seleção de progênies interpopulacionais ... - iac.sp.gov.br · campinas, sp fevereiro de 2015...
TRANSCRIPT

Seleção de progênies interpopulacionais de milho e
estimativas de parâmetros genéticos relacionados com
tolerância à seca
Cristiani Santos Bernini
CAMPINAS, SP
FEVEREIRO DE 2015

INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
Seleção de progênies interpopulacionais de milho e
estimativas de parâmetros genéticos relacionados com
tolerância à seca
Cristiani Santos Bernini
Orientadora: Maria Elisa Ayres Guidetti Zagatto Paterniani
Campinas, SP
Fevereiro de 2015
Tese submetida como requisito parcial
para obtenção do título do grau de
Doutora em Agricultura Tropical e
Subtropical, Área de Concentração em
Genética, Melhoramento Vegetal e
Biotecnologia.



Aos meus pais José e Maria
Ao meu filho João
Aos meus irmãos Fabiano e Marcos
Dedico
À minha orientadora Maria Elisa
Ofereço

AGRADECIMENTOS
A Deus.
Ao Instituto Agronômico (IAC).
À CAPES pela concessão da bolsa de estudos.
À avó Jovita, pelo carinho e, essencialmente, pelas orações.
Aos companheiros e irmãos, Fabiano e Janayna, pelo imenso apoio.
À professora, amiga e orientadora, Maria Elisa, pelos ensinamentos transmitidos, dedicação,
conselhos, competência e alegria que nos ensina a lidar com a pesquisa.
Aos docentes da pós-graduação e pesquisadores do Instituto Agronômico (IAC) pelos
ensinamentos transmitidos.
Ao Centro de Grãos e Fibras do IAC por disponibilizar os equipamentos.
Ao Centro de Ecofisiologia e Biofísica, especialmente a Dra. Regina Célia de Matos Pires e o
técnico Leonardo Teixeira.
À secretária Célia Terra pela atenção, apoio e amizade.
Aos professores Gonçalo Amarante Guimarães Pereira e Anete Pereira de Souza pelos
ensinamentos transmitidos e dedicação no estádio docência.
À pesquisadora Luciana Carlini pela ajuda na análise de dados dos experimentos e apoio
fundamental neste trabalho.
Aos funcionários da Fazenda Santa Elisa Carlinhos, Noberto, Fátima, Iolanda, Osvaldo,
Adelino, Claret, Eduardo (Baiano), Rafael, Arnaldo e José Carlos pelo auxílio na condução
dos experimentos, dedicação e amizade.
À Agência Paulista de Tecnologia dos Agronegócios – APTA, da cidade de Mococa – SP,
pela instalação e condução dos experimentos, especialmente ao Pesquisador Paulo Gallo e aos
funcionários Ronaldo e Geraldo.
Às amigas Paula, Sara, Renata e Andrea pelo apoio em todos os momentos de alegria e
tristeza e sobretudo pela amizade.
Aos membros da banca Dr. Eduardo Sawazaki, Dr. Antônio Paulino da Costa Netto, Dr.
Deoclecio Domingos Garbuglio e Dr. Walter José Siqueira pelas valiosas sugestões que muito
contribuíram neste trabalho.
Ao professor Antônio Paulino, meus sinceros agradecimentos, principalmente pelos
ensinamentos, orientação, dedicação e principalmente pela amizade. Aos pesquisadores
Aildson Pereira Duarte, Eduardo Sawazaki e orientadora Maria Elisa Paterniani pelos
conhecimentos transmitidos e pela oportunidade de crescer profissionalmente.

i
SUMÁRIO
LISTA DE TABELAS................................................................................... ii
LISTA DE FIGURAS.................................................................................... vi
LISTA DE ANEXOS..................................................................................... vii
LISTA DE ABREVIATURAS...................................................................... vii
RESUMO....................................................................................................... 1
ABSTRACT................................................................................................... 3
1 INTRODUÇÃO.......................................................................................... 5
2 REVISÃO DE LITERATURA................................................................... 7
2.1 Efeito do déficit hídrico em milho........................................................... 7
2.2 Parâmetros fisiológicos em resposta ao déficit hídrico........................... 13
2.3 Caracteres secundários............................................................................. 18
2.4 Melhoramento genético para tolerância à seca........................................ 22
2.4.1 Seleção recorrente................................................................................. 23
2.4.2 O fator latente....................................................................................... 27
2.5 Estimativas de parâmetros genéticos....................................................... 29
3 MATERIAL E MÉTODOS........................................................................ 31
3.1 Descrição do material.............................................................................. 31
3.2 Métodos................................................................................................... 32
3.2.1 Seleção recorrente de progênies interpopulacionais em campo........... 32
3.2.1.1 Obtenção das progênies interpopulacionais (ciclo 0)........................ 32
3.2.1.2 Avaliação de progênies interpopulacionais em campo...................... 33
3.2.1.3 Avaliação dos caracteres biométricos e secundários em campo........ 34
3.2.2 Análises biométricas............................................................................. 36
3.2.2.1 Análises de variâncias e estimativas de parâmetros genéticos.......... 36
3.2.2.2 Estimativa de heterose em relação à média dos pais.................... 39
3.2.3 Avaliação em casa de vegetação das progênies interpopulacionais
selecionadas..............................................................................................
40
3.2.3.1 Avaliação fisiológica em casa de vegetação...................................... 42
3.2.3.2 Avaliação biométrica em casa de vegetação...................................... 43
3.2.4 Determinação do índice de valor relativo de tolerância (RVT)............ 44
3.2.5 Análise de variância do experimento de casa de vegetação................. 44
4 RESULTADOS E DISCUSSÃO................................................................ 46
4.1 Experimentos de avaliação das progênies interpopulacionais em
campo...................................................................................................
46
4.1.1 Análises de variâncias, médias e heterose............................................. 46
4.1.2 Estimativas de parâmetros genéticos interpopulacionais...................... 64
4.2 Experimento de avaliação das progênies interpopulacionais em casa de
vegetação...................................................................................................
70
4.2.1 Análises de variâncias e médias........................................................... 70
5.0 CONCLUSÕES....................................................................................... 86
6.0 REFERÊNCIAS BIBLIOGRÁFICAS..................................................... 88
ANEXOS....................................................................................................... 103

ii
LISTA DE TABELAS
Tabela 1 - Descrição dos estádios de desenvolvimento do milho....................... 9
Tabela 2 - Esquema da análise de variância individual e esperanças dos
quadrados médios da análise do látice quadrado parcialmente
balanceado, utilizando as médias ajustadas de tratamentos e a
variância efetiva do látice....................................................................
36
Tabela 3 -
Período de déficit hídrico, temperaturas média de máxima e mínima
e umidade relativa dentro da casa de vegetação, potencial matricial
do solo e intervalo de variação (IV) durante o período de seca no
estádio fenológico de florescimento (VT)...........................................
42
Tabela 4 -
Resumo da análise de variância no delineamento em látice dos
caracteres altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (AC+Q), florescimento masculino (FM),
florescimento feminino (FF), intervalo entre florescimentos (IF) e
stay-green (SG) de 196 progênies interpopulacionais em Campinas.
Safrinha 2012......................................................................................
47
Tabela 5 -
Resumo da análise de variância no delineamento em látice dos
caracteres massa de grãos (MG), prolificidade (PROL),
comprimento de espiga (CE), número de fileiras de grãos na espiga
(NFE) e número de ramificações do pendão (NRP) de 196
progênies interpopulacionais em Campinas. Safrinha
2012.....................................................................................................
47
Tabela 6 -
Resumo da análise de variância no delineamento em látice dos
caracteres altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (AC+Q), florescimento masculino (FM),
florescimento feminino (FF), intervalo entre florescimentos (IF) e
stay-green (SG) de 196 progênies interpopulacionais em Mococa.
Safrinha 2012......................................................................................
48
Tabela 7 -
Resumo da análise de variância no delineamento em látice dos
caracteres massa de grãos (MG), prolificidade (PROL),
comprimento de espiga (CE), número de fileiras de grãos na espiga
(NFE) e número de ramificações de pendão (NRP) de 196
progênies interpopulacionais em Mococa. Safrinha
2012.....................................................................................................
48
Tabela 8 -
Resumo da análise de variância conjunta dos caracteres altura de
planta (AP), altura de espiga (AE), plantas acamadas e quebradas
(AC+Q), florescimento masculino (FM), florescimento feminino
(FF), intervalo entre florescimentos (IF) e stay-green (SG) de 196
progênies interpopulacionais em Campinas e Mococa. Safrinha,
2012.....................................................................................................
49
Tabela 9 -
Resumo da análise de variância conjunta dos caracteres massa de
grãos (MG), prolificidade (PROL), comprimento de espiga (CE),

iii
número de fileiras de grãos na espiga (NFE) e número de
ramificações do pendão (NRP) de 196 progênies
interpopulacionais em Campinas e Mococa. Safrinha,
2012.....................................................................................................
49
Tabela 10 -
Médias de massa de grãos (MG), estimativas de heterose em
relação à média dos pais ( H ) e heterose relativa ( %H ) de 20% das
progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e
testemunhas em Campinas, na safrinha de 2012.................................
52
Tabela 11 - Médias de massa de grãos (MG), estimativas de heterose em
relação à média dos pais ( H ) e heterose relativa ( %H ) de 20% das
progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e
testemunhas em Mococa, na safrinha de 2012....................................
53
Tabela 12 - Médias de altura de planta (AP), altura de espiga (AE),
florescimento masculino (FM), florescimento feminino (FF) e
intervalo entre florescimentos (IF) de 20% das progênies
interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas
em Campinas, na safrinha de 2012.....................................................
59
Tabela 13 - Médias de plantas acamadas e quebradas (A+Q), número de
ramificações do pendão (NRP), stay-green (SG), de prolificidade
(PROL), comprimento de espiga (CE) e número de fileiras de grão
na espiga (NFE) de 20% das progênies interpopulacionais (PI(1,2))
selecionadas, no ciclo 0 e testemunhas em Campinas, na safrinha de
2012.....................................................................................................
60
Tabela 14 - Médias de altura de planta (AP), altura de espiga (AE),
florescimento masculino (FM), florescimento feminino (FF) e
intervalo entre florescimentos (IF) de 20% das progênies
interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas
em Mococa, na safrinha de 2012.........................................................
61
Tabela 15 - Médias de plantas acamadas e quebradas (A+Q), número de
ramificações do pendão (NRP), stay-green (SG), prolificidade
(PROL), comprimento de espiga (CE) e número de fileiras de grãos
na espiga (NFE) de 20% das progênies interpopulacionais (PI(1,2))
selecionadas, no ciclo 0 e testemunhas em Mococa, na safrinha de
2012.....................................................................................................
62
Tabela 16 - Estimativas da variância fenotípica (2
)2,1(ˆ
F ), da variância genética
entre progênies ( 2
)2,1(ˆ
p ), da variância aditiva ( )2,1(
2ˆA ), da
variância do erro (2
)2,1(ˆ
E ), da herdabilidade em sentido amplo
( 2
)2,1(ˆ
xh ), do coeficiente de variação genética (gVC ˆ ) e experimental
( eVC ˆ ), índice de seleção (b) e ganho de seleção ( )2,1(ˆsG ) para doze
caracteres avaliados em 196 progênies interpopulacionais, em
Campinas na Safrinha 2012................................................................
68

iv
Tabela 17 - Estimativas da variância fenotípica (2
)2,1(ˆ
F ), da variância genética
entre progênies ( 2
)2,1(ˆ
p ), da variância aditiva ( )2,1(
2ˆA ), da
variância do erro (2
)2,1(ˆ
E ), da herdabilidade em sentido amplo
( 2
)2,1(ˆ
xh ), do coeficiente de variação genética (gVC ˆ ) e experimental
(eVC ˆ ), índice de seleção (b) e ganho de seleção ( )2,1(
ˆsG ) para doze
caracteres avaliados em 196 progênies interpopulacionais, em
Mococa na Safrinha 2012....................................................................
69
Tabela 18 - Resumo da análise de variância para potencial de água na folha
( w ), clorofila total (Chl), condutância estomática (gs(1)
) (horário
de 9:00 a 10:30h), condutância estomática (gs(2)
) (horário de 11:00 a
12:30h), altura de planta (AP), altura de espiga (AE), florescimento
masculino (FM) e florescimento feminino (FF) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC Maya Latente
e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos
de irrigação, irrigação normal e déficit hídrico, no estádio de
florescimento. Campinas, 2013...........................................................
72
Tabela 19 - Resumo da análise de variância para intervalo entre florescimento
(IF), stay-green (SG), massa de grãos (MG), prolificidade (PROL),
matéria seca total (MST) e índice de colheita (IC) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC Maya Latente
e progênie sensível à seca – P.Inter-141), submetidas a dois
manejos de irrigação, irrigação normal e déficit hídrico, no estádio
de florescimento. Campinas, 2013......................................................
73
Tabela 20 - Média de massa de grãos (MG) e índice relativo de tolerância
(RVT) de 8 progênies interpopulacionais e 3 testemunhas
(DAS2B707, IAC Maya Latente e progênie sensível à seca – PI(1,2)-
141), submetidas a dois manejos de irrigação, irrigação normal e
déficit hídrico, no estádio de florescimento (VT). Campinas,
2013.....................................................................................................
74
Tabela 21 - Média de prolificidade (PROL), matéria seca total (MST), índice de
colheita (IC) e índice relativo de tolerância (RVT) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC Maya Latente
e progênie sensível à seca – P.Inter-141), submetidas a dois
manejos de irrigação, irrigação normal e déficit hídrico, no estádio
de florescimento (VT). Campinas, 2013.............................................
77
Tabela 22- Média de altura de planta (AP), altura de espiga (AE) e índice
relativo de tolerância (RVT) de 8 progênies interpopulacionais e 3
testemunhas (DAS2B707, IAC Maya Latente e progênie sensível à
seca – PI(1,2)-141), submetidas a dois manejos de irrigação, irrigação
normal e déficit hídrico, no estádio de florescimento (VT), em
Campinas, 2013...................................................................................
78

v
Tabela 23 - Média de florescimento masculino (FM), florescimento feminino
(FF) e índice relativo de tolerância (RVT) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC Maya Latente
e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos
de irrigação, irrigação normal e déficit hídrico, no estádio de
florescimento (VT), em Campinas, 2013............................................
79
Tabela 24-
Média de intervalo entre florescimentos feminino e masculino (IF),
stay-green (SG) e índice relativo de tolerância (RVT) de 8
progênies interpopulacional e 3 testemunhas (DAS2B707, IAC
Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a
dois manejos de irrigação, irrigação normal e déficit hídrico, no
estádio de florescimento (VT), em Campinas, 2012...........................
80
Tabela 25 - Média de potencial de água na folha ( w ), clorofila total (Chl) e
índice relativo de tolerância (RVT) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC Maya Latente
e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos
de irrigação, irrigação normal e déficit hídrico, no estádio de
florescimento (VT), em Campinas, 2013............................................
84
Tabela 26- Média de condutância estomática medida das 9:00 à 10:30h (gs(2)
) e
das 11:00 à 12:30h (gs(3)
) e índice relativo de tolerâcia (RVT) de 8
progênies interpopulacionais e 3 testemunhas (DAS2B707, IAC
Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a
dois manejos de irrigação, irrigação normal e déficit hídrico, no
estádio de florescimento (VT), em Campinas, 2013...........................
85

vi
LISTA DE FIGURAS
Figura 1 - Obtenção de progênies interpopulacionais de milho obtida do
intercruzamento da Pop.Tol.1 e IAC Maya Latente, na safra de
verão de 2011/12. Campinas, SP........................................................
32
Figura 2 - Temperaturas máxima e mínima e precipitação pluvial média da
localidade de Campinas, no período de 03/03/2012 à 03/08/2012....
34
Figura 3 - Temperaturas máxima e mínima e precipitação pluvial média da
localidade de Mococa no período de 29/03/2012 à 29/08/2012.........
34
Figura 4 -
Disposição do experimento realizado em casa de vegetação no
Centro Experimental Central do Instituto Agronômico (Campinas,
2013)...................................................................................................
41
Figura 5 - Aspecto visual das plantas em déficit hídrico (a) e (b); figura (b)
mostra os tratamentos irrigados (direita) e déficit hídrico
(esquerda), em casa de vegetação. Centro Experimental do Instituto
Agronômico, Campinas, 2013............................................................
43

vii
LISTA DE ANEXOS
Anexo 1 - Médias de massa de grãos (MG), estimativas de heterose em
relação à média dos pais ( H ), heterose relativa ( %H ) de 20% das
progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e
testemunhas em Campinas e Mococa, na safrinha de
2012....................................................................................................
104
Anexo 2 - Médias de altura de planta (AP), altura de espiga (AE),
florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), estimativas de heterose em
relação à média dos pais ( H ) e heterose relativa ( %H ) de 20%
das progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e
testemunhas em Campinas e Mococa, na safrinha de 2012...............
105
Anexo 3 - Médias de plantas acamadas e quebradas (A+Q), número de
ramificações do pendão (NRP), stay-green (SG), prolificidade
(PROL), comprimento de espiga (CE), número de fileiras de grãos
na espiga (NFE), estimativas de heterose em relação à média dos
pais ( H ) e heterose relativa ( %H ) de 20% das progênies
interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas
em Campinas e Mococa, na safrinha de 2012....................................
106
Anexo 4 -
Médias de massa de grãos (MG), altura de planta (AP), altura de
espiga (AE), florescimento masculino (FM), florescimento
feminino (FF), intervalo entre florescimentos (IF), plantas
acamadas e quebradas (A+Q), número de ramificações do pendão
(NRP), stay-green (SG), prolificidade (PROL), comprimento de
espiga (CE), número de fileiras de grãos na espiga (NFE),
estimativas de heterose em relação à média dos pais ( H ) e
heterose relativa ( %H ) de 8 progênies interpopulacionais
selecionadas em Campinas e Mococa, na safrinha de
2012....................................................................................................
108
Anexo 5 - Médias de altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (A+Q), florescimento masculino (FM),
florescimento feminino (FF), intervalo entre florescimentos (IF),
stay-green (SG), massa de grãos (MG) de 196 progênies
interpopulacionais (PI(1,2)) avaliadas em Campinas, na safrinha de
2012....................................................................................................
109
Anexo 6 - Médias de altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (A+Q), florescimento masculino (FM),
florescimento feminino (FF), intervalo entre florescimentos (IF),
stay-green (SG), massa de grãos (MG) de 196 progênies
interpopulacionais (PI(1,2)) avaliadas em Mococa, na safrinha de
2012....................................................................................................
114

viii
LISTA DE ABREVIATURAS
AP- Altura da planta
AE - Altura da espiga
A+Q - Plantas acamadas e quebradas
CE - Comprimento da espiga
FM - Florescimento masculino
FF - Florescimento feminino
IC - Índice de colheita
IF - Intervalo entre florescimentos
MG - Massa de grãos
MST - Matéria seca total
NFE - Número de fileiras de grãos na espiga
NRP - Número de ramificações do pendão
PROL - Prolificidade
RVT - Índice de valor relativo de tolerância
SG - Stay-green
Chl - Clorofila total
gs - Condutância estomática
w Potencial de água na folha

1
BERNINI, C.S. Seleção de progênies interpopulacionais de milho e estimativas de parâmetros
genéticos relacionados com tolerância à seca. 2015. 118p. Tese (Doutorado) – Instituto
Agronômico – IAC, Campinas (SP).
RESUMO
A seca é uma das maiores limitações no cultivo de plantas e um grande desafio para as
pesquisas científicas em agricultura. O estresse hídrico afeta a produtividade das culturas
especialmente o milho, ocasionando ameaça à segurança alimentar de vários continentes,
sendo de importância primordial os estudos de déficit hídrico aliados ao melhoramento
genético, bem como o entendimento de mecanismos genéticos e fisiológicos envolvidos com
a tolerância à seca. Diante isso, os objetivos do trabalho foram: 1) Avaliar progênies
interpopulacionais de milho em campo, quanto aos caracteres agronômicos e secundários
relacionados com tolerância à seca, em condições de safrinha; 2) Estimar parâmetros
genéticos das progênies interpopulacionais; 3) Estudar a introgressão do IAC Maya Latente
na Pop.Tol.1 quanto aos caracteres relacionados a tolerância à seca e 4) Avaliar alterações
fisiológicas de progênies tolerantes à seca selecionadas em campo e da variedade IAC Maya
Latente, a fim de identificar as respostas gerais de milho sob estresse e os mecanismos de
tolerância, em casa de vegetação. Em condições de campo foram avaliadas 196 progênies
interpopulacionais, oriundas do cruzamento da Pop.Tol.1 e o Maya Latente, em Campinas e
Mococa, na safrinha de 2012. Foram avaliados os caracteres florescimento masculino (FM) e
feminino (FF), intervalo entre florescimentos (IF), stay-green (SG), número de ramificações
de pendão (NRP), altura da planta (AP) e da espiga (AE), plantas acamadas e quebradas
(A+Q), prolificidade (PROL), comprimento de espiga (CE) e número de fileiras de grãos na
espiga (NFE) e massa de grãos (MG). O delineamento experimental foi o de látice 14x14 com
duas repetições. Foram estimados os parâmetros genéticos: variância genética entre progênies,
variância fenotípica entre as médias dos cruzamentos, variância aditiva interpopulacionais,
herdabilidade no sentido amplo ao nível de média de progênie e índice de seleção. Com base
nos resultados dos experimentos de campo, foram selecionadas 8 progênies interpopulacionais
tolerantes à seca, para serem avaliadas em casa de vegetação. Foram avaliados os seguintes
parâmetros fisiológicos: potencial de água na folha ( w ), clorofila total (Chl) e condutância
estomática (gs), e os biométricos: florescimento masculino (FM) e feminino (FF), stay-green
(SG), altura da planta (AP) e da espiga (AE), prolificidade (PROL), matéria seca total (MST),
índice de colheita (IC) e massa de grãos (MG). O delineamento experimental na casa de

2
vegetação foi o de blocos casualizados, em parcela subdividida com duas repetições. As
parcelas foram constituídas de tratamentos de estresse hídrico e as subparcelas foram
formadas pelas progênies. Foi obtido o índice de valor relativo de tolerância (RVT) e as
médias dos genótipos e tratamentos hídricos foram agrupadas pelo teste de Scott-Knott
(P≤0,05). Os resultados obtidos nos experimentos de campo indicaram progênies
interpopulacionais promissoras, sendo indicadas como fonte de tolerância à seca para
obtenção de novas populações tolerantes em programas de seleção recorrente e de híbridos.
Obtiveram-se altas estimativas de herdabilidade em sentido amplo ( 2
)2,1(ˆ
xh ) para os seguintes
caracteres: NFE, AP, AE, FM e SG, demonstrando elevado potencial a ser explorado pela
seleção interpopulacional. As progênies PI(1,2)-59, PI(1,2)-100, PI(1,2)-109, PI(1,2)-117 e PI(1,2)-
147 obtiveram as melhores performances de MG e parâmetros fisiológicos em casa de
vegetação e podem ser exploradas para extração de linhagens ou para obtenção de nova
população tolerante à seca. Os parâmetros fisiológicos w , gs e Chl, permitiram a
caracterização das progênies PI(1,2)-59, PI(1,2)-100, PI(1,2)-109, PI(1,2)-147, da variedade Maya
Latente e do DAS2B707, possibilitando que o melhorista utilize estas ferramentas na seleção
de genótipos tolerantes. Confirmou-se o padrão de sensibilidade da PI(1,2)-141 (Progênie
Sensível) pelos valores mais negativos de w , menor Chl, comportamento precoce de
senescência de folhas e colmos e baixa MG, em déficit hídrico. A variedade IAC Maya
Latente apresenta comportamento produtivo estável nos ambientes de irrigação normal e
déficit hídrico, devido à maior sensibilidade (fechamento) dos estômatos, resultando no maior
acúmulo de matéria seca total e pouca redução do índice de colheita, em estresse hídrico.
Palavras-chave: Zea mays L.; Seleção recorrente interpopulacional, parâmetros genéticos e
fisiológicos; estresse hídrico.

3
BERNINI, C.S. Selection of interpopulation progenies of maize and estimates of genetic
parameters related of drought tolerance. 2014. 118p. Agronomic Institute – IAC, Campinas
(SP).
ABSTRACT
Drought is one of the major limitations for the cultivation of plants and a great challenge for
the scientific research in agriculture. The drought stress affects the productivity of crops
especially maize, causing threat to the food security of several continents, being of main
importance the studies of water deficit together to genetic improvement, as well as the
understanding of genetic mechanisms and physiological involved with tolerance to drought.
The objectives of this study were: 1) Evaluate of interpopulation progenies of maize in the
field, regarding the agronomic traits and secondary related to drought tolerance, in conditions
of second crop; 2) Estimate genetic parameters of interpopulation progenies; 3) Study of
introgression to IAC Maya Latent in Pop.Tol.1 as the traits related to drought tolerance and 4)
Evaluate physiological alterations of progenies tolerant to drought selected in the field, and
the IAC Maya Latent, in order to identify the general responses of maize under stress and the
mechanisms used to achieve the tolerance, in the greenhouse. In field conditions were
evaluated 196 interpopulation progenies, derived from the cross of Pop.Tol.1 and the Maya
Latent, in Campinas and Mococa, in the second crop of 2012. The traits male (FM) and
female flowering (FF), interval between flowering (IF), stay-green (SG), tassel branches
number (NRP), plant (AP) and ear (AE) height, lodging and broken plants (A+Q), prolificacy
(PROL), ear length (CE) and number of kernels rows per ear (NFE) and grains mass (MG)
were evaluated. The experimental design was the lattice 14x14 with two repetitions. Were
estimated the genetics parameters: genetic variance between progeny, phenotypic variance
between mean of crosses, additive variance interpopulation, broad sense heritability at the
level of mean progeny and selection index. Based on the results of field experiments, were
selected 8 interpopulation progenies tolerant to drought, to be evaluated in a greenhouse. The
following physiological parameters were evaluated: leaf water potential ( w ), total
chlorophyll (Chl) and stomatal conductance (gs), and the biometric: male (FM) and female
flowering (FF), stay-green (SG), plant (AP) and ear (AE) height, prolificacy (PROL), total dry
matter (MST), index harvest (IC) and grains mass (MG). The experimental design of the
greenhouse was randomized blocks in a split plot scheme in two repetitions. The plot

4
consisted of treatments of drought stress and the sub-plots were formed by progenies. It was
obtained the index of relative value of tolerance (RTV) and the mean of genotypes and water
treatments were grouped by Scott-Knott test (P≤0.05). The results obtained in field
experiments indicated interpopulation progenies promising, being indicated as source of
drought tolerance for obtaining to new tolerant populations in programs of recurrent selection
and hybrid. Obtained higher estimates of heritability in the broad sense ( 2
)2,1(ˆ
xh ) for the
following characters: NFE, AP, AE, FM and SG, demonstrating high potential to be explored
by the selection interpopulation. The progenies PI(1,2)-59, PI(1,2)-100, PI(1,2)-109, PI(1,2)-117 and
PI(1,2)-147 obtaining the best performance of MG and physiological parameters in greenhouse
and can be exploited for the extraction of lines inbred or obtainment of new population
tolerant to drought. The physiological parameters w , gs and Chl, allowed the
characterization of progenies PI(1,2)-59, PI(1,2)-100, PI(1,2)-109, PI(1,2)-147, of the variety Maya
Latent and DAS2B707, allowing the breeder to use these tools in the selection of genotypes
tolerant. It was confirmed that the standard of sensitivity of PI(1,2)-141 (Progeny Sensitive) by
more negative values of w , lower Chl, behavior early senescence of leaves and stems and
low MG in water deficit. The IAC Maya Latent present productive behavior in stable
environments of normal irrigation and water deficit, due to the higher sensitivity (closure) of
stomata resulting in greater accumulation of total dry matter and little reduction in the index
of harvest, in hydric stress.
Keywords: Zea mays L.; interpopulation recurrent selection, genetics and physiological
parameters, drought stress.

