1979a)
and Mottet (1978), red abalone larvae, held in cultures at
15°C, become competent within 6-7 days following the
development of eyespots, a muscular foot, and cephalic
tentacles. Morse et (1979a; 1979b) noted that mari-
culture of this species was not economically feasible
because the processes of settlement and metamorphos were
retarded in the absence of a naturally required specific
biochemical cue. This resulted in high rates of mortality

when the larva's yolk supplies were exhausted. However,
juvenile abalone {l-20 mm) were most commonly observed in
natural "nursery habitats" of crustose red algae including
Lithophyllum, Lithotharnnion, and Hildenbrandia, suggesting
the settling cues might be associated with these species
(Morse et al., 1979a; 1980). Many invertebrate species,
including tubeworms (Gee, 1965), chitons (Barnes and Gonar,
1973; Rumrill and cameron, 1983), limpets (Steneck, 1982),
and asteroids (Barker, 1977) have also been reported to
settle preferentially on crustose red algae.
Morse et al. (l979a; 1980) subsequently discovered
that a /-aminobutyric acid (GABA) mimetic peptide and
phycoerythobilin were the settlement inducing agents
sequestered at the surface of the crustose red algae.
Foliose reds, greens, browns, and cyanobacteria were also
found to contain a GABA mimetic peptide and biliproteins
(Morse et ~~ 1984; Morse and Morse, 1984). However, the
larvae required contact with the biochemical cues in order
to trigger settlement and metamorphosis (Morse et
1980) . The morphogenetic inductive molecules were freed
during mucus sloughing by the epithelial cells of crustose
reds and contact occurred when larvae randomly tested the
substrate (Morse and Morse, 1984). The requirement of
3

4
larval contact assured that the abalone would recruit to a
desirable habitat which provided inductive molecules,
shelter, camouflaging pigments, and nutrition (Morse et al.,
1979a; 1979b; 1980).
Three methods have been most often used to initiate
settlement and metamorphosis. The first involved the use
of a 1 ~M solution of GABA in filtered seawater. In the
presence of antibiotics, Morse et (1979a; l979b)
achieved 93% settlement and metamorphosis of abalone larvae
at that concentration. They noted that higher concentra
tions inhibited metamorphosis and lower concentrations
slowed the process. However, in the absence of antibiotics,
complete mortality of the larvae was reported (Morse et al.,
1979b). A second method used diatom mats, the predominant
diet of newly settled larvae, as the inductive substrate
(Grant, 1981; Ina, 1966; Ebert and Houk, 1984). An average
of 7.5% of the larvae settled, by the California Department
of Fish and Game's Marine Culture Lab, on diatom mats
survived to 3 months age (Ebert and Houk, 1984). Finally,
the mucous material secreted by the foot of juvenile and
adult abalones has been used to successfully induce settle
ment and metamorphosis of larval abalone in Japanese
hatcheries with a 12% survival rate (Kan-no, 1975; Seki,

1980; Seki and Kan-no, l98lb).
The advantage of GABA for use in rnariculture is the
high percentage of metamorphosis, whereas the advantage of
diatoms or mucus is that they are simple, inexpensive
methods. However, there is little agreement within the
scientific community as to which method is best for use in
rnariculture. Akashige et al. (1981) reported that GABA
actually narcotized the velar cilia of the planktonic
5
larvae causing the veligers to "fall" out of solution and
die on the substrate. Morse (pers. corn.) noted the "weakly
inductive" potential of mucus, but suggested that mucus
was a breeding ground for a "microbial overgrowth" that
could kill the larvae. To further complicate the situation,
there has been little consistency in experimental design
among the investigators, making comparisons difficult.
Results were obtained from experiments involving several
hundred (Morse et al., 1979b) to several million (Grant,
1981) larvae in vessels of variable volumes (Mottet, 1978).
The importance in determining the best substrate for
settlement and metamorphosis of abalone larvae is unequiv
ocal. In addition, the need for standardization in determi
nation of a "best method" is clear. The purpose of this
study was to compare settlement, metamorphosis, and

6
survival rates of red abalone larvae on mucous, diatom, and
clean substrates in the presence and absence of GABA without
the benefit of antibiotics. The experimental design
addresses the need for development of low cost mariculture
systems through questions of settlement substrate pref
erence, inductive cue strength, and survival rates of
abalone larvae in standard culture vessels.

