serological analysis of the deoxyribonucleic acid...
TRANSCRIPT

JOURNAL OF VIROLOGY, Nov. 1972, p. 951-958Copyright © 1972 American Society for Microbiology
Vol. 10, No. 5Printed in U.S.A.
Serological Analysis of the Deoxyribonucleic AcidPolymerase of Avian Oncornaviruses
I. Preparation and Characterization of Monospecific Antiserum withPurified Deoxyribonucleic Acid Polymerase
K. F. WATSON, R. C. NOWINSKI, A. YANIV, AND S. SPIEGELMANIinstitute of Cancer Research, Columbia University, College of Physicians and Surgeons, New York, New York
10032, and McArdle Laboratory for Cancer Research, University of Wisconsin, Madison, Wisconsin 53706
Received for publication 11 July 1972
Monospecific antiserum was prepared against purified deoxyribonucleic acid(DNA) polymerase from avian myeloblastosis virus (AMV). Immunodiffusionassay with purified DNA polymerase revealed that the anti-DNA polymeraseserum formed one precipitation band, whereas no reaction with any of the sevenmajor structural proteins of AMV was observed. The antiserum also demon-strated enzyme-neutralizing antibody activity that was associated with the immuno-globulin G fraction. There was no difference in the neutralization of DNA poly-merase activity directed by ribonucleic acid (RNA), DNA, or RNA-DNA hybridtemplates.
The ribonucleic acid (RNA) tumor or oncorna-viruses (10) have been demonstrated to containRNA-instructed deoxyribonucleic acid (DNA)polymerase activity (1, 4-6, 12-14). Serologicalrelationships of avian C-type viral DNA polym-erases "reverse transcriptase" have been describedby Parks et al. (11) by using partially purifiedenzyme from the Schmidt-Ruppin strain of Roussarcoma virus (RSV) for immunization and pro-duction of antisera. These authors demonstratedthat the polymerase preparations of avian on-cornaviruses contained a common antigen thatappeared to be group-specific (gs) since it wasnot found in similar preparations from mam-malian oncornaviruses. Furthermore, antigensfound in the polymerase appeared to be sero-logically distinct from the known viral gs anti-gens. However, limitations exist in such a sero-logical analysis, as it is not possible with apolyvalent antiserum to distinguish adequatelyreactions directed against the polymerase or toother closely associated viral structural gs pro-teins.
Recently, Kacian et al. (7) reported the puri-fication of the RNA-instructed DNA polymeraseactivity from avian myeloblastosis virus (AMV).This purified enzyme preparation was shown tocontain two protein units in equimolar amounts,one of 110,000 and the other of 69,000 molecularweight. Throughout the purification procedure,the responses to homologous AMV 70S RNA,double-stranded DNA, and the synthetic homo-
polymer duplexes seemed to be mediated by asingle protein entity. The availability of purifiedenzyme preparations has enabled us to prepare amonospecific precipitating antiserum against theDNA polymerase of AMV.
In this report we describe the preparation of anantiserum against highly purified DNA polym-erase of AMV, with an analysis of its precipi-tating and neutralizing properties against thepurified enzyme.
MATERIALS AND METHODSAMV, BAI strain A, was obtained from J. W.
Beard, Duke University, Durham, N.C., and purifiedby a method previously described (2, 7). Unlabeleddeoxynucleoside triphosphates and dithiothreitol wereproducts of P-L Biochemicals. Polydeoxythymidylate-polyriboadenylate (poly dT. poly rA) was supplied byMiles Laboratories. Tritiated thymidine triphosphateand tritiated poly rA were obtained from Schwarz-Mann. Nonidet P40 detergent was a gift of ShellChemical Co. Bovine serum albumin (BSA) andrabbit antisera against rat immunoglobulin G (IgG)were purchased from Miles Laboratories. "Activated"myeloblast DNA, prepared by mild deoxyribonucle-ase treatment, was kindly supplied by A. Kiessling,Sloan-Kettering Institute for Cancer Research, NewYork, N.Y. AMV 70S RNA was prepared as de-scribed previously (7). Isolated AMV proteins werekindly donated by E. Fleissner, Sloan-Kettering In-stitute. The proteins were separated by gel filtrationfrom virus solubilized in 6 M guanidine hydrochlorideand reducing agents (3, 9).