5
INTRODUÇÃO
O milho (Zea mays L.) é uma das principais culturas de grãos de maior interesse no
mundo, ocupando uma área de 177 milhões de hectares e produção de 872 milhões de
toneladas (FAOSTAT, 2012). No Brasil, a produção total é de 78 milhões de toneladas, sendo
cultivado principalmente na safrinha, representando 59% da produção total de grãos
(CONAB, 2014). A safrinha pode ter sua produtividade bastante afetada devido às perdas que
acontecem em função do estresse hídrico nos períodos críticos da cultura. Isto ocorre pelo fato
dessa safra de milho (implantada no período de janeiro a março) ser cultivada após uma
cultura de verão, portanto, em condições de risco de instabilidade climática devido aos
períodos de estiagem denominados veranicos. O comportamento climático, principalmente, o
déficit hídrico pode variar de safrinha a safrinha, o que resulta em flutuações na
produtividade.
A seca é a causa mais severa da redução da produção agrícola e o aquecimento global
previsto agravará, consideravelmente, o impacto do déficit hídrico (RIBAUT et al., 2009).
Alterações climáticas, os custos elevados da irrigação e a escassez de água para esta
finalidade somados a degradação da capacidade de retenção da água do solo, devido à erosão
ou compactação são outros fatores que podem ampliar a frequência da seca (EDMEADES et
al., 2006), aumentando a necessidade de melhoramento genético para tolerância à seca em
milho (BÄNZIGER & ARAUS, 2007). Uma estratégia seria a obtenção de cultivares
tolerantes que possam escapar do estresse hídrico durante os períodos mais críticos da cultura,
com a seleção de germoplasma que possua variabilidade genética para os caracteres
adaptativos relacionados à seca (BECK et al., 1997; VASAL et al., 1997).
Nos estudos de MIRANDA et al. (1982) com a variedade IAC IAC Maya Latente, que
foi desenvolvida com a incorporação do fator “latente”, os autores relataram uma resposta
distinta à seca e ao frio. O mecanismo de tolerância à seca denominado fator “latente”,
consiste em prolongar o estádio vegetativo sob estresse hídrico permitindo que o
florescimento ocorra logo após as chuvas iniciarem, tendo o rápido desenvolvimento para
assim completar o ciclo reprodutivo (FISHER et al., 1982; HAYANA-KANASHIRO et al.,
2009).
A tolerância à seca é um caráter complexo, que depende da ação e interação de
diferentes processos fisiológicos, bioquímicos e moleculares que permitem à planta

6
sobreviver e produzir satisfatoriamente. O desenvolvimento de uma cultivar tolerante, em
uma abordagem promissora, requer, entre outros, o conhecimento dos mecanismos
fisiológicos e do controle genético que contribuem para o fenótipo nos diferentes estádios de
crescimento das plantas. Apesar dos mecanismos de tolerância à seca variarem de espécie
para espécie nos diferentes estádios (FOOLAD & LIN, 2001), as respostas celulares a um
estresse abiótico específico se mantêm conservadas em muitas espécies de plantas (ZHU,
2002). Assim, caracteres diferentes têm sido propostos como critério de seleção para melhorar
os níveis de tolerância à seca.
Vários autores relatam que os principais caracteres secundários são: intervalo entre
florescimentos feminino e masculino, senescência retardada de colmos e folhas (stay-green),
número de ramificações de pendão, altura de planta e prolificidade, e os fisiológicos:
potencial de água na folha, condutância estomática, conteúdo de clorofila e matéria seca total.
Com isso é sugerido identificar quais respostas fisiológicas e secundárias das plantas estão
envolvidas no processo de seleção em déficit hídrico para inseri-los nos programas de
melhoramento de milho. Contudo, o conhecimento sobre mecanismos de tolerância
fisiológica ainda é incipiente para a contribuição do aumento da produtividade de grãos
quando ocorre déficit hídrico (BÄNZIGER et al., 2000).
O melhoramento de populações visa essencialmente ao aumento dos genes favoráveis
nas populações base. Os acréscimos nas frequências alélicas são funções que dependem da
magnitude da ação gênica, do processo de seleção, da população base, da intensidade de
seleção e da precisão experimental. A variabilidade genética, contudo, pode ser encontrada a
partir de novas combinações e intercruzamentos daqueles genótipos que possuem caracteres
desejáveis (ASHRAF, 1994). Diante disso, diferentes procedimentos de seleção recorrente
foram desenvolvidos e a eficácia de cada método depende da população submetida à seleção,
das características selecionadas e do objetivo do programa de melhoramento (VASAL et al.,
1997). O método interpopulacional é efetivo para o melhoramento de duas populações e da
heterose que se manifesta no cruzamento, assim como na seleção de híbridos tolerantes à seca
e no aumento da probabilidade de obtenção de linhagens superiores. Desta forma, é necessário
identificar quais são os atributos fisiológicos que contribuem para a adaptação à seca,
objetivando combiná-los seletivamente, nos materiais altamente produtivos e adaptados aos
sistemas de produção atuais, pelo melhoramento genético.
Ante o exposto, o presente trabalho teve por objetivos: 1) Avaliar progênies
interpopulacionais de milho em campo, quanto aos caracteres agronômicos e secundários

7
relacionados com a tolerância à seca, em condições de safrinha; 2) Estimar parâmetros
genéticos das progênies interpopulacionais; 3) Estudar a introgressão do IAC IAC Maya
Latente na Pop.Tol.1 quanto aos caracteres relacionados a tolerância à seca e 4) Avaliar
alterações fisiológicas de progênies tolerantes à seca selecionadas em campo e da variedade
IAC IAC Maya Latente, a fim de identificar as respostas gerais de milho sob estresse e os
mecanismos de tolerância, em casa de vegetação.
2 REVISÃO DE LITERATURA
2.1 Efeito do déficit hídrico em milho
O termo “estresse” é definido como desvio significativo das condições ótimas para a
vida, que induz mudanças e respostas em todos os níveis funcionais do organismo, as quais
são reversíveis a princípio, mas podem se tornar permanentes (LARCHER, 2004). Para
SCHULZE et al. (2005) o termo é usado em situações onde há o desvio do normal fisiológico
em resposta aos fatores ambientais (subótimo ou quantidades danosas). Para esses autores, o
estresse abiótico restringe o desenvolvimento das plantas, provocando um desequilíbrio
termodinâmico das células que aumenta seus gastos enérgicos.
Todos os processos vitais das plantas, da multiplicação e crescimento das células à
fotossíntese, dependem do estado hídrico celular. Assim, um déficit de água leva ao
desequilíbrio fisiológico, que repercute negativamente sobre o desenvolvimento e
produtividade da planta. A queda de produtividade do milho que ocorre em função do estresse
hídrico depende do estádio de desenvolvimento em que se encontra a planta, sendo que a
consequência e o impacto de um estresse hídrico específico dependem, também, do nível
máximo de déficit hídrico (BLUM, 2010).
O estresse hídrico pode reduzir a produtividade das culturas afetando as áreas de
produção, denominadas fontes, e as áreas de metabolismo ou armazenamento, chamadas de
drenos. A capacidade da fonte é reduzida como resultado do efeito do estresse na área foliar,
trocas gasosas, armazenamento de carbono disponível para o enchimento dos grãos e aumento
da senescência foliar (BLUM, 2010). As fontes incluem qualquer órgão exportador, como por
exemplo, as folhas maduras, que são capazes de produzir fotoassimilados. Os drenos incluem
órgãos como as raízes, frutos em desenvolvimento e folhas imaturas, que devem importar
carboidratos para seu desenvolvimento (TAIZ & ZEIGER, 2006).

8
O milho é uma espécie C4, com mecanismos específicos de concentração de CO2, que
inicialmente envolvem a fixação de CO2 pela fosfoenolpiruvato carboxilase (PEPC) nas
células do mesofilo, formando um ácido de quatro carbonos, seguido pela sua descarboxilação
nas células da bainha que resulta a liberação de CO2 reassimilado pela ribulose-1-5-bifosfato
carboxilase/oxigenase (Rubisco). O aumento da concentração de CO2 nas células da bainha
resulta em uma baixa atividade da ribulose 1-5-bifosfato oxigenase e consequentemente baixa
taxa de fotorrespiração e aumento da taxa fotossintética (TAIZ & ZEIGER, 2006). Essa
espécie apresenta potencial para alcançar altas produtividades quando a máxima área foliar
coincidir com a maior disponibilidade de radiação solar, utilizando a água eficazmente. O
requerimento de água de qualquer cultura é muito dependente do ambiente (temperatura e
umidade do solo), na qual é cultivada. O milho requer 500-800 mm de água durante seu ciclo
de 80 a 110 dias (CRITCHLEY & KLAUS, 1991). De acordo com JAMIESON et al. (1995),
o requerimento de água para o milho no período de pré-florescimento é 135 mm/mês (4,5
mm/dia), podendo ter um aumento acima de 195 mm/mês (6,5 mm/dia), nos dias com
temperaturas elevadas.
O desenvolvimento do milho é dividido em duas fases principais: (1) estádio vegetativo,
da emergência das plântulas até o pendoamento, (2) estádio reprodutivo, do embonecamento e
polinização até a maturidade fisiológica (RITCHIE et al., 1993). Os estádios vegetativos e
reprodutivos do milho se encontram esquematizados na tabela 1. As subdivisões dos estádios
vegetativos são designadas pela letra “V”, mais um valor numérico, que representa o número
de folhas totalmente expandidas até o momento da observação. Assim, nos estádios
vegetativos de V1 até Vn, ṉ representa o número total de folhas expandidas antes do estádio
VT (pendoamento). Os estádios reprodutivos são designados pela letra “R”.

9
Tabela 1. Descrição dos estádios de desenvolvimento do milho.
Estádio Descrição
VE germinação e emergência
V1 primeira folha desenvolvida; alongamento das primeiras raízes
V3
três folhas desenvolvidas; todas as folhas e espigas que a planta irá
produzir estão sendo formadas; ocorre aproximadamente duas
semanas após a emergência
V5
cinco folhas completamente desenvolvidas; a iniciação do pendão já
pode ser observada microscopicamente na extremidade de formação
do caule, logo abaixo da superfície do solo
V6 o ponto de crescimento e o pendão estão acima do nível do solo; o
colmo está iniciando um período de alongação acelerada
V7 e V8 fases de rápido crescimento; definição do número das fileiras de grãos
V9 e V10 alta taxa de desenvolvimento de órgãos florais; rápido e contínuo
crescimento com acumulação de nutrientes e matéria seca
V12 são definidos o número de óvulos e o tamanho da espiga
V15 e V17 estilos-estigmas iniciam o crescimento nas espigas; espigas visíveis
no caule; a extremidade do pendão já pode ser observado (V17)
V18 a planta de milho encontra-se a uma semana do florescimento
VT pendoamento; o último ramo do pendão está completamente visível e
os estilos-estigmas ainda não emergiram
R1 embonecamento e polinização; estilos-estigmas estão visíveis, para
fora das espigas
R2 grão bolha d’água
R3 grão leitoso; ocorre normalmente de 12 a 15 dias após a polinização;
o grão tem aparência amarela, e no seu interior, um fluído leitoso
R4 grão pastoso; os grãos encontram-se com cerca de 70% de umidade
R5 formação de dente
R6 maturidade fisiológica; ocorre cerca de 50 a 60 dias após polinização
FONTE: RITCHIE et al. (1993)

10
O milho é mais suscetível ao déficit hídrico nos estádios de pré-florescimento,
florescimento e enchimento de grãos, a ponto de não conseguir recuperar em reidratação.
Segundo BERGAMASCHI et al. (2006) a susceptibilidade do milho nessas fases é devido à
ocorrência dos processos fisiológicos ligados a formação do zigoto e início do enchimento de
grãos, além da elevada transpiração decorrente da máxima área foliar e da elevada carga
energética proveniente da radiação solar.
A maior parte dos efeitos do déficit hídrico sob o crescimento da cultura é devido às
reduções na área foliar, que afetarão a exposição à luz incidente durante o período crítico da
floração (WESTGATE et al., 2004). SALAH & TARDIEU (1997) relataram que o
crescimento é mais afetado do que o desenvolvimento, sendo a redução na área foliar
explicada, essencialmente, pela redução na taxa de expansão foliar devido à perda de turgor.
A inibição da expansão celular provoca um reduzido consumo de carbono e energia e uma
proporção maior de assimilados vegetais que pode ser distribuída ao sistema subterrâneo,
onde pode sustentar o crescimento posterior de raízes. A área foliar menor transpira menos,
conservando efetivamente um suprimento de água limitado no solo por um período mais
longo (TAIZ & ZEIGER, 2006).
Em muitas espécies a redução ou paralização da expansão celular é um dos processos
mais sensíveis, se não for o mais sensível, ocasionado na planta pelo estresse hídrico, quando
uma pequena redução no potencial de pressão causa, aparentemente, uma desaceleração da
elongação celular (HSIAO et al., 1973). Os autores acrescentam que o estresse hídrico reduz o
turgor das células e, consequentemente, o seu crescimento, além de reduzir a atividade da
redutase do nitrato e ocasionar um incremento na síntese de ácido abscísico (ABA), que
resulta no fechamento estomático e diminuição na assimilação de CO2.
A importância da determinação da área foliar é devido a correlação entre esta e as taxas
fotossintéticas e transpiração das plantas, uma vez que reflete a capacidade da planta em
interceptar as radiações e efetuar trocas gasosas com o ambiente, constituindo-se, assim, em
um importante indicativo da produtividade da cultura (SOUZA et al., 2011). DOW (1981)
concluiu que genótipos tolerantes à seca são aqueles relacionados a menor área foliar,
sugerindo que o desenvolvimento da área foliar pode afetar o desenvolvimento da espiga.
Quanto ao tamanho da planta, FISCHER et al. (1982) relatam que o milho tolerante ao
estresse hídrico tem pequeno porte em relação ao não tolerante. ÇAKIR (2004) estudando o
efeito do déficit hídrico no milho na fase vegetativa, entre V6 e VT, e reprodutiva, entre VT e
R3 e entre R3 e R6, acrescido de todas as combinações possíveis entre essas épocas, observou

11
drástica redução da altura de plantas quando o corte no suprimento de água ocorreu a partir do
estádio V6.
O estresse hídrico no período do florescimento reduz, consideravelmente, a produção de
grãos (DWIVEDI et al., 2010). A grande susceptibilidade do milho neste estádio, quando
comparada a outros cereais exceto o arroz, está associada à própria estrutura floral, que
apresenta as flores femininas e masculinas separadas em uma única haste, cujas espiguetas se
desenvolvem quase ao mesmo tempo, numa mesma espiga; e à competição entre os órgãos
reprodutivos masculino e feminino pelos fotossíntatos translocados (MONNEVEUX &
RIBAULT, 2006; BÄNZIGER & ARAUS, 2007).O déficit hídrico provoca atraso na
emergência dos estilos-estigmas, aumentando o intervalo entre os florescimentos masculino e
feminino seguido de uma alta porcentagem de zigotos abortados após a fertilização. O aborto
é controlado pela inflorescência feminina, que apresenta maior sensibilidade do que o pólen
ao déficit hídrico (BOYER & WESTGATE, 2004; WETSGATE et al., 2004). Dependendo da
intensidade do estresse pode não ocorrer o desenvolvimento do tubo polínico, levando a
esterilidade completa da planta. A falha reprodutiva está relacionada ao baixo status hídrico e
alto conteúdo de ácido abscísico. ZINSELMEIER et al. (1995) verificaram que a atividade da
enzima invertase foi inibida e o teor de amido nos ovários foi reduzido. Ao fazer a
suplementação de sacarose nos ovários, o amido e a atividade da invertase foram restaurados.
Isto demonstrou que a deficiência de açúcar nos ovários de milho foi determinante para causar
aborto e a invertase está diretamente implicada como fator limitante para produção de grãos
sob estresse hídrico (BLUM, 2010).
O déficit hídrico durante a fase de florescimento, além de afetar o sincronismo entre
florescimentos feminino e masculino, pode reduzir a chance de aparecimento de uma segunda
espiga em materiais considerados prolíficos (EDMEADES et al., 1999). O período
correspondente a alguns dias antes e cerca de 25 dias após o embonecamento foi identificado
como sensível ao déficit hídrico, reduzindo drasticamente o número de grãos na espiga
(BARKER et al., 2005).
Cerca de duas a três semanas após a polinização, os grãos entram em uma fase linear de
acumulação de biomassa, tornando-se drenos fortes para as reservas acumuladas nas folhas e
colmo. Se os grãos conseguem atingir esse estádio, eles normalmente atingem pelo menos
30% do peso observado em plantas não submetidas a estresse hídrico (BOLAÑOS &
EDMEADES, 1996; MAGALHÃES & SOUZA, 2012). Geralmente, o estresse hídrico
durante o enchimento dos grãos resulta na produção de sementes pequenas. Dentre as

12
possíveis explicações fisiológicas para este fato pode-se citar a perda do fornecimento pelo
fluxo dos fotossíntatos devido à senescência prematura das folhas e a diminuição na
capacidade dos grãos, em processo de enchimento, de estocar e metabolizar os açúcares
importados (capacidade-dreno). Nesse estádio, as reservas de carboidratos do caule podem ser
translocadas para apoiar o desenvolvimento do grão (WESTGATE et al., 2004). Os milhos
tropicais são mais dreno limitantes do que fonte limitantes. Visando produtividade de grãos, a
parte aérea ainda é considerada importante e fundamental para o bom desenvolvimento e
crescimento das plantas (BORRÁS et al., 2004).
No milho, o estresse hídrico também leva à senescência prematura das folhas, que
prejudica severamente a quantidade total de compostos carbonatos sintetizados na fotossíntese
(GRANIER & TARDIEU, 1999; EDMEADES et al., 2006; MASCLAUX-DAUBRESSE et
al., 2007). A senescência é um processo ativo de desenvolvimento, caracterizado pela perda
de clorofila e regulada por genes específicos (DWIVEDI et al., 2010). Neste processo
estruturas celulares e macromoléculas são decompostas e translocadas do órgão senescente
para regiões de crescimento ativo que servem como drenos de nutrientes (TAIZ & ZEIGER,
2006; BLUM, 2010).
Os hormônios de plantas estão envolvidos no controle da senescência das folhas, onde
geralmente o ácido abscísico (ABA), que é importante por controlar as respostas ao estresse
hídrico, promove a senescência e o atraso da citocinina. Consequentemente, a atuação da
citocinina melhora o stay-green e o teor de clorofila nas plantas (GAN & AMASINO, 1995).
A alta expressão de produção de citocinina produziu em plantas de tabaco transgênico durante
o estádio reprodutivo um retardo da senescência das folhas e resultou no melhor status hídrico
da folha, atuando na recuperação do déficit hídrico e alta produtividade quando comparado
com a variedade convencional (RIVERO et al., 2009). Em contraste da citocinina, o ácido
abscísico atua no avanço da senescência e exporta carbono e nitrogênio das folhas de cereais
como o arroz (YANG et al., 2003). O efeito do ABA na senescência das folhas é mediada
pela produção de etileno, sendo este hormônio conhecido como responsável pela senescência
das folhas (BUCHANAN-WOLLASTON, 1997).
Os déficits hídricos moderados afetam o desenvolvimento do sistema radicular.
Geralmente, o sistema radicular profundo das plantas é maior em cultivares tolerantes à seca e
diversos trabalhos relatam esse aprofundamento em déficit hídrico (KUCHANUR, 2010).
Antes do florescimento, as raízes de milho crescem extensivamente em comprimento,
densidade e peso seco e sua capacidade de capturar água normalmente alcança o máximo

13
quando próximo do estádio de florescimento (MENGEL & BARBER, 1974). LIEDGENS et
al. (2000) estudaram raízes em milho, onde observaram que a máxima densidade de raízes
não ocorria ao mesmo tempo na época de florescimento, em contraste da máxima exposição
da área foliar. Na menor profundidade foi encontrada a maior densidade de raízes que
coincidiu ao mesmo tempo com o florescimento masculino, enquanto que na profundidade de
100 cm, a máxima densidade de raízes ocorria com 2 semanas após o florescimento
masculino. Os autores observaram a máxima exposição das raízes em 10 dias antes do
florescimento e concluíram que a máxima densidade de raízes foi alcançada com algum atraso
do desenvolvimento vegetativo na cultura. O comprimento e a densidade das raízes são
bastante utilizados em testes de campo e considerados como caráter secundário ideal. Estas
são importantes na aquisição de água e um componente significativo de tolerância a estresse
hídrico (MCCULLY, 1999).
2.2 Parâmetros fisiológicos em resposta ao déficit hídrico
Os parâmetros fisiológicos são usados na seleção de genótipos tolerantes à seca, pois
são mensuráveis e herdáveis; responsivos à seleção e correlacionados com o desenvolvimento
da cultura (RICHARDS, 1978). Alguns parâmetros fisiológicos de plantas e respostas
adaptativas à seca levam ao melhor entendimento de mecanismos fisiológicos utilizados pelos
genótipos tolerantes e sensíveis, a fim de discriminá-los para inseri-los em programas de
melhoramento para tolerância à seca.
Vários mecanismos são úteis para amenizar os danos causados pelo estresse hídrico, e
incluem escape, evitância e tolerância à seca em baixo status hídrico na planta. No escape à
seca é observada a atuação de mecanismos morfofisiológicos, principalmente a precocidade e
redução da área foliar. Na tolerância à seca, as plantas reduzem a transpiração ou aumentam a
absorção de água (CAVATTE et al., 2011). A tolerância em baixo status hídrico é definido
como a habilidade da planta em manter relativamente alto nível de hidratação sob condições
de estresse hídrico atmosférico ou do solo, sendo medida a partir do status hídrico da folha ou
tecido, que expressa o conteúdo relativo de água, sendo controlada por caracteres
constitutivos e adaptativos de plantas. A manutenção do turgor é também um componente de
tolerância a baixo status hídrico da planta.
LEVITT (1980) preconiza dois tipos de plantas com respeito à tolerância em baixo
status hídrico de plantas: i) as que evitam a desidratação pela reduzida transpiração e eficiente
uso da água e ii) as plantas que usam outros meios de reduzir a transpiração, evitando a

14
desidratação das células. A importância dessas implicações de cada fenótipo para evitar a
desidratação leva a melhor compreensão da produção no campo. A evidência sugerida no
cultivo de plantas sugere que a combinação de respostas pode existir em uma espécie,
provavelmente como resultado de seleção de melhoramento (BLUM, 2010).
GRANT (1992) considera que o desenvolvimento de mecanismos de adaptação das
plantas é influenciado pelo nível de CO2, radiação solar, temperatura e umidade relativa do ar.
As cultivares tolerantes à seca possuem distintos caracteres fisiológicos adaptativos que
asseguram a produtividade em condições de déficit hídrico. Os estudos fisiológicos têm
identificado esses caracteres como sendo: menor decréscimo do potencial hídrico, redução nas
taxas de assimilação de CO2, prolongado fechamento dos estômatos e maiores conteúdo de
clorofila (WANG et al., 2008). Devido às variações no conteúdo de água das células e tecidos,
algumas vias de sinalização são ativadas em decorrência das alterações ambientais. Outras
vias são ativadas por hormônios de plantas que sinalizam alterações hídricas (CHAVES et al.,
2003).
O potencial hídrico ( w ) é influenciado por vários componentes importantes para a
célula, dentre esses efeitos cita-se potencial de: osmótico ( s ), pressão (p ), matricial ( m )
e gravitacional (g ), sendo o potencial gravitacional desprezível em plantas de pequeno
porte. Portanto, o potencial hídrico e seus componentes são expressos como segue, eq. (1):
mpsw (1)
onde: os subscritos s, p e m correspondem aos potenciais de osmótico, pressão e matricial,
respectivamente. Cada componente é algebricamente aditivo conforme se aumenta (positivo)
ou diminui (negativo) o potencial hídrico quando comparado pela referência de energia livre
da água pura. Considerando que w da energia livre da água é nula, o w da célula vegetal é
sempre negativo.
O potencial hídrico é considerado um parâmetro confiável para medir a resposta da
planta ao déficit hídrico. Isso varia de acordo com o tipo de planta e das condições ambientais
(SILVA et al., 2011). De acordo com BLUM (1988), o potencial hídrico representa a
diferença de energia livre da água contida no citosol e a água pura nas mesmas condições de
temperatura e pressão, medido em unidades de pressão negativa em Bars ou Mega Pascals
(MPa) (BLUM, 2010). A importância da mesma refere-se à absorção da água e íons do solo;
transporte de água entre partes da planta; controle da abertura e fechamento dos estômatos;
fotossíntese; transporte de fotoassimilados; expansão e crescimento celular.

15
Em condições de déficit hídrico o potencial hídrico das plantas e o turgor são reduzidos
suficientemente para alterar suas funções vitais. Segundo HSIAO et al. (1973), a maioria das
respostas das plantas ao estresse hídrico (crescimento da célula, síntese de parede celular e
proteína, atividade enzimática, etc.) é influenciada por reduções de potencial hídrico
inferiores a -1,5 MPa. BERGONCI (1997) observou que acima de um potencial mínimo de
água na folha de -1,5 MPa, as plantas de milho recuperavam sua condição hídrica após o
fechamento estomático, sendo esse valor possível de ser utilizado como indicador de estresse
hídrico. BONO et al. (2001) classificaram as condições hídricas em três intervalos
representativos: w > -1,5 MPa como alto déficit, w entre -1,5 e -0,8 MPa como médio
déficit e w < -0,8 MPa como sem déficit. É observado, em estresse hídrico moderado
(potencial hídrico de -1,0 MPa), o acúmulo de solutos como glicina-betaína, prolina,
aminoácidos solúveis, proteínas e açúcares solúveis (FARHAD et al., 2011). Esses solutos
podem se acumular em grandes quantidades sob condições de baixo potencial hídrico,
protegendo a célula contra a desidratação por meio do ajustamento osmótico (ZHOU & YU,
2010).
O ajustamento osmótico mantém o conteúdo de água na célula pelo acúmulo de solutos
orgânicos no citoplasma, contribuindo para sustentar a alta taxa fotossintética e expansão para
o crescimento. O mecanismo do ajuste osmótico é importante para evitar a diminuição do
potencial hídrico celular, mediante a biossíntese de solutos intracelulares, que possuem baixo
peso molecular, se acumulam em altas quantidades no vacúolo ou no citosol e mantêm a
turgidez e o volume celular para preservar a integridade de compostos e estruturas celulares
fundamentais para o adequado metabolismo vegetal (ABDUL JALEEL et al., 2007). Um
exemplo é o aminoácido prolina que se acumula nas células e tem função osmoprotetora. Isto
previne a desnaturação de proteínas, preserva a estrutura de enzimas e protege as membranas
dos efeitos deletérios causados pelas espécies reativas de oxigênio e age como um tampão
para regular o potencial redox celular (ASHRAF & FOOLAD, 2007).
Os estômatos podem ser considerados como responsáveis pela superfície das folhas
auxiliando na abertura para permitir a absorção de CO2 e fechamento para evitar a perda de
água. O controle da abertura dos estômatos é influenciado por estímulos ambientais,
principalmente, intensidade e qualidade de luz, temperatura, umidade relativa, concentração
intracelular de CO2 e da quantidade de água disponível nas raízes das plantas (BLUM, 2010).
Os estômatos são células altamente especializadas na troca de gás e perdem água por meio da
alta transpiração foliar. O mecanismo que regula entrada, saída de CO2 e água nas folhas é

16
chamado de condutância estomática, que constitui outro parâmetro fisiológico importante.
Conforme LARCHER (2004) pode-se considerar a condutância como uma medida da
permeabilidade com que a água e o CO2 difundem-se através da folha. A condutância foliar
ao vapor de água é constituída pelas condutâncias estomática, cuticular e dos espaços
intercelulares. Dentre estas, a condutância estomática assume maior importância por ser a
principal via por onde ocorrem as trocas gasosas entre planta e atmosfera. A alta densidade de
estômatos tem um papel importante na condutividade da folha principalmente em condições
irrigadas. Quando o estresse hídrico ocorre, o fechamento estomático é o principal controle da
resistência. As células guarda sentem os sinais do ambiente, funcionando como células motor
dentro do complexo estomático. O controle estomático da condutância foliar à água é um
importante mecanismo através do qual o vegetal limita a perda de água, podendo ser utilizada
como indicador de déficit hídrico (PAIVA et al., 2005).
Para adaptação da planta à seca, o fechamento estomático pode ser considerado como
uma resposta direta da redução do potencial hídrico da folha induzido pelo déficit hídrico.
Conforme JONES (1985) o fechamento estomático pode ocorrer dentro de uma larga faixa de
potencial hídrico na folha em torno de -1,7 MPa. A preservação de água pelo fechamento
estomático é um importante mecanismo de tolerância à seca que conserva a fotossíntese da
perda de água (TEARE et al., 1973). Em plantas, na qual passaram por murcha e reidratação,
os estômatos permanecem fechados por vários dias, mesmo se há rápida recuperação da
turgescência celular (HIRON & WRIGHT, 1973). Em baixa disponibilidade hídrica há um
grande acúmulo de ácido abscísico (ABA) regulando a condutância estomática. Seu acúmulo
em folhas estressadas exerce grande papel na redução da perda de água pela transpiração
(ZHU, 2002) e também reduz a extensão das folhas (murcha das folhas). O fechamento
estomático também pode ser causado pelo aumento do transporte de ABA (através do
xilema), produzido nas raízes, em contato com o solo seco, para a parte aérea
(SCHACHTMAN & GOODGER, 2008; MAGALHÃES & SOUZA, 2012).
Entretanto, o melhoramento para tolerância à seca pode ser direcionado na seleção de
fenótipos com eficiência das funções estomáticas (comportamento e frequência), somente
quando outros processos importantes estão envolvidos na produção de grãos sob déficit
hídrico. A atividade fotossintética é relativamente menos sensível ao déficit hídrico foliar
quando comparado ao estômato (SACCARDY et al., 1996). No entanto, a manutenção da
condutância estomática durante o período de déficit hídrico é crucial para fotossíntese e
produção de grãos. É visto que a elevada produtividade de grãos obtida pelas culturas como

17
algodão, trigo e arroz mantida pela alta condutância estomática e transpiração em condições
de estresse e não-estresse, é indicativo da eficiência dos genótipos em manter as trocas
gasosas nas folhas ou temperatura foliar. O papel de evitar a desidratação das células em
relação aos estômatos pelas trocas gasosas está relacionado por manter o suprimento de água,
da sustentação da hidratação foliar e da turgescência, observado pelo lento fechamento dos
estômatos em condições de déficit hídrico (BLUM, 2010). Uma maior sensibilidade dos
estômatos (maior controle estomático) frente ao estresse hídrico resulta em maior eficiência
no uso da água (EUA), sendo uma das mais importantes estratégias em plantas tipo C4
(LOPES et al., 2011).
GU et al. (1990) relataram que a cultivar de milho Yuvu n°3 foi tolerante à seca por
apresentar reduzida perda de água utilizando os mecanismos de resistência estomática e
potencial osmótico. GUIMARÃES et al. (2014) estudaram diversos mecanismos fisiológicos
em resposta à seca, e observaram que as reduções da condutância estomática (gs) ao longo do
período experimental foram bastante nítidas entre os híbridos estudados. Os autores indicaram
que em estresse severo uma das primeiras respostas dos híbridos foi o fechamento estomático,
de forma a minimizar a perda de água, reduzindo também a taxa de fotossíntese líquida.
O parâmetro fisiológico teor de clorofila mede, de forma dinâmica, a proporção de
energia luminosa absorvida através da fluorescência das clorofilas. Grande parte da radiação
incidente sobre a superfície da folha não é utilizada nos processos fotoquímicos. Esta radiação
é perdida de várias formas, uma parte é refletida e outra porção é transmitida, ou seja,
atravessa a folha sem ser absorvida, enquanto que uma terceira fração da radiação é absorvida
pelos pigmentos fotossintetizantes. No entanto, nem todos esses fótons absorvidos pelos
pigmentos são utilizados nos processos fotoquímicos (TAIZ & ZEIGER, 2006).
As clorofilas são pigmentos verdes especializados na absorção luminosa contida nos
cloroplastos. Nos cloroplastos, a energia luminosa é convertida em química por meio de
unidades funcionais chamadas de fotossistemas, processo este chamado de fotossíntese (TAIZ
& ZEIGER, 2006). A alta eficiência fotossintética pode levar ao incremento de produtividade
agrícola, e essa relação está diretamente relacionada com o aproveitamento da radiação
disponível por esses pigmentos. Além das clorofilas, os cloroplastos contêm outros pigmentos
denominados de acessórios, tais como os carotenoides (carotenos e xantofilas). No entanto, os
pigmentos essenciais para conversão de energia luminosa em química armazenada são as
clorofilas a e b presentes nas folhas. A clorofila a é o pigmento utilizado para realizar a
fotoquímica (o primeiro estágio do processo fotossintético), enquanto que os demais