7
Materials and Methods
The studies were conducted at the California Department
of Fish and Game's Marine Culture Laboratory over an eight
month period from June 1985 to February 1986. The lab is
located at Granite canyon, an exposed section of coastline,
approximately 12 miles south of Monterey. The existing
seawater system, assembled by Ebert et al. (1974), was
modified with the addition of 5, 3, 1, and 0.5 micron
in-line cartridge filters. This effectively reduced culture
contamination by copepods, nematodes and bacteria. Lab
oratory broodstock were utilized for all larval production
(Ebert and Houk, 1984).
Abalone were spawned with the ultraviolet irradiated
seawater technique described by Kikuchi and Uki (1974). The
ova were fertilized, washed, and left to develop to the
veliger stage for 30 hours in 15°C UV treated seawater
(Ebert and Hamilton, 1983). The "healthiest" larvae, as
determined by their swimming behavior, were held in con
centrations of 5 per ml in 8 liter culture tubes and
maintained for 6-7 days at 15°C (Ebert and Houk, 1984).
The competent larvae, which exhibited the characteristic
behavioral (exploration and orientation) and morphological

8
(sense organ development and cephalic tentacle protrusion)
changes described by Mottet (1978) and Seki and Kan-no
{l98la), were utilized in the experiments. Subsamples of
larvae (McCallum, 1979) from the cultures, were collected to
estimate total numbers of individuals in the ten substrate
preference trials (5/28/85, 6/17/85, 7/1/85, 7/15/85,
8/5/85, 8/20/85, 9/10/85, 9/23/85, 10/6/85, and 12/4/85) and
three survival trials (5/28/85, 9/10/85, and 12/4/85).
Settlement
The substrate preference of settling abalone larvae was
tested in two plastic troughs. Each settling trough was
147.32 em long, 30.48 em wide, and 25.40 em deep. Seawater
filtered (FSW) through 1 micron filters at 15°C was added
to each trough. A solution of GABA was added to one trough
to a final concentration of 1 pM, as described by Morse
al. (1979b), but without the antibiotics.
Substrates for these troughs were prepared by first
placing 64 plastic petri dishes 60.96 em below two Chroma-50
flourescent tubes. Forty eight dishes were innoculated with
100 ml of a diatom slurry solution (Ebert and Houk, 1984).
The remaining 24 dishes were innoculated with 100 ml of

filtered seawater. The diatoms settled within three days
and coated the petri dishes with a thin tan film.
All 64 petri dishes were transferred to a water table
after the first three days. Five juvenile abalone (8 rnrn)
were placed in each of 16 dishes containing a diatom
9
film. The dishes were covered with a plastic screen to
contain the grazing abalone. Filtered seawater was deliver
ed to the water table and all dishes remained immersed for
72 hours. A second batch of juvenile abalone was placed in
16 additional dishes containing the diatom film. These
animals were allowed to graze their designated dishes for 24
hours. The remaining 16 petri dishes were left with an
ungrazed diatom film.
After 72 hours on the water table the abalones were
removed, leaving four substrates available for use in the
settlement experiment: 16 petri dishes with a diatom film,
16 with a 24 hour mucous film, 16 with a 72 hour mucous
film, and 16 clean dishes. Eight petri dishes, represent
ing each substrate, were randomly positioned in each
settling trough. Approximately 5000 larvae were added to
each trough and a nytex screen was used to cover the
cultures.
The petri dishes were removed from each settling trough

after 72 hours. A Wild dissecting microscope was used to
count the number of larvae that settled in each dish.
Metamorphosis
10
Twelve small tanks were utilized to test the survival
of abalone larvae on various substrates. Each tank was
constructed by welding a PVC sheet to a section of 15.24 em
diameter PVC pipe. A capillary filtration standpipe was
fashioned for each tank from 1.27 em diameter PVC pipe and
90 micron mesh screen. The tank volumes were 1.2 liters
and provided 540.5 sq ern of wetted surface area. Seawater
filtered with 0.5 micron screen was delivered to each tank
at a rate of 100 rnl per min.
The twelve tanks were placed on a wet table 91.44 em
under two Chroma-50 flourescent tubes. Each tank was
innoculated with 1000 rnl of a diatom slurry. Within three
days the diatoms had settled and produced a tan film on
each tank surface. Ten juvenile abalone (8-15 rnrn) were
added to each of four tanks and allowed to graze the
substrate for three days. The abalones were removed from
the tanks after providing a fine mucous film on the sub
strate. Four additional tanks were inoculated with a

11
solution of GABA to a final concentration of 1 jlM. Three
treatments were therefore available: 1) diatoms, 2) diatoms
+ mucus, and 3) diatoms + GABA.
Two hundred larvae were added to each tank during the
three trials (5/28/85, 9/10/85, and 12/4/85) of the survival
experiment. The tanks were covered with a screen to prevent
contamination of the cultures. Sampling for mortalities
occurred after 3, 6, 9, and 11 weeks (12 weeks in trial 1).
The numbers of abalones exhibiting new shell growth were
counted at these times.
A Three-Way factorial mixed effect model ANOVA was
utilized to determine the effects of water treatment,
substrates, and time on the settling of larval abalone
(Winer, 1971). A Tukey's Studentized Range test was used to
compare the means. A Two-Way ANOVA was used to examine the
effect of period and substrate on larval survival (Zar,
1984). A nonparametric Tukey type test was used to deter
mine significant differences between the samples.