Purification of DNA polymerase. The polymerase951
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

WATSON ET AL.
from AMV was purified as described by Kacian et al.(7); approximately 400 mg of purified, viral proteinfrom chick plasma was used for the preparation.After the carboxymethyl-Sephadex chromatographicstep, the enzyme activity was concentrated by ammo-nium sulfate precipitation (50% saturation) andfractionated further by glycerol gradient centrifuga-tion. Figure 1 shows a sodium dodecyl sulfate-poly-acrylamide gel of the enzyme after velocity sedimenta-tion in a glycerol gradient. As reported previously, theenzyme contains two protein subunits in equimolaramounts and responds to homologous AMV viralRNA, double-stranded DNA, and synthetic homo-polymer duplexes (7). The polymerase preparationshown in Fig. 1 was used for production of antiseraand for the neutralization studies.
Standard polymerase assay. Reaction mixtures (100,uliters) for homopolymer-templated reactions con-tained the following in micromoles: tris(hydroxy-methyl)aminomethane (Tris)-hydrochloride (pH 8.2at 25 C), 5.0; potassium chloride, 10.0; magnesiumchloride, 0.8; dithiothreitol, 0.04; deoxyadenosine
FIG. 1. Sodium dodecyl sulfate-polyacrylamide gelelectrophoresis of putrified DNA polymerase. A saniple(18 ,g) of purified AMV DNA polymerase used forthese immunological studcies was characterized as de-scribed previously (7).
5'-triphosphate, 0.02; radioactive thymidine 5'-tri-phosphate, 0.01; synthetic poly dT-poly rA template,1.2 X 103 pmoles of polymer phosphate per strand;and purified AMV-DNA polymerase, 0.01 ug.
After incubation at 37 C for a specified time, thereactions were terminated at 0 C with the addition ofcold 5% trichloroacetic acid. The acid-precipitableradioactivity was collected on nitrocellulose filtersand counted in 0.4%,, 2,5-bis-2-(5-tcrt-butvlbenzo-axozolyl) thiophene in toluene.
Assays using AMV 70S viral RNA and activatedmyeloblast DNA were prepared identically except that0.02 ,umole each of deoxycytidine 5'-triphosphate anddeoxyguanosine 5'-triphosphate were included. Tem-plates were added at concentrations of 1 to 2 Ag per100-p,liter assay volume.The specific activity of 3H-labeled thymidine tri-
phosphate was 300 to 400 counts per min per pmole.Preparation of antisera. Antiserum against purified
enzyme or viral proteins was prepared in (W Fu XBN)F, hybrid rats. The initial immunizing materialwas antigen (0.3 mg of protein) emulsified with anequal volume of complete Freund's adjuvant andinjected in multiple subcutaneous sites. IncompleteFreund's adjuvant was used for the subsequent inocu-lations (each containing 0.3 mg of protein); these weregiven at triweekly intervals, subcutaneously at multiplesites. Precipitating antisera against the AMV polym-erase were obtained after the fifth immunization; pre-cipitating antisera against the major viral structuralproteins were obtained after the third immunization.The normal rat serum used as a control in the neu-tralization assays was obtained from (W/Fu X BN)Fthybrid rats.
For enzyme neutralization assays, rat sera werefractionated by chromatography on Sephadex G-200in 0.1 M Tris-hydrochloride, pH 8.0, buffer. Figure 2shows a typical chromatographic separation; 1.5 to2.0 ml of serum was applied to a column (2.5 by 90cm), and 2-ml fractions were collected at a rate of 6 mlper hr. Rat gamma globulins were identified sero-logically by immunodiffusion with rabbit anti-ratIgG antiserum. The relevant fractions were then con-centrated by ammonium sulfate precipitation (50%osaturation) and dialyzed against 0.1 xi Tris-hydro-chloride, pH 8.0. The protein concentration of IgGfractions was measured by the method of Lowry et al.(8).
Neutralization assay. In a volume of 60 /Aliters, 0.01pg of purified AMV DNA polymerase was mixed withserum protein (50-150 4g) or purified IgG fraction(3-50 ,g); the reaction mixture also contained 0.01M Tris-hydrochloride (pH 8.2), 0.15 xi potassiumchloride, 20% (v/v) glycerol, and 50 ,ug of BSA. After15 min at 37 C, standard polymerase assay conditionswere established as described above. The acid-precipi-table radioactivity was taken as a measure of the DNApolymerase activity not neutralized by added serumor IgG fraction.
Double-diffusion (Ouchterlony) tests were per-formed on slides in 2% Noble agar (Immunoplate,pattern C; Hyland Laboratories). The slides wereleft at room temperature in a humidified chamber.Optimal precipitation occurred within 24 hr.
Nuclease assay. Nucleolytic activities in sera and
952 J. VIROL.
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

AVIAN ONCORNAVIRUS DNA POLYMERASES. I.
RT
. -
2.....