18
pigmentos auxiliam na absorção de luz e na transferência da energia radiante para os centros
de reação, sendo assim chamados de pigmentos acessórios (STREIT et al., 2005). Os
carotenoides são pigmentos amarelos ou alaranjados, mas normalmente sua coloração é
mascarada pelas clorofilas. Esses pigmentos situam-se nas lamelas dos cloroplastos, em
íntima associação com as clorofilas, o que permite a transferência de energia para as clorofilas
(TAIZ & ZEIGER, 2006).
Durante o déficit hídrico ocorre o fechamento estomático e também estresse luminoso
secundário pela falta de CO2, fazendo com que uma pequena proporção de energia luminosa
seja usada para fotossíntese e todas as energias disponíveis sejam utilizadas como forma de
sobrevivência do aparato fotossintético (SCHULZE et al., 2005).
MEENA-KUMARI et al. (2004) relataram que o teor de clorofila reduziu sob déficit
hídrico em 60% e 50% quando o estresse foi no florescimento e enchimento de grãos,
respectivamente. As linhagens tolerantes à seca resistiram à redução do teor de clorofila e
mostraram pouca variação, enquanto nas linhagens suscetíveis, esse caráter foi drasticamente
reduzido. Sob estresse hídrico, a diminuição do teor de clorofila foi também relatado por
diversos pesquisadores (GUTIERREZ et al., 1998; CHAVES et al., 2003).
2.3 Caracteres secundários
Devido às dificuldades em selecionar diretamente para produtividade de grãos sob
estresse hídrico, o uso de caracteres secundários tem sido sugerido para complementar a
seleção fenotípica. As características secundárias podem ser definidas como outras
características da planta, morfológicas e/ou fisiológicas, que fornecem informações adicionais
sobre as alterações da produção sob estresse hídrico (MONNEVEUX & RIBAULT, 2006).
Estas são importantes na seleção porque, usualmente, a herdabilidade do caráter “produção”
diminui enquanto a herdabilidade de algumas características secundárias permanecem altas e
a correlação genética entre produtividade e essas características aumentam nitidamente
(BOLAÑOS & EDMEADES, 1997; BÄNZIGER & LAFITTE, 1997).
Segundo MONNEVEUX & RIBAULT (2006) uma característica secundária deve ser
(1) geneticamente relacionada com a produtividade de grãos sob estresse hídrico; (2)
geneticamente variável; (3) altamente herdável; (4) facilmente mensurável e de baixo custo;
(5) não destrutiva; (6) estável dentro do período de mensuração; (7) não associada com o
decréscimo na produtividade de grãos em condições não-estressadas. Número de espigas por
planta (prolificidade), intervalo entre florescimentos feminino e masculino, stay-green, e

19
número de ramificações de pendão são características secundárias efetivamente usadas para
melhorar a produção de milho em ambientes sob estresse hídrico (BÄNZIGER et al., 2000). O
uso de caracteres secundários melhora a resposta da seleção que foca no efeito direto da seca
e evita fatores confundíveis (por exemplo, fertilidade do solo, deficiência de micronutrientes e
patógenos) que também influenciam na determinação final da produtividade de grãos
(MONNEVEUX & RIBAULT, 2006).
A seleção de populações de milho tropical sob estresse hídrico no florescimento resultou
na redução do intervalo entre os florescimentos feminino e masculino (IF) e no aumento do
número de grãos (EDMEADES et al., 2006). O menor IF assegura que um número maior de
plantas receberá pólen no momento da emergência dos estilos-estigmas, garantindo a
polinização e a fertilização (WESTGATE et al., 2004). Este caráter secundário, de fácil
observação e de herdabilidade alta, também apresenta alta relação com rendimento de grãos
sob estresse severo no florescimento (MONNEVEUX & RIBAUT, 2006). LIZASO et al.
(2003) obteve sucesso ao usar a coincidência da liberação do pólen e a emergência dos
estilos-estigmas (sincronismo), com baixa concentração de pólen típica de campos
estressados, aumentando o número de grãos (EDMEADES et al., 2006). O intervalo entre os
florescimentos feminino e masculino reduzido é indicativo da melhor partição de fotossíntatos
para a espiga, uma vez que está associado com o rápido desenvolvimento desta (GOWDA et
al., 2009) em detrimento do crescimento do pendão e colmo (DWIVEDI et al., 2010).
Reduzido IF é parcialmente consequência de um maior potencial hídrico da planta durante o
florescimento. Se ocorrer deficiência hídrica no florescimento, as plantas tolerantes podem ser
simplesmente as que florescerem mais cedo que a média e, assim escapar da seca que tende a
se intensificar com o tempo (CAMPOS et al., 2004). MEENA KUMARI et al. (2004)
relataram que, sob estresse hídrico severo, o intervalo entre os florescimentos masculino e
feminino aumentou em 3-5 dias nas linhagens tolerantes à seca e de 9-17 dias em linhagens
sensíveis de milho.
Para garantir progressos na seleção para tolerância à seca, a solução pode estar na
manipulação dos órgãos-dreno (MONNEVEUX et al., 2008), uma vez que o conjunto dos
componentes de produção constitui o dreno (BLUM, 2010). Assim, sugere-se que a seleção
para prolificidade, grãos maiores e pendão menor pode colaborar com o aumento da
produtividade sob estresse hídrico em curto prazo (MONNEVEUX et al., 2008). A
prolificidade apresenta alta herdabilidade, aumentando com a intensidade do estresse e alta
relação com produtividade de grãos sob estresse no florescimento. WESTGATE (1997)

20
enfatiza que a habilidade de genótipos produzirem maior número de espigas sob estresse
constitui uma das características mais importantes associadas com a tolerância à seca, uma
vez que a prolificidade pode aumentar eficazmente a produção de fotoassimilados para a
espiga.
Mudanças no tamanho do pendão em cultivares de milho têm sido observadas, desde os
anos 1930, como característica a ser melhorada visando o aumento na produção de grãos. A
redução no tamanho do pendão pode ser empregada uma vez que a produção de pólen
geralmente não é um fator limitante na produção de grãos em milho (GERALDI et al., 1985;
MICKELSON et al., 2002). O pendão reduz a produção de grãos tanto em função da
interceptação de luz pela copa da planta, mas principalmente, pela competição por
fotoassimilados (GERALDI et al., 1985; CHAPMAN & EDMEADES, 1999; MICKELSON
et al., 2002). Germoplasma de clima tropical parece ser mais sensível à redução nos níveis de
fotossíntese no florescimento que aqueles de clima temperado (FISCHER et al., 1987).
Assim, a competição por fotoassimilados entre pendão e espiga pode reduzir a produção de
grãos, mais pronunciadamente em material de clima tropical.
Dentre os mecanismos responsáveis pela competição de fotoassimilados está a produção
de ácido indol acético (AIA) no pendão que proporcionaria uma dominância apical por
nutrientes inibindo o desenvolvimento da espiga. Assim a espiga superior só se desenvolveria
após a redução dos níveis de AIA, causando, desta maneira, um pronunciado protandrismo
característico da planta de milho. Por sua vez, a espiga superior inibiria da mesma forma, o
desenvolvimento da espiga inferior (SOUZA Jr. et al., 1985). Para expressão da prolificidade,
faz-se necessária uma redução na quantidade de AIA, que pode ser conseguida pela
diminuição do tamanho do pendão, como forma de minimizar os efeitos da dominância apical.
Assim, mais de uma espiga pode se desenvolver simultaneamente e em menor espaço de
tempo, induzindo a coincidência da antese com a emergência de estilo-estigmas, isto é,
reduzindo o intervalo entre florescimentos feminino e masculino. O caráter número de
ramificações de pendão afeta de forma direta, portanto, dois caracteres com reconhecida
atuação na tolerância à seca.
O stay-green é uma característica importante dos programas de melhoramento à seca,
particularmente pela facilidade de visualização e seleção, principalmente em pós-
florescimento. Tem sido utilizado, por vários melhoristas na seleção de híbridos de milho
produtivos, o retardo da senescência foliar com notáveis folhas verdes escuras, além de outros
caracteres. Assim, a retenção de maior área foliar verde das folhas no estádio de enchimento

21
de grãos é um fenótipo buscado por melhoristas (BLUM, 2010) por estar associada com a
melhoria do desempenho produtivo em condições de estresse hídrico (EDMEADES et al.,
2006).
O caráter senescência retardada ou stay-green pode ser dividido em cinco tipos,
denominados A, B, C, D e E (THOMAS & SMART, 1993; THOMAS & HAWARTH, 2000).
Os tipos A e B são stay-green funcionais, que mantêm as folhas verdes e a atividade
fotossintética por mais tempo, devido ao início da senescência tardiamente (tipo A) ou ao
desenvolvimento da senescência de forma mais lenta (tipo B). Os tipos C, D e E são stay-
green não funcionais, que mantêm as folhas verdes, ou seja, há retenção da clorofila durante a
senescência, porém sem capacidade fotossintética (PARK et al., 2007; BEKAVAC et al.,
2007). O stay-green funcional é mensurado pela concentração de clorofila (BARKER et al.,
2005) e tem potencial para aumentar a produtividade das culturas (HÖRTENSTEINER,
2009). DWIVEDI et al. (2010) relataram que uma cultivar mutante de trigo duro com stay-
green funcional possui esse potencial devido à extensão do período da atividade fotossintética
durante o enchimento dos grãos. No milho, o stay-green funcional é um mecanismo
alternativo para reverter reduções de produtividade de grãos decorrentes do estresse hídrico
durante o enchimento de grãos (BARKER et al., 2005). Este caráter secundário é de fácil
observação e mensuração, herdabilidade moderadamente alta, apresentando média relação
com rendimento de grãos sob estresse, durante o enchimento de grãos (MONNEVEUX &
RIBAUT, 2006).
De acordo com CAMPOS et al. (2004) a seleção para stay-green, baseada em amplas
áreas de experimentação, tem obtido sucesso na melhoria da produtividade sob estresse
hídrico em milho temperado quando o estresse ocorre no período do florescimento até o início
do enchimento de grãos. Contudo esses ganhos são consideravelmente reduzidos quando o
período de seca ocorre entre a fase intermediária até a fase final de enchimento de grãos.
Genótipos com stay-green apresentam tolerância à seca justamente nos períodos finais de
enchimento de grãos permitindo que a planta não interrompa o fornecimento de
fotoassimilados durante esse período.
WILLMAN et al. (1987) enfatizou que correlações entre produtividade e stay-green são
definidas pela constituição genética do material e duração de seu ciclo de desenvolvimento.
Devido à relação competitiva de fotoassimilados entre a espiga e as partes da planta, o
melhoramento simultâneo de ambos os caracteres pode ser conseguido em programas de
seleção recorrente (BEKAVAC et al., 2007). Correlação positiva foi observada entre stay-

22
green e teor de umidade dos grãos indicando que o melhoramento para stay-green pode trazer
mudanças na maturidade fisiológica das plantas. Tais respostas serão indesejáveis em muitos
programas que objetivam o desenvolvimento de híbridos superiores, dentro de grupos de
maturidade (ETA-NDU & OPENSHAW, 1992; BEKAVAC et al., 2007).
Baseado nos progressos de melhoramento para tolerância à seca em milho tropical, o
ideótipo de milho tolerante é caracterizado por porte de planta baixo, área foliar reduzida
(especialmente na parte superior do colmo), menor tamanho do pendão, folhas eretas,
senescência retardada, raiz profunda (com menos ramificação lateral), melhor capacidade no
enchimento dos grãos, desenvolvimento de sementes vigorosas e bom ajuste osmótico
(RIBAUT et al., 2009).
A produtividade e caracteres secundários são incorporados nas progênies pelas
numerosas combinações das fontes de variação para alelos de vários e alguns locos e na
geração de grande número de eventos de recombinação (HALLAUER & MIRANDA FILHO,
1988; PANDEY & GARDNER, 1992).
2.4 Melhoramento genético para tolerância à seca
O melhoramento de plantas é arte e ciência de melhorar a hereditariedade das plantas em
benefício da humanidade (PHOELMAN & SLEPER, 1995). O aumento da produção de grãos
é o objetivo principal do melhoramento em milho. No entanto, em ambientes sujeitos à seca, o
progresso do melhoramento é lento devido a grande interação entre genótipos x ambientes do
caráter produção de grãos, decorrente de chuvas imprevisíveis (MONNEVEUX & RIBAULT,
2006).
A avaliação da interação genótipos por ambientes (GxA) torna-se de grande
importância, considerando que o melhor genótipo em um ambiente pode não o ser em outro.
Esse fato influencia o ganho de seleção e dificulta a recomendação de cultivares em ampla
região de cultivo. Portanto, cabe ao melhorista avaliar sua magnitude e significância e
fornecer subsídios que possibilitem adotar procedimentos para sua minimização, e, ou seu
aproveitamento (CRUZ et al., 2004). Contudo, o sucesso de uma cultivar depende do
potencial produtivo e estabilidade da mesma, diante das variações ambientais.
A principal causa das diferenças entre genótipos em suas estabilidades de produção é a
grande ocorrência da GxA. A variação do ranking e diferenças relativas dos genótipos em
uma longa extensão de ambientes é definida estatisticamente como interação GxA, definida
como a expressão genotípica diferencial através dos ambientes. Consequentemente, o

23
melhoramento para tolerância a estresses tem sido um dos responsáveis na diminuição da
interação (KANG, 1998; GEBRE, 2005). No entanto, o limitado sucesso do melhoramento
para tolerância à seca é devido, principalmente, ao fato desta característica ser controlada por
muitos genes de efeito aditivo e com forte interação existente entre genes de tolerância e
aqueles envolvidos no potencial produtivo.
A tolerância à seca é definida como a capacidade do genótipo ser mais produtivo em
condições de estresse hídrico (RIBAULT et al., 2009). O genótipo tolerante à seca possui
maior produtividade de grãos do que o genótipo suscetível em diversos ambientes sob estresse
hídrico. O genótipo ideal deve apresentar alta produtividade de grãos em ambas condições, a
não-estressante e sob estresse hídrico. Portanto, o melhoramento para tolerância à seca busca
obter o máximo potencial para produtividade de grãos e diminuir a lacuna entre o potencial
produtivo e a produtividade sob estresse (MESSMER & STAMP, 2010). A “expressão” da
tolerância depende do estádio de desenvolvimento quando ocorre o déficit hídrico e da
intensidade e duração deste. EDMEADES et al. (1995) ressaltam que a condução dos
experimentos, em ambientes com estresse hídrico, causa padrões similares de discriminação
entre genótipos adaptados e não adaptados. Assim, torna-se necessário buscar uma abordagem
mais eficiente para selecionar cultivares tolerantes à seca.
2.4.1 Seleção recorrente
Muitos métodos de melhoramento resultam em uma redução substancial de
variabilidade genética de determinado genótipo. Em contraste, os programas de seleção
recorrente têm por objetivo encontrar uma resposta à seleção por longo prazo para a
característica de interesse pelo aumento da frequência de alelos favoráveis, enquanto mantêm
simultaneamente a variabilidade genética do germoplasma para seleções posteriores
(HALLAUER, 1985).
Cada ciclo de um programa de seleção recorrente consiste de três fases: (1)
desenvolvimento de progênies (meios-irmãos, irmãos germanos, S1, S2), (2) avaliação das
progênies em ensaios com repetições que podem ser conduzidos em diferentes ambientes e
serem selecionadas com base nos caracteres de interesse e (3) recombinação das progênies
selecionadas, baseada em avaliação de ensaios, para formar uma nova população para o
próximo ciclo de seleção (HALLAUER & MIRANDA FILHO, 1988).
Os esquemas de seleção recorrente são classificados em duas categorias: seleção
recorrente intrapopulacional e interpopulacional, também denominada de seleção recorrente

24
recíproca. Na seleção recorrente intrapopulacional, o objetivo é melhorar as performances per
se das populações, enquanto na interpopulacional o objetivo é o melhoramento do cruzamento
de duas populações, ou seja, o híbrido interpopulacional (SOUZA Jr., 2001).
Existem vários métodos de seleção recorrente intrapopulacional, dentre eles, a seleção
massal, baseada no fenótipo dos indivíduos, e seleção com uso de progênies, incluindo
progênies de meios-irmãos, irmãos germanos ou de autofecundação (S1 ou S2). Os diferentes
métodos não se diferenciam apenas quanto ao tipo de progênie utilizada na seleção, mas
também quanto ao tipo de progênie utilizada na recombinação dos genótipos superiores. Desta
forma, no método de seleção com progênies de meios-irmãos, a recombinação dos genótipos
superiores pode ser feita com sementes remanescentes e com progênies S1 que exige plantas
prolíficas para obter dois tipos de progênies. O mesmo vale para a seleção com progênies de
irmãos germanos, que pode ter como unidades de recombinação, progênies de irmãos
germanos e progênies S1. Teoricamente, as famílias de irmãos germanos são mais eficientes
do que as de meios-irmãos, devido ao melhor controle parental e, consequentemente, o ganho
predito na seleção deverá ser maior.
O melhoramento interpopulacional iniciou após COMSTOCK et al. (1949) sugerirem a
seleção recorrente recíproca. Este método utiliza-se de duas populações, onde genótipos de
cada população são avaliados em cruzamentos com a outra população e, os genótipos de cada
população que apresentaram melhor capacidade de combinação com a população recíproca
são recombinados entre si. Assim procura-se melhorar a heterose do cruzamento de duas
populações. Os esquemas mais utilizados de seleção recorrente recíproca são aqueles
envolvendo progênies de meios-irmãos ou de irmãos germanos interpopulacionais para a
avaliação e progênies S1 para a recombinação (HALLAUER, 1985). No entanto, diversos
autores propuseram alterações nos procedimentos originais de seleção recorrente.
PATERNIANI (1967) propôs a seleção recorrente recíproca com base em família de
meios-irmãos. Progênies de meios-irmãos de cada uma das populações são cruzadas com
populações contrastantes, fazendo despendoamento das progênies. Para recombinação das
progênies selecionadas utilizam-se as sementes remanescentes. A vantagem do método
consiste na facilidade e simplicidade de execução, permitindo executar grande número de
progênies.
Outros esquemas envolvendo a avaliação de progênies de meios-irmãos ou de
testcrosses de meios-irmãos e recombinação de progênies de meios-irmãos foram sugeridos
por PATERNIANI & VENCOVSKY, 1977, 1978. Estes autores em 1977 sugeriram os

25
testcrosses de progênies de meios-irmãos onde as progênies de meios-irmãos de duas
populações são utilizadas, sendo que cada população serve de testador recíproco para a outra.
As sementes remanescentes das progênies de meios-irmãos de cada população, dos testcrosses
selecionados com base em ensaios com repetições são utilizadas para recombinação
(PATERNIANI & VENCOVSKY, 1977).
Comparações teóricas entre os métodos de seleção intra, interpopulacionais e
modificado foram realizadas por SOUZA Jr. (1993). Este autor propôs um método alternativo
de seleção recorrente usando a população de baixa produção (B) para avaliar outra população
com performance superior (A) e ela mesma (B), resultando no aumento médio das duas
populações per se e também da heterose devido a população (A) ser melhorada pela
capacidade de combinação com a população (B). Portanto, o aumento da heterose ocorre na
menor extensão do que na seleção recorrente recíproca tradicional, porém duas populações
são melhoradas com ganho genético adequado (ARIAS & SOUZA Jr., 1998). Como o
aumento da média do híbrido interpopulacional é função do melhoramento de populações per
se e da heterose, o método de seleção recorrente modificado denominado seleção em
testcrosses (interpopulacional) e meios-irmãos (intrapopulacional) seja tão eficiente quanto o
método tradicional.
Nos programas de seleção recorrente têm-se desenvolvido novas fontes de tolerância,
com o uso de progênies parcialmente endogâmicas (S1 e S2) e populações F2 (FALKE, 2007)
como unidade de seleção, principalmente, quando o estresse hídrico atinge os estádios de pré-
florescimento, florescimento e enchimento de grãos. Assim, a diversidade genética é um fator
essencial para o processo de seleção e o milho tem servido como cultura exemplar para o
aproveitamento dessa variabilidade.
No trabalho desenvolvido por BOLAÑOS & CÓRDOVA (1997) foram obtidos
variedades sintéticas de alta produtividade de grãos, utilizando-se a população tolerante ao
estresse hídrico Tuxpeño Séquia C6 (TS6) x BS19, através de progênies de meios-irmãos com
recombinação de plantas S2, seguidos de 4 ciclos de seleção. EDMEADES et al. (1997b)
avaliaram a produtividade de grãos de três populações de milho (La Posta Sequía, Pool 26
Sequía e Tuxpeño Sequía) selecionadas para tolerância à seca, através de seleção recorrente
baseada em progênies S1. A produtividade variou de 1,0 a 4,5 kg ha-1
sob estresse hídrico e de
5,8 a 10,4 kg ha-1
em boas condições hídricas, respectivamente. Os ganhos com a seleção em
ambientes com estresse hídrico, variaram de 3,8 a 12,6% por ciclo, enquanto para ambientes
com boas condições hídricas, variaram de 0,5 a 2,3% por ciclo. BADU-APRADU et al.

26
(2013) obtiveram uma taxa média de aumento de produtividade de grãos de 13,5 kg ha-1
ano-1
,
correspondendo a 1,1% de ganho genético anual em condições de seca, em cultivares de
milho precoce.
GUIMARÃES (2013) avaliou a massa de grãos e caracteres secundários em progênies
interpopulacionais de milho, de ciclo 0 de seleção recorrente recíproca, observando alta
variabilidade genética para esses caracteres. Destacaram-se a precocidade e os menores
números de ramificações do pendão dessas progênies e alto ganho com a seleção entre
progênies para prolificidade e intervalo entre florescimentos.
GOMES E GAMA et al. (2004) estudaram progênies S2 oriundas de cada uma das duas
populações sintéticas de milho, Syndent e Synflint, com o objetivo de obter sintéticos com
alto potencial produtivo e com comportamento agronômico sob condições de seca da região
semi-árida do Brasil. Os autores obtiveram ganho de seleção sob estresse hídrico
considerando os caracteres secundários devido a variabilidade desses germoplasmas para
tolerância à seca. Eles indicaram que a utilização dessas progênies para a seleção
interpopulacional visando obter híbridos de sintéticos ou top cross é uma opção viável porque
requer menos tempo na obtenção de uma cultivar e menor custo de sementes quando
comparado com os híbridos de linhagens.
SRINIVASAN et al. (1997) utilizaram o método de seleção recorrente modificado para
tolerância à seca e baixo teor de nitrogênio nas populações 901 e 902 (milho branco precoce e
semi-dentado) obtendo progênies interpopulacionais com boa capacidade combinatória para
serem utilizadas como testadores e também na obtenção de sintéticos. A principal estratégia
deste programa de melhoramento foi a piramidação de genes que conferem tolerância à seca
(menor intervalo entre florescimentos, produtividade e outros caracteres secundários) e a
eficiência no uso de nitrogênio nos diferentes sintéticos formados e verificação de sua
performance em combinações.
MONNEVEUX et al. (2006) demonstraram que o uso da seleção recorrente, em
populações de milho tropical sob condições de estresse hídrico, apresentou ganhos
significativos para produtividade de grãos associados ao aumento de prolificidade e número
de grãos por espiga; reduções do intervalo entre florescimentos e taxa de abortos durante o
enchimento de grãos.

27
2.4.2 O fator latente
Alguns tipos de milho sob a influência da seleção natural em várias partes do mundo
têm desenvolvido mecanismos de tolerância à seca. As variedades de milho mexicanas
apresentam diferentes mecanismos de tolerância, que têm sido relatados por pesquisadores
desde 1960 pela Fundação Rockefeller (MIRANDA, 1972). Dentre essas variedades, cita-se o
Cateto Criolo cultivado no Estado de Oaxaca no México, que possui alto nível de tolerância à
seca e caracteriza-se por ter longo ciclo vegetativo, com lento desenvolvimento até as chuvas
iniciarem, quando há uma rápida resposta em termos de desenvolvimento e recuperação. A
variedade Michoacán 21 possui o mecanismo de tolerância à seca denominado “latente”, que
consiste no prolongamento do estádio vegetativo, em condições de estresse hídrico, retomada
rápida do crescimento normal e do estádio reprodutivo, assim que se inicia o período das
chuvas (HAYANO-KANASHIRO et al., 2009). PALACIOS DE LA ROSA (1959) descreveu
o Michoacán 21, como responsável por ter uma resposta distinta à seca e ao frio. Segundo
MUÑOZ (1975), citado por FISHER et al. (1982), a coleção de Michoacán mantida em
condições de seca, apresentou notável recuperação após reidratação, sendo mais resistente à
murcha permanente no estádio de plântulas e reduzida transpiração, devido ao fechamento de
estômatos. Segundo LARQUE-SAAVEDRA & WAIN (1974), esta resposta pode ser devido
a altos níveis de ácido abscísico.
HAYANO-KANASHIRO et al. (2009) estudaram dois genótipos mexicanos de milho
tolerantes à seca (Cateto Criolo e Michoácan 21 do CIMMYT), quanto às diferenças nas
respostas fisiológicas tais como fotossíntese, condutância estomática e potencial hídrico em
comparação ao genótipo suscetível (85-2). Os resultados confirmaram que o Michoácan 21 foi
associado ao rápido fechamento dos estômatos em comparação ao suscetível, e que a
hipersensibilidade dos estômatos foi uma característica em comum nas linhagens tolerantes à
seca. A estratégia do Michoácan em diminuir a taxa fotossintética pode ser vantajosa quando
ocorre curto período de déficit hídrico. Enquanto que o Cateto Criolo apresentou reduzida
taxa fotossintética e maiores valores de potencial hídrico, permitindo a planta sobreviver por
longos períodos de déficit hídrico. Após a reidratação, ambos os genótipos Cateto Criolo e
Michoácan 21 demonstraram uma rápida recuperação pelo aumento da fotossíntese quando
comparado com o genótipo suscetível, sugerindo que genótipos tolerantes à seca possuem
mecanismos distintos após déficit hídrico, visto que o suscetível não apresentou esse
comportamento.

28
Porém o caráter latente forneceu dificuldade para transferir alta produtividade,
característica agronomicamente desejável, particularmente nos trópicos. Entretanto,
pesquisadores dos Estados Unidos obtiveram sucesso usando o Michoacán 21 como fonte de
genes para o melhoramento de híbridos tolerantes para a região do Corn Belt nos Estados
Unidos (CASTLEBERRY & LERETTE, 1979; FISHER et al., 1982). MONNEVEUX et al.
(2006) relataram que o Michoácan 21 foi utilizado como fonte de fenótipo latente para o
desenvolvimento da população de milho tropical DTP1 (Drought Tolerant Population) pelo
Cimmyt. O caráter latente não possui herança simples e o desenvolvimento de híbridos
tolerantes à seca requer seleção de linhagens sob condições ambientais controladas para
produtividade e outras características associadas com a tolerância à seca (FISHER et al.,
1982).
MIRANDA (1972) estudou a herança do caráter latente em milho nas populações IAC
IAC Maya Latente e IAC 1 latente, em relação à tolerância a geada e calor. Esse estudo
indicou que o efeito fisiológico exibido no milho latente (Michoácan 21 comp.1-104) pode ser
devido a atuação de um ou dois locus. Indicaram que nas populações provenientes de
cruzamentos com milho latente e selecionados por meio de geada, as sementes revelaram
maior tolerância quando testadas ao calor do que as populações originais.
Outros estudos relacionados com o fator latente foram realizados por BRUNINI et al.
(1981, 1982 a, b) e CAMARGO et al. (1982) citado por MIRANDA & MIRANDA (1993),
onde a comparação de caracteres fisiológicos do IAC IAC Maya Latente e IAC Phoenyx
latente com cultivares de milho comum, relacionou o latente por possuir um mecanismo de
abertura e fechamento de estômatos mais eficiente; os estômatos abriram mais cedo, com
menos luz de manhã, e fecharam antes, durante a parte mais quente do dia como verificado
pelo aumento da resistência estomática. BRUNINI et al. (1981) demonstrou o menor consumo
de água do IAC Maya Latente em déficit hídrico, verificam que seu consumo sob condições
de excesso de chuva (mais de 5 mm/dia) foi superior, e com falta de água (menos de 5
mm/dia) seu consumo baixou até 57% em relação às cultivares normais.
SAWAZAKI et al. (1984) estudaram as cultivares IAC IAC Maya Latente e IAC 8222
quanto ao comportamento destas em relação às cultivares comerciais e o efeito da bordadura
na parcela para avaliação de cultivares com diferenças de porte e consumo de água. Os
autores obtiveram maior peso de grãos para o IAC 8222 para a maioria dos locais avaliados,
destacando-se os locais de Arandu e Cesário Lange, que apresentaram o maior efeito de seca

29
na produção. Atribuíram este fato aos fatores do complexo latente da variedade IAC Maya
Latente (MIRANDA et al., 1984) que é o progenitor fêmea deste híbrido (IAC 8222).
2.5 Estimativas de parâmetros genéticos
A seleção de plantas sob déficit hídrico requer a atuação do melhoramento de plantas no
entendimento de mecanismos genéticos. Estudos mostram que há grande variação para
tolerância à seca em várias espécies cultivadas como o trigo, sorgo e milheto. Por causa da
ampla variabilidade genética encontrada para seca, as estimativas de parâmetros genéticos
ajudam a desenvolver cultivares tolerantes à seca, permitindo predizer o efeito da seleção e
planejar os procedimentos de melhoramento, já que estes têm suas origens nos quadrados
médios da análise de variância, de dados experimentais e suas esperanças matemáticas
(VENCOVSKY & BARRIGA, 1992).
A variância genotípica presente na herdabilidade leva o melhorista a uma melhor
compreensão dos padrões de herança de vários caracteres. A herdabilidade pode ser de
sentido amplo ou restrito e se refere o quanto dos valores genéticos ou aditivos são atribuídos
à variância fenotípica, respectivamente (FALCONER, 1989). A relação das variâncias
genética e fenotípica ( 22
FG ) é denominada de herdabilidade em sentido amplo. Essa se
refere à extensão dos fenótipos dos indivíduos que são determinados pelo genótipo. Por outro
lado, a relação das variâncias aditivas e fenotípica ( 22
FA ) é denominada herdabilidade em
sentido restrito, sendo a proporção da variabilidade observada em razão dos efeitos aditivos
dos genes, que é a parte herdável da variância genética, ou seja, a única aproveitável na
seleção. Essa expressa à extensão dos fenótipos que são determinados pelos genes
transmitidos aos seus pais. Também representa a magnitude da variância genética da
população, que é a principal responsável por mudanças na composição genética da população
por meios de seleção (FALCONER, 1989).
A estimativa da herdabilidade em sentido amplo é baseada na informação de vários
componentes das variâncias genética e ambiental do germoplasma (DUDLEY & MOLL,
1969). Contudo, o tipo de ação gênica envolvendo a expressão de um caráter tem papel
significativo na determinação da herdabilidade. Os caracteres que são controlados por muitos
genes de forma aditiva geralmente têm maiores valores de herdabilidade do que os
governados por genes de efeito não-aditivos (FALCONER, 1989).