12
Results
Settlement
Larval settlement was significantly affected by
substrate (P< .001) and by period (P< .001) (3 Way ANOVA;
Table 1). However, the effect of water treatment on
larval settlement was not significant. All two way inter-
action effects (substrate x treatment, substrate x period,
and treatment x period) had a significant influence on
larval settlement (P< .001). The three way interactions
effect was not significant.
There were significant differences between larval
settlement on mucous substrates and on diatom and clean
plastic substrates (Tukey Test; Table 2a). Mean settling
differences were not significant between the 72 hour mucous
(X= 63.99; SD = 32.20) and 24 hour mucous (X= 57.99; SD =
31.55) substrates. Similiarly, no difference was noted
between the clean plastic (X= 44.23; SD = 31.74) and diatom
(X= 43.35; SD = 33.27) substrates.
Larval settlement varied with time (3 Way Anova;
Table 1). A significantly high number of larvae (X ~ 76.31;
SD = 25.36) settled during the 8/20/85 trial (Tukey . '

13
Table 2b and Figure 1) . Larval settlement was significantly
lower (X= 20.59; SD = 25.66) during the 5/28/85 trial
(Tukey Test; Table 2b and Figure 1).
Metamorphosis
Survival of larvae was markedly different (2 Way Anova;
P<O.Ol) among the three substrates (Table 3; Figure 2).
Approximately 50% of the larvae, in tanks with a mucous
treatment, survived through development of the first
respiratory pore, at 11 to 12 weeks, {Figure 2). The 0%
larval survival in the GABA treatments was significantly
lower (0.005<P<0.01) than the larval survival in the mucous
treatments (nonparametric Tukey Test q= 4.30). The survival
of larvae in tanks with a mucous substrate was not signifi
cantly different from survival on diatom substrates (
2.36). No signifcant difference was noted between survival
in the diatom and GABA treatments ( 1.94). All treatment
tanks, in each trial run, exhibited a high initial mortality
through week 3 of the experiment.

14
Discussion
Settlement
The results obtained suggest that red abalone larvae
will settle on several substrates including diatoms, mucus,
and clean plastic, but settlement on mucus was signifi
cantly higher. This agrees with the findings of Seki and
Kan-no (1981b) who found the larvae of Haliotis discus
hannai settle preferentially on the mucous trails of adult
abalones. Based on these findings it seems larval red
abalone utilize mucus, or some associated component of the
mucus, as an inducer of settlement and metamorphosis. Mucus
discrimination has been reported in adult gastropods
(Peters, 1964; Lowe and Turner, 1976) that used their
cephalic tentacles to "taste" the substrate. Seki and
Kan-no (1981a) have observed Haliotis discus hannai veligers
testing substrates with their cephalic tentacles prior to
settlement. However, the presence of ungrazed diatoms in
the mucous film may also provide some inductive cue to the
larvae.
The amount of mucus present did not significantly
affect the number of larvae that settled. This might be

15
due to a decrease in mucous production over time by the
grazers held for 72 hours. Culley and Sherman (1985)
reported mucous production was related to substrate texture.
Production was decreased when the pedal surface was protect
ed from abrasion. Another possibility is that the inductive
agent within the mucus might degrade or change over time
periods longer than 24 hours. For example, Seki and Kan-no
(1981b) noted different settling rates of abalone larvae on
mucus collected from grazing, crawling, and stimulated
adults. This suggested that the secreted mucus was of a
different chemical nature. These hypotheses could fee-
tively explain the similar inductive potential of 72 hour
and 24 hour mucus in my experiments.
Larval settlement on each substrate was unaffected by
the presence of GABA. Akashige et (1981) noted the
velar cilia of the larvae were paralyzed by GABA and
they suggested that it caused the larvae to settle un
naturally. I observed no difference in settlement of larvae
on mucous substrates in FSW and on mucous substrates in FSW
+ GABA. This seems to suggest that if larvae are narco
tized by GABA they are still capable of a certain degree
of substrate selectivity or GABA does not incapacitate the
larvae as reported. The fact that higher settlement was

16
noted on mucous substrates than diatom or clean substrates
suggests the larvae are actively testing the substrate for a
preferred inductive agent. Morse et al. (1979a) described
the induction of metamorphosis by GABA as a stereochemically
specific system. However, I observed larval selection of a
preferred substrate in the presence of GABA. Perhaps the
inductive cue in mucus utilizes a separate pathway to
trigger larval settlement behavior or GABA is not blocking
all the receptor sites. The lack of statistically signifi
cant differences of larval settlement on clean plastic in
the two water treatments is consistent with the findings of
Morse et al. (1979b) and indicates the importance of
antibiotics when using GABA as an inductive cue.
The most surprising result suggested that larval
settlement varied with time. This is confusing since all
trials were subjected to a controlled set of conditions.
Lannan (1980) observed increased larval survival when
fertilization occured during an optimal period in the
adult's gametogenic cycle and confirmed a genetic component
(in addition to environmental factors) in the role of larval
settlement success. Abalones exhibit extreme variability in
gametogenic cycles (Mottet, 1978). The April to July peak
spawning season in red abalones (Ault, 1982) suggests