TZ i gs
G°o
0.5-
70 90 1O 130 150FRACTION NUMBER
FIG. 2. Gel filtration chromatography of rat anti-AMVDNA polymerase serum. A 1.5-ml amount of ratantiserum was applied to a Sephadex G-200 column(2.5 by 90 cm) and chromatographed at 6 ml/hr with0.1 M Tris-hydrochloride (pH 8) elution buffer. IgGwas identified serologically.
IgG fractions were measured by the solubilization ofacid-precipitable, radioactively labeled nucleic acids.3H-AMV RNA, 3H-Escherichia coli DNA, and 3H-poly rA duplex with oligo-dT were used as substrates.A 150-pg amount of serum protein or 50 pug of purifiedIgG was incubated for 60 min at 37 C with each la-beled nucleic acid under standard polymerase assayconditions (minus deoxynucleoside triphosphates andDNA polymerase). Samples (50 pliters) of the incu-bation mixture were removed at 0 and 60 min andtreated with 5% trichloroacetic acid, and the acid-precipitable radioactivity was determined.
RESULTS
Immunodiffusion studies with antiserum pre-
pared against AMV DNA polymerase. Rat anti-AMV polymerase serum reacted in immuno-diffusion with concentrated preparations (600 jigper ml) of purified viral DNA polymerase (Fig. 3).One major precipitation band was generally ob-served, although occasionally a second morerapidly diffusing minor component was alsodetected. As would be expected for such a largemolecule, diffusion of the polymerase into 2%agar was slow, and the precipitin band oftenformed an arc about the antigen well.The major structural proteins of AMV were
isolated by chromatography in 6 M guanidinehydrochloride (Fig. 4) and tested by immuno-diffusion with rat anti-AMV DNA polymeraseserum. In all tests, these proteins did not reactwith the polymerase antiserum; these same pro-
FIG. 3. Immunodiffusiont tests wit/i rat anti-AMVDNA polymerase antisernim. Central well a, rat anti-AMV DNA polymerase antiserum; central wvell b,rabbit anti-AMV serum. The peripheral wells of bothpatterns are derived from the same antigen pools. Ab-breviations: AMV, ether-treated; RT, viral reverse-transcriptase enzyme; gsl and gs2 refer to group-
specific antigens of AMV isolated by gel filtration inguanidine hiydrochloride.
gs 1
300 4
U)200
~~~~~~~~gs3
0 ~~~~~~~gs2
100-m2
~~~~gs4p5
30 40 50 60 70 80 90 100 110
FRACTION
FIG. 4. Clhromatographic separation of AMV pro-teinis by gel filtrationi in 6 M guantiditie hydrochlorideand reducing agents. A small sample of MC29 virus,radiolabeled with 'I4C-amino acid mixture, was intcludedwith the AMV in order to identify the proteint peaks.Seven distinct viral proteins are observed. Two of thesepolypeptides (ml and m2) were glycoproteins andidentified immunologically as viral membrane antiigens.Four of the viral proteins (gsl, gs2, gs3, and gs4) are
serologically distinct grouip-specific anitigens, localizedin the interior of the virus. The seventh and smallestprotein, pS, has yet to be serologically definedl. Fordetails see references 3 and 9.
teins, however, were precipitated by their homolo-gous antisera (unpublished data). Figure 3 illus-trates tests with two of these isolated proteins.
Polyvalent rabbit antisera prepared againstether-degraded AMV and hamster sera fromanimals bearing transplants of RSV-inducedsarcomas (CoFAL reference antisera) precipi-tated the major structural viral antigens, but didnot react with preparations of purified enzyme.
Neutralization of DNA polymerase activity by
AMV .
953VOL. 10, 1972
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

WATSON ET AL.