30
Em condições de estresse hídrico, os valores de herdabilidade são baixos em relação aos
encontrados nas condições favoráveis (BLUM, 1988). Neste caso, a estimativa da
herdabilidade permite identificar ambientes favoráveis para seleção direta ou indireta do
caráter, em condições irrigadas e de déficit hídrico e a correlação genética existente entre
esses ambientes. Contudo, pesquisas revelam declínio da herdabilidade para produtividade de
grãos em condições de déficit hídrico, devido a baixa variância genética e a alta variação
ambiental (BLUM, 2010). Um estudo de feijão realizado por FRAHM et al. (2004),
concluíram que o estresse hídrico e baixa produções contribuíram para o alto coeficiente de
variação (CV).
A magnitude da herdabilidade sob estresse foi associado com os estudos de distância
genética entre genitores quando em cruzamento (CECCARELLI et al., 1998), indicando que a
utilização de raças de cevada adaptadas a seca, asseguram a variabilidade genética para esse
fator, ao invés de apenas potencial produtivo. Similarmente, VENUPRASAD et al. (2007)
encontraram para seleção de produtividade de grãos em arroz, avaliada sob estresse hídrico
severo, altos níveis de tolerância a seca derivadas de genes doadores das populações com
caracteres de adaptação à seca. Portanto, as estruturas genética e fisiológica da população em
termos de tolerância a estresses é crucial na determinação da herdabilidade para produtividade
de grãos e da eficiência de seleção sob déficit hídrico (BLUM, 2010).
É sugerido que a seleção em ambientes ótimos de irrigação não é efetiva na
identificação de genótipos superiores para condições de seca, devido a baixa variância
genética e herdabilidade da produtividade de grãos (BÉTRAN et al., 2003). De acordo com
BLUM (2010), duas considerações podem melhorar a herdabilidade e a eficiência da seleção
para produtividade sob déficit hídrico e/ou compensar a baixa herdabilidade da produção sob
estresse: (1) a condução do campo experimental sob déficit hídrico deve apresentar
homogeneidade das condições de umidade do solo e assim reduzir o erro experimental e (2)
utilizar populações que contenham variabilidade genética para produtividade de grãos,
tolerância a seca e ideotipo de plantas que contenham os caracteres secundários.
No trabalho de ZIMOYO & BERNARDO (2013) a herdabilidade em sentido amplo
para produtividade foi 0,60 no experimento irrigado e 0,37 em déficit hídrico. A diminuição
da herdabilidade para produtividade sob déficit hídrico corresponde a diminuição da média da
produção de grãos, uma relação que tem sido verificado em vários trabalhos (BÄNZIGER &
LAFITTE, 1997; BOLAÑOS & EDMEADES, 1996). A alta herdabilidade e variância

31
genética para intervalo entre florescimentos e stay-green, sob déficit hídrico, se destacam para
necessidade de seleção desses caracteres secundários em ambientes sujeitos à seca.
3 MATERIAL E MÉTODOS
3.1 Descrição do material
Foram utilizadas duas populações de milho: uma população com potencial para
tolerância à seca, oriunda do intercruzamento de híbridos comerciais tolerantes, sintetizada no
programa de melhoramento de milho do Instituto Agronômico (IAC), denominada População
Tolerante 1 (Pop.Tol.1) e a variedade de milho IAC IAC Maya Latente. A População
Tolerante 1 é uma população sintética de meios-irmãos, que apresenta características de
menor altura de plantas, stay-green acentuado e alta prolificidade (GUIMARÃES, 2013).
A variedade IAC Maya foi obtida pela combinação das seguintes linhagens e variedades
conforme MIRANDA (1972), citado por SAWAZAKI (1980): (a) 14 linhagens originárias de
Asteca que é um composto de coleções de San Luis Potosi; ( b) Ip 701-1, linhagem obtida por
autofecundação da variedade Tuxpan, originária do Texas, selecionada do cruzamento
Tuxpeño branco com Creole Yellow Flint, sendo precoce, mais baixa e de endosperma
amarelo; (c) Tx 303, linhagem comercial tardia do Texas, obtida da variedade Yellow
Surecropper; (d) Llera III branca; (e) Vera Cruz 226 e 227, variedades de Tuxpeño branco,
típicas, muitos tardias, altas e de espigas brancas. Estes materiais foram combinados por
cruzamentos manuais em cadeia, planta a planta, seguindo-se de uma geração de polinização
livre em campo isolado e em 1963/64 essa população foi sintetizada e melhorada pelo método
de seleção entre e dentro de progênies de meios-irmãos (SAWAZAKI, 1980).
O IAC Maya Latente é uma variedade sintética de grãos dentados e amarelos, derivada
do cruzamento entre o IAC Maya com o Michoacán 21 Composto 1 - 104 “latente”, que é
fonte de tolerância à seca e geada (PALACIOS DE LA ROSA, 1959; FISCHER et al., 1982)
e de calor e geada (MIRANDA et al., 1982). No desenvolvimento desta cultivar na década de
70 foram realizados: cruzamento entre IAC Maya x Michoacán 21 (Composto 1-104) e duas
autofecundações para obter famílias puras e segregantes. Estas famílias foram submetidas a
ensaios em regiões com incidência de geada. As sementes remanescentes das plantas mais
tolerantes a geada foram semeadas, recombinadas e comparadas com o material adaptado para
tolerância ao calor em teste realizado com sementes. O IAC Maya foi retrocruzado mais três
vezes e autofecundado duas vezes entre cada retrocruzamento. Muitas plântulas foram

32
avaliadas em safras normais e em ocorrência de geada, que confirmou a tolerância ao calor e à
geada desta cultivar (MIRANDA et al., 1982).
3.2 Métodos
3.2.1 Seleção recorrente de progênies interpopulacionais em campo
3.2.1.1 Obtenção das progênies interpopulacionais (ciclo 0)
Em campo isolado, foram semeadas a variedade IAC IAC Maya Latente nas linhas
masculinas e a População Tolerante 1 nas femininas, que foram despendoadas no estádio de
pendoamento (VT). A proporção utilizada foi de 2 ♂: 3♀ (Figura 1), ambas contendo 20 m de
comprimento e 0,80 cm de espaçamento entre as linhas. A semeadura das fileiras masculinas
foi no dia 20 de julho de 2011 e femininas no dia 30 de julho de 2011. Os demais tratos
culturais foram os normalmente recomendados para a cultura.
Na colheita, foram tomadas aleatoriamente espigas de plantas das linhas femininas, que
foram colhidas e debulhadas separadamente. Dessa forma, foram obtidas 196 progênies
interpopulacionais de milho, para avaliação na safrinha de 2012. Uma quantidade suficiente
de sementes de todas as progênies foi guardada para posterior recombinação das progênies
interpopulacionais selecionadas, visando à obtenção de uma nova população tolerante à seca.
Figura 1. Obtenção de progênies interpopulacionais de milho obtida do intercruzamento da
Pop.Tol.1 e IAC IAC Maya Latente, na safra de verão de 2011/12. Campinas, SP.

33
3.2.1.2 Avaliação de progênies interpopulacionais em campo
As 196 progênies interpopulacionais obtidas do cruzamento entre Pop.Tol.1 e IAC IAC
Maya Latente e 4 testemunhas foram avaliadas na safrinha de 2012, em dois locais do Estado
de São Paulo: no Centro Experimental Central do IAC, em Campinas (latitude 22°54'S,
longitude 47°03'W e altitude de 600m) e na Apta Regional do Nordeste Paulista, em Mococa
(latitude 21°28’S, longitude 47°01’W e altitude de 665 m). O delineamento experimental
utilizado foi o de látice quadrado parcialmente balanceado, 14 x 14, com duas repetições. As
parcelas foram constituídas de uma linha de 5 m, espaçadas de 0,90 m, com densidade de
semeadura de 24 plantas por linha após o desbaste. Como testemunhas, foram utilizadas a
População Tolerante 1, uma população sensível à seca (obtida do intercruzamento de um
sintético de híbridos comerciais sensíveis, sendo estes identificados pelos ensaios regionais de
safrinha no Estado de São Paulo) (GUIMARÃES, 2013), o híbrido DAS2B707 (tolerante à
seca) (GUIMARÃES et al., 2014) e a variedade IAC IAC Maya Latente. O DAS2B707 é um
híbrido simples, precoce, de elevada produtividade de grãos, alta prolificidade e stay-green
acentuado (DOW SEMENTES, 2014/2015).
Os experimentos de Campinas e Mococa foram semeados nos dias 03 e 29 março de
2012, respectivamente. Na adubação de plantio foram utilizados 350 kg ha-1
da fórmula
8:28:16 de N:P:K, em Campinas e 400 kg ha-1
de 4:14:8, em Mococa. A adubação de
cobertura foi realizada com a aplicação de 200 kg ha-1
de sulfato de amônio, parcelada nos
estádios fenológicos de 4 e 8 folhas completamente expandidas, nos dois locais. Durante o
cultivo, foram realizadas práticas recomendadas para condução da cultura, tais como o
controle de plantas daninhas e da lagarta-do-cartucho (Spodoptera frugiperda).
Os dados climáticos referentes aos meses de março a agosto de 2012 em Campinas e
Mococa encontram-se nas Figuras 2 e 3, sendo para precipitação pluvial total dos
experimentos de Campinas e Mococa, foram de 676 e 421 mm, respectivamente (CIIAGRO,
2013). Em Campinas, o florescimento masculino (FM) ocorreu durante o período de 7 a 14 de
maio 2012 e, em Mococa, no período de 2 a 9 de junho de 2012. Os valores médios de
temperatura máxima e mínima, durante o período do experimento em Campinas, foram de
26,2°C e 14,5°C, e em Mococa, foram de 27,3°C e 15°C, respectivamente. Nesse período da
experimentação ocorreu estresse hídrico em Campinas e Mococa, principalmente, nos
estádios de florescimento e enchimento de grãos (Figuras 2 e 3).

34
Figura 2. Temperaturas máxima e mínima e precipitação pluvial média da localidade
de Campinas, no período de 03/03/2012 à 03/08/2012. (Fonte: CIIAGRO, 2013).
Figura 3. Temperaturas máxima e mínima e precipitação pluvial média da
localidade de Mococa no período de 29/03/2012 à 29/08/2012. (Fonte: CIIAGRO,
2013).
3.2.1.3 Avaliação dos caracteres biométricos e secundários em campo
(a) Florescimento masculino (FM) e feminino (FF): obtidos pelo número de dias da
semeadura até que 50% das plantas de cada parcela estivessem com antese e estilos-
estigmas visíveis, respectivamente; O intervalo entre florescimentos feminino e
masculino (IF) correspondeu a diferença, em dias, entre o florescimento feminino e o
florescimento masculino. Os dados de IF foram transformados em x para a realização
das análises de variância.
Tem
per
atura
(°C
)
Chuva (m
m)
Tem
per
atura
(°C
)
Chuva (m
m)
FM
FM

35
(b) Altura da planta (AP) e da espiga (AE): obtidas pela medida tomada do nível do
solo até a inserção da folha bandeira e da espiga principal em cinco plantas da parcela,
respectivamente, em cm;
(c) porcentagem de plantas acamadas e quebradas (A+Q): obtida através da
contagem do número de plantas cujo ângulo com o solo seja <45° e das plantas
quebradas, respectivamente. Os dados de (A+Q) foram transformados em arcoseno da
100)5,0( x para realizar as análises de variância;
(d) Senescência retardada de folhas e colmo ou “stay-green” (SG): avaliado cerca de
120 dias após a semeadura após a maturidade fisiológica dos grãos, em cinco plantas
competitivas na parcela, a partir de uma escala de notas variando de 1 a 5, onde as notas
seguiram o seguinte padrão: 1- plantas com as folhas acima da primeira espiga e pelo
menos duas folhas abaixo da primeira espiga verdes; 2- plantas com todas as folhas
acima da primeira espiga verdes; 3- plantas com até duas folhas acima da primeira
espiga secas e as demais verdes; 4- plantas com até duas folhas verdes; 5- plantas com
todas as folhas secas (COSTA et al., 2008). Os dados de SG foram transformados em
x para a realização das análises de variância.
(e) Número de ramificações do pendão (NRP): número de ramificações primárias de
cinco plantas na parcela. Os dados de NRP foram transformados em x para a
realização das análises de variância;
(f) Prolificidade (PROL): medida pela relação do número de espigas com número de
plantas da parcela;
(g) Massa de grãos (MG): corresponde ao peso de grãos da parcela, em kg, resultantes
da debulha total de espigas de uma parcela, com auxílio de balança eletrônica. A MG
foi corrigida para umidade de 14% e estande ideal (24 plantas/ parcela). Para as análises
genético-estatísticas a MG foi convertida em g planta-1
, dividindo-se o valor médio de
cada parcela pelo estande ideal do respectivo experimento. Para correção do estande
utilizou-se o método da covariância descrito por VENCOSVSKY & BARRIGA (1992).
(h) Componentes de produção: comprimento de espiga (CE), comprimento médio, em
cm, de 5 espigas sem palha obtidas aleatoriamente de cada parcela, medida com auxílio
de régua; número de fileiras de grãos na espiga (NFE), contagem do número médio de
fileiras de grãos de 5 espigas obtidas aleatoriamente de cada parcela.

36
3.2.2 Análises Biométricas
3.2.2.1 Análises de variâncias e estimativas de parâmetros genéticos
A análise do látice utilizando as médias ajustadas dos tratamentos da análise com
recuperação da informação interblocos, e tendo como quadrado médio do resíduo a
variância efetiva média desta mesma análise com recuperação da informação
interblocos (tabelas 2) foi realizada de acordo com a metodologia COCHRAN & COX
(1966).
Para as análises de variância o efeito de progênies considerado aleatório e adotou-
se o seguinte modelo estatístico, eq. (2):
ijkjkjiijk ebrpY )( (2)
em que ijkY é o valor observado da i ésima progênie, na j ésima repetição, no k ésimo
bloco; é o efeito da média geral do experimento; ip é o efeito aleatório da progênie
interpopulacional i; jr é o efeito aleatório da repetição j;
)( jkb é o efeito aleatório do
bloco k dentro da repetição j, sendo k = 1,..., 16;ijke é o erro experimental associado a
observação ijkY .
Tabela 2. Esquema da análise de variância individual e esperanças dos quadrados
médios da análise do látice quadrado parcialmente balanceado, utilizando as médias
ajustadas de tratamentos e a variância efetiva do látice.
Fontes de Variação GL QM E(QM)
Repetições (r-1) 4Q 22
)2,1( rE p
Blocos/rep.(aj.) r(k-1) 3Q 2
/
2
)2,1( rbE p
Progênies (aj.) (k2-1) 2Q
)2,1(
22
)2,1( pE r
Erro intrabloco (k-1)(rk-k-1) 1Q 2
)12(E
Fonte: SILVA et al. (1999)
Nesse caso, o ( *
1Q ) é a variância efetiva média, da análise do látice com
recuperação da informação interblocos ( 'rV ), que foi dada por, eq.(3):
r
b
rb
r VV
VV
kr
rVQ .
)(.
)1)(1(1'*
1
(3)

37
em que r é o número de repetições; k é o número de parcelas em cada bloco; bV é o
quadrado médio de blocos/repetições (ajustado), obtido na análise intrabloco; e rV é o
quadrado médio do resíduo intrablocos.
A eficiência relativa (%)Ef dos experimentos em látice com recuperação da
informação de interblocos, em relação aos blocos casualizados completos, foi calculada
pela fórmula, eq. (4):
100.'
(%)rV
QMREf (4)
em que 'rV é a variância efetiva média da análise do látice com recuperação de
informação interblocos, e QMR é o quadrado médio do resíduo da análise do látice
como blocos casualizados completos.
Com as médias ajustadas e os quadrados médios dos erros efetivos obtidos das
análises individuais (FEDERER, 1955), foram realizadas análises de variâncias
conjuntas dos ambientes, utilizando o seguinte modelo, eq. (5):
ililliil epaapY )( (5)
em que ilY é a média ajustada da i ésima progênie, no l ésimo ambiente; é o efeito da
média geral do experimento; ip é o efeito aleatório da progênie interpopulacional i,
sendo i = 1,2,...,196; la é o efeito fixo do ambiente l, sendo l = 2; ilpa)( é o efeito il
ésima interação progênies por ambientes; ile é o erro efetivo médio associado à
observação ilY .
De posse das esperanças de quadrados médios, apresentadas na Tabela 2, é
possível estimar os componentes de variância da análise individual. Foram utilizados os
seguintes estimadores descritos por SILVA et al. (1999), eq. (6, 7 8):
2
)2,1(ˆ
p , estimador da variância genética entre progênies interpopulacionais, em
nível de médias dos tratamentos.
r
QMQM EP
p
)(2
)2,,1(
(6)

38
2
)2,1(ˆ
E , estimador da variância ambiental, em nível de médias dos tratamentos.
r
QM E
E 2 (7)
2ˆF , estimador da variância fenotípica entre progênies, em nível de médias dos
tratamentos.
r
QM P
F 2 (8)
2ˆA , estimador da variância genética aditiva.
2
)2,1(
2
)2,1(4
1pA (9)
2
)2,1(xh , herdabilidade em sentido amplo em nível de média de progênies
interpopulacionais.
r
hE
p
p
x 2
)2,1(
)2,1(
2
)2,1(
2
2
)2,1(
(10)
em que QMP e QME referem-se, respectivamente, aos quadrados médios das fontes de
variação progênies e do erro efetivo médio; r refere-se ao número de repetições.
O erro da estimativa da variância genética entre progênies foi obtido seguindo a
metodologia proposta por VENCOVSKY & BARRIGA (1992), eq. (11):
22
2).(
2
2
2
1
2
1
22)2,1(
2
n
QM
n
QM
rkVar p (11)
em que r é o número de repetições; k é o número de plantas por parcela; 1QM e
2QM referem-se aos quadrados médios relativos às fontes de variação de progênies e do
resíduo, respectivamente; 1n e 2n referem-se ao número de graus de liberdade de 1QM e
2QM , respectivamente.
O erro da estimativa da herdabilidade em sentido amplo em nível de média de
progênies (2
)2,1(xh ) foi estimado conforme proposto por VENCOVSKY & BARRIGA
(1992), eq. (12):
2
1
2
1)1(2).(
21
22
)2,1(
2
)2,1(nn
hhVar xx (12)

39
Os coeficientes de variação genética ( gVC ˆ ), experimental (EVC ˆ ), o índice de
seleção (b) e a predição de ganho ()2,1(
ˆSG ) foram obtidos de acordo com
VENCOSVSKY (1987), eq. (13, 14, 15, 16, 17):
100.(%)ˆ )2,1(
XVC
p
g
(13)
100.(%)ˆ )2,1(
XVC
E
E
(14)
E
g
CV
CVb (15)
)2,1(
)2,1
2
(
2,1
41.ˆ
F
A
ksG
(16)
100.ˆ
(%)ˆ 2,1
2,1X
GsG
S (17)
em que )2,1(
ˆp ,
)2,1(ˆ
F e )2,1(
ˆE referem-se, respectivamente, aos desvios padrão genético
de progênies, fenotípica e erro efetivo médio; )2,1
2
(A é a variância aditiva da progênie
interpopulacional 1,2 (Pop.Tol.1 e IAC Maya Latente); k é função da intensidade de
seleção aplicada entre progênies interpopulacionais (intensidade de seleção
estandardizada); X é a média do caráter; 1,2 referem-se, respectivamente, a Pop.Tol.1 e
o IAC IAC Maya Latente.
Para a realização das análises genético-estatística foi utilizado o programa
computacional Genes versão Windows (CRUZ, 2006).
3.2.2.2 Estimativa de heterose em relação à média dos pais
A heterose foi obtida pela média das progênies interpopulacionais em relação a
média dos pais (Pop.Tol.1 e IAC Maya Latente) para os caracteres secundários (AP, IF,
NRP e SG) e massa de grãos (MG), conforme eq.(18, 19):
PMInterPH . (18)
100..
%PM
PMInterPH
(19)

40
em que H é a heterose média das progênies interpopulacionais; %H é a heterose
relativa da progênie interpopulacional; InterP. é a média das progênies
interpopulacionais e PM é a média dos pais (Pop.Tol.1 e IAC Maya Latente).
3.2.3 Avaliação em casa de vegetação das progênies interpopulacionais
selecionadas
Com base nas informações dos experimentos de campo, foram selecionadas oito
progênies tolerantes à seca. Na seleção destas progênies foram considerados os
caracteres: massa de grãos, altura de planta, intervalo entre os florescimentos e stay-
green, principalmente, no experimento de Mococa. As progênies selecionadas,
denominadas: PI(1,2)-49, PI(1,2)-52, PI(1,2)-59, PI(1,2)-72, PI(1,2)-100, PI(1,2)-109, PI(1,2)-117
e PI(1,2)-147, foram avaliadas quanto aos parâmetros fisiológicos e biométricos,
juntamente com as testemunhas: DAS2B707 (híbrido tolerante à seca), IAC IAC Maya
Latente e uma progênie sensível à seca (PI(1,2)-141, selecionada pelos experimentos de
campo).
O experimento foi instalado no dia 2 de setembro de 2013, no Centro
Experimental do Instituto Agronômico, Campinas (Figura 4), em casa de vegetação,
com semeadura direta no solo. O delineamento experimental foi o de blocos ao acaso,
com parcelas subdivididas e duas repetições. As parcelas foram constituídas de dois
tratamentos de estresse hídrico submetidos em estádio de desenvolvimento da cultura:
(1) não-estressada, em que as plantas foram mantidas sob irrigação de acordo com o
recomendável e (2) sob estresse hídrico severo, em que a irrigação foi retirada quando
50% das plantas apresentavam antese visíveis (VT). As subparcelas foram formadas
pelas 8 progênies e 3 testemunhas. Cada parcela foi constituída de uma linha de 3,5m,
com espaçamento de 0,50 cm entre plantas e de 1,20 m entre linhas, totalizando 14
plantas na parcela.

41
Figura 4. Disposição do experimento realizado em casa de vegetação no Centro
Experimental Central do Instituto Agronômico (Campinas, 2013).
Os tratos culturais foram realizados de acordo com a necessidade da cultura, tais
como o controle de plantas daninha por meio de capina manual e o controle das
lagartas: lagarta-do-cartucho (Spodoptera frugiperda) e lagarta-rosca (Agrotis ipsilon).
A irrigação foi fornecida por um sistema de gotejamento com vazão de 3,75 L h-1
. O
potencial matricial da água no solo foi medido por tensiômetros de coluna de mercúrio,
instalados a 45 cm de profundidade de cada parcela. Foram fornecidos de três a quatro
irrigações semanais com duração de 10 minutos quando as plantas se encontravam no
estádio vegetativo e de vinte minutos no estádio de florescimento e enchimento de
grãos. O fornecimento de água no tratamento irrigado foi realizado no momento em que
os tensiômetros indicaram potencial matricial do solo próximo da capacidade de campo
(-0,10 MPa) (BERNARDO et al., 2006). O período de déficit hídrico, temperatura
máxima e mínima e da umidade relativa dentro da casa de vegetação e potenciais
matriciais no solo estão apresentadas na tabela 3.

42
Tabela 3. Período de déficit hídrico, temperaturas média de máxima e mínima e
umidade relativa dentro da casa de vegetação, potencial matricial do solo e amplitude de
variação (AV) durante o período de seca no estádio fenológico de florescimento (VT)
até o estádio reprodutivo (R4/R5).
Casa de vegetação Estádio fenológico de
florescimento (VT)
Período de déficit hídrico (dias) 02/11/2012 a 20/12/2012
(48 dias)
Temperatura máxima (°C)
(AV)
33°C
(27,2;39,1)
Temperatura mínima (°C)
(AV)
21°C
(17,7;23,9)
Umidade relativa (%)
(AV)
58%
(42;73)
Potencial matricial do solo (MPa)
(AV)
-0,90 MPa
(-0,80;-1,00)
3.2.3.1 Avaliação fisiológica em casa de vegetação
Realizaram-se avaliações de parâmetros fisiológicos na folha da espiga principal
(critério de avaliação), no dia de máximo déficit hídrico (Figuras 5a e b). O dia de
máximo déficit hídrico foi determinado pelo aspecto visual das plantas, ou seja, quando
essas plantas apresentavam sintomas de murcha acentuada no início do dia. Os
parâmetros fisiológicos avaliados foram:
(a) Clorofila total (Chl): as avaliações do teor de clorofila a e b foram realizadas com o
equipamento SPAD-502 Plus – Konica Minolta, sendo obtidas as concentrações das
clorofilas a e b totais, em cinco plantas por repetição.
(b) Condutância estomática (gs): foi utilizado o porômetro (Type AP4 – Delta T
Devices) em estado de equilíbrio dinâmico, em 2 avaliações (gs(1)
- 9:00 às 10:30h e gs(2)
- 11:00 às 12:30h). Para esta avaliação escolheu-se a primeira folha acima da espiga
principal exposta à radiação solar, em todas as parcelas. Os dados de gs(1)
foram
transformados em 2,0x para a realização da análise de variância.
(c) Potencial de água na folha ( w ): medido na primeira folha acima da espiga
principal, em cada parcela, com o uso de câmara tipo Scholander (Modelo 3115 da
Soilmoisture Equipment Corp), no período de manhã (7:00 às 8:00 h). No momento
dessa avaliação a temperatura máxima indicava 21°C, umidade relativa do ar de 54% e
do potencial matricial do solo de -1,00 MPa.

43
Figuras 5. Aspecto visual das plantas em déficit hídrico (a) e mostra os tratamentos
irrigados (direita) e déficit hídrico (esquerda) (b). Centro Experimental do Instituto
Agronômico, Campinas, 2013.
3.2.3.2 Avaliação biométrica em casa de vegetação
(a) Florescimento masculino (FM) e feminino (FF): obtidos pelo número de dias da
semeadura até que 50% das plantas de cada parcela estivessem com antese e estilos-
estigmas visíveis, respectivamente; O intervalo entre florescimentos feminino e
masculino (IF) correspondeu a diferença, em dias, entre o florescimento feminino e o
florescimento masculino; Os dados de IF foram transformados em x para a realização
das análises de variância.
(b) Altura da planta (AP) e da espiga (AE): obtidas pela medida tomada do nível do
solo até a inserção da folha bandeira e da espiga principal em cinco plantas da parcela,
respectivamente, em cm;
(c) Senescência retardada de folhas e colmo ou “stay-green” (SG): avaliado cerca de
120 dias após a semeadura após a maturidade fisiológica dos grãos, em cinco plantas
competitivas na parcela, a partir de uma escala de notas variando de 1 a 5, onde as notas
seguiram o seguinte padrão: 1- plantas com as folhas acima da primeira espiga e pelo
menos duas folhas abaixo da primeira espiga verdes; 2- plantas com todas as folhas
acima da primeira espiga verdes; 3- plantas com até duas folhas acima da primeira
a b

44
espiga secas e as demais verdes; 4- plantas com até duas folhas verdes; 5- plantas com
todas as folhas secas. Os dados de SG foram transformados em x para a realização
das análises de variância;
(d) Prolificidade (PROL): corresponde à relação número de espigas pelo número de
plantas da parcela;
(e) Massa seca total (MST): pesagem das frações folha, caule, sabugo e produção de
grãos de sete plantas de cada parcela, em kg parcela-1
. Essas amostras foram coletadas e
desidratadas em casa de vegetação de circulação forçada a 60˚C até a obtenção da massa
seca constante;
(f) Índice de colheita (IC): peso seco de grãos/(peso seco de grãos + peso seco da
planta)*100.
(g) Massa de grãos (MG): corresponde ao peso de grãos da parcela, em kg, resultantes
da debulha total de espigas de uma parcela, com auxílio de balança eletrônica. A MG
foi corrigida para umidade de 14% e estande ideal (24 plantas/ parcela). Para as análises
genético-estatísticas a MG foi convertida em g planta-1
dividindo-se o valor médio de
cada parcela pelo estande ideal do respectivo experimento. Para correção do estande
utilizou-se o método da covariância descrito por VENCOSVSKY & BARRIGA (1992).
3.2.4 Determinação do índice de valor relativo de tolerância (RVT)
Para expressar a tolerância das progênies e testemunhas foi utilizado um índice de
valor relativo de tolerância (RVT), onde o valor do caráter avaliado na condição
estressada foi dividido pelo valor da condição irrigada (SOUZA et al., 2011).
3.2.5 Análise de variância do experimento de casa de vegetação
O experimento de casa de vegetação foi em delineamento de parcelas
subdivididas, sendo o efeito de subparcela representado pelos genótipos e o efeito de
parcela os tratamentos hídricos, de acordo com o modelo, eq. (20):
ijkikkijijijk PSSPBY (20)
em que é a média geral; jB é o efeito do j ésimo bloco, sendo j= 1,2; iP é o efeito da i
ésima parcela, sendo i = 1, 2; ij é o erro aleatório a; kS é o efeito da k ésima

45
subparcela, sendo k = 1, 2; ..., 11; ikPS é o efeito da interação da i ésima parcela com a k
ésima subparcela; ijk é o erro aleatório b.
Todas as análises estatísticas foram realizadas utilizando-se o programa
computacional Genes versão Windows (CRUZ, 2006). Os dados fisiológicos e
biométricos foram agrupados pelo teste Scott-Knott a 5% de probabilidade.

46
4.0 RESULTADOS E DISCUSSÃO
4.1 Experimentos de avaliação das progênies interpopulacionais em campo
4.1.1 Análises de variâncias, médias e heterose
Os resumos das análises de variância individuais dos caracteres avaliados são
apresentados nas tabelas 4, 5, 6 e 7. FEDERER (1955) e MIRANDA FILHO (1987)
sugerem a análise de variância a partir do látice, somente quando sua eficiência for
superior a 110%. Para todos os caracteres o delineamento em látice foi mais eficiente do
que o de blocos casualizados (exceto para CE), indicando que o delineamento foi
adequado para se obter melhor controle local, sendo possível isolar o efeito da variação
de blocos dentro de repetições.
O coeficiente de variação experimental (CV%) da análise de variância conjunta
(Tabela 9) para MG foi de 10,6%, valor próximo aos encontrados na literatura. Para
todos os caracteres estes apresentaram nível satisfatório para a cultura do milho, de
acordo com a classificação proposta por SCAPIM et al. (1995). Os CV% variaram de
2,18% para NRP a 27,65% para A+Q (Tabelas 8 e 9), indicando que os experimentos
foram conduzidos com boa precisão experimental.
As análises de variância individuais para todos os caracteres apresentaram
diferenças altamente significativas entre progênies (p<0,01) para todos os caracteres em
todos os ambientes avaliados, exceto para CE em Mococa (Tabelas 4, 5, 6 e 7),
indicando a existência de ampla variabilidade genética das progênies para os diversos
caracteres. As análises de variâncias conjuntas apresentaram diferenças altamente
significativas (p<0,01) para os efeitos de ambientes, progênies e para a interação
progênies por ambientes para todos os caracteres, evidenciando respectivamente,
elevada variação entre os ambientes, presença de variabilidade genética na população e
performances diferenciais das progênies nos diferentes ambientes (Tabelas 8 e 9). Deste
modo, foi analisado o desempenho das progênies para cada um dos locais, descritos a
seguir.