17
genetic variation may be important in my experiments. It is
interesting to note that larvae from the third settlement
(6/30/85) group were collected during a natural spawning of
abalones held in raw seawater in the lab. These larvae did
not exhibit a high degree of settlement suggesting other
factors were also affecting the larval set.
Metamorphosis
Although the results suggest red abalone larvae can
settle quite successfully on many substrates, meta
morphosis and survival were highly variable on each. The
high rate of survival on the mucous substrate seems signifi
cant as this was the preferred inductive cue for larval
settlement. The mucus of gastropods has been well studied
(Calow, 1974; Crisp, 1967; Cook, 1971; Grenon and Walker;
1980; Hughes, 1978; Lowe and Turner, 1976; Peters, 1964) and
is probably quite important ecologically. Seki and Kan-no
(1981b) noted abalone larvae of Haliotis discus hannai
settled on mucus produced by adults of the same species, as
well as other abalone species, suggesting a common agent
within the mucus. It follows that the mucus is an important
inductive agent for larval abalone settlement. However,

there is little evidence to support a gregarious settling
strategy in natural populations of abalone larvae. It
appears this system may have been historically important,
perhaps as a precursor to the coraline algae inductive
system described by Morse et ~ (1980).
18
Survival noted on the diatom substrate is interesting
as it compares quite favorably with the survivorship
observed by Ebert and Houk (1984) in their hatchery tanks.
They reported yields ranging from 1.9 to 13.5 % dependent on
larval stocking densities. Similar survivorship was
reported in Japanese hatcheries utilizing mass culture
methods (Kan-no, 1975). Most researchers agree the initial
diet is most important in determining survival of post
settled larvae (Garland et al., 1984; Imai, 1967; Seki,
1980). This might help explain the initial high mortality
observed during the first three weeks of the experiments.
The evidence suggests the larvae are feeding in-
e iciently in the culture containers. Recent studies have
shown newly settled larvae were incapable of digesting large
pennate diatoms (>10 microns) due to the slow development
of the radula and fed almost exclusively on bacteria
grazed from the substrate (Garland et al., 1984). Th
suggests another food source, absent in my cultures, might

be crucial during the early (1-3 weeks) juvenile develop
ment.
19
The higher rate of larval survival in mucous cultures
compared to diatom cultures indicates the nutritional value
of mucous material (Calow, 1974). Larvae may be utilizing
mucoproteins, mucopolysaccharides, bacteria, or some as yet
undetermined component of the mucus during the initial 3
weeks of settlement. The poor survival of larvae settling
in the presence of GABA superficially supports the argument
of Akashige et al. (1981) that narcotized larvae are incap
able of feeding. However it seems more likely the absence
of antibiotics, a requisite component of the GABA inductive
system (Morse, pers. com.), had a greater effect on larval
survival,
The o % survival recorded in tanks containing GABA is
consistent with the results reported by Morse et al. (1979b,
1980) in which they compared survival in penicillin and
streptomycin treated cultures with untreated cultures. I
used no antibiotics in my cultures; however, my experimental
design allowed for equal infection of all cultures. It is
surprising that larvae in mucous and diatom cultures
persisted, with varying degrees of success, while larvae in
GABA cultures did not survive. Characteristic grazing of

20
the diatom~ilm (Ebert and Houk, 1984~ Morse et , 1979a;
Mottet, 1978) was evident in the mucous and diatom cultures
but not in the GABA culture. I observed stunting of larvae
and a lack of new shell growth in the GABA cultures.
Approximately 10 % of the larvae in the diatom and mucous
cultures also exhibited stunting; this probably represent
ed the "normal 11 situation or was an artifact of the hypo
thesized incomplete diet. These results confirm the
importance of antibiotics in culture work utilizing GABA.
My results indicate red abalone larvae settle prefer
entially on the mucous trails left by grazing juvenile and
adult abalone. In addition, the highest rates of meta
morphosis and survival were recorded on the mucous sub
strate. Many invertebrate species have larvae which settle
preferentially in the optimal adult habitats (Burke, 1983;
Crisp, 1974; Scheltema, 1974). Morse and his colleagues
(1980) proposed an evolutionary bond between abalone larvae
and certain species of crustose red algae which appeared to
be optimal "nursery habitats" for juvenile abalones. My
results suggest this coevolution was secondary in develop
ment to the mucus inductive system. Furthermore, my results
strongly suggest mucus is the most important inductive agent
for the settlement and metamorphosis of larval red abalone.