antisera. To test for enzyme neutralization, in-creasing levels of antiserum and control rat serumwere incubated with purified DNA polymerase.Enzyme inhibition was determined by the loss ofpolymerase activity. A 50-,g amount of antiserumprotein inhibited the enzymatic activity by morethan 90% (Fig. 5A). However, as a function ofincreased serum protein, normal rat serum alsoexhibited an inhibitory effect, although less thanthat observed with the specific antiserum. Theseresults suggest the presence of both a specificantibody against the enzyme and a nonimmuno-logical inhibitor of DNA polymerase activity.To eliminate this inhibitor, all sera were chro-
matographed on Sephadex G-200, and the IgGfraction was isolated. Figure 5B shows enzymenCutralization assays with isolated IgG of normalrat serum tested against the AMV DNA polym-erase. After this purification, no inhibition of thepolymerase activity was observed in the presenceof up to 50 ,ug of normal rat IgG.To examine the nature of the nonspecific in-
hibitory factor, normal rat serum, antipolym-erase serum, and their corresponding IgG frac-tions were tested for their effect on componentsof the DNA polymerase assay. Since nucleolyticactivity could account for the observed inhibition,assays detecting degradation of nucleic acid tem-plates and DNA product were performed. Incu-bation of normal rat serum or antipolymeraseserum with radioactively labeled AMV 70S RNAresulted in a 40% loss of acid-precipitable radio-
B
-75
50 100 ;5C'
US serum ,role n
-25
+50 _
-75
rl-
1 2C0 040 50 'mlcroqrars IgO
FiG. 5. Enzyine neultralization assays With whole.serumn. A, Standard neultralization assay mixtutres were
prepared containing 0.01 Mg ofpurifiedDNA polymeraseantd increasing amounts of rat anti-AMV DNA poly-mnerase serum (closed circles) or normwal rat serum (opencircles). Preincubation of the serunm and enzyme was
for 15 min at 37 C, and the polymerase activity assaywvas performed at 37 C tbr 60 min. B, Smandard neu-
traclization assays were prepared as described abovewith 0.01 Mg ofpurifiedDNA polymerase and increasingamouints of normal rut seriion1 IgG fraction.
activity; however, nuclease tests of serum withlabeled E. coli DNA or with the hybrid template-labeled poly rA annealed with oligo-dT did notresult in a loss of acid-precipitable template. Thus,whole rat serum had ribonuclease activity, butno appreciable nuclease activity for DNA orhybrid nucleic acids. Similar tests with isolatedIgG from normal rat serum and rat antisera wereconsistently negative for ribonuclease activity,indicating the effectiveness of the isolation pro-cedure.
Conditions for neutralization of DNA polym-erase activity. Addition of purified rat anti-AMVDNA polymerase IgG fraction to a standardpolymerase assay (at zero time) resulted in nosignificant neutralization of DNA polymeraseactivity (Fig. 6). This observation suggested thata preincubation of antibody with antigen may
20
1 5
0~I
CL
0 10 20 30 40 50
micrograms IgG
FIG. 6. Effect of anti-AMV DNA polymerase IgGand normal rat IgG on DNA polymerase activity with-out preincubation. Standard polymerase assay mixtureswere prepared containing 0.01 uAg ofpurifiedDNA poly-merase and increasing concentrations of norm?al ratserum IgG (open circles) or anti-AMV DNA polymer-ase IgG (closed circles). The reaction mixtures were in-cubated 40 minz at 37 C anud terminated as described inMaterials and Methods. Bovine seruim albumin and 20%(v/v) glycerol were not present in the inclubalion m?lix-tures.
A
954 J. VIROL.