47
Tabela 4. Resumo da análise de variância no delineamento em látice dos caracteres altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (A+Q), florescimento masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF) e stay-green (SG)
de 196 progênies interpopulacionais de milho em Campinas. Safrinha 2012.
Quadrados Médios
Fonte de Variação GL AP AE A + Q(1)
FM FF IF(2)
SG(3)
(cm) (cm) (%) (d.a.s.) (d.a.s.) (dias) (nota)
Repetições 1 3211,43 26,023 2,185 68,61 107,21 0,23 0,051
Blocos/rep. 26 1497,34 819,33 44,66 24,83 34,80 0,45 0,096
Progênies (aj.) 195 512,02** 475,47** 25,44** 9,46** 14,58** 0,17** 0,048**
Erro efetivo 169 114,31 39,75 11,73 1,24 3,56 0,076 0,022
Média 245,2 150,4 10,91 68,8 71,3 2,5 1,66
CV(%) 4,36 4,19 31,4 1,61 2,6 11,02 8,91
Eficiência do látice (%) 251,73 231,34 129,41 263,81 207,45 157,32 154,88 ns
e **
: não significativo e significativo a 1%, respectivamente, pelo teste F; (1)
: dados transformados para arcoseno 100)5,0( x ; (2,3)
: dados transformados para x .
Tabela 5. Resumo da análise de variância no delineamento em látice dos caracteres massa de grãos (MG), prolificidade (PROL), comprimento
de espiga (CE), número de fileiras de grãos na espiga (NFE) e número de ramificações do pendão (NRP) de 196 progênies interpopulacionais de
milho em Campinas. Safrinha 2012.
Quadrados Médios
Fonte de Variação GL
MG PROL CE NFE NRP(1)
(g planta-1
) (espigas.
planta-1
) (cm) (f.p.e.)
(ramificações
pendão-1
)
Repetições 1 7716,23 0,046 5,86 3,20 0,018
Blocos/rep. 26 1750,14 0,145 6,49 10,80 0,588
Progênies (aj.) 195 789,08** 0,075** 3,19** 5,310** 0,287**
Erro efetivo 169 262,81 0,023 0,647 0,35 0,140
Média 139,1 0,87 14,33 14,3 15,44
CV (%) 11,64 17,32 5,61 4,15 2,42
Eficiência do látice (%) 166,49 237,57 210,95 212,72 134,45
ns e
**: não significativo e significativo a 1%, respectivamente, pelo teste F; f.p.e.: fileiras por espiga;
(1)dados transformados para x .

48
Tabela 6. Resumo da análise de variância no delineamento em látice dos caracteres altura de planta (AP), altura de espiga (AE), plantas
acamadas e quebradas (A+Q), florescimento masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF) e stay-green (SG)
de 196 progênies interpopulacionais de milho em Mococa. Safrinha 2012.
Quadrados Médios
Fonte de Variação GL AP AE A + Q
(1) FM FF IF
(2) SG
(3)
(cm) (cm) (%) (d.a.s.) (d.a.s.) (dias) (nota)
Repetições 1 57,39 462,94 546,01 4,93 14,34 0,03 1,01
Blocos/rep. 26 922,15 515,71 94,07 16,70 20,06 0,57 5,44
Progênies (aj.) 195 431,72** 260,21** 46,69** 6,63** 8,30** 0,26** 0,033**
Erro efetivo 169 64,30 64,51 7,68 1,45 2,20 0,116 0,009
Média 241,8 134,9 11,44 69,2 71,3 2,1 1,48
CV (%) 3,31 5,97 24,23 1,74 2,08 16,23 6,65
Eficiência do látice (%) 268,24 184,08 240,36 230,34 198,78 143,94 151,08 ns
e **
: não significativo e significativo a 1%, respectivamente, pelo teste F. (1)
: dados transformados para arcoseno 100)5,0( x ; (2,3)
: dados transformados para x .
Tabela 7. Resumo da análise de variância no delineamento em látice dos caracteres massa de grãos (MG), prolificidade (PROL), comprimento
de espiga (CE), número de fileiras de grãos na espiga (NFE) e número de ramificações de pendão (NRP) de 196 progênies interpopulacionais de
milho em Mococa. Safrinha 2012.
Quadrados Médios
Fonte de Variação GL
MG PROL CE NFE NRP(1)
(g planta-1
) (espigas
planta-1
) (cm) (f.p.e.)
(ramificações
pendão-1
)
Repetições 1 1209,8 0,001 3,33 2,14 0,16
Blocos/rep. 26 353,44 1,365 1,62 9,46 0,93
Progênies (aj.) 195 189,73** 0,007** 1,56ns
3,79** 0,41**
Erro efetivo 169 102,25 0,005 1,27 0,49 0,08
Média 117,00 1,00 16,57 14,68 15,36
CV(%) 8,64 7,28 6,79 4,79 1,92
Eficiência do látice (%) 125,00 114,11 101,03 255,98 218,93
ns e
**: não significativo e significativo a 1%, respectivamente, pelo teste F; f.p.e: fileiras por espiga;
(1)dados transformados para x .

49
Tabela 8. Resumo da análise de variância conjunta dos caracteres altura de planta (AP), altura de espiga (AE), plantas acamadas e quebradas
(A+Q), florescimento masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF) e stay-green (SG) de 196 progênies
interpopulacionais em Campinas e Mococa. Safrinha, 2012.
Quadrados Médios
Fonte de Variação GL AP AE A+Q
(1) FM FF IF
(2) SG
(3)
(cm) (cm) (%) (d.a.s.) (d.a.s.) (dias) (nota)
Repetições/Locais 2 - - - - - - -
Locais 1 2931,44** 45110,7** 9129,73** 35,15** 1,14ns
2,19** 0,045**
Progênies (aj.) 195 559,80** 336,33** 34,86** 8,83** 14,09** 0,23** 0,804**
Progênies x Locais 195 383,94** 399,35** 37,28** 7,21** 8,80** 0,21** 0,034**
Resíduo Médio 338 89,30 52,13 9,71 1,35 2,88 0,096 0,016
Média 244,0 142,3 11,27 68,9 71,2 2,2 1,57
CV(%) 3,87 5,08 27,65 1,68 2,38 14,08 8,00 ns
e **
: não significativo e significativo a 1%, respectivamente, pelo teste F; (1)
: dados transformados para arcoseno 100)5,0( x ; (2,3)
: dados transformados para x .
Tabela 9. Resumo da análise de variância conjunta dos caracteres massa de grãos (MG), prolificidade (PROL), comprimento de espiga (CE),
número de fileiras de grãos na espiga (NFE) e número de ramificações do pendão (NRP) de 196 progênies interpopulacionais em Campinas e
Mococa. Safrinha, 2012.
Quadrados Médios
Fonte de Variação GL
MG PROL CE NFE NRP(1)
(g planta-1
) (espigas
planta-1
) (cm) (f.p.e.)
(ramificações
pendão-1
)
Repetições/Locais 2 - - - - -
Locais 1 83936,5** 3,339** 986,87** 24,07** 0,089ns
Progênies (aj.) 195 538,02** 0,041** 2,52** 4,72** 0,314**
Progênies x Locais 195 440,79** 0,041** 2,24** 4,38** 0,383**
Resíduo Médio 338 182,53 0,014 0,96 0,42 0,114
Média 127,35 0,93 15,45 14,50 15,40
CV(%) 10,60 12,72 6,33 4,49 2,18 ns
e **
: não significativo e significativo a 1%, respectivamente, pelo teste F; f.p.e.: fileiras por espiga; (1)
dados transformados para x .

50
As Tabelas 10 a 15 apresentam as médias das 40 progênies selecionadas para
todos os caracteres avaliados, considerando-se uma intensidade de seleção de 20%. Para
as 40 melhores progênies selecionadas em campo, a média geral de massa de grãos foi
de 159,81 g planta-1
(8.524 kg ha-1
), com amplitude de variação de 135,46 g planta-1
(7.225 kg ha-1
) a 181,12 g planta-1
(9.656 kg ha-1
) em Campinas (Tabela 10), e em
Mococa a média geral foi de 117,23 g planta-1
(6.252 kg ha-1
), com amplitude de
variação de 97,44 g planta-1
(5.197 kg ha-1
) a 149,07 g planta-1
(7.951 kg ha-1
) (Tabela
11). Em Mococa foi observado valores menores de MG para algumas progênies e
testemunhas indicando que a menor quantidade de precipitação pluvial total, podem ter
afetado o desenvolvimento e a produtividade desse experimento (Figuras 2 e 3). Houve
uma redução média de 27% da MG em relação ao experimento de Campinas (Tabelas
10 e 11).
As progênies mais promissoras apresentaram percentagens de MG(1)
em kg ha-1
,
em relação à média da variedade IAC Maya Latente, com amplitude de variação de
113,1% (PI(1,2)-52) a 151,3% (PI(1,2)-100) em Campinas (Tabela 10), e com amplitude de
variação de 102,9% (PI(1,2)-174) a 157,4% (PI(1,2)-52) em Mococa (Tabela 11). Estas
mesmas progênies também apresentaram desempenho semelhante em relação à
testemunha DAS2B707, com percentagens de MG(2)
que variaram de 74,4% (PI(1,2)-52)
a 99,51 (PI(1,2)-100) em Campinas, e de 80,7% (PI(1,2)-174) a 123,5% (PI(1,2)-52) em
Mococa. Com isso, verifica-se o potencial produtivo destas progênies obtidas do
cruzamento entre a Pop.Tol.1 e IAC Maya Latente, mesmo estando em seu primeiro
ciclo de seleção, em condições de safrinha. BÉTRAN et al. (2003) obtiveram híbridos
com bons desempenhos produtivos em níveis de estresse hídrico, indicando que é
possível combinar tolerância ao estresse com potencial produtivo em híbridos de milho
tropical.
GUIMARÃES (2013) obteve valores semelhantes de MG que variaram de 120,97
a 134,55 g planta-1
utilizando progênies de irmãos germanos interpopulacionais de
milho, na média de dois locais do Estado de São Paulo, Campinas e Mococa.
PATERNIANI et al. (2012) ressaltaram a importância da escolha adequada das
populações com potencial para tolerância à seca, cujas progênies da Pop.Tol.1
apresentaram valores de MG entre 111,02 a 177,02 g planta-1
e percentagem de
produção de 87,8% em relação ao híbrido comercial DAS2B710.
Os métodos de seleção recorrente interpopulacional dão ênfase às performances
dos cruzamentos, tentando aproveitar ao máximo o fenômeno da heterose nos

51
cruzamentos. Estes métodos surgiram a partir das hipóteses explicativas da heterose, ou
seja, da sobredominância versus dominância completa ou parcial atuando nos diversos
locos (HALLAUER et al., 1988). As estimativas de heterose média ( H ) e relativa
( %H ) do caráter MG em g planta-1
estão apresentados nas tabelas 10 e 11. Para o
caráter massa de grãos (MG), a H foi de 29,6 g planta-1
(22,7%) em Campinas, e de
30,7 g planta-1
(35,5%) em Mococa. Na média dos dois locais a H foi de 30,1 g planta-1
(27,82%) (Anexo 1).
No trabalho de REIS et al. (2014) os autores obtiveram para peso de espiga, valor
de H de 12,3% no ciclo 0 e de 24,9% no ciclo 3, utilizando progênies
interpopulacionais oriundas de híbridos comerciais. SOUZA Jr. et al. (1993) obtiveram
para H valor de 17,9% para peso de espigas em t/ha utilizando progênies
interpopulacionais derivadas das populações BR-105 x BR-106. BETRAN et al. (2003)
avaliaram linhagens e seus híbridos separadamente em experimentos sob estresse
hídrico, baixo teor de nitrogênio e condições ótimas de irrigação totalizando 12
ambientes. As diferenças de produtividade de grãos entre híbridos e suas linhagens
(heterose) aumentou com a intensidade do déficit hídrico (BETRAN et al., 2003). Este
resultado está de acordo com encontrado neste trabalho, onde Mococa foi o local que
obteve a maior heterose.
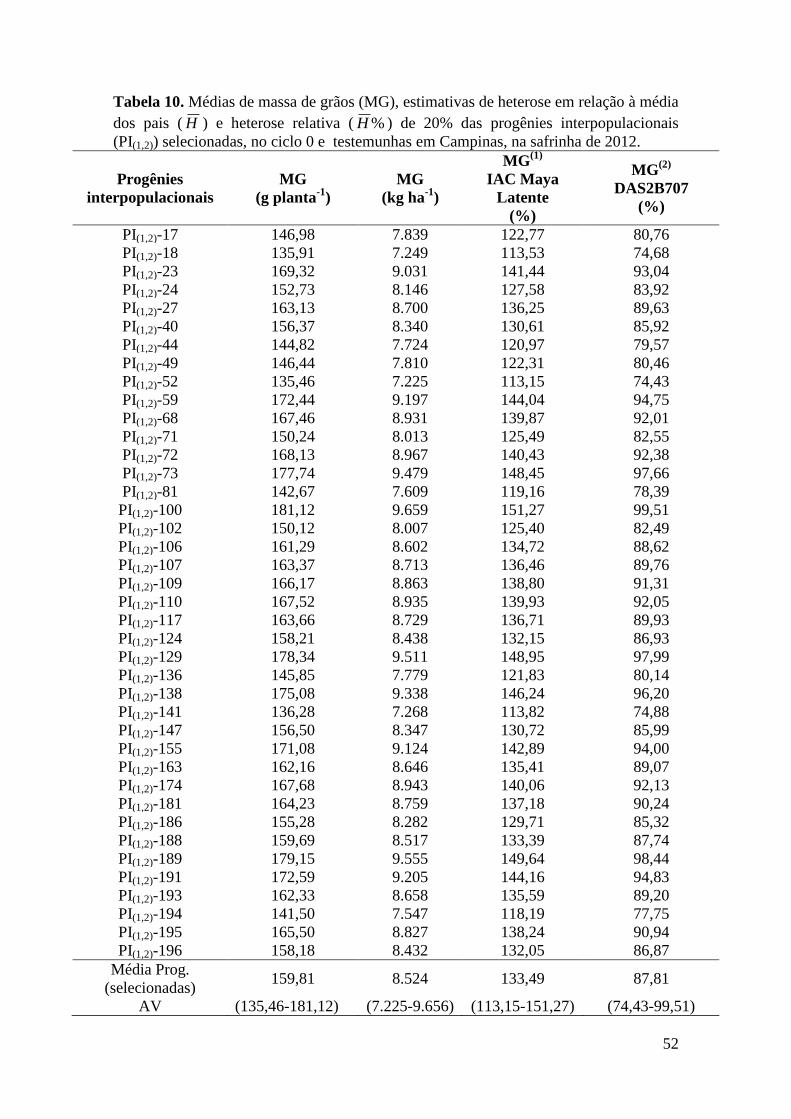
52
Tabela 10. Médias de massa de grãos (MG), estimativas de heterose em relação à média
dos pais ( H ) e heterose relativa ( %H ) de 20% das progênies interpopulacionais
(PI(1,2)) selecionadas, no ciclo 0 e testemunhas em Campinas, na safrinha de 2012.
Progênies
interpopulacionais
MG
(g planta-1
)
MG
(kg ha-1
)
MG(1)
IAC Maya
Latente
(%)
MG(2)
DAS2B707
(%)
PI(1,2)-17 146,98 7.839 122,77 80,76
PI(1,2)-18 135,91 7.249 113,53 74,68
PI(1,2)-23 169,32 9.031 141,44 93,04
PI(1,2)-24 152,73 8.146 127,58 83,92
PI(1,2)-27 163,13 8.700 136,25 89,63
PI(1,2)-40 156,37 8.340 130,61 85,92
PI(1,2)-44 144,82 7.724 120,97 79,57
PI(1,2)-49 146,44 7.810 122,31 80,46
PI(1,2)-52 135,46 7.225 113,15 74,43
PI(1,2)-59 172,44 9.197 144,04 94,75
PI(1,2)-68 167,46 8.931 139,87 92,01
PI(1,2)-71 150,24 8.013 125,49 82,55
PI(1,2)-72 168,13 8.967 140,43 92,38
PI(1,2)-73 177,74 9.479 148,45 97,66
PI(1,2)-81 142,67 7.609 119,16 78,39
PI(1,2)-100 181,12 9.659 151,27 99,51
PI(1,2)-102 150,12 8.007 125,40 82,49
PI(1,2)-106 161,29 8.602 134,72 88,62
PI(1,2)-107 163,37 8.713 136,46 89,76
PI(1,2)-109 166,17 8.863 138,80 91,31
PI(1,2)-110 167,52 8.935 139,93 92,05
PI(1,2)-117 163,66 8.729 136,71 89,93
PI(1,2)-124 158,21 8.438 132,15 86,93
PI(1,2)-129 178,34 9.511 148,95 97,99
PI(1,2)-136 145,85 7.779 121,83 80,14
PI(1,2)-138 175,08 9.338 146,24 96,20
PI(1,2)-141 136,28 7.268 113,82 74,88
PI(1,2)-147 156,50 8.347 130,72 85,99
PI(1,2)-155 171,08 9.124 142,89 94,00
PI(1,2)-163 162,16 8.646 135,41 89,07
PI(1,2)-174 167,68 8.943 140,06 92,13
PI(1,2)-181 164,23 8.759 137,18 90,24
PI(1,2)-186 155,28 8.282 129,71 85,32
PI(1,2)-188 159,69 8.517 133,39 87,74
PI(1,2)-189 179,15 9.555 149,64 98,44
PI(1,2)-191 172,59 9.205 144,16 94,83
PI(1,2)-193 162,33 8.658 135,59 89,20
PI(1,2)-194 141,50 7.547 118,19 77,75
PI(1,2)-195 165,50 8.827 138,24 90,94
PI(1,2)-196 158,18 8.432 132,05 86,87
Média Prog.
(selecionadas) 159,81 8.524 133,49 87,81
AV (135,46-181,12) (7.225-9.656) (113,15-151,27) (74,43-99,51)

53
Média Geral 139,10 7.344
Testemunhas
Pop. Tol. 1 138,79 6.961 109 71,70
Pop. Sensível 97,54 5.572 87,26 57,40
DAS2B707 181,58 9.709 152,01 100
IAC Maya Lat. 121,66 6.385 100 65,78
H 29,59
%H 22,72
CV(%) 11,64 11,77
(1,2): porcentagem em relação às testemunhas IAC Maya Latente e DAS2B707, respectivamente (em kg ha-1
).
Tabela 11. Médias de massa de grãos (MG), estimativas de heterose em relação à média
dos pais ( H ) e heterose relativa ( %H ) de 20% das progênies interpopulacionais
(PI(1,2)) selecionadas, no ciclo 0 e testemunhas em Mococa, na safrinha de 2012.
Progênies
interpopulacionais
MG
(g planta-1
)
MG
(kg ha-1
)
MG(1)
IAC Maya
Latente
(%)
MG(2)
DAS2B707
(%)
PI(1,2)-17 131,67 7.022 138,99 109,09
PI(1,2)-18 113,04 6.029 119,34 93,66
PI(1,2)-23 125,14 6.675 132,13 103,70
PI(1,2)-24 115,95 6.184 122,41 96,07
PI(1,2)-27 126,29 6.736 133,33 104,65
PI(1,2)-40 115,43 6.156 121,85 95,63
PI(1,2)-44 117,08 6.245 123,61 97,02
PI(1,2)-49 113,51 6.054 119,83 94,05
PI(1,2)-52 149,07 7.951 157,38 123,52
PI(1,2)-59 110,80 5.909 116,96 91,80
PI(1,2)-68 121,28 6.468 128,03 100,48
PI(1,2)-71 128,86 6.873 136,05 106,77
PI(1,2)-72 118,12 6.300 124,70 97,87
PI(1,2)-73 123,09 6.565 129,95 101,99
PI(1,2)-81 106,28 5.668 112,19 88,05
PI(1,2)-100 107,47 5.732 113,46 89,05
PI(1,2)-102 98,41 5.249 103,90 81,54
PI(1,2)-106 113,80 6.069 120,13 94,28
PI(1,2)-107 124,27 6.628 131,20 102,97
PI(1,2)-109 114,80 6.122 121,18 95,11
PI(1,2)-110 122,27 6.522 129,10 101,32
PI(1,2)-117 132,50 7.067 139,89 109,79
PI(1,2)-124 125,39 6.687 132,36 103,88
PI(1,2)-129 118,59 6.325 125,20 98,26
PI(1,2)-136 108,39 5.781 114,43 89,81
PI(1,2)-138 130,42 6.956 137,69 108,06
PI(1,2)-141 122,93 6.556 129,77 101,85
PI(1,2)-147 109,75 5.853 115,86 90,93
PI(1,2)-155 103,12 5.499 108,85 85,43
PI(1,2)-163 124,89 6.661 131,85 103,48
PI(1,2)-174 97,44 5.197 102,87 80,74
Tabela 10. (Continuação)

54
PI(1,2)-181 121,33 6.471 128,09 100,53
PI(1,2)-186 128,64 6.861 135,81 106,59
PI(1,2)-188 101,50 5.413 107,15 84,09
PI(1,2)-189 103,72 5.532 109,50 85,94
PI(1,2)-191 110,75 5.907 116,92 91,77
PI(1,2)-193 110,13 5.874 116,27 91,25
PI(1,2)-194 114,00 6.080 120,35 94,45
PI(1,2)-195 118,25 6.307 124,84 97,98
PI(1,2)-196 110,95 5.917 117,12 91,92
Média Prog.
(selecionadas) 117,23 6.252 123,75 97,13
AV (97,44-149,07) (5.197-7.951) (102,87-157,38) (80,74-123,52)
Média Geral 117,0 6.241
Testemunhas
Pop. Tol. 1 92,39 5.137 101,68 79,80
Pop. Sensível 88,04 5.120 101,35 79,54
DAS2B707 117,54 6.437 127,41 100,00
IAC Maya Lat. 80,66 5.052 100,00 78,48
H 30,70
%H 35,48
CV(%) 8,64 9,68
(1,2): porcentagem em relação às testemunhas IAC Maya Latente e DAS2B707, respectivamente (em kg
ha-1
).
Em Campinas, as médias das progênies interpopulacionais para os caracteres AP,
AE, FM, FF, IF, A+Q, NRP, SG, PROL, CE e NFE foram, respectivamente, de 241 cm,
153 cm, 68,9 dias, 71,7 dias, 2,9 dias, 7,01%, 15,17 ramificações pendão-1
, nota de 1,74,
0,88 espiga planta-1
, 14,55 cm e 14,3 fileira espiga-1
(Tabelas 12 e 13). Para Mococa, as
médias destes caracteres foram, respectivamente, de 237 cm, 132 cm, 69,5 dias, 71,8
dias, 2,4 dias, 13,79%, 14,9 ramificações pendão-1
, nota de 1,49, 1,00 espiga planta-1
,
16,48 cm e 15,0 fileira espiga-1
(Tabelas 14 e 15).
Quanto aos caracteres AP e AE, as médias foram de 241 e 153 cm para as
progênies interpopulacionais, de 272 e 150 cm para o IAC Maya Latente e de 249 e 124
cm para a Pop.Tol.1, respectivamente, indicando uma redução de AP das progênies em
relação aos parentais, em Campinas (Tabela 12). Em Mococa, o caráter AP também
apresentou este comportamento, com média de 237 cm para progênies
interpopulacionais e de 250 cm para o IAC Maya Latente (Tabela 14). No anexo 2,
encontra-se a heterose média e relativa de AP das progênies interpopulacionais,
indicando média de 2 cm (0,84%). No melhoramento genético de milho procuram-se
Tabela 11. (Continuação)

55
materiais que exibem pouca ou nenhuma heterose para o caráter altura de plantas, já que
as cultivares comerciais deve ser preferencialmente de porte baixo.
Para FM e FF, verificaram-se ampla variabilidade genética das progênies
interpopulacionais selecionadas, em Campinas e Mococa, com valores de amplitude de
variação de 60,9 e 72,6 d.a.s., e de 65,7 e 74,6 d.a.s, respectivamente (Tabelas 12 e 14).
Para IF, as progênies interpopulacionais obtiveram amplitude de variação de 0,6 a 6,4
dias e o IAC Maya Latente obteve valor de 2,5 dias, em Campinas (Tabela 12). Em
Mococa, foi verificado que o IF apresentou amplitude de variação de -1,2 a 5,5 dias para
as progênies e o IAC Maya Latente obteve valor de 2,8 dias (Tabela 14). Os resultados
de FM, FF e IF indicam comportamento precoce a tardia das progênies
interpopulacionais. O mecanismo de diminuir o intervalo entre florescimentos em
ambientes sujeitos a déficit hídrico apresenta de extrema vantagem do “escape à seca”,
resultando na melhor adaptação dos genótipos nesses ambientes devido ao sincronismo
dos florescimentos.
O reduzido IF assegura que um número maior de plantas receberá pólen no
momento da emergência dos estilos-estigmas, garantindo a polinização e a fertilização
(WESTGATE et al., 2004). GAMA et al. (2001) enfatizaram a necessidade da utilização
de caracteres secundários como o IF, além da seleção per se para produtividade de
grãos, a fim de obter progresso mais rápido dentro do programa de melhoramento. O
sincronismo floral é influenciado pelas condições ambientais adversas, principalmente
pelo estresse hídrico, onde a emergência dos estilos-estigmas pode atrasar em relação à
liberação do pólen. Conjuntamente, este resultado sugere que a seleção para tolerância à
seca utilizando IF pode resultar na seleção direta para protoginia (negativo valor de IF),
sendo esta abordagem bastante prática nos programas de melhoramento. WESTGATE
(1997) indica grande vantagem desta prática de seleção onde o menor IF indica maior
sincronia entre máxima exposição dos pólens durante a emergência dos estilos-estigma,
levando assim maior estabilidade da produção dos grãos.
BOLAÑOS & EDMEADES (1996) obtiveram para o caráter IF média de 1,32
dias (variando de -1,84 a 5,71 dias), enquanto GUIMARÃES (2013) obteve para IF
valor de 1,06 dias na Pop.Tol.1, com amplitude de variação de 0,33 a 1,67 dias, em
Campinas e Mococa. Outros autores relataram que a seleção recorrente para tolerância à
seca diminuiu o IF em 1,3 dias por ciclo na população LPS (variedade sintética do
Tuxpeño) (EDMEADES et al., 1999), e de 0,6 e 0,2 dias por ciclo nas populações DTP1
e DTP2, respectivamente. Devido à relação existente entre IF e MG (BOLAÑOS &

56
EDMEADES, 1996), observa-se que a alta variação do intervalo entre florescimentos
pode resultar em baixa variação da produtividade de grãos quando o IF é alto (seja por
causa do nível de déficit hídrico ou da suscetibilidade do germoplasma)
(MONNEVEUX et al., 2008).
Para o caráter SG, foi verificado amplitude de variação de notas (1,03 a 3,00) para
as progênies interpopulacionais, nota de 1,61 para o IAC Maya Latente e, nota de 0,94
para o híbrido DAS2B707, em Campinas (Tabela 13) e, em Mococa o amplitude de
variação de notas foi de (0,97 a 2,38) para as progênies, nota de 1,03 para o IAC IAC
Maya Latente e, nota de 0,97 para o híbrido DAS2B707 (Tabela 15). Observando que a
nota 1 era atribuída à planta com “stay-green” e a nota 5 à planta sem “stay-green”,
esses resultados indicam que as progênies selecionadas podem ser utilizadas para o
desenvolvimento de variedades sintéticas e de linhagens com elevado “stay-green”.
Em Campinas, a média de A+Q foi de 7,01%, com amplitude de variação de 1,21
a 18,33%, para as progênies interpopulacionais, destacaram as progênies PI(1,2)-68,
PI(1,2)-194 e PI(1,2)-196, pela tolerância ao acamamento e quebramento de plantas
(Tabela 13). Em Mococa (Tabela 15), foi observada maior média de A+Q de 13,8%,
com amplitude de variação de 3,14 a 21,04%, para progênies, que também apresentaram
menores valores de plantas acamadas + quebradas, com destaque para PI(1,2)-40 e PI(1,2)-
106 e o híbrido DAS2B707.
A PROL apresentou amplitude de variação de 0,52 (PI(1,2)-23) a 1,51 espigas
planta-1
(PI(1,2)-102), destacando a PI(1,2)-102, em Campinas (Tabela 13). Em Mococa a
PROL variou de 0,83 (PI(1,2)-117) a 1,1 espigas planta-1
(PI(1,2)-102) para as progênies
interpopulacionais, e de 1,58 espigas planta-1
para o IAC IAC Maya Latente (Tabela
15). Esses resultados mostram que a PI(1,2)-102 obteve os maiores valores de PROL nos
dois locais e a variedade IAC IAC Maya Latente destacou-se por ser bastante prolífica
em ambientes sujeitos a déficit hídrico (Mococa). BOLAÑOS & EDMEADES (1996)
encontraram para prolificidade valor médio de 0,98 (variando de 0,65 a 1,37 espigas por
planta-1
). Resultados semelhantes foram obtidos por CÂMARA et al. (2007) em
populações derivadas de milho tropical, que indicaram baixa prolificidade (0,70 espiga
planta-1
), comportamento protogínicas com valores negativos de intervalo entre
florescimentos e pouco retardamento da senescência de folhas e colmos (nota de 3,44).
A relação de competição por fotoassimilados dos órgãos reprodutivos, feminino
(espiga) e masculino (pendão), está principalmente relacionada à própria organização
floral do milho que é monóica e protândrica, que assegura a produção de pólen e sua

57
dispersão nas espigas (BOLAÑOS et al, 1993). Nesse estudo, a redução do número de
ramificações de pendão (NRP) pode levar a um importante aumento da produtividade
em condições de safrinha. Em Campinas e Mococa, o caráter NRP apresentou média de
15 ramificações pendão-1
(Tabelas 13 e 15) para as progênies interpopulacionais, com
amplitude de variação de (10 a 21) e (9 a 21) ramificações pendão-1
, respectivamente.
GUIMARÃES (2013) encontrou para NRP valor médio de 12,3 ramificações pendão-1
,
do qual foi menor ao deste trabalho com valor médio de 15 ramificações pendão-1
,
indicando que a incorporação de germoplasma tropical (IAC Maya Latente) tende a
refletir em aumentos de NRP. DE LEON & COORS (2002) relataram que a seleção
baseada na prolificidade diminuiu o comprimento e número de ramificações de pendão,
confirmando, assim, a competição desses caracteres.
Com relação à heterose H das 40 progênies para os caracteres IF, NRP e SG
foram, respectivamente, de 0,29 dias (12,4%), de 1,0 ramificações pendão-1
(7,1%) e de
0,38 nota (28%) (Anexos 2 e 3). AMINU et al. (2014) relataram valores de
heterobeltiose de -38,33 a 5,27% para IF em híbridos top-cross de milho. Neste trabalho
foram observados desejáveis níveis de heterose para IF, NRP e SG indicando favoráveis
para seleção interpopulacional.
Para os caracteres CE e NFE (Tabelas 13 e 15) das progênies interpopulacionais
foram observados valores similares aos encontrados para o híbrido comercial
DAS2B707, indicando uma tendência das progênies mais produtivas apresentarem
também elevados componentes de produção. Com isso, a seleção indireta para produção
de grãos com base nestes caracteres, que apresentam correlação alta e significativa com
MG, poderia proporcionar maiores ganhos do que a seleção direta para este caráter.
Os resultados obtidos para as progênies interpopulacionais indicam
comportamento produtivo superior em relação aos parentais (Pop.Tol.1 e IAC Maya
Latente), evidenciando que as progênies selecionadas superaram os parentais,
manifestando elevada heterose. A identificação dessas progênies interpopulacionais
evidencia ainda mais o fato de que as duas populações são divergentes, possibilitando a
extração de linhagens e desenvolvimento de híbridos superiores com as características
desejadas. Ganhos constantes no melhoramento de milho para tolerância à seca têm sido
obtidos (BÄNZINGER et al., 2006; SETIMELA et al.; 2012). Pesquisadores do
CIMMYT (BOLAÑOS & EDMEADES, 1993; EDMEADES et al., 1997a) obtiveram
ganho de produtividade utilizando como fonte de tolerância as populações La Posta
Sequia e DTP desenvolvidas nos anos 1970, 1980 e 1990, através de seleção recorrente.