\
References
Akashige, s., Seki, T., Kan-no, H. and Nomura, T., 1981. Effects ofl'-aminobutyric acid and certain neurotransmitters on the settlement and the metamorphosis of the larvae of Ha1iotis discus hannai Ino (Gastropoda). Bull. Tohoku Reg. Fish. Res. Lab., 43:37-45.
Ault, J.S., 1982. Aspects of the laboratory reproduction of the red abalone, Haliotis rufescens Swainson. M.S. Thesis, Humboldt State University. 77pp.
Baloun, A.J. and Morse, D.E., 1984. Ionic control of settlement and metamorphosis in larval Haliotis rufescens (Gastropoda). Biol. Bull., 167:124-138.
21
Barker, M.F., 1977. Observations on the settlement of the brachiolaria larvae of Stichaster australis (Verrill) and Coscinasterias calamaria (Gray) (Echinodermata: asteroidea) in the laboratory and on the shore. J. Exp. Mar. Biol. Ecol., 30:95-108.
Barnes, J.R. and Goner, J.J., 1973. The larval settling response of the lined chiton Tonicella lineata. Marine Biol., 20:259-264.
Bonar, D.B., 1976. Molluscan metamorphosis: A study in tissue tranmsformation. Amer. Zool. 16:573-591.
Burke, R.D., 1983. The induction of marine invertebrate larvae: Stimulus and response. can. J. Zool., 61:1701-1719.
Calow, P., 1974. Some observations on locomotary strategies and their metabolic effects in two species of freshwater gastropods, Ancylus fluviatilis Mull. and Planorbis contortus Linn. Oecologia (Berl.), 16: 149-161.
Chia, F.S., 1978. Perspectives: Settlement and metamorphosis of marine invertebrate larvae. in: Settlement and metamorphosis of marine invertebrate larvae. (ed. Chia, F.S. & Rice, M.E.). Elsevier, New York. 290 pp.
Crisp, D.J., 1967. Chemical factors inducing settlement in Crassostrea virginica (Gmelin). J. Anim. Eco1., 36:329-335.

22
Crisp, D.J., 1974. Factors influencing settlement of marine invertebrate larvae. in Chemorecption in Marine Organisms (edited by Grant, P.T. and Mackie, A.M.), 177-265. Academic Press: New York.
Cook, S.B., 1971. A study of homing behavior in the limpet Siphonaria alternata. Biol. Bull., 141: 449-457.
Culley, M. and Sherman, K., 1985. The effect of substrate particle size on the production of mucous in Haliotis tuberculata L. and the importance of this in a culture system. Aquaculture 47: 327-334.
Ebert, E.E., Haseltine, A.W. and Kelly, R.O., 1974. Seawater system design and operations of the Marine Culture Laboratory, Granite canyon. Calif. Fish and Game, 60(1) :4-14.
Ebert, E.E. and Hamilton, R.M., 1983. Ova fertility relative to temperature and to the time of gamete mixing in the red abalone, Haliotis rufescens. Calif. Fish and Game, 69(2): 115-120.
Ebert, E.E. and Houk, J.L., 1984. Elements and innovations in the cultivation of red abalone Haliotis rufescens. Aquaculture, 39:375-392.
Garland, C.D., Cook, S.L., Grant, J.F. and McMeekin, T.A., 1984. Ingestion of bacteria and cuticle on non-articulated coralline algae by juvenile abalone (Haliotis ruber) . in press. 16pp.
Gee, J.M., 1965. Chemical stimulation of settlement in larvae of Spirorbis rupestris (Serpulidae) . Animal Behavior, 13:181-186.
Grant, J.F., 1981. Abalone culture in Japan: Development and current commercial practice. Tasmanian Fish. Res. 23:2-17.
Genon, J.F., and Walker, G., 1980. Biochemical and rheological properties of the pedal mucous of the limpet, Patella vulgata L. Comp. Biochem. Physiol., 66(B): 451-458.

23
Hadfield, M.G., 1978 . Metamorphosis in marine molluscan larvae: An analysis of stimulus and response. in Settlement and Metamorphosis of Marine Invertebrate Larvae (edited by Chia, F. and Rice, M.E.), 165-175. Elsevier: New York.
Hadfield, M.G., 1984. settlement requirements of molluscan larvae: New data on chemical and genetic roles. in Advances in Aquaculture and Fisheries Science: Recent Advances in Cultivation of Pacific Molluscs (edited by Morse, D.E., Chew, K.K. and Mann, R.), 283-298. Elsevier: New York.
Hughes, R.N., 1978. The biology of Dendropoma corallinaceum and Serpulorbis natalensis two south African vermetid gastropods. Zool. J. Linn. Soc., 64(2): 111-128.
Imai, T., 1967. Mass production of molluscs by means of rearing the larvae in tanks. Venus, 25(3&4): 159-167.
Ino, T., 1966. Abalone science and its propagation in Japan. Fish. Res. Bd. Can. Trans. Serv. No. 1078. 209pp.
Kan-no, H., 1975. Recent advances in abalone culture in Japan. Proceedings of the First International Conference on Aquaculture Nutrition, October,l975:195-211.
Kikuchi, s. and Uki, N., 1974. Technical study on the artificial spawning of abalone, genus Haliotis II: Effect of irradiated sea water with ultraviolet rays on inducing to spawn. Bull. Tohoku Reg. Fish. Res. Lab., 33:79-86.
Lannan, J.E., 1980. Broodstock management of crassostrea gigas. I. Genetic and environmental variation in survival in the larval rearing system. Aquaculture, 21: 323-336.
Leighton, D.L., 1972. Laboratory observations on the early growth of the abalone, Haliotis sorenseni, and the effect of temperature on larval development and settling success. Fish. Bull., 70(2) :373-381.
Leighton, D.L., Byhower, M.J., Kelly, J.c., Hooker, G.N. and Morse, D.E., 1981. Acceleration of development and growth in young green abalone (Haliotis fulgens) using warmed effluent seawater. J. World Maricult. Soc., 12(1) :170-180.