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
AVIAN ONCORNAVIRUS DNA POLYMERASES. T.
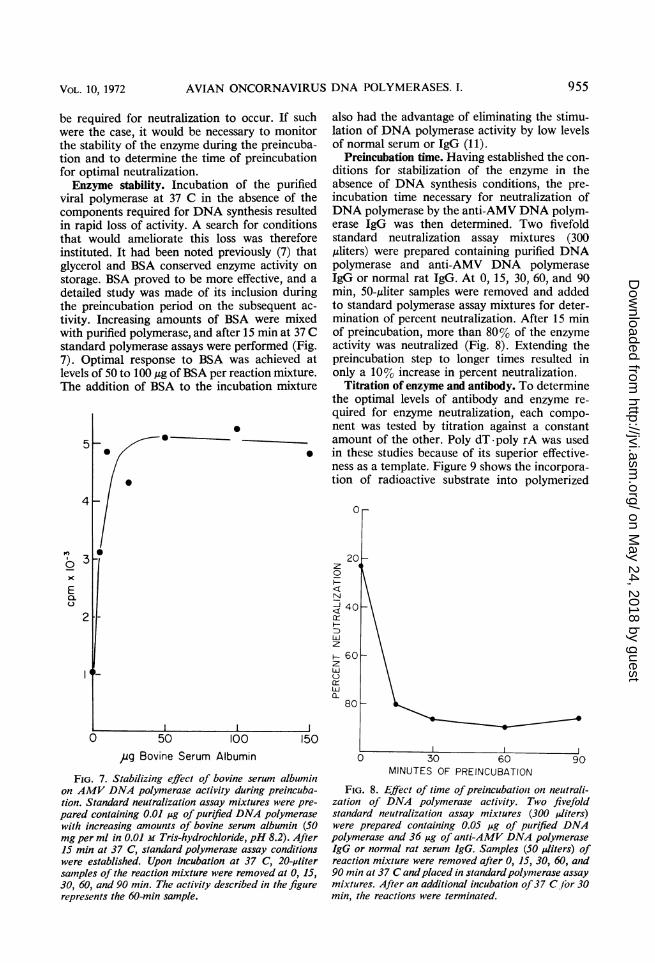
be required for neutralization to occur. If suchwere the case, it would be necessary to monitorthe stability of the enzyme during the preincuba-tion and to determine the time of preincubationfor optimal neutralization.Enzyme stability. Incubation of the purified
viral polymerase at 37 C in the absence of thecomponents required for DNA synthesis resultedin rapid loss of activity. A search for conditionsthat would ameliorate this loss was thereforeinstituted. It had been noted previously (7) thatglycerol and BSA conserved enzyme activity onstorage. BSA proved to be more effective, and adetailed study was made of its inclusion duringthe preincubation period on the subsequent ac-tivity. Increasing amounts of BSA were mixedwith purified polymerase, and after 15 min at 37 Cstandard polymerase assays were performed (Fig.7). Optimal response to BSA was achieved atlevels of 50 to 100 ,ug ofBSA per reaction mixture.The addition of BSA to the incubation mixture
aS -
,^o3
E
2
z0
N'I-J
IDLLJz
zLLJC-)
LUJa-
,ug Bovine Serum Albumin
FIG. 7. Stabilizing effect of bovine serum albuminon AMV DNA polymerase activity during preincuba-tion. Standard neutralization assay mixtures were pre-pared containing 0.01 ,ug of purified DNA polymerasewith increasing amounts of bovine serum albumin (50mg per ml in 0.01 M Tris-hydrochloride, pH 8.2). After15 min at 37 C, standard polymerase assay conditionswere established. Upon incubation at 37 C, 20-plitersamples of the reaction mixture were removed at 0, 15,30, 60, and 90 min. The activity described in the figurerepresents the 60-min sample.
also had the advantage of eliminating the stimu-lation of DNA polymerase activity by low levelsof normal serum or IgG (11).
Preincubation time. Having established the con-ditions for stabilization of the enzyme in theabsence of DNA synthesis conditions, the pre-incubation time necessary for neutralization ofDNA polymerase by the anti-AMV DNA polym-erase IgG was then determined. Two fivefoldstandard neutralization assay mixtures (300,uliters) were prepared containing purified DNApolymerase and anti-AMV DNA polymeraseIgG or normal rat IgG. At 0, 15, 30, 60, and 90min, 50-,uliter samples were removed and addedto standard polymerase assay mixtures for deter-mination of percent neutralization. After 15 minof preincubation, more than 80% of the enzymeactivity was neutralized (Fig. 8). Extending thepreincubation step to longer times resulted inonly a 10% increase in percent neutralization.
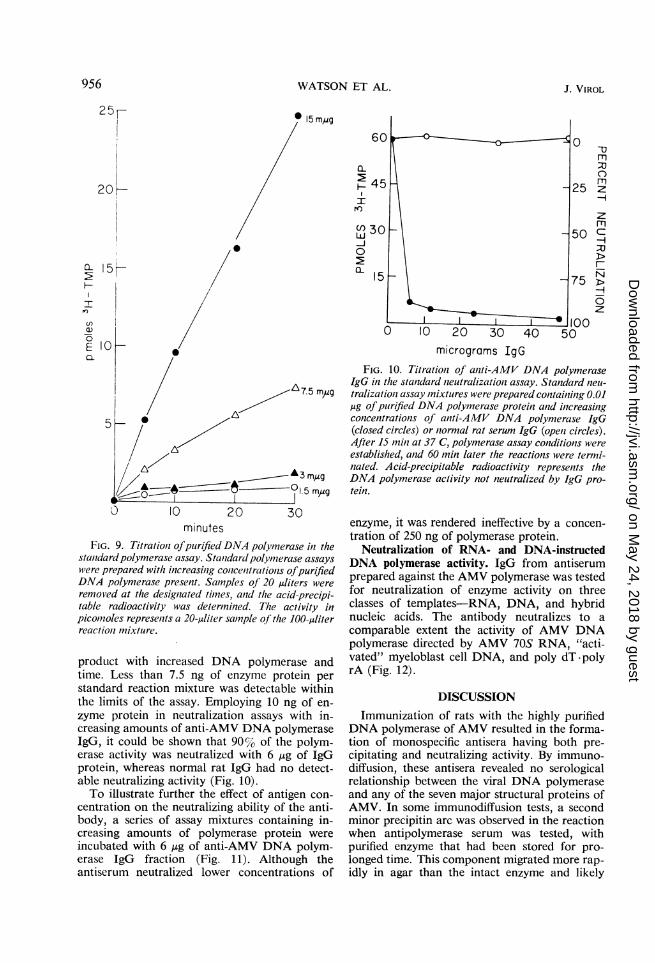
Titration of enzyme and antibody. To determinethe optimal levels of antibody and enzyme re-quired for enzyme neutralization, each compo-nent was tested by titration against a constantamount of the other. Poly dT -poly rA was usedin these studies because of its superior effective-ness as a template. Figure 9 shows the incorpora-tion of radioactive substrate into polymerized
0
90MINUTES OF PREINCUBATION
FIG. 8. Effect of time ofpreincubation on neutrali-zation of DNA polymerase activity. Two fivefoldstandard neutralization assay mixtures (300 ,diters)were prepared containing 0.05 ,ug of purified DNApolymerase and 36 pAg of anti-AMV DNA polymeraseigG or normal rat serum IgG. Samples (50 Aliters) ofreaction mixture were removed after 0, 15, 30, 60, and90 min at 37 C andplaced in standardpolymerase assaymixtures. After an additional incubation of37 C/tor 30min, the reactions were terminated.