58
Esses autores indicaram que as linhagens obtidas dessas populações foram
desenvolvidas e melhoradas com estudos de fisiologia em conjunto o melhoramento de
plantas durante 15 anos.
Neste trabalho foi destacada a importância de obter novas populações com fontes
de tolerância para o desenvolvimento contínuo de novas cultivares com níveis mais
elevados de tolerância à seca como os apresentados pelas 40 progênies selecionadas.
Diante disso, também foram discutidos os dados médios de seleção das 8 progênies
interpopulacionais como alternativa para obtenção de sintéticos, sendo estas
selecionadas entre as 40 PI(1,2), quanto a MG, SG e IF no experimento de Mococa para
serem avaliadas em casa de vegetação (item 4.2). As médias das 8 progênies estão
apresentadas no Anexo 4 para todos caracteres avaliados. Para MG o valor médio foi de
139,18 g planta-1
(7.408 kg ha-1
) com heterose ( H ) de 30,8 g planta-1
(28,4%). Para os
caracteres AP, AE, FM, FF, IF, A+Q, NRP, SG, PROL, CE e NFE foram,
respectivamente, de 244 cm, 145 cm, 69,5 d.a.s., 71,1 d.a.s., 1,7 dias, 11,3%, 15,5
ramificações pendão-1
, nota 1,7, 0,96 espiga planta-1
, 15,5 cm e 14,1 fileira por espiga
(Anexo 4). Em relação à média original (196 progênies) os caracteres MG e IF das 8
progênies se destacaram para maior produtividade de grãos e menor IF.
A heterose média ( H ) obtida para as 8 progênies foi de 6 cm (2,52%) para AP,
de -0,65 dias (-27,9%) para IF, de 1,5 ramificações pendão-1
(10,7%) e de nota 0,45
(36%) para SG (Anexo 4), indicam que esses caracteres contidos nas 8 progênies podem
ser recomendados para obtenção de variedade sintética tolerante à seca, sendo este
método de melhoramento indicado no desenvolvimento de sintéticos que concentram
genes específicos. Portanto, a heterose encontrada neste trabalho indica grande
diversidade entre as progênies interpopulacionais, o que é um indicativo do grande
potencial na síntese de populações para melhoramento de sintéticos ou compostos.
Essas populações vão possuir uma grande variabilidade genética, ou o aproveitamento
direto de informações de cruzamentos específicos em seleção recorrente ou de parentais
(testadores) de híbridos intervarietais do programa de melhoramento de milho do
Instituto Agronômico.

59
Tabela 12. Médias de altura de planta (AP), altura de espiga (AE), florescimento
masculino (FM), florescimento feminino (FF) e intervalo entre florescimentos (IF) de
20% das progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas em
Campinas, na safrinha de 2012.
Progênies
interpopulacionais
AP
(cm)
AE
(cm)
FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
PI(1,2)-17 261 163 65,2 69,1 3,9
PI(1,2)-18 229 158 66,1 69,5 3,3
PI(1,2)-23 256 153 66,3 70,2 3,9
PI(1,2)-24 252 155 65,2 67,4 2,2
PI(1,2)-27 267 149 68,4 71,6 3,1
PI(1,2)-40 256 179 72,1 74,6 2,5
PI(1,2)-44 244 159 71,0 72,4 1,4
PI(1,2)-49 260 168 68,3 70,2 1,9
PI(1,2)-52 257 178 68,2 69,2 1,0
PI(1,2)-59 235 156 68,2 72,9 4,8
PI(1,2)-68 236 158 71,9 74,5 2,5
PI(1,2)-71 261 163 66,8 69,4 2,6
PI(1,2)-72 243 161 67,0 68,9 1,9
PI(1,2)-73 219 146 65,6 69,9 4,3
PI(1,2)-81 255 162 72,6 74,5 1,9
PI(1,2)-100 249 125 67,6 70,7 3,1
PI(1,2)-102 225 167 70,7 77,1 6,4
PI(1,2)-106 272 156 69,4 70,3 0,9
PI(1,2)-107 228 150 69,8 74,3 4,5
PI(1,2)-109 241 145 72,6 73,3 0,6
PI(1,2)-110 246 147 70,4 72,7 2,3
PI(1,2)-117 237 147 70,9 75,4 4,5
PI(1,2)-124 229 166 72,6 73,9 1,4
PI(1,2)-129 241 157 67,8 69,6 1,8
PI(1,2)-136 223 121 70,9 72,5 1,6
PI(1,2)-138 260 154 72,6 74,7 2,1
PI(1,2)-141 236 159 66,0 68,8 2,7
PI(1,2)-147 249 182 66,1 68,7 2,6
PI(1,2)-155 254 139 69,6 73,4 3,8
PI(1,2)-163 242 127 71,5 74,9 3,4
PI(1,2)-174 254 142 70,3 73,1 2,9
PI(1,2)-181 239 123 70,9 75,6 4,7
PI(1,2)-186 217 167 66,3 69,5 3,2
PI(1,2)-188 217 142 60,9 64,7 3,7
PI(1,2)-189 232 150 70,0 75,5 5,5
PI(1,2)-191 223 145 67,4 70,2 2,8
PI(1,2)-193 256 160 69,8 71,7 1,9
PI(1,2)-194 223 159 70,6 73,7 3,1
PI(1,2)-195 210 144 67,6 69,6 2,0
PI(1,2)-196 225 140 68,9 70,9 2,1
Média Prog.
(selecionadas) 241 153 68,9 71,7 2,9
IV (210-272) (121-182) (60,9-72,6) (64,7-77,1) (0,6-6,4)
Média geral 245 150 68,8 71,3 2,5

60
Testemunhas
Pop.Tol.1 249 124 68,2 70,9 2,8
Pop. Sensível 205 130 67,9 71,2 3,4
DAS2B707 232 113 69,9 73,3 3,4
Maya Lat. 272 150 68,5 71,1 2,5
CV(%) 4,36 4,19 1,61 2,6 50,5
Tabela 13. Médias de plantas acamadas e quebradas (A+Q), número de ramificações do
pendão (NRP), stay-green (SG), de prolificidade (PROL), comprimento de espiga (CE)
e número de fileiras de grão na espiga (NFE) de 20% das progênies interpopulacionais
(PI(1,2)) selecionadas, no ciclo 0 e testemunhas em Campinas, na safrinha de 2012.
Progênies
Interp.
A+Q(1)
(arcoseno)
NRP
(ramif.
pend.-1
)
SG
(nota)
PROL
(espigas
planta-1
)
CE
(cm)
NFE
(f.p.e.)
PI(1,2)-17 13,55 18 1,86 1,08 13,85 16,9
PI(1,2)-18 12,82 15 1,80 1,05 14,41 10,3
PI(1,2)-23 11,62 14 1,70 0,52 13,32 14,4
PI(1,2)-24 10,22 13 2,45 0,83 15,41 16,1
PI(1,2)-27 18,33 12 1,95 0,97 14,78 13,4
PI(1,2)-40 3,67 20 1,70 0,92 13,71 14,5
PI(1,2)-44 6,84 11 1,04 0,70 13,14 16,6
PI(1,2)-49 5,73 14 1,05 0,77 14,91 12,6
PI(1,2)-52 8,39 12 3,00 1,39 13,32 14,3
PI(1,2)-59 3,75 17 1,06 0,96 14,98 12,9
PI(1,2)-68 1,21 14 1,06 0,57 13,75 13,5
PI(1,2)-71 3,84 16 2,07 0,92 15,49 11,9
PI(1,2)-72 6,79 18 1,13 0,63 13,26 12,5
PI(1,2)-73 5,71 17 1,31 1,26 14,52 11,7
PI(1,2)-81 13,82 21 1,28 0,71 14,83 15,9
PI(1,2)-100 6,60 15 1,51 1,10 11,73 14,3
PI(1,2)-102 8,72 17 1,03 1,51 13,29 13,1
PI(1,2)-106 2,99 18 2,02 0,73 14,97 12,3
PI(1,2)-107 2,48 13 1,73 1,21 14,39 12,9
PI(1,2)-109 8,53 19 1,66 0,83 15,49 15,3
PI(1,2)-110 5,59 18 2,09 0,60 13,12 12,2
PI(1,2)-117 5,36 13 2,15 0,95 13,44 16,3
PI(1,2)-124 3,88 16 2,16 0,71 14,14 17,2
PI(1,2)-129 4,99 17 1,56 1,21 12,00 15,9
PI(1,2)-136 11,67 12 2,35 0,84 14,22 16,8
PI(1,2)-138 4,96 13 2,06 0,84 14,75 17,4
PI(1,2)-141 5,04 16 2,33 0,94 16,35 14,9
PI(1,2)-147 6,04 19 2,12 0,66 15,53 12,7
PI(1,2)-155 7,96 15 1,74 0,79 16,80 13,9
PI(1,2)-163 4,82 13 1,03 0,67 13,80 13,7
PI(1,2)-174 2,63 21 1,21 0,80 15,47 12,3
PI(1,2)-181 12,51 18 1,38 0,62 16,22 15,8
PI(1,2)-186 7,26 14 1,31 1,10 15,94 13,7
PI(1,2)-188 6,90 16 1,99 0,65 14,78 14,6
Tabela 12. (Continuação)

61
PI(1,2)-189 10,39 10 1,61 0,78 13,39 16,7
PI(1,2)-191 4,57 12 2,31 0,90 15,22 14,7
PI(1,2)-193 9,22 14 1,74 0,85 16,10 15,4
PI(1,2)-194 1,44 12 2,67 0,97 14,61 13,0
PI(1,2)-195 7,95 10 1,76 0,75 18,24 13,3
PI(1,2)-196 1,71 14 1,83 0,95 14,61 14,7
Média Prog.
(selecionadas) 7,01 15,17 1,74 0,88 14,55 14,3
IV (1,21-18,33) (10-21) (1,03-3,00) (0,52-1,51) (11,73-18,24) (10,3-17,4)
Média geral 10,9 15,44 1,66 0,87 14,33 14,3
Testemunhas
Pop. Tol. 1 8,28 15 1,36 1,01 13,64 18,0
Pop. Sensível 5,10 14 1,41 0,86 14,18 14,0
DAS2B707 8,60 17 0,94 0,56 14,75 16,0
Maya Lat. 14,11 16 1,61 1,10 14,18 16,0
CV(%) 31,4 2,42 8,91 17,32 5,61 4,15
(1)
: dados transformados para arcoseno 100)5,0( x ; f.p.e: fileiras por espiga;
Tabela 14. Médias de altura de planta (AP), altura de espiga (AE), florescimento
masculino (FM), florescimento feminino (FF) e intervalo entre florescimentos (IF) de
20% das progênies interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas em
Mococa, na safrinha de 2012.
Progênies
interpopulacionais
AP
(cm)
AE
(cm)
FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
PI(1,2)-17 240 132 65,9 69,5 3,5
PI(1,2)-18 235 145 65,7 69,2 3,5
PI(1,2)-23 236 139 66,1 69,3 3,1
PI(1,2)-24 233 133 69,1 72,7 3,6
PI(1,2)-27 241 142 66,9 69,5 2,6
PI(1,2)-40 219 132 69,0 71,5 2,5
PI(1,2)-44 259 135 68,2 71,7 3,5
PI(1,2)-49 248 144 69,1 69,8 0,8
PI(1,2)-52 244 136 71,6 72,9 1,4
PI(1,2)-59 243 135 67,9 69,5 1,6
PI(1,2)-68 218 147 70,2 72,6 2,4
PI(1,2)-71 252 139 68,3 71,2 2,9
PI(1,2)-72 261 147 71,6 71,4 -0,2
PI(1,2)-73 237 132 71,3 73,9 2,6
PI(1,2)-81 250 145 68,3 69,6 1,3
PI(1,2)-100 244 126 71,3 72,3 0,9
PI(1,2)-102 222 121 68,3 71,5 3,3
PI(1,2)-106 233 128 71,0 73,1 2,1
PI(1,2)-107 216 117 67,7 71,1 3,3
PI(1,2)-109 219 109 68,5 69,4 0,9
PI(1,2)-110 207 129 68,6 71,8 3,2
PI(1,2)-117 245 130 69,4 70,1 1,0
PI(1,2)-124 240 133 68,0 72,1 4,1
PI(1,2)-129 249 138 68,3 71,4 3,1
Tabela 13. (Continuação)

62
PI(1,2)-136 232 139 70,5 74,6 4,2
PI(1,2)-138 227 132 68,6 70,9 2,3
PI(1,2)-141 258 131 71,4 72,1 0,6
PI(1,2)-147 231 126 74,6 74,5 -0,2
PI(1,2)-155 243 121 68,1 71,8 3,7
PI(1,2)-163 235 139 68,3 73,8 5,5
PI(1,2)-174 234 139 67,7 67,7 -0,01
PI(1,2)-181 236 129 70,8 69,7 -1,2
PI(1,2)-186 223 123 69,4 72,5 3,1
PI(1,2)-188 234 118 72,6 74,9 2,3
PI(1,2)-189 247 145 72,4 74,4 2,0
PI(1,2)-191 242 133 68,9 71,1 2,2
PI(1,2)-193 255 129 69,7 72,9 3,2
PI(1,2)-194 238 147 72,0 76,3 4,3
PI(1,2)-195 249 117 73,7 76,3 2,6
PI(1,2)-196 238 121 69,8 72,4 2,6
Média Prog.
(selecionadas) 237 132 69,5 71,8 2,4
IV (207-261) (109-147) (65,5-74,6) (67,7-76,3) (-1,2-5,5)
Média geral 241,8 134,9 69,2 71,3 2,1
Testemunhas
Pop. Tol. 1 180 118 69,0 70,3 1,3
Pop. Sensível 200 117 69,9 72,1 2,1
DAS2B707 205 114 69,0 70,7 1,7
IAC Maya Lat. 250 142 69,0 71,8 2,7
CV(%) 3,31 5,97 1,74 2,08 59,55
Tabela 15. Médias de plantas acamadas e quebradas (A+Q), número de ramificações do
pendão (NRP), stay-green (SG), prolificidade (PROL), comprimento de espiga (CE) e
número de fileiras de grãos na espiga (NFE) de 20% das progênies interpopulacionais
(PI(1,2)) selecionadas, no ciclo 0 e testemunhas em Mococa, na safrinha de 2012.
Progênies
interpopulacionais
A+Q(1)
(arcsen)
NRP
(ramif.
pendão-1
)
SG
(nota)
PROL
(espigas
planta-1
)
CE
(cm)
NFE
(f.p.e.)
PI(1,2)-17 13,15 13 1,26 1,01 15,88 15,1
PI(1,2)-18 11,99 16 1,83 1,01 17,05 15,4
PI(1,2)-23 18,15 14 1,20 1,00 14,73 15,7
PI(1,2)-24 12,02 17 1,64 1,10 15,94 13,9
PI(1,2)-27 9,17 11 1,41 1,03 16,78 15,7
PI(1,2)-40 4,91 19 1,51 0,93 15,89 15,5
PI(1,2)-44 19,28 16 1,35 0,99 16,98 14,8
PI(1,2)-49 14,81 14 2,18 1,04 15,98 14,1
PI(1,2)-52 13,47 15 1,52 0,99 18,15 12,8
PI(1,2)-59 21,04 10 0,97 1,05 17,29 15,1
PI(1,2)-68 14,27 10 1,10 1,00 16,84 15,0
PI(1,2)-71 6,82 9 1,70 1,09 15,93 15,1
PI(1,2)-72 16,08 11 1,48 1,00 16,35 13,0
Tabela 14. (Continuação)

63
PI(1,2)-73 11,64 12 1,28 1,01 16,91 14,1
PI(1,2)-81 20,22 9 1,42 1,01 16,89 13,2
PI(1,2)-100 18,13 19 1,26 1,03 14,98 13,7
PI(1,2)-102 15,72 21 1,32 1,10 14,21 16,1
PI(1,2)-106 3,14 12 1,18 1,09 16,19 15,5
PI(1,2)-107 16,45 18 1,79 1,01 16,69 16,3
PI(1,2)-109 14,24 14 1,30 0,99 16,69 15,9
PI(1,2)-110 7,66 20 1,29 0,95 16,88 15,7
PI(1,2)-117 18,65 19 2,11 0,83 15,54 15,0
PI(1,2)-124 9,61 13 1,70 0,95 16,18 15,7
PI(1,2)-129 16,62 13 1,64 1,04 17,23 14,0
PI(1,2)-136 13,28 15 1,71 0,98 16,52 14,8
PI(1,2)-138 11,43 19 1,17 0,98 17,87 16,9
PI(1,2)-141 13,64 19 2,08 0,98 16,71 14,7
PI(1,2)-147 17,85 15 2,38 1,01 17,93 13,8
PI(1,2)-155 12,99 16 1,85 1,03 18,15 13,0
PI(1,2)-163 19,83 13 1,35 0,99 16,39 14,6
PI(1,2)-174 20,91 13 1,03 1,07 16,55 13,9
PI(1,2)-181 15,95 15 1,03 1,04 16,64 12,6
PI(1,2)-186 8,12 17 1,46 1,00 14,67 16,9
PI(1,2)-188 9,87 20 1,63 1,04 16,49 16,1
PI(1,2)-189 12,37 15 1,61 1,00 16,54 15,7
PI(1,2)-191 16,51 16 1,62 1,00 16,71 17,6
PI(1,2)-193 14,86 13 1,34 1,00 18,72 18,2
PI(1,2)-194 11,82 15 1,52 0,99 17,01 13,3
PI(1,2)-195 9,27 14 1,43 0,97 16,31 15,4
PI(1,2)-196 16,05 16 1,32 1,03 14,20 16,7
Média Prog.
(selecionadas) 13,79 14,9 1,49 1,00 16,48 15,0
IV (3,14-21,04) (9-21) (0,97-2,38) (0,83-1,1) (14,2-18,7) (12,6-18,2)
Média geral 11,44 15,36 1,48 1,48 16,57 14,68
Testemunhas
Pop. Tol. 1 9,93 10 1,00 1,10 18,24 14,0
Pop. Sensível 10,66 12 0,99 1,50 17,43 14,0
DAS2B707 7,37 9 0,97 0,96 16,94 16,0
IAC Maya Lat. 10,22 15 1,03 1,58 17,26 16,0
CV (%) 24,23 1,92 6,65 7,28 6,79 4,79
(1)
: dados transformados para arcoseno 100)5,0( x ; f.p.e: fileiras por espiga.
Tabela 15. (Continuação)

64
4.1.2 Estimativas de parâmetros genéticos interpopulacionais
No caso que se deseja maximizar os ganhos para uma série de ambientes, o
apropriado é estimar este ganho utilizando o componente de variância genética da
análise conjunta (CRUZ et al., 2004). No entanto, neste estudo pretende-se selecionar
progênies interpopulacionais que venham a formar populações com adaptações
específicas aos locais avaliados. Assim, será realizada uma análise dos parâmetros
genéticos obtidos a partir das estimativas de cada ambiente.
As estimativas dos parâmetros genéticos, com base nas análises individuais, para
todos os caracteres avaliados são apresentadas, nas Tabelas 16 e 17, para os
experimentos realizados em Campinas e Mococa, respectivamente.
O componente de variância de progênies ()2,1(
2ˆp ) é resultado das diferenças
genéticas entre os indivíduos. Um valor elevado desse componente é indicativo de
ampla variabilidade genética para o caráter, sendo de importância na identificação de
genótipos superiores. As estimativas para massa de grãos (MG) foram de 394,5 (g
planta-1
)2 e 94,8 (g planta
-1)2, em Campinas e Mococa, respectivamente. Resultados
obtidos por GUIMARÃES et al. (2012) e PATERNIANI et al. (2012) em híbridos inter
e intrapopulacionais de milho, respectivamente, são menores aos encontrados neste
trabalho, obtendo para o componente de variância genética de progênies para MG 20,01
(g planta-1
)2 nos híbridos interpopulacionais e de 28,89 (g planta
-1)2 nos
intrapopulacionais.
A variância aditiva é relativa ao efeito médio dos alelos, sendo a de maior
importância no melhoramento. Esta variância é a causa de semelhança entre parentes, já
que os pais passam seus alelos e não os seus genótipos para sua progênie, ou seja, é o
efeito médio dos alelos parentais, que determina a média do valor genotípico de sua
progênie (FALCONER, 1989). A variância genética aditiva apresentada pelas progênies
interpopulacionais é o de alta magnitude e constitui indicativo da possibilidade de
ganho em todos os locais (tabelas 16 e 17). Porém, as estimativas da variância genética
aditiva em Campinas apresentaram valores muito próximos e superaram a estimativa
obtida em Mococa para os caracteres avaliados. O valor da estimativa de massa de grãos
(MG) foi de 194,9 (g.planta-1
)2 e 1052,5 (g.planta
-1)2 em Mococa e Campinas,
respectivamente, indicando que estes valores foram de magnitude de baixa a alta,
quando comparada com valores apresentados a outros pesquisadores em outros estudos.
PATERNIANI et al. (2004) e NASS & MIRANDA FILHO (1999) obtiveram para

65
variância genética aditiva, valores oscilando de 502,00 a 821,56 (g planta-1
) 2, para peso
de espiga.
PATERNIANI (1968) ressalta que é do máximo interesse que a variância genética
aditiva permaneça tão alta quanto possível, para permitir progressos substanciais por
seleção. PATERNIANI et al. (2012), encontraram estimativas menores de 2ˆA de
115,55 (g planta-1
)2 para MG, utilizando ciclo 0 da Pop.Tol.1. Valores altos de variância
aditiva foram encontrados em populações com ciclos iniciais de seleção, onde toda a
variabilidade genética livre é liberada (RAMALHO, 1977), que corresponde à
segregação de blocos poligênicos. A partir do primeiro ciclo de seleção, é utilizada a
variabilidade genética, presente dentro de blocos poligênicos, que é liberada
gradativamente por meio da permuta genética.
As estimativas do coeficiente de variação genética (gVC ˆ ) permitem ao melhorista
ter uma noção de grandeza relativa das mudanças, que podem ser obtidas por meio da
seleção, ao longo de um programa de melhoramento. Em Campinas, os caracteres A+Q,
PROL, MG e NFE apresentaram as maiores estimativas do coeficiente de variação
genético (CVg) de 23,9%, 11,6%, 18,5% e 11,0%, respectivamente. Em Mococa,
destacaram-se os caracteres A+Q e IF com maiores estimativas de CVg de 38,6% e
12,7%, respectivamente.
Para a relação entre o coeficiente de variação genético e o coeficiente de variação
experimental (CVg/CVe) o destaque é para todos os caracteres avaliados, em Campinas
(tabela 16) e Mococa (tabela 17), exceto para PROL e CE, em Mococa. As estimativas
do índice b foram, em geral, alta para os caracteres, indicando baixa influência do
ambiente sobre a variação. Observa-se, portanto, que a população envolvida neste
estudo apresenta um valor superior demonstrando boa possibilidade para a sua
exploração em programas de melhoramento. Além disso, estimativas de b na média de
dois ou mais locais, normalmente, apresentam-se inferiores às estimativas com base em
apenas um local, no qual é mais comum encontrar valores bem próximos ou superiores
a 1. Para MG o índice de seleção foi de 1,41, em Campinas e de 0,92, em Mococa, que
representam um valor significativo para seleção em termos de seleção em fase inicial
(C0) das progênies. FALUBA (2008) encontrou valor de índice de seleção de 0,64 para
MG e PATERNIANI et al. (2012), obtiveram valores de b de 0,53 para MG. As
progênies interpopulacionais de milho, avaliadas por GUIMARÃES et al. (2012),
apresentaram estimativas de índice de seleção inferior de MG de 0,33.

66
Para cada um dos locais, as maiores estimativas de herdabilidade em sentido
amplo, foram obtidas para AP, AE, FM, FF, NFE, FF, MG, PROL, CE e NFE em
Campinas (Tabela 16) e para AP, AE, A+Q, FM, FF, NRP, SG e NFE em Mococa
(Tabela 17), indicando maior variabilidade genética a ser explorada pela seleção destes
caracteres. Estas herdabilidades consideradas altas para cada caráter, acima citado,
associadas com a variabilidade genética da população indicam eficiência da seleção
para essas características, o que proporcionará ganhos satisfatórios, em cada um dos
locais. Essas estimativas de herdabilidade foram superiores às obtidas por EDMEADES
et al. (1997a), em estudo de seleção recorrente de progênies S1, com 57% no ambiente
sem estresse e 43% no ambiente com estresse. Estimativas de herdabilidade de trabalhos
diferentes são difíceis de serem comparadas visto que elas tendem a variar, dentre
outros, com o tipo de progênie, o número de repetições e número de ambientes
utilizados na estimação.
Para massa de grãos (MG) a herdabilidade foi de 80% em Campinas e de 63% em
Mococa. Este resultado foi semelhante ao encontrado por RIBAULT et al. (1997), de 61
a 74% para produção de grão, em experimento sob condições de estresse intermediário e
severo estresse, respectivamente. BÉTRAN et al. (1997) e BOLAÑOS & EDMEADES
(1997) encontraram resultados similares (h2
> 50%), avaliando linhagens de milho
derivadas do CIMMYT para tolerância à seca. GAMA et al. (2001) obtiveram valores
de herdabilidade em sentido amplo para peso de espigas em condições de estresse
hídrico e de não estresse, de 38% e 75% para progênies “Synflint” de irmãos germanos.
SILVA et al. (2008) observaram nas progênies de meios-irmãos da UFVM100,
estimativas de herdabilidade em nível de médias ( 2ˆmh ) de 34,8% e 56,2%, nos
experimentos conduzidos sob déficit hídrico e irrigação normal, respectivamente.
CAIRNS et al. (2013) encontraram para produtividade estimativas de herdabilidade em
sentido amplo que variaram de 64% a 84% em condições irrigadas e 47% a 80% em
déficit hídrico.
Maiores estimativas da relação CVg/CVe e da herdabilidade, além da variância
genética aditiva, indicam ser essas as características alvo para seleção direta, pois
proporcionaria estimativas mais precisas de ganho nas progênies, em todos locais
avaliados. Entretanto, é importante também conhecer a relação entre os demais
caracteres de importância agronômica, já que se espera ao final do processo seletivo
para obter nova população.

67
O ganho de seleção para MG foi de 13% e 5%, em Campinas e Mococa,
respectivamente. A diferença no ganho de seleção está relacionada com o ambiente,
onde Mococa teve a maior influência sobre a variância de progênies, reduzindo a
herdabilidade e o ganho. Em média os ganhos para todos os caracteres foram adequados
demonstrando potencial das progênies visando tolerância à seca. Com destaque do
ganho de seleção para A+Q, PROL, NRP, MG e NFE, em Campinas (tabela 16) e para
A+Q, IF, SG e NFE, em Mococa (tabela 17). O ganho genético é um parâmetro de
eficiência na seleção que relaciona com a variabilidade genética, variâncias aditiva e
dominante, herdabilidade e intensidade de seleção.
EDMEADES et al. (1999) obtiveram ganhos com a seleção direta para MG, nas
populações La Posta Sequía, Pool 26 Sequía e Tuxpeño Sequía, de 12,4%, 12,7% e
3,8% respectivamente, em ambiente de estresse hídrico. No ambiente sem estresse
hídrico, os percentuais de ganho foram de 0,6%, 2,3% e 0,5%, respectivamente.
MONNEVEUX et al. (2006) obtiveram ganhos com a seleção para MG em resposta
correlacionada à seleção recorrente nas populações DTP1 e DTP2. Neste caráter o
ganho direto foi de 14,3% e 4% nas DTP1 e DTP2, respectivamente. Os autores
concluíram que nestas populações, os ganhos de produtividade significativos
observados em condições de seca foram associados a um aumento significativo do
número de espigas por planta e grãos por espiga, reduções significativas do IF, do
número de óvulos e taxa de abortamento.