Lowe, E.F., and Turner R.D. 1976. Aggregation and trail following in juvenile Bursat~lla leachii pleii. Veliger, 19(2): 153-155.
24
McCallum, I.D., 1979. A simple method of taking a subsample of zooplankton. N.Z. J. Mar. & FW. Res., 13(4): 559-560.
Morita, H., 1972., Primary processes of insect chemoreception. Adv. Biophys. 3: 161-198.
Morse, A.N.c., Froyd, c.A. and Morse, D.E., 1984. Molecules from cyanobacteria and red algae that induce larval settlement and metamorphosis in the mollusc Haliotis rufescens. Mar. Biol., 81:293-298.
Morse, A.N. and Morse, D.E., 1984. Recruitment and metamorphosis of Haliotis larvae induced by molecules uniquely available at the surfaces of crustose red algae. J. Exp. Mar. Biol. Ecol., 75:191-215.
Morse, D.E., 1984. Biochemical and genetic engineering for improved production of abalones and other valuable molluscs. in Advances in Aquaculture and Fisheries Science: Recent Advances in the Cultivation of Pacific Molluscs (ed. by Morse, D.E., Chew, K.K., and Mann, R.) Elsevier: New York. 263-282.
Morse, D.E., Duncan, H., Hooker, N., and Morse, A., 1977. Hydrogen peroxide induces spawning in molluscs, with activation of prostaglandin endoperoxide synthetase. Science, 196: 298-300.
Morse, D.E., Duncan, H. Hooker, N., and Morse, A., 1979a. /f-aminobutyric acid, a neurotransmitter, induces planktonic abalone larvae to settle and begin metamorphosis. Science 204: 407-410.
Morse, D.E., Hooker, N., Jensen, L. and Duncan, H., 1979b. ~~duction of larval abalone settling and metamorphosis by {-aminobutyric acid and it's congeners from crustose red
algae: II: Applications to cultivation, seed-production and bioassays; principle causes of mortality and interference. Proc. World Maricul. Soc. 10:81-91.

25
Morse, D.E., Tegner, M., Duncan, H., Hooker, .N., Trevelyan, G. and cameron, A., 1980. Induct~on of settling and metamorphosis of planktonic molluscan (Haliotis) larvae. III: signaling by metabolites of intact algae is dependent on contact. in Chemical Signals (edited by Muller-Schwarze, D. and Silverstein, R.M.), 67-86. Plenum Press: New York.
Mottet, M.G., 1978. A review of the fishery biology of abalone. Wash. state Dept. of Fish. Tech. Rep. 37. 81 pp.
Peters, R.s., 1964. The function of cephalic tentacles in Littorina. Veliger, 7: 143-148.
Rumrill, s.s. and cameron, R.A., 1983. Effects of gamma-aminobutyric acid on the settlement of larvae of the black chiton Katharina tunicata. Mar. Biol., 72:243-247.
Scheltema, R.S., 1974. Biological interactions determining larval settlement of marine invertebrates. Thallasia Jugoslav., 10:263-296.
Seki, T., 1980. An advanced biological engineering system for abalone seed production. in Proc. Intl. Symp. on Coast. Pac. Mar. Life. 45-54.
Seki, T. and Kan-no, H., 1977. Synchronized control of early life in the abalone, Haliotis discus hannai Ino, Haliotidae, Gastropoda. Bull. Tohoku Reg. Fish. Res. Lab., 38:143-153.
Seki, T. and Kan-no, H., 198la. Observations on the settlement and metamorphosis of the veliger of the Japanese abalone, Haliotis discus hannai Ino, Haliotidae, Gastropoda. Bull. Tohoku Reg. Fish. Res. Lab., 42:31-39.
Seki, T. and Kan-no, H., 198lb. Induced settlement of the Japanese abalone, Haliotis discus hannai, veliger by the mucous of the juvenile and adult abalones. Bull. Tohoku Reg. Fish. Res. Lab., 43:29-36.
Steneck, R.S., 1982., A limpet-coralline alga association: adaptations and defenses between a selective herbivore and it 1 s prey. Ecology, 63: 507-522.