955VOL. 10, 1972
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
WATSON ET AL.
* 15mug
20+
a-
a)0
0-
I
-J00~
/0
1O
Za7.5 m#ug
5+
5) 10 20 30minutes
FIG. 9. Titrationz ofpurified DNA polyimierase in thestanidardpolymerase assay. Stantdarc polymerase assayswere prepared with increasinzg conlcenttrationis ofpurifiedDNA polymerase present. Sanmples of 20 ,uliters were
removed at the designiated tines, anid the acid-precipi-table radioactivity was determined. The activity inpicomoles representts a 20-,uliter sample of the 100-jiliterreactiont mixture.
product with increased DNA polymerase andtime. Less than 7.5 ng of enzyme protein perstandard reaction mixture was detectable withinthe limits of the assay. Employing 10 ng of en-zyme protein in neutralization assays with in-creasing amounts of anti-AMV DNA polymeraseIgG, it could be shown that 90% of the polym-erase activity was neutralized with 6 ,ug of IgGprotein, whereas normal rat IgG had no detect-able neutralizing activity (Fig. 10).To illustrate further the effect of antigen con-
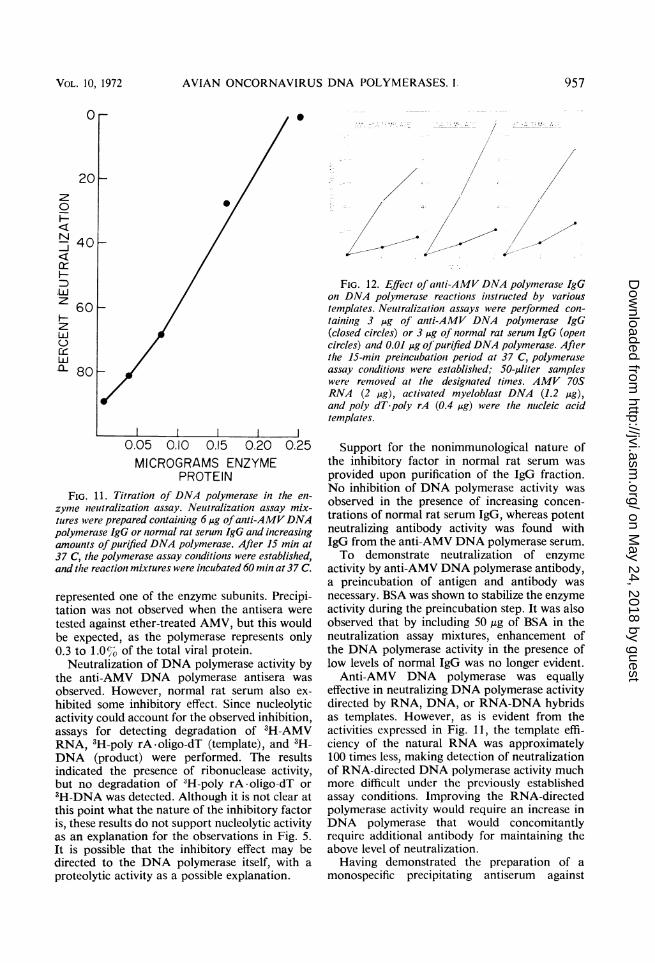
centration on the neutralizing ability of the anti-body, a series of assay mixtures containing in-creasing amounts of polymerase protein wereincubated with 6 ,ug of anti-AMV DNA polym-erase IgG fraction (Fig. 11). Although theantiserum neutralized lower concentrations of
-tJmC)mz-Hi
zrric
r-
z
micrograms IgG
FIG. 10. Titrationl of aniti-AMV DNA polymeraseIgG in the sianudard nieutralizationz assay. Standard nieu-tralizationi assay mixtures were prepared containinlg 0.01.ug of purified DNA polymerase proteit an2d inicreasingconcentrations of aniti-AMV DNA polymerase IgG(closed circles) or niormal rat serum IgG (opeil circles).After 15 niiii at 37 C, polymerase assay conditions wereestablished, anid 60 min later the reactions were termi-nated. Acid-precipitable radioactivity represenzts theDNA polymerase activity not neutralized by IgG pro-tein.
enzyme, it was rendered ineffective by a concen-tration of 250 ng of polymerase protein.