68
Tabela 16. Estimativas da variância fenotípica (2
)2,1(ˆ
F ), da variância genética entre progênies ( 2
)2,1(ˆ
p ), da variância aditiva ( )2,1(
2ˆA ), da
variância do erro (2
)2,1(ˆ
E ), da herdabilidade em sentido amplo ( 2
)2,1(ˆ
xh ), do coeficiente de variação genética (gVC ˆ ) e experimental ( eVC ˆ ),
índice de seleção (b) e ganho de seleção ( )2,1(ˆsG ) para doze caracteres avaliados em 196 progênies interpopulacionais, em Campinas na Safrinha
2012.
Parâmetro
s
genéticos ª
AP
(cm)
AE
(cm)
A+Q
(%)
FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
NRP
(ram.
pend.-1
)
SG
(nota)
MG
(g.pl.-1
)
PROL
(espiga.
plant.-1
)
CE
(cm)
NFE
(n.f.e)
2
)2,1(ˆ
F 256,0
1 237,73 12,72 4,73 7,29 0,085 0,143 0,024 394,54 0,037 1,59 2,65
2
)2,1(ˆ
p
DP
198,8
5 217,86 6,85 4,11 5,51 0,047 0,073 0,013 263,13 0,026 1,27 2,48
±1,10 ±1,00 ±0,06 ±0,02 ±0,03 ±3,9.10-4
±6,7.10-4
±1,1.10-4
±2,42 ±1,6.10-4
±6,8.10-3
±0,01
)2,1(
2ˆA 795,4 871,44 27,5 16,44 22,04 0,188 0,292 0,052 1052,52 0,104 5,08 9,92
2
)2,1(ˆ
E 57,15 19,87 5,86 0,62 1,78 0,038 0,07 0,011 131,40 0,011 0,323 0,16
2
)2,1(ˆ
xh (%)
DP
87,43 95,63 70,04 92,98 86,09 71,21 67,59 70,27 80,02 82,54 88,75 96,87
±0,02 ±0,006 ±0,04 ±0,01 ±0,02 ±0,04 ±0,05 ±0,04 ±0,03 ±0,02 ±0,02 ±0,005
gVC ˆ (%) 5,75 9,81 23,98 2,96 3,29 8,67 3,92 6,86 11,66 18,53 7,86 11,01
eVC ˆ (%) 3,08 2,96 22,18 1,14 1,87 7,79 1,71 6,32 8,24 12,05 3,96 2,79
b 1,86 3,31 1,08 2,57 1,76 1,11 2,29 1,08 1,41 1,54 1,98 3,94
)2,1(ˆsG 17,39 19,28 2,69 2,12 2,85 0,226 2,70 0,117 18,55 0,189 1,41 2,13
)2,1(ˆsG (%) 7,09 12,81 24,65 3,08 4,01 9,03 17,51 7,07 13,33 21,75 9,85 14,89
ªAP, AE, A+Q, FM, FF, IF, NRP, SG, MG, PROL, CE e NFE referem a altura de planta, altura de espiga, porcentagem de plantas acamadas e quebradas, florescimento
masculino, florescimento feminino, intervalo entre florescimento feminino e masculino, número de ramificações de pendão, stay-green, massa de grãos, prolificidade,
comprimento de espiga e número de fileiras de grãos na espiga, respectivamente. DP: desvio-padrão.

69
Tabela 17. Estimativas da variância fenotípica (2
)2,1(ˆ
F ), da variância genética entre progênies ( 2
)2,1(ˆ
p ), da variância aditiva ( )2,1(
2ˆA ), da
variância do erro (2
)2,1(ˆ
E ), da herdabilidade em sentido amplo ( 2
)2,1(ˆ
xh ), do coeficiente de variação genética (gVC ˆ ) e experimental ( eVC ˆ ),
índice de seleção (b) e ganho de seleção ( )2,1(ˆsG ) para doze caracteres avaliados em 196 progênies interpopulacionais, em Mococa na Safrinha
2012.
Parâmetros
genéticos ª
AP
(cm)
AE
(cm)
A+Q
(%)
FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
NRP
(ram.
pend.-1
)
SG
(nota)
MG
(g .pl.-1
)
PROL
(espiga.
plant.-1
)
CE
(cm)
NFE
(n.f.e)
2
)2,1(ˆ
F 215,86 130,10 23,34 3,31 4,15 0,13 0,205 0,016 94,86 0,003 0,78 1,89
2
)2,1(ˆ
p
DP
183,71 97,85 19,50 2,59 3,05 0,072 0,165 0,012 43,74 0,001 0,145 1,65
±0,63 ±0,56 ±0,09 ±0,01 ±0,02 ±6.10-4
±8,7.10-4
±7,2.10-5
±0,46 ±1,8.10-5
±0,004 ±0,008
)2,1(
2ˆA 734,84 379,4 78,0 10,36 12,2 0,288 0,66 0,048 194,96 0,004 0,58 6,6
2
)2,1(ˆ
E 32,15 32,25 3,84 0,72 1,1 0,058 0,04 4,5.10-3
51,12 2,5.10-3
0,635 0,24
2
)2,1(ˆ
xh (%)
DP
91,95 85,85 91,04 87,79 84,72 71,28 89,19 84,21 63,12 44,44 31,35 93,22
±0,01 ±0,02 ±0,13 0,02± ±0,02 ±0,04 ±0,02 ±0,02 ±0,05 ±0,08 ±0,1 ±0,01
gVC ˆ (%) 5,60 7,33 38,60 2,32 2,45 12,77 2,64 7,40 5,65 3,16 2,26 8,75
eVC ˆ (%) 2,34 4,21 17,13 1,22 1,47 11,47 1,30 4,53 6,11 5,00 4,79 3,34
b 2,39 1,74 2,25 1,90 1,66 1,11 2,03 1,63 0,92 0,63 0,47 2,62
)2,1(ˆsG 17,51 12,01 5,65 1,99 2,09 0,28 0,51 0,13 6,29 0,026 0,22 1,68
)2,1(ˆsG (%) 7,24 8,90 49,41 2,87 2,94 13,33 3,32 8,98 5,37 2,6 1,34 11,48
ªAP, AE, A+Q, FM, FF, IF, NRP, SG, MG, PROL, CE e NFE referem a altura de planta, altura de espiga, porcentagem de plantas acamadas e quebradas, florescimento
masculino, florescimento feminino, intervalo entre florescimento feminino e masculino, número de ramificações de pendão, stay-green, massa de grãos, prolificidade,
comprimento de espiga e número de fileiras de grãos na espiga, respectivamente. DP: desvio-padrão.

70
4.2 Experimento de avaliação das progênies interpopulacionais em casa de
vegetação
4.2.1 Análises de variâncias e médias
As análises de variância (Tabelas 18 e 19) apresentaram diferenças altamente
significativas (p<0,01) para genótipos e tratamentos hídricos, em todos os parâmetros
fisiológicos e biométricos (exceto para gs(2)
, AE e PROL no efeito de tratamento
hídrico), evidenciando, respectivamente, presença de variabilidade genética das
progênies e variação dos tratamentos hídricos. Na interação tratamentos hídricos versus
genótipos foram detectadas diferenças significativas, para w e AP, indicando
desempenhos diferenciais dos genótipos nos tratamentos hídricos.
A massa de grãos (MG) das progênies e testemunhas foi substancialmente
reduzida em condições de déficit hídrico (Tabela 20). A média geral de MG foi de
232,96 g planta-1
com amplitude de variação de 196,18 g planta-1
a 276,33 g planta-1
no
tratamento irrigado, e em déficit hídrico a média geral foi de 149,78 g planta-1
, com
amplitude de variação de 113,29 g planta-1
a 184,44 g planta-1
. Pelo teste Scott-Knott as
médias dos genótipos foram agrupadas em três grupos (a, b, c) nas condições irrigadas e
dois grupos (a, b) nas condições de déficit hídrico, enquanto os tratamentos hídricos
foram agrupados em dois grupos (A, B). A redução da massa de grãos do tratamento de
déficit hídrico em relação ao irrigado representou amplitude de variação de 22,9% (IAC
Maya Latente) a 52,4% (PI(1,2)-109), com redução média de 35,7% (Tabela 20). A queda
de produção ocorre pela ocorrência da competição fonte-dreno, resultando em falhas na
reprodução e também falhas nas pontas das espigas devido ao aborto dos óvulos, tendo
em vista que o enchimento ocorre da base para as pontas. A progênie PI(1,2)-59
destacou-se pelo elevado valor de MG no tratamento irrigado, e em déficit hídrico a MG
foi satisfatória representando redução de apenas 30%. Destacaram também as progênies
PI(1,2)-49, PI(1,2)-72, PI(1,2)-147 e o híbrido DAS2B707 pelas menores reduções de MG
em déficit hídrico com 30,7, 29,1, 30,7 e 27,9%, respectivamente (Tabela 20).
GUIMARÃES (2013) avaliou a MG de progênies de irmãos germanos em casa de
vegetação sob déficit hídrico no estádio de florescimento, e encontrou reduções
significativas de 89% a 28%. BOLAÑOS & EDMEADES (1996) analisaram o
desempenho de progênies S1, obtendo média de produtividade de grãos (PG) de 2,49,
0,95 e 0,35 ton ha-1
em condições irrigadas, com estresse intermediário e estresse

71
severo, respectivamente. Segundo esses autores, o déficit hídrico imposto reduziu a PG
em 14% a 38%. SILVA et al. (2008), encontraram valores médios para PG em famílias
de meios-irmãos da UFVM 100, de 5,6 ton ha-1
em irrigação normal e, em déficit
hídrico de 3,4 ton ha-1
. A redução da PG do experimento irrigado, em relação ao ensaio
de déficit hídrico, foi de 40%. Segundo BÄNZINGER et al. (2000) em estudos de nível
de déficit hídrico é desejável que seja suficiente para reduzir a produção de grãos em, no
mínimo, 40% em relação ao ambiente sem estresse hídrico.
As progênies interpopulacionais tolerantes (PI(1,2)-49, PI(1,2)-59, PI(1,2)-72, PI(1,2)-
100, PI(1,2)-117 e PI(1,2)-147) e as testemunhas (IAC Maya Latente e DAS2B707),
diferiram estatisticamente da Progênie Sensível para MG, em déficit hídrico (Tabela
20). As progênies e as testemunhas acima listadas, também apresentaram alto valor de
índice relativo de tolerância (RVT), indicando possuir potencial produtivo mesmo em
déficit hídrico. O IAC Maya Latente é uma variedade que não apresentou uma melhor
performance em condições irrigadas, porém seu desempenho produtivo em condições
de déficit hídrico se manteve constante (alto valor de RVT). Por definição a variedade
IAC Maya Latente é tolerante à seca, mesmo tendo um baixo potencial produtivo.
Aparentemente, o IAC Maya Latente possui características adaptativas que não existem
na Progênie Sensível, que permite assegurar a produtividade tanto em condições
estressantes e não estressantes. DUVICK (1995) propõe que o principal objetivo dos
programas de melhoramento de milho tropical deve ser a estabilidade de produção dos
genótipos e a ampla adaptação dos mesmos através do aumento da tolerância para
vários estresses.
Condições de seca levam a menor ocorrência do aparecimento de polens e
também da diminuição dos filamentos dos etilos-estigmas, que pode resultar na redução
do número de grãos na espiga e peso de espigas. Portanto, a variação das progênies
interpopulacionais em relação à massa de grãos (MG), pode ser atribuída a alterações de
muitos processos metabólicos e fisiológicos em plantas durante o déficit hídrico (IZGE
& DUGJE, 2011).

72
Tabela 18. Resumo da análise de variância para potencial de água na folha ( w ), clorofila total (Chl), condutância estomática (gs(1)
)(horário de
9:00 a 10:30h), condutância estomática (gs(2)
)(horário de 11:00 a 12:30h), altura de planta (AP), altura de espiga (AE), florescimento masculino
(FM) e florescimento feminino (FF) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707, IAC IAC Maya Latente e progênie sensível
à seca – PI(1,2)-141), submetidas a dois manejos de irrigação, irrigação normal e déficit hídrico, no estádio de florescimento. Campinas, 2013.
Fontes de variação
Quadrados médios
GL w
(MPa)
Chl
(SPAD)
gs(1)
(mmol m
-2
s-1
)
gs(2)
(mmol m
-2
s-1
)
AP
(cm)
AE
(cm)
FM
(d.a.s.)
FF
(d.a.s.)
Blocos 1 0,027 39,06 0,06 195,17 0,56 0,35 8,20 13,09
Tratamentos hídricos (TH) 1 0,020** 11,52** 253,91* 65,46ns
213,84** 3,27ns
0,56** 1,45**
Erro a 1 0,001 8,204 336 108,23 46,02 15,36 0,56 1,45
Genótipos (G) 10 1,226** 1373,4** 5,78** 19125,8** 60570,2** 17557,1** 2629,2** 2766,2**
TH x G 10 0,009* 5,21ns
1,59ns
47,19ns
139,74** 3,27ns
2,36ns
2,45ns
Erro b 20 0,003 13,85 607 98,06 24,94 10,62 3,08 4,17
Total 43 - - - - - - - -
Média geral -1,12 44,72 112,65 211,45 296 167 63,5 65,5
CV (%) Erro a 2,82 6,40 16,27 4,92 2,29 2,34 1,18 1,84
CV (%) Erro b 4,89 8,32 21,87 4,68 1,68 1,95 2,76 3,12 ns
,* e
**: não significativo, significativo a 5% e 1%, respectivamente, pelo teste F;
(1) dados transformados para 2,0x .

73
Tabela 19. Resumo da análise de variância para intervalo entre florescimento (IF), stay-green (SG), massa de grãos (MG), prolificidade (PROL),
matéria seca total (MST) e índice de colheita (IC) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707, IAC IAC Maya Latente e
progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de irrigação, irrigação normal e déficit hídrico, no estádio de florescimento.
Campinas, 2013.
Fontes de variação
Quadrados médios
GL IF
(1)
(dias)
SG(2)
(nota)
MG
(g planta-1
)
PROL
(espiga
planta-1
)
MST
(g planta-1
) IC
(3)
Blocos 1 0,049 0,004 1104,86 0,065 144,24 2.10-5
Tratamentos hídricos (TH) 1 0,015** 0,004** 444,48** 0,006ns
941,56** 1,7.10-3
**
Erro a 1 0,015 0,004 5,01 0,012 44,72 1,5.10-3
Genótipos (G) 10 1,671** 0,999** 24129,78** 1,230** 72935,39** 0,089**
TH x G 10 0,007ns
0,002ns
207,31ns
0,003ns
424,35ns
9.10-4ns
Erro b 20 0,019 0,005 376,86 0,019 255,64 4.10-4
Total 43 - - - - - -
Média geral 1,95 1,8 191,37 1,27 343,08 0,375
CV (%) Erro a 6,41 3,51 1,16 8,62 1,95 10,33
CV (%) Erro b 7,08 3,93 10,14 10,85 4,66 5,33 (1,2)
dados transformados para x ; (3)
IC: peso seco de grãos/ (peso seco de grãos + peso total da planta) ns
,* e
**: não significativo, significativo a 5% e 1%, respectivamente,
pelo teste F.

74
Tabela 20. Média de massa de grãos (MG) e índice relativo de tolerância (RVT) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707,
IAC IAC Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de irrigação, irrigação normal e déficit hídrico, no
estádio de florescimento (VT). Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico
MG
(g planta-1
)
MG(1)
(%)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico Redução
(2)
(%)
RVT(3)
Irrigado Déficit
hídrico
PI(1,2)-49 241,314 bA 167,319 aB 30,7 0,69 123,0 110,7
PI(1,2)-52 212,392 cA 126,317 bB 40,5 0,59 108,3 83,6
PI(1,2) -59 263,616 aA 184,447 aB 30,0 0,69 134,4 122,0
PI(1,2) -72 221,734 cA 157,276 aB 29,1 0,71 113,0 104,1
PI(1,2) -100 209,799 cA 143,207 aB 31,7 0,68 106,9 94,7
PI(1,2) -109 237,838 bA 113,291 bB 52,4 0,47 121,2 74,9
PI(1,2) -117 276,334 aA 155,057 aB 43,9 0,56 140,8 102,6
PI(1,2) -147 228,923 cA 158,642 aB 30,7 0,69 116,7 104,9
Média progênies 236,493 150,694 36,3 0,63 120,5 99,7
IAC Maya Latente 196,184 cA 151,116 aA 22,9 0,77 100,0 100,0
DAS2B707 242,946 bA 175,000 aB 27,9 0,72 123,8 115,8
Progênie Sensível 231,517 cA 116,001 bB 49,9 0,50 118,0 76,7
Média geral 232,963 149,788 35,7 0,63 118,4 99,1 (1)
: porcentagem em relação à testemunha IAC Maya Latente (em g planta-1
); (2)
Redução da MG do tratamento de déficit hídrico em relação ao irrigado; (3)
RVT: (déficit
hídrico/irrigado);. Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo teste Scott-Knott entre os genótipos na mesma condição hídrica e
letras maiúsculas distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.

75
Tendo em vista que a translocação de carboidratos é menos sensível que a fotossíntese e
a respiração (KRAMER & BOYER, 1995), a habilidade de uma cultivar quanto ao
armazenamento e mobilização de reservas no caule, para posterior translocação para os grãos
pode ser uma característica de tolerância à seca (BLUM, 2010). Os valores de massa seca
total (MST) estão mostrados na Tabela 21. Observou-se que o déficit hídrico influenciou esse
caráter e revelou diferenças significativas entre os genótipos, as quais foram agrupadas em
quatro grupos distintos pelo teste Scott-Knott (P≤0,05). O valor médio de MST, em condição
irrigada, foi de 390,84 g planta-1
, com amplitude de variação de 313,15 g planta-1
a 458,32 g
planta-1
, e em déficit hídrico a média geral foi de 295,32 g planta-1
, com amplitude de variação
de 252,61 g planta-1
a 381,94 g planta-1
. No geral, as progênies PI(1,2) -59, PI(1,2) -117 e PI(1,2) -
147 e as testemunhas tolerantes IAC Maya Latente e DAS2B707 tiveram maiores valores de
RVT, indicando que a biomassa se manteve constante nos dois tratamentos hídricos.
GUIMARÃES (2013) obteve para MST na Pop.Tol.1, valor médio de 337,14 g planta-1
em
condição irrigada e 327,43 g planta-1
em déficit hídrico, com redução de apenas 2,8%.
BARNABY et al. (2013) detectaram uma redução de 53% de MST em híbridos de milho
tolerantes em relação aos híbridos intermediários e suscetíveis. Estes autores ainda
demonstraram que na seleção de híbridos em condição extremas de déficit hídrico, as plantas
possuem menor altura de plantas devido aos menores potenciais de água no solo e na folha.
Para o índice de colheita (IC) a progênie PI(1,2) -147 e IAC Maya Latente, destacaram
pela maior alocação diferencial de fotoassimilados para a espiga durante seu ciclo de vida
(Tabela 21). Pelos valores de RVT do IC foi confirmado a maior tolerância das progênies
PInter-49, PInter-72, P-Inter147 e do IAC IAC Maya Latente e DAS2B707. HUSSAIN
(2009) encontrou para IC valores de 0,36 a 0,67 em condições irrigadas e de 0,32 a 0,51 sob
déficit híbrido em linhagens de milho.
A imposição do déficit hídrico proporcionou aumentos do intervalo entre florescimentos
feminino e masculino (IF) e stay-green (SG), e reduções da prolificidade (PROL) e altura de
planta (AP) (Tabelas 21, 22, 23 e 24). Em condições irrigadas, a média geral para IF, SG,
PROL e AP foram, respectivamente, de 1,8 dias, nota de 1,5, 1,4 espiga planta-1
e 304 cm.
Para condições de déficit hídrico, as médias destes caracteres foram, respectivamente, de 2,1
dias, nota de 2,08, 1,12 espiga planta-1
e 289 cm. No geral, as seguintes progênies PI(1,2) -59 e
PI(1,2) -117 destacaram-se pelos menores intervalos entre florescimentos. Sendo para médias
de genótipos do caráter IF o teste Scott-Knott, agrupou em quatro grupos (a, b, c, d) nos dois
tratamentos hídricos, e para tratamentos hídricos o teste agrupou em dois grupos (A, B)

76
(Tabela 24). De acordo com ZAIDI et al. (2008), os valores de IF menores que cinco dias
podem identificar linhagens tolerantes a estresses, seja por déficit ou excesso de água. Para o
caráter PROL foi observado a formação de dois grupos (a, b) para genótipos e, também para
tratamentos hídricos (Tabela 21). Para PROL, as PI(1,2) -49 e PI(1,2) -117 obtiveram maiores
valores desse caráter com 1,38 e 1,28 espiga planta-1
, respectivamente. Enquanto para as PI(1,2)
-49 e PI(1,2) -147 e o DAS2B707, destacaram-se para stay-green acentuado (menores notas de
SG de 1,6, 1,4 e 1,5) e menores alturas de planta (278, 271 e 246 cm), respectivamente. Para
SG as médias de genótipos foram agrupadas em dois grupos (a, b) na condição irrigada, e de
três grupos (a, b, c) na condição de déficit hídrico (Tabela 24). Enquanto para AP o teste
Scott-Knott separou genótipos em quatro grupos (a, b, c, d) nos dois tratamentos hídricos e,
para tratamentos hídricos o teste agrupou em dois grupos (A, B) (Tabela 22). Para ALI et al.
(2011) a redução na estatura das plantas é devido a adaptação das mesmas para enfrentar o
déficit hídrico. Com os efeitos iniciais do déficit hídrico, as plantas de milho passam a desviar
os assimilados do colmo e utilizá-los para maior crescimento das raízes, a fim de aumentar a
absorção de água.
CAIRNS et al. (2013) encontraram para IF valores que variaram de 1,79 dias na
localidade do Kenya a 5,11 dias na Tailândia, sob déficit hídrico, e 2,64 dias a 12,01 dias no
México, sob déficit hídrico e altas temperaturas. Para altura de plantas, os autores
encontraram reduções de 38,2 e 19,2 cm, em dois experimentos sob déficit hídrico e altas
temperaturas, em híbridos simples de milho. Para DURÃES et al. (2004) a redução da altura
de plantas sob déficit hídrico é consequência da diminuição da abertura de estômatos e,
consequentemente, da absorção de CO2 afetando assim negativamente a taxa de fotossíntese.

77
Tabela 21. Média de prolificidade (PROL), matéria seca total (MST), índice de colheita (IC) e índice relativo de tolerância (RVT) de 8 progênies
interpopulacionais e 3 testemunhas (DAS2B707, IAC IAC Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de
irrigação, irrigação normal e déficit hídrico, no estádio de florescimento (VT). Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico
PROL
(espiga planta-1
)
MST
(g planta-1
) IC
(1)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico Irrigado Déficit hídrico RVT(2)
Irrigado Déficit
hídrico RVT
(2)
PI(1,2) -49 1,485 aA 1,384 aA 354,830 cA 252,611 dB 0,71 0,440 aA 0,358 aB 0,81
PI(1,2) -52 1,393 aA 1,208 aA 403,907 bA 305,304 bB 0,75 0,396 aA 0,263 aB 0,66
PI(1,2) -59 1,526 aA 1,117 bB 368,235 cA 325,638 bA 0,88 0,455 aA 0,354 aB 0,78
PI(1,2) -72 1,031 bA 1,275 aA 450,539 aA 279,605 cB 0,62 0,417 aA 0,346 aB 0,83
PI(1,2) -100 1,151 bA 1,003 bA 420,082 bA 290,940 cB 0,69 0,416 aA 0,303 aB 0,73
PI(1,2) -109 1,603 aA 1,000 bB 390,748 cA 232,609 dB 0,59 0,329 aA 0,245 aB 0,74
PI(1,2) -117 1,459 aA 1,285 aA 458,323 aA 381,941 aA 0,83 0,402 aA 0,285 aB 0,71
PI(1,2) -147 1,708 aA 0,940 bB 369,034 cA 300,762 bB 0,81 0,424 aA 0,427 aA 1,00
Média progênies 1,419 1,151 401,962 296,176 0,74 0,409 0,322 0,79
IAC Maya Latente 1,381 aA 1,000 bB 410,390 bA 356,606 aB 0,86 0,356 aA 0,336 aA 0,94
DAS2B707 1,409 aA 1,191 aA 313,150 dA 261,512 dB 0,83 0,526 aA 0,424 aB 0,81
Progênie Sensível 1,425 aA 1,000 bA 360,023 cA 260,985 dB 0,72 0,446 aA 0,308 aB 0,69
Média geral 1,415 1,127 390,841 295,319 0,75 0,418 0,332 0,79 (1)
IC: peso seco de grãos/ (peso seco de grãos + peso seco da planta). (2)
RVT: (déficit hídrico/irrigado). Letras minúsculas distintas nas colunas indicam diferenças estatísticas
(P<0,05) pelo teste Scott-Knott entre os genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas representam diferenças estatísticas entre os tratamentos
hídricos.

78
Tabela 22. Média de altura de planta (AP), altura de espiga (AE) e índice relativo de tolerância
(RVT) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707, IAC IAC Maya Latente e
progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de irrigação, irrigação normal
e déficit hídrico, no estádio de florescimento (VT), em Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico(1)
AP (cm) AE (cm)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico RVT(2) Irrigado Déficit hídrico RVT(2)
PI(1,2) -49 305,5 cA 278,5 bB 0,91 169,5 cA 160,0 cB 0,94
PI(1,2) -52 315,0 bA 317,5 aA 1,01 163,5 dA 161,0 cA 0,98
PI(1,2) -59 296,5 cA 283,0 bB 0,95 179,0 bA 171,0 bA 0,95
PI(1,2) -72 312,0 bA 301,5 aB 0,96 174,0 bB 180,0 aA 1,03
PI(1,2) -100 327,0 aA 287,5 bB 0,88 167,0 cA 167,5 bA 1,00
PI(1,2) -109 296,5 cA 268,5 cB 0,90 168,0 cA 160,0 cB 0,95
PI(1,2) -117 314,0 bA 304,5 aA 0,97 178,5 bA 172,0 bA 0,96
PI(1,2) -147 290,0 cA 271,0 cB 0,93 184,0 aA 128,0 dB 0,69
Média progênies 307,0 288,0 0,94 173,0 164,0 0,95
IAC Maya Latente 326,0 aA 316,0 aA 0,97 176,0 bA 162,0 cA 0,92
DAS2B707 252,5 dA 246,0 dA 0,97 157,0 dA 129,0 dB 0,82
Progênie Sensível 315,5 bA 309,0 aA 0,98 173,5 bB 182,0 aA 1,04
Média geral 304,0 289,0 0,95 170,0 165,0 0,97
RVT: (déficit hídrico/irrigado). Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05)
pelo teste Scott-Knott entre os genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas
representam diferenças estatísticas entre os tratamentos hídricos.

79
Tabela 23. Média de florescimento masculino (FM), florescimento feminino (FF) e índice
relativo de tolerância (RVT) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707,
IAC IAC Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de
irrigação, irrigação normal e déficit hídrico, no estádio de florescimento (VT), em Campinas,
2013.
Genótipos
Variáveis/ Tratamento hídrico(1)
FM (d.a.s.) FF (d.a.s.)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico RVT(2) Irrigado Déficit hídrico RVT(2)
PI(1,2) -49 65,0 aA 63,0 aA 0,97 66,5 aA 65,5 bA 0,98
PI(1,2) -52 60,5 bB 65,0 aA 1,07 61,5 bB 66,5 bA 1,08
PI(1,2) -59 62,0 bA 65,0 aA 1,05 64,5 bA 66,0 bA 1,02
PI(1,2) -72 61,0 bB 66,0 aA 1,08 62,0 bB 68,5 bA 1,10
PI(1,2) -100 62,0 bB 68,0 aA 1,10 64,0 bB 72,5 aA 1,13
PI(1,2)-109 61,0 bA 57,0 bB 0,93 61,5 bA 58,5 cA 0,95
PI(1,2) -117 65,5 aA 64,5 aA 0,98 66,0 aA 65,0 bA 0,98
PI(1,2) -147 63,5 aA 64,5 aA 1,01 67,0 aA 66,5 bA 0,99
Média progênies 62,6 64,1 1,02 64,1 66,1 1,03
IAC Maya Latente 65,5 aA 65,0 aA 0,99 69,0 aA 68,0 bA 0,98
DAS2B707 61,5 bA 63,5 aA 1,03 63,5 bA 65,5 bA 1,03
Progênie Sensível 65,0 aA 64,0 aA 0,98 66,5 aA 66,5 bA 1,00
Média geral 63,0 64,1 1,02 64,7 66,3 1,02
(1)Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo teste Scott-Knott entre os
genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas representam diferenças estatísticas
entre os tratamentos hídricos; (2)
RVT: déficit hídrico/irrigado.

80
Tabela 24. Média de intervalo entre florescimentos feminino e masculino (IF), stay-green
(SG) e índice relativo de tolerância (RVT) de 8 progênies interpopulacional e 3 testemunhas
(DAS2B707, IAC IAC Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a
dois manejos de irrigação, irrigação normal e déficit hídrico iniciado no estádio de
florescimento (VT), em Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico(1)
IF (dias) SG (nota)(2)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico RVT(3) Irrigado Déficit hídrico RVT(3)
PI(1,2) -49 1,5 cA 2,5 bA 1,66 1,2 bA 1,6 cA 1,33
PI(1,2) -52 1,0 dA 1,5 cA 1,50 1,4 bB 2,0 bA 1,43
PI(1,2) -59 2,5 bA 1,0 dB 0,40 1,8 aA 2,0 bA 1,11
PI(1,2) -72 1,0 dB 2,5 bA 2,50 1,6 aB 2,7 aA 1,70
PI(1,2) -100 2,0 cB 4,5 aA 2,25 1,6 aB 3,0 aA 1,87
PI(1,2) -109 0,5 dA 1,5 cA 3,00 2,0 aB 2,6 aA 1,30
PI(1,2) -117 0,5 dA 0,5 dA 1,00 1,9 aA 2,1 bA 1,10
PI(1,2) -147 3,5 aA 2,0 cB 0,57 1,4 bA 1,4 cA 1,00
Média progênies 1,6 2,0 1,28 1,6 2,2 1,35
Maya Latente 3,5 aA 3,0 bA 0,85 1,2 bA 1,4 cA 1,16
DAS2B707 2,0 cA 2,0 cA 1,00 1,4 bA 1,5 cA 1,07
Progênie Sensível 1,5 cA 2,5 bA 1,66 1,0 bB 2,6 aA 2,60
Média geral 1,8 2,1 1,21 1,5 2,08 1,39
(1)Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo teste Scott-Knott entre os
genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas representam diferenças estatísticas
entre os tratamentos hídricos; (2)
dados transformados para x ; (3)
RVT: déficit hídrico/irrigado.