Winer, B. J. 1971. Statistical Principles In Experimental Design, 2nd. 'Edition. McGraw-Hill Inc., New York. 907 pp.
Zar, J. H., 1984. Biostatistical Analysis, 2nd. Edition. Prentice-Hall Inc., New Jersey. 718 pp.
26

27
Source of Variation DF ss MS F p
Total 639 733936.34
Cells 79 349270.84 4421.15 6.44 <0.001
Treatment 1 8555.63 8555.63 2.52 <0.5
Substrate 3 50270.53 16756.85 9.48 <0.001
T X S 3 16589.56 5529.85 8.35 <0.001
Period 9 177707.38 19745.26 28.75 <0.001
T X p 9 30551.09 3394.57 4.94 <0.001
s X p 27 47719.81 1767.40 2.57 <0.001
T X s X p 27 17876.84 662.11 0.96 <0.5
Error
Table 1.
560 384665.50 686.90
ANOVA summary Table showing the effects of water treatment (FSW and FSW + GABA), substrate (clean plastic, diatoms, 24 hour mucus, and 72 hour mucus), period, and interactions (between treatment, substrate, and period) on larval settlement.

a)
N Mean
160 63.99
160 57.94
160 44.23
160 43.35
b)
N Mean
64 76.31
64 72.19
64 64.67
64 61.56
64 54.67
64 53.39
64 48.48
64 38.11
64 33.92
64 20.59
Table 2.
28
SD Substrate Tukey Grouping
32.20 72 hour mucus
J 31.55 24 hour mucus
31.75 clean plastic
J 33.27 diatoms
SD Period/(date) Tukey Grouping
25.36 6 (08/20/85)
J 13.93 7 (09/10/85)
J 20.14 5 (08/05/85)
26.49 10 (12/04/85)
40.60 9 ( 10/06/85)
39.43 4 (07/15/85)
33.49 8 (09/23/85)
] 21.97 3 (07/01/85)
25.77 1 (05/28/85) ] 25.66 2 (06/17/85)
Tukey's studentized Range Groupings for mean larval settlement with respect to substrate (a), and period (b);~= 0.05, DF = 560, MS = 686.9027. Means with overlapping bars are not significantly different.

Source of Variation
Total
Cells
Substrate
Period
S X p
Error
Table 3.
29
DF ss MS F p
143 404809.49 2830.84
11 370001.41 33636.49 127.56 <0.001
2 283742.76 141871.38 538.01 <0.001
3 79849.24 26616.41 100.94 <0.001
6 6409.40 1068.23 4.05 <0.001
132 34808.08 263.70
ANOVA Summary Table showing the effects substrate (diatoms, diatoms + GABA, and diatoms+ mucus), period, and the interactions (between substrate and period) on larval survival.
160 160
140 a 140 b
120 120
100
eo
' 0~~~~~~~/~~~~~~~~~~
160
140 c
120
100
Figure 1.
2 3 4 s s 1 a s 10 2 3 4 5 a 1 a 9 10
160
140 d lili£1 FSW
120 ~ FSW+GABA
100
2: 3 4 5 s 1 a g 10 2 3 4 s 6 7 ll 9 10
Period Period
Mean Settlement of abalone larvae in FSW and FSW + GABA on 72 hour mucous (a), 24 hour mucous (b), clean plastic (c), and diatoms (d) substrates during ten periods from 5/28/85 to 12/4/85. Error bars = + 1 SD. Histograms =X larvae from 8 petri dishes.
w 0
200
180
160 CIJ
140 ro t: ro -1 120 CIJ :> :J 100 =1:1: c co 80 CIJ :E
60
40
20
0 0
Figure 2.
31
-o- Dlatoms+Mucus -+- Diatoms -1!8- DlatomS+GABA
3 6 9 12 Weeks
Mean Survival of abalone larvae on diatoms + mucous, diatoms, and diatoms + GABA substrates during three trials; 5/28/85, 9/10/85, and 12/4/85. Error bars = ± 1 SD. Symbols = X of 12 replicates for each substrate.