Neutralization of RNA- and DNA-instructedDNA polymerase activity. IgG from antiserumprepared against the AMV polymerase was testedfor neutralization of enzyme activity on threeclasses of templates-RNA, DNA, and hybridnucleic acids. The antibody neutralizes to acomparable extent the activity of AMV DNApolymerase directed by AMV 70S RNA, "acti-vated" myeloblast cell DNA, and poly dT -polyrA (Fig. 12).
DISCUSSIONImmunization of rats with the highly purified
DNA polymerase of AMV resulted in the forma-tion of monospecific antisera having both pre-cipitating and neutralizing activity. By immuno-diffusion, these antisera revealed no serologicalrelationship between the viral DNA polymeraseand any of the seven major structural proteins ofAMV. In some immunodiffusion tests, a secondminor precipitin arc was observed in the reactionwhen antipolymerase serum was tested, withpurified enzyme that had been stored for pro-longed time. This component migrated more rap-idly in agar than the intact enzyme and likely
956 J. VIROL-
25rF
1 5.
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
AVIAN ONCORNAVIRUS DNA PlOLYMERASES. 1.
Or-
20Fz
0
N
:D
z
fLz
CRLLQ
0
40F
60K
80K
I
0.05 0.10 0.15 0.20 0.25MICROGRAMS ENZYME
PROTEINFIG. 11. Titration of DNA polymerase in the en-
zyme neutralization assay. Neutralizationi assay mix-
tures were prepared containzinig 6 jig ofantti-AMVDNApolynmerase IgG or normal rat serum IgG an2d increasinlgamounts ofpurified DNA polymerase. After 15 min at37 C, the polymerase assay coniditions were established,and the reaction mixtures were incubated 60 min at 37 C.
represented one of the enzyme subunits. Precipi-tation was not observed when the antisera were
tested against ether-treated AMV, but this wouldbe expected, as the polymerase represents only0.3 to 1.0% of the total viral protein.
Neutralization of DNA polymerase activity bythe anti-AMV DNA polymerase antisera was
observed. However, normal rat serum also ex-
hibited some inhibitory effect. Since nucleolyticactivity could account for the observed inhibition,assays for detecting degradation of 3H-AMVRNA, 3H-poly rA .oligo-dT (template), and 3H-DNA (product) were performed. The resultsindicated the presence of ribonuclease activity,but no degradation of 3H-poly rA oligo-dT or
3H-DNA was detected. Although it is not clear atthis point what the nature of the inhibitory factoris, these results do not support nucleolytic activityas an explanation for the observations in Fig. 5.It is possible that the inhibitory effect may bedirected to the DNA polymerase itself, with a
proteolytic activity as a possible explanation.
..FIG. 12. Effect of anti-AMV DNA polymerase IgG
on DNA polymerase reactionis inistructed by varioustemplates. Neuftralization assays were performed con-taininig 3 uig of aniti-AMV DNA polymerase IgG(closed circles) or 3 jig of normal rat serutm IgG (opelzcircles) and 0.01 jg ofpurified DNA polymerase. Afterthe 15-mimz preincubationi period at 37 C, polymeraseassay contditionts were established; 50-lAliter sampleswere removed at the designated times. AMV 70SRNA (2 ig), activated myeloblast DNA (1.2 jig),anzd poly dT-poly rA (0.4 jAg) were the nueleic acidtemplates.