81
O potencial hídrico é uma medida do grau de hidratação de uma planta e, assim, fornece
um índice relativo do estresse hídrico ao qual a planta está submetida. Esse parâmetro
fisiológico tem recebido merecida atenção como mecanismo de adaptação de plantas, para
manutenção do turgor celular e de funções fisiológicas. Os valores médios de potencial de
água na folha ( w ) foram, respectivamente, de -0,55 MPa em condições irrigadas, e de -1,69
MPa em déficit hídrico (Tabela 25). O teste Scott-Knott (P≤0,05) separou a média dos
genótipos em dois grupos (a, b) em condição irrigada e em quatro grupos (a, b, c, d) em
condição de déficit hídrico. Os resultados de w discriminaram apenas as PI(1,2)-59 e PI(1,2)-
100 pelos valores menos negativos de -1,42 e -1,45 MPa, respectivamente, e confirmou-se o
padrão de sensibilidade da Progênie Sensível com valor mais negativo de -2,02 MPa.
GUTIÉRREZ et al. (1996) encontraram valores semelhantes de w de -1,7 MPa avaliados
em populações de milho Tuxpeño Sequía sob estresse hídrico, e em condições irrigadas, a
média de w foi de -0,38 MPa.
GUIMARÃES et al. (2014) também encontraram para w avaliado em déficit hídrico,
valores de -0,30 (DAS2B707) a -0,99 (DAS2B710); -0,28 (DAS2B710) a -0,60 MPa
(AS1522) e de -0,23 (DAS2B707) a -0,40 MPa (AS1522), respectivamente, para estádio
vegetativo, florescimento e enchimento de grãos. Os autores indicaram que a manutenção do
status hídrico do DAS2B707 foi relacionada ao fechamento parcial dos estômatos e aumento
da densidade e profundidade das raízes, garantindo maior exploração do solo.
A condutância estomática mede a taxa de passagem de dióxido de carbono que sai, ou o
vapor de água que entra através dos estômatos. A taxa de condutância estomática, ou seu
inverso, resistência estomática, é diretamente relacionada com a resistência da camada
limítrofe da folha e gradiente de concentração absoluto de vapor de água da folha para a
atmosfera. A cutícula que recobre as superfícies expostas das plantas serve de barreira efetiva
à perda de água, protegendo assim, a planta contra a dessecação. A condutância estomática
frequentemente está relacionada com o status hídrico da folha. O porômetro é um aparelho
capaz de estimar a condutância foliar (mmol m-2
s-1
) à difusão gasosa, aumentando o
entendimento do processo transpiratório da planta.
Em relação à condutância estomática gs(1)
e (2)
, medida nos horários de 9:00 a 10:30h e
11:00 a 12:30h (Tabela 26), as médias foram, respectivamente, de 156,7 e 332,2 mmol m-2
s-1
,
em condições irrigadas, e de 68,6 e 90,6 mmol m-2
s-1
, em déficit hídrico. O gs(1)
foi separado
em quatro grupos distintos (a, b, c, d) em condição irrigada, e em dois grupos distintos (a, b)

82
em déficit hídrico. Enquanto que as médias de genótipos de gs(2)
foram separadas cinco grupos
distintos em condição irrigada e em três grupos em condição de déficit hídrico. Neste trabalho
foi observado que o horário de alta demanda evaporativa de condutância estomática gs(2)
pode
discriminar melhor os genótipos, como descrito a seguir. Os genótipos PI(1,2)-147 e
DAS2B707 apresentaram maiores status hídricos e alto valor de condutância estomática (gs(2)
)
em déficit hídrico, indicando que eles não apresentaram fechamento dos estômatos. Esses dois
parâmetros fisiológicos (gs e w ) estão intimamente ligados, porque um maior status hídrico
pode favorecer uma maior condutância estomática, levando a um fluxo de CO2 e resfriamento
da folha pela transpiração (SOUZA, 2012). De acordo com JONES et al.(2009) e MUTAVA
et al. (2008) citados por SOUZA (2012), genótipos tolerantes à seca com alta produtividade
de grãos têm sido identificados pelo resfriamento do dossel e por escaparem da seca, pois
necessitam de maior gasto de água.
GUTIÉRREZ et al. (1996) estudaram três ciclos de seleção recorrente (C0, C4 e C8) de
populações Tuxpeño selecionados para tolerância à seca pelos melhoristas do CIMMYT no
intuito de identificar mudanças da condutância estomática medida ao meio-dia e acúmulo de
ácido abscísico (ABA). Os autores encontraram maiores valores de gs de 96,9 mmol m-2
s-1
para as plantas de C8, e menores para as plantas de C0 e C4 de 71,2 e 64,6 mmol m-2
s-1
,
respectivamente, relacionando os menores valores de gs a reduzida perda de água pela
transpiração, porém não proporcionou em mudanças diretas da concentração de ABA.
Limitada disponibilidade hídrica no solo em geral faz as espécies reduzirem a
condutância estomática, o que leva a menor assimilação de CO2. O fechamento estomático é
uma das respostas mais proeminente resposta à seca, levando à diminuição das trocas gasosas
(FAROOQ et al., 2009), para evitar a desidratação dos tecidos. Esse comportamento
estomático pode estar associado a uma rápida adaptação ao estresse hídrico. SOUZA et al.
(2011) citam que o maior número de estômatos pode favorecer a maior entrada de gases,
diminuindo assim a resistência estomática. O agrupamento de médias separou os genótipos
em dois grupos distintos (a, b) em déficit hídrico, e também para tratamentos hídricos (Tabela
26). Destacaram-se as seguintes progênies: PI(1,2)-49, PI(1,2)-52, PI(1,2)-72 e PI(1,2)-100 e IAC
Maya Latente pelo fechamento parcial dos estômatos em déficit hídrico, indicando possuir
mecanismos de adaptação à seca. GUIMARÃES et al. (2014) relacionaram o híbrido FT510
por apresentar maior valor de gs quando submetido à seca no estádio de enchimento de grãos,
devido a maior fotossíntese e respiração do mesmo.

83
A clorofila total (Chl) demonstrou ser indicativa de área verde foliar em déficit hídrico
possibilitando a identificação das PI(1,2)-49, PI(1,2)-59, PI(1,2)-100, PI(1,2)-109 e PI(1,2)-147, visto
que é uma das responsáveis pela manutenção da taxa fotossintética. Os valores médios de Chl
foram de 54,63 SPAD, em condição irrigada e de 34,81 SPAD em déficit hídrico.
MAGALHÃES et al. (2009) verificaram redução média do índice de SPAD de 42,36 para
35,32 sob deficiência hídrica. Segundo TORRES NETTO et al. (2005), leituras SPAD
inferiores a 40 indicam deficiência de clorofila, o que afeta o processo fotossintético. Com
exceção da PI(1,2)-72, todas as progênies que apresentaram altos teores de clorofila total em
déficit hídrico, também possuíram maiores massas de grãos (Tabela 25). Neste estudo os
genótipos tolerantes à seca apresentaram maiores níveis de clorofila em relação à Progênie
Sensível, sugerindo que este parâmetro é bom indicador fisiológico na seleção de genótipos
tolerantes. O atraso da senescência foliar e da manutenção do conteúdo de clorofila em déficit
hídrico como observado pelas progênies estudadas, podem estar relacionados ao aumento da
produção de carboidratos pela interceptação de radiação com maior absorção da fração de luz
incidente (MUCHOW & SINCLAIR, 1994; ZAIDI et al., 2008).
O medidor de clorofila (portátil) permitiu medições instantâneas do valor
correspondente ao seu teor na folha sem destruí-la, e constitui uma alternativa para estimar o
teor relativo desse pigmento na folha. Os valores são calculados pelo equipamento com base
na quantidade de luz transmitida pela folha, em dois comprimentos de ondas, com diferentes
absorbâncias das clorofilas a e b. Destaca-se pela rapidez, simplicidade e por possibilitar uma
avaliação não destrutiva do tecido foliar.

84
Tabela 25. Média de potencial de água na folha ( w ), clorofila total (Chl) e índice relativo
de tolerância (RVT) de 8 progênies interpopulacionais e 3 testemunhas (DAS2B707, IAC
IAC Maya Latente e progênie sensível à seca – PI(1,2)-141), submetidas a dois manejos de
irrigação, irrigação normal e déficit hídrico iniciado no estádio de florescimento (VT), em
Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico(1)
w (MPa) Chl (SPAD)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico RVT(2) Irrigado Déficit hídrico RVT(2)
PI(1,2)-49 -0,70 bA -1,92 dB 2,75 57,56 aA 36,61 aB 0,63
PI(1,2)-52 -0,47 aA -1,67 bB 3,52 52,25 aA 30,66 bB 0,53
PI(1,2)-59 -0,62 bA -1,42 aB 2,28 56,18 aA 38,75 aB 0,68
PI(1,2)-72 -0,52 aA -1,75 cB 3,33 50,61 aA 28,65 bB 0,56
PI(1,2)-100 -0,52 aA -1,45 aB 2,76 56,24 aA 40,81 aB 0,72
PI(1,2)-109 -0,52 aA -1,77 cB 3,38 62,01 aA 36,63 aB 0,59
PI(1,2)-117 -0,60 bA -1,62 bB 2,71 55,63 aA 33,53 bB 0,60
PI(1,2)-147 -0,55 aA -1,72 cB 3,14 52,49 aA 35,16 aB 0,66
Média progênies -0,56 -1,66 2,96 55,37 35,10 0,63
Maya Latente -0,52 aA -1,67 bB 3,19 50,99 aA 31,65 bB 0,62
DAS2B707 -0,52 aA -1,62 bB 3,09 51,33 aA 40,56 aB 0,79
Progênie Sensível -0,57 aA -2,02 dB 3,52 55,71 aA 29,89 bB 0,53
Média geral -0,55 -1,69 3,07 54,64 34,81 0,64
(1)Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo teste Scott-Knott entre os
genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas representam diferenças estatísticas
entre os tratamentos hídricos; (2)
RVT: déficit hídrico/irrigado.
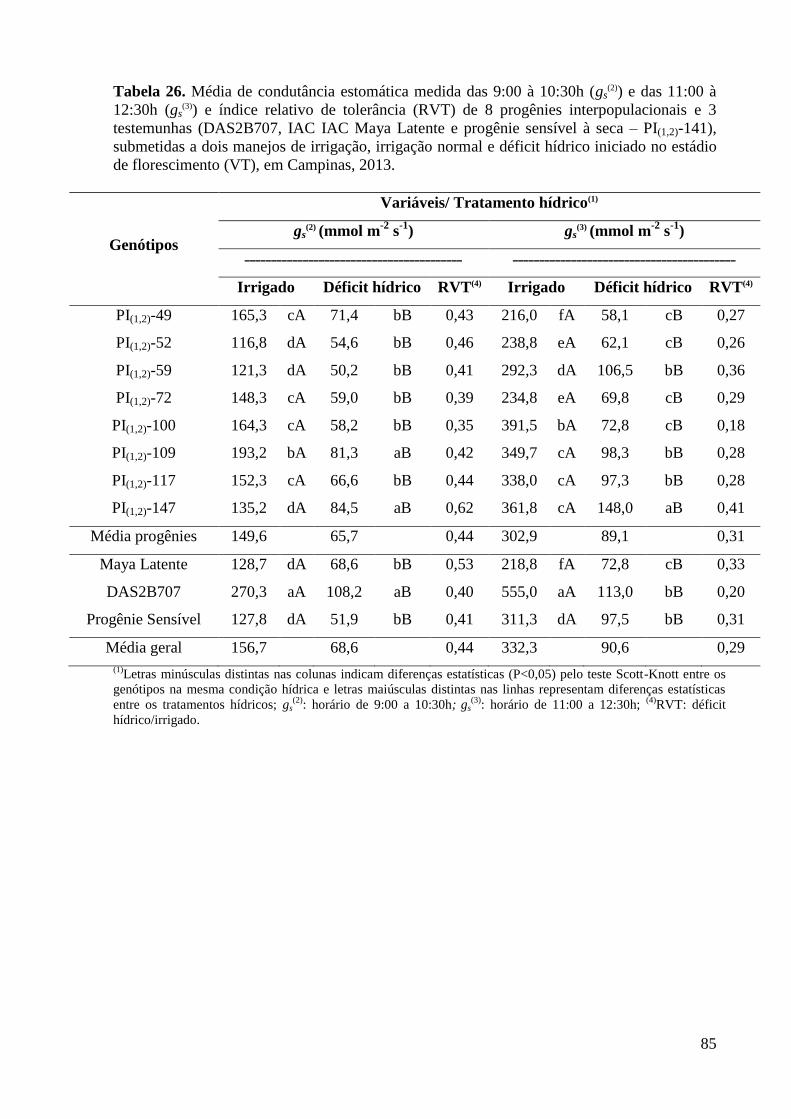
85
Tabela 26. Média de condutância estomática medida das 9:00 à 10:30h (gs(2)) e das 11:00 à
12:30h (gs(3)) e índice relativo de tolerância (RVT) de 8 progênies interpopulacionais e 3
testemunhas (DAS2B707, IAC IAC Maya Latente e progênie sensível à seca – PI(1,2)-141),
submetidas a dois manejos de irrigação, irrigação normal e déficit hídrico iniciado no estádio
de florescimento (VT), em Campinas, 2013.
Genótipos
Variáveis/ Tratamento hídrico(1)
gs(2) (mmol m
-2 s
-1) gs
(3) (mmol m-2
s-1
)
˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗ ˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗˗
Irrigado Déficit hídrico RVT(4) Irrigado Déficit hídrico RVT(4)
PI(1,2)-49 165,3 cA 71,4 bB 0,43 216,0 fA 58,1 cB 0,27
PI(1,2)-52 116,8 dA 54,6 bB 0,46 238,8 eA 62,1 cB 0,26
PI(1,2)-59 121,3 dA 50,2 bB 0,41 292,3 dA 106,5 bB 0,36
PI(1,2)-72 148,3 cA 59,0 bB 0,39 234,8 eA 69,8 cB 0,29
PI(1,2)-100 164,3 cA 58,2 bB 0,35 391,5 bA 72,8 cB 0,18
PI(1,2)-109 193,2 bA 81,3 aB 0,42 349,7 cA 98,3 bB 0,28
PI(1,2)-117 152,3 cA 66,6 bB 0,44 338,0 cA 97,3 bB 0,28
PI(1,2)-147 135,2 dA 84,5 aB 0,62 361,8 cA 148,0 aB 0,41
Média progênies 149,6 65,7 0,44 302,9 89,1 0,31
Maya Latente 128,7 dA 68,6 bB 0,53 218,8 fA 72,8 cB 0,33
DAS2B707 270,3 aA 108,2 aB 0,40 555,0 aA 113,0 bB 0,20
Progênie Sensível 127,8 dA 51,9 bB 0,41 311,3 dA 97,5 bB 0,31
Média geral 156,7 68,6 0,44 332,3 90,6 0,29
(1)Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo teste Scott-Knott entre os
genótipos na mesma condição hídrica e letras maiúsculas distintas nas linhas representam diferenças estatísticas
entre os tratamentos hídricos; gs(2)
: horário de 9:00 a 10:30h; gs
(3): horário de 11:00 a 12:30h;
(4)RVT: déficit
hídrico/irrigado.

86
5 CONCLUSÕES
1. Ressaltam-se progênies interpopulacionais promissoras oriundas do cruzamento da
Pop.Tol.1 com o IAC IAC Maya Latente, nos experimentos de campo em condições de
safrinha, com destaque para menor intervalo entre florescimentos (IF) e maiores valores de
massa de grãos (MG) e componentes de produção (CE e NFE), sendo indicadas como fonte
de tolerância à seca para obtenção de novas populações tolerantes em programas de seleção
recorrente e de híbridos.
2. A introgressão do IAC Maya em germoplasma da população tolerante 1 avaliada nas
progênies interpopulacionais resultou em aumentos da altura de planta, de prolificidade, do
número de ramificações de pendão e massa de grãos.
3. Existe alta estimativa de herdabilidade em sentido amplo em nível de média de progênie
( 2
)2,1(ˆ
xh ) para os seguintes caracteres: número de fileiras de grãos na espiga (NFE), altura de
planta (AP) e de espiga (AE), florescimento masculino (FM) e stay-green (SG), indicando
elevados ganhos a serem obtidos em programas de seleção interpopulacional.
4. As progênies PI(1,2)-59, PI(1,2)-100, PI(1,2)-109, PI(1,2)-117 e PI(1,2)-147 obtiveram as melhores
performances de MG e parâmetros fisiológicos em casa de vegetação e podem ser exploradas
para a extração de linhagens ou para obtenção de nova população tolerante à seca.
5. Os parâmetros fisiológicos, potencial de água na folha ( w ), condutância estomática (gs(2)
)
e clorofila total (Chl), permitiram a caracterização das progênies PI(1,2)-59, PI(1,2)-100, PI(1,2)-
109, PI(1,2)-147 e da variedade IAC Maya Latente e do DAS2B707, possibilitando que o
melhorista utilize estas ferramentas na seleção de genótipos tolerantes. Confirmou-se o padrão
de sensibilidade da PI(1,2)-141 (Progênie Sensível), pelos valores mais negativos de potencial
de água na folha ( w ), menor conteúdo de clorofila total (Chl), comportamento precoce de
senescência de folhas e colmos e baixa massa de grãos, em déficit hídrico.
6. A variedade IAC IAC Maya Latente apresenta comportamento produtivo estável nos
ambientes de irrigação normal e déficit hídrico, devido à maior sensibilidade (fechamento)
dos estômatos, resultando no maior acúmulo de matéria seca total e pouca redução do índice

87
de colheita, em estresse hídrico. Esta variedade pode ser indicada para compor populações
tolerantes à seca, pelos seus caracteres intrínsecos em ambientes sujeitos ao déficit hídrico.

88
6 REFERÊNCIAS BIBLIOGRÁFICAS
ABDUL JALEEL, C.; MANIVANNAN, P.; KISHOREKUMAR, A.; SANKAR, B.; GOPI,
R.; SOMASUNDARAM, R.; PANEERSELVAM, R. Alterations in osmoregulation,
antioxidant enzymes and indole alkaloid levels in Catharanthusroseus exposed to water
deficit. Colloids and Surfaces B: Biointerfaces, Amsterdam, v.59, p.150-157, 2007.
AMINU, D.; GARBA, Y.M.; MUHAMMAD, A.S. Combining ability and heterosis for
phonologic and agronomic traits in maize (Zea mays L.) under drought conditions in the
Nothern Guinea Savanna of Borno State, Nigeria. Full Length Research Paper. 2014.
ALI, Q.; ELAHI, M.; AHSAN, M.; NADEEM TAHIR, M.H.; BASRA, S.M.A. Genetic
evaluation of maize (Zea mays L.) genotypes at seedling stage under moisture stress.,
International Journal for Agro Veterinary and Medical Sciences,v.5, n.2, p.184-
193, 2011.
ARIAS, C.A.A.; SOUZA Jr., C.L. Genetic variance and covariance componentes related to
intra and interpopulation recurrent selection in maize (Zea mays L.). Genetics and Molecular
Biology, v.21, p.537-544, 1998.
ASHRAF, M. Breeding for salinity tolerance in plants. Crit Rev Plant Sci, v. 13, p. 17–42,
1994.
ASHRAF, M.; FOOLAD, M.R. Roles of glycine betaine and proline in improving plant
abiotic stress resistance. Environmental and Experimental Botany, Amsterdam, v.59, p.
206-216, 2007.
BADU-APRAKU, B.; OYEKUNLE, M.; MENKIR, A.; OBENG-ANTWI, K.; YALLOU,
C.G.; USMAN, I.S.; ALIDU, H. Comparative performance of early-maturing maize cultivars
developed in three eras under drought stress and well-watered environments in west Africa.
Crop Science, Madison, v. 53, n. 4, p. 1298-1311, 2013.
BÄNZIGER, M.; LAFITTE, H.R. Efficiency of secondary traits for improving maize low-
nitrogen target environments. Crop sci., Madison, v. 39, p. 1035-1040, 1997.
BÄNZINGER, M.; EDMEADES, G.O.; BECK, D.; BELLON, M. Breeding for drought
and nitrogen stress tolerance in maize: from theory to practice. Mexico, D.F.: CIMMYT,
2000. 68p.
BÄNZIGER, M.; SENTIMELA, P.S.; HODSON, D.; VIVEK, B. Breeding for improved
abiotic stress tolerance in Africa in maize adapted to southern Africa. Agriculture and Water
Management, v.80, p.212-214, 2006.
BÄNZIGER, M.; ARAUS, J.L. Recent advances in breeding maize for drought and salinit
stress tolerance. In: JENKS, M.A.; HASEGAWA, P.M.; JAIN, S.M. (Eds.) Advances in
molecular breeding toward drought and salt tolerance crops. Netherlands: Springer, 2007.
p. 587-601.

89
BARBANY, J.Y.; KIM, M.; BAUCHAN, G.; BUNCE, J.; REDDY, V.; SICHER, R.C.
Drought responses of foliar metabolites in three maize hybrids differing in water stress
tolerance. Plos One, San Francisco. v.8, n.10, 2013.
BARKER, H.; CAMPOS, M.; COOPER, M.; DOLAN, D.; EDMEADES, G.; HABBEN, J.S.;
WRIGHT, D.; ZINSELMEIER, C. Improving drought tolerance in maize. In: JANICK, J.
(Ed.). Plant breeding reviews. v. 25. N.J.: John Wiley &Sons, 2005. p. 173-235.
BECK, D.L.; BETRAN, F.J.; BÄNZIGER, M.; WILLCOX, M.; EDMEADES, G.O. From
landrace to hybrid: Strategies for the use of source populations and lines in the development
of drought tolerant cultivars. In: PROCEEDING OF A SYMPOSIUM, 1996, El Batán.
Developing Drought and Low N-Tolerant Maize: proceedings… Mexico,DF.: CIMMYT,
1997. p. 369-382.
BEKAVAC, B.P.; STOJAKOVC, M.; IVANOVIC, M.; NASTASIC, A. Genetic analysis of
stay-green trait in broad-based maize populations. Cereal research communications,
Szeged, Hungary, v. 35, n. 1, p. 31-41, 2007.
BERGAMASCHI, H.; DALMAGO, G.A.; COMIRAN, F.; BERGONCI, J.I.; MÜLLER,
A.G; FRANÇA, S.; SANTOS, A.O.; RADIN, B.; BIANCHI, C.A.M; PEREIRA, P.G. Déficit
hídrico e produtividade na cultura do milho. Pesquisa Agropecuária Brasileira, Brasília, v.
41, n. 2, p. 243-249, 2006.
BERGONCI, J.I. Avaliação de indicadores de déficit hídrico em milho. 1997. 111 f. Tese
(Doutorado em Fitotecnia) - Universidade Federal do Rio Grande do Sul, Porto Alegre.
BERNARDO, S.; SOARES, A.A.; MANTOVANI, E.C. Manual de Irrigação. 8.ed. Viçosa:
UFV, 2006. 625p.
BETRÁN, F.J.; BÄNZIGER, M.; BECK, D.L. Relationship between line and topcross
performance under drought and non-stressed conditions in tropical maize. In: PROCEEDING
OF A SYMPOSIUM, 1996, El Batán. Developing Drought and Low N-Tolerant Maize:
proceedings… Mexico,DF.: CIMMYT, 1997. p. 383-386.
BÉTRAN, F.J.; BECK, D.; BÄNZIGER, M.; EDMEADES, G.O. Genetics analysis of inbred
and hybrid grain yield under stress and non stress environments in tropical maize. Crop
Science, Madison, v. 43, p.807-817, 2003.
BLUM , A. Plant Breeding for Stress Environments. Florida:CRC. Press , 1988. 223 p.
BLUM, A. Plant breeding for water-limit environments. 1.ed. New
York/Dordrech/Heidelberg/London: Springer, 2010. 258 p.
BOLAÑOS, J.; CÓRDOVA, H. Desarrollo y mejoramiento de germoplasma de maíz com
tolerancia a sequia para las zonas tropicales de América Central. In: PROCEEDING OF A
SYMPOSIUM, 1996, El Batán. Developing Drought and Low N-Tolerant Maize:
proceedings… Mexico,DF.: CIMMYT, 1997. p. 442-447.

90
BOLAÑOS, J.; EDMEADES, G.O.; MARTINEZ, L. Eight cycles of selection for drought
tolerance in lowland tropical maize. III. Responses in drought-adaptive physiological and
morphological traits. Field Crops Research, Amsterdam. v.31, n.3-4, p.269-286, 1993.
BOLAÑOS, J.; EDMEADES, G.O. Eight cycles of selection for water stress tolerance in
lowland tropical maize. I. Responses in grain yield, biomass and radiation utilization. Field
Crops Research, Amsterdan, v.31, p.233-252, 1993.
BOLAÑOS, J.; EDMEADES, G.O. The importance of the anthesis-silking interval in
breeding for drought tolerance in tropical maize. Field Crops Research, Amsterdam, v. 48, p.
65-80, 1996.
BOLAÑOS, J.; EDMEADES, G.O. The importance of anthesis-silking interval in breeding
for drought tolerance in tropical maize. In: PROCEEDING OF A SYMPOSIUM, 1996, El
Batán. Developing Drought and Low N-Tolerant Maize: proceedings…Mexico,DF.:
CIMMYT, 1997. p. 355-368.
BONO, L.; BERGAMASCHI, H.; ROSA, L.M.G.; FRANÇA, S.; RADIN, B.; SANTOS,
A.O.; BERGONCI, J.I. Alterações no padrão de resposta à luz da condutância estomática do
milho causados pelo déficit hídrico. Revista Brasileira de Agrometeorologia, Santa Maria.
v.9, n.1, p.27-34, 2001.
BORRÁS, L.; SLAFER, G.A.; OTEGUI, M.E. Seed dry weight response to source-sink
manipulations in wheat, maize and soybean: quantitative reappraisal. Field Crop. Res.,
Amsterdam, v. 86, p. 131-146, 2004.
BOYER, J.S.; WESTGATE, M.E. Grain yields with limited water. Journal of Experimental
Botany, London. v.55, p.2385-2394, 2004.
BRUNINI, O.; CAMARGO, M.B.P.de; MIRANDA, L.T.; SAWAZAKI, E. Resistência
estomatal e potencial da água em variedades de milho em condições de campo. In:
CONGRESSO BRASILEIRO DE AGROMETEOROLOGIA, 2, 1981, Pelotas. Anais…
Pelotas, 1981. p. 130-134.
BRUNINI, O. DONZELI, J.L.; ANGELOCCI, L.R.; MAGALHÃES, H.H.S.; SAWAZAKI,
E.; MIRANDA, L.T.de; VOLPE, C.A.; FISCH, G.F. Sensibilidade estomatal de cultivares de
milho em condições de campo. In: CONGRESSO NACIONAL DE MILHO E SORGO, 14,
1982, Florianópolis. Resumos... Florianópolis, 1982a. p. 103.
BRUNINI, O.; CAMARGO, M.B.P.de; CABRAL, O.M.P.; SAWAZAKI, E.; MIRANDA,
L.T.de; VOLPE, C.A.; ANGELOCCI, L.R.; FISCH, G.F. Balanço hídrico em condições de
campo para cultivares de milho. In: CONGRESSO NACIONAL DE MILHO E SORGO, 14,
1982, Florianópolis. Resumos... Florianópolis, 1982b. p. 104.
BUCHANAN-WOLLASTON, V. The molecular biology of leaf senescence. Journal
Experimental Botany, London. v.48, p.181-199, 1997.
CAIRNS, J.E.; HELLIN, J.; SONDER, K.; ARAUS, J.L.; MACROBERT, J.F.;
THIERFELDER, C.; PRASANNA, B.M. Adapting maize production to climate change in
sub-Saharan Africa. Food Security, London, v.5, p.345–360, 2013.

91
ÇAKIR, R. Effect of water stress at different development stages on vegetative and
reproductive growth of corn. Field Crops Research, Amsterdam, v. 89, p. 1-6, 2004.
CÂMARA, T.M.M.; BENTO, D.A.V.; ALVES, G.F.; SANTOS, M.F.; MOREIRA, J.U.V.;
SOUZA JÚNIOR, C.L. Parâmetros genéticos de caracteres relacionados à tolerância à
deficiência hídrica em milho tropical. Bragantia, Campinas. v. 66, n. 4, p. 595-603, 2007.
CAMARGO, M.B.P.; BRUNINI, O.; MACHADO, E.C.; DONZELLI, J.L.; MIRANDA,
L.T.de; SAWAZAKI, E.; ANGELOCCI, L.R.; VOLPE, C.A.; JANUÁRIO, M. Eficiência do
uso da água por cultivares de milho. In: CONGRESSO NACIONAL DE MILHO E SORGO,
14, 1982, Florianópolis. Resumos… Florianópolis, 1982. p. 117.
CAMPOS, H.; COOPER, M.; HABBEN, J.E.; EDMEADES, G.O.; SCHUSSLER, J.R.
Improving drought tolerance in maize: a view from industry. Field Crops Research,
Amsterdam, v. 90, p.19-34, 2004.
CASTLEBERRY, R.M.; LERETTE, R.J. Latente, a new type of drought tolerance? In:
ANNUAL CORN AND SORGHUM RES. CONF., 34, 1979, Chicago.
Anais…Washington:ASTA, 1979. p. 46-56.
CAVATTE, P.C.; MARTINS, S.C.V.; MORAIS, L.E.; SILVA, P.E.M.; SOUZA, L.T.;
DaMATTA, F.M. A Fisiologia dos Estresses Abióticos. In: FRITSCHE-NETO, R.; BORÉM,
A. (Eds.). Melhoramento de plantas para condições de estresses abióticos. Viçosa:UFV,
2011. 250p.
CECCARELLI, S.; GRANDO, S.; IMPIGLIA, A. Choice of selection strategy in breeding
barley for stress environments. Euphytica, Netherlands, v.10, p.307–318, 1998.
CHAPMAN, S.C.; EDMEADES, G.O. Selection improves drought tolerance in tropical
maize populations: II. Direct and correlated responses among secondary traits. Crop Science,
Madison. v.39, n.5, p.1315-1324, 1999.
CHAVES, M.M.; MAROCO, J.P.; PEREIRA, J.S. Understanding plant responses to drought-
from genes to the whole plant. Functional Plant Biology, Collingwood, v. 30, p. 239-264,
2003.
CIIAGRO (CENTRO INTEGRADO DE INFORMAÇÕES AGROMETEORÓGICOS).
2013. Disponível em:<http://www.ciiagro.sp.gov.br/ciiagroonline/MenuBH.htm>. Acesso
em: 13 out. 2013.
COCHRAN, W.G.; COX, G.M. Experimental designs. 2.ed. New York: John Wiley, 1966.
611p.
CONAB – COMPANHIA NACIONAL DE ABASTECIMENTO. Acompanhamento da
safra brasileira de grãos 2013/14 – décimo levantamento, 2014. Disponível em:<hppt://
http://www.conab.gov.