Substrate
olsan plaliltic
diatoms
24 hr mucus
7:il hr mucus
Appendix A.
32
Treatment
FSW FSW + GABA
Period N Miiii<!W so Mean so
1 a 13.75 4.87 32.38 12.60 2 8 11.00 11. OJ 22.38 9.49 3 a 24.13 18.57 41.38 9.77 4 8 32.38 14.84 49.00 17.36 5 8 84.88 45.63 74.63 73.68 6 8 61.38 28.36 57.38 58.20 7 e 61.13 19.89 66.25 23.18 8 a 49,50 12.85 44.75 25,61 9 a 33.00 10.58 47.13 13.62
10 a u.88 10.43 33.38 10.00
1 8 5.25 2.76 16.88 14.52 2 8 13.00 9.15 31.50 19.84 3 8 22.38 19.70 35.00 10.85 4 8 34.38 13.07 46.50 13.75 5 B 59.63 21.96 36,00 33.45 6 8 90.38 28.55 85.13 57.68 7 B 75.00 :n. 79 49.75 32.39 a 8 32.88 19.85 38.25 20.01 9 a 48.38 15.77 44.88 24.29
10 8 53.00 U.62 48.88 17.78
1 8 42.50 5.61 43.75 16.48 2 8 13.00 7.!56 21.63 20.56 3 B 46.50 13.14 44.88 14.05 4 8 52.25 23.96 60.88 19.58 5 8 97.75 29.03 49.00 31.43 6 8 83.63 25.29 53.63 40.57 7 B 90.75 23.06 76.50 43.45 8 8 48.50 18.72 55.50 23.18 9 8 74.13 20.50 49.63 27.93
10 B 53.00 19.62 48.88 17.78
1 8 73.13 34.26 44.13 19.23 2 8 21.50 13.94 30.75 13.72 3 8 4 6. B 8 31.52 43.75 26.67 4 8 eo. 38 27.45 71.38 38,64 5. 8 74.63 36.21 40.88 25.80 6 7 8 9
10
B 8·L50 29.52 94.50 36.15 8 116.75 51.73 61.38 23.72 8 68.50 24.10 50,00 16.63 8 91.38 22.35 48.88 27.75 8 99.63 16.517 56.88 14.47
Mean Settlement of abalone larvae in water treatments (FSW and FSW + GABA) over four substrates (clean plastic, diatoms, 24 hour mucus, and 72 hour mucus) during ten periods (5/28/85 to 12/4/85). N = number of dishes sampled.
Substrate
Mucus
Diatoms
GABA
Appendix B.
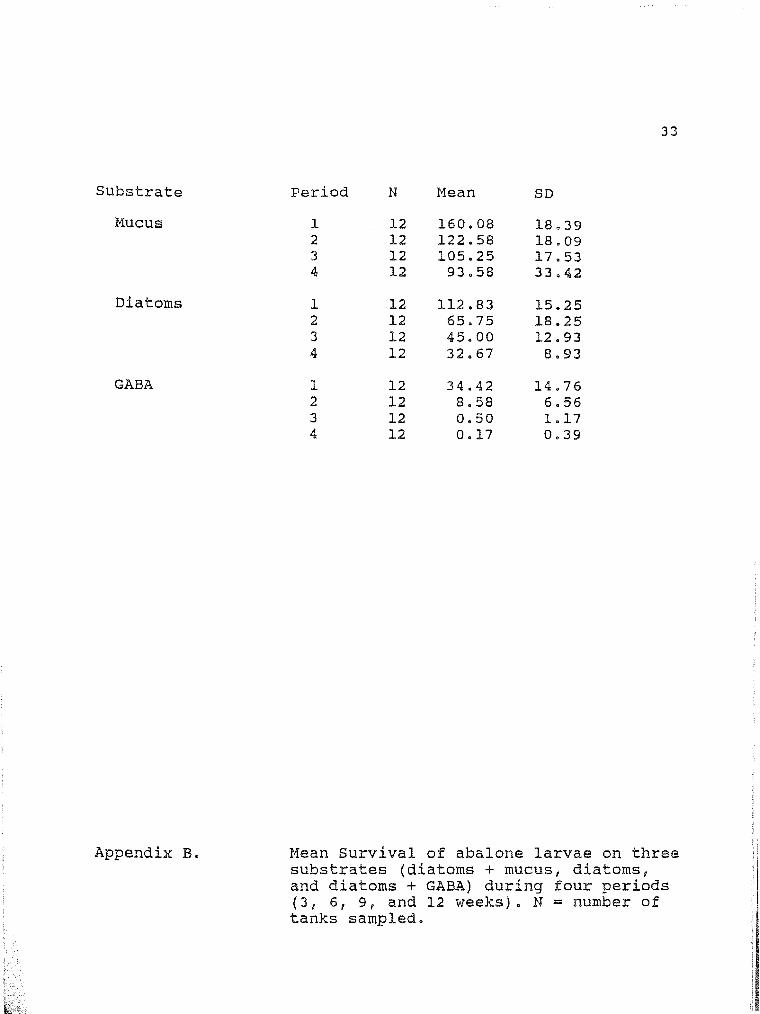
33
Period N Mean SD
1 12 160.08 18.39 2 12 122.58 18.09 3 12 105.25 17.53 4 12 93.58 33.42
1 12 112.83 15.25 2 12 65.75 18.25 3 12 45.00 12.93 4 12 32.67 8.93
l 12 34.42 14.76 2 12 8.58 6.56 3 12 0.50 1.17 4 12 0.17 0.39
Mean Survival of abalone larvae on three substrates (diatoms + mucus, diatoms, and diatoms + GABA) during four periods (3, 6, 9, and 12 weeks). N = number of tanks sampled.