Support for the nonimmunological nature ofthe inhibitory factor in normal rat serum wasprovided upon purification of the IgG fraction.No inhibition of DNA polymerase activity wasobserved in the presence of increasing concen-trations of normal rat serum IgG, whereas potentneutralizing antibody activity was found withIgG from the anti-AMV DNA polymerase serum.To demonstrate neutralization of enzyme
activity by anti-AMV DNA polymerase antibody,a preincubation of antigen and antibody wasnecessary. BSA was shown to stabilize the enzymeactivity during the preincubation step. It was alsoobserved that by including 50,g of BSA in theneutralization assay mixtures, enhancement ofthe DNA polymerase activity in the presence oflow levels of normal IgG was no longer evident.Anti-AMV DNA polymerase was equally
effective in neutralizing DNA polymerase activitydirected by RNA, DNA, or RNA-DNA hybridsas templates. However, as is evident from theactivities expressed in Fig. 11, the template effi-ciency of the natural RNA was approximately100 times less, making detection of neutralizationof RNA-directed DNA polymerase activity muchmore difficult under the previously establishedassay conditions. Improving the RNA-directedpolymerase activity would require an increase inDNA polymerase that would concomitantlyrequire additional antibody for maintaining theabove level of neutralization.Having demonstrated the preparation of a
monospecific precipitating antiserum against
957VOL. 10, 1972
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

WATSON ET AL.
AMV DNA polymerase and established neutrali-zation assay conditions, it is now possible toextend these studies to include a serological com-parison of the AMV DNA polymerase to DNApolymerases from other avian and mammaliantumor viruses.
ACKNOWLEDG MENTS
This work was supported by PuLblic Health Ser-vice SpecialVirus Cancer Program Contract 70-2049, traininag grant CA-0501 1,research grant CA-02332, and Postdoctoral Fcllowship CA-38924, all fromi the National Cancer InstituLte.
LITERATURE CITEI)
]. Baltimore, D. 1970. Viral RNA-depenident DNA polymiierase.Nature (London) 226:1209-121 1.
2. Carnegie, J. W., A. O'C. Deeney, K. C. Olson, ziild G. S.Beaudreu. 1969. An RNA fraction tifromi imyeloblastosisvirus having properties similar to tranisfer RNA. Biochim.Biophys. Acta 190:274-284.
3. Fleissner, E. 1971. Chromatogratphic separaitioni and antigeniCanalysis of proteins of the oncornaviruses. I. Avian leuLke-mia-sarcoma viruses. J. Virol. 8:778-785.
4. Garapin, A., J. P. McDonnell, W. Levinisoni, N. Qulintrell, L.Fanshier, and J. M. Bishop. 1970. Deoxyribonucleic acidpolymerase associated with Rous sarcomiia virLus anid avianmyeloblastosis virus: properties of the enizymiie anid its prod-uct. J. Virol. 6:589-598.
5. Green, M., M. Rokutanda, K. FuLjinaga, R. K. Ray, H.Rokutanda, and G. Gurgo. 1970). Mechainismii of carcino-
genesis by RNA tumor viruses. 1. An RNA-dependentDNA polymerase in murine sarcoima viruses. Proe. Nat.Acad. Sci. U. S.A. 67:385-393.
6. Hatanaka, M., R. J. Huebner, and R. V. Gildeni. 1970. DNApolymerase activity associated with RNA tuLmor viruses.Proc. Nait. Acad. Sci. U.S.A. 67:143-147.
7. Kacian, D. L., K. F. Watson, A. Burny, and S. Spiegelman.1971. Purification of the DNA polymerase of avian miiyelo-blaistosis virus. Biochim. Biphys. Acta 246:365-383.
8. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Rand-all. 1951. Protein measurement with the Folin phenol re-
aigent. J. Biol. Chem. 193:265-275.9. Nowinski, R. C., E. Fleissner, N. H. Sarkar, anid T. Aoki.
1972. Chromatographic separation and antigenic anialysisof proteins of the oncornaviruses. II. Mammalian leuLkemia-sarcoma viruses. J. Virol. 9:359-366.
10. Nowinski, R. C., L. J. Old, N. H. Sarkar, and D. H. Moore.1970. Common properties of the oncogenic RNA viruses(oncornaviruses). Virology 42:1152-1157.
11. Parks, W. P., E. M. Scolnick, J. Ross, G. J. Todaro, and S.A. Aaronson. 1972. Immiiunological relationships of reverse
tralnscriptases from ribonucleic acid tumor viruses. J. Virol.9: 110-115.
12. Scolinick, E. M., S. A. Aaronson, and G. J. Todiaro. 1970.DNA synthesis by RNA-containing tumiior viruses. Proc.Nat. Acad. Sci. U.S.A. 67:1034-1041.
13. Spiegelmiian, S., A. Burny, M. R. Das, J. Keydar, J. Schlom,M. Travnicek, aInd K. Waitson. 1970. Characterization ofthe products of RNA-directed DNA polymerases in onco-
genic RNA viruses. Nature (London) 227:563-567.14. Temin, H. M., and S. Mizutani. 1970. RNA-dependent DNA
polymerase in virions of Rous sarcoma virus. Nature(London) 226:1211-1213.
958 J. VIROL.
on May 24, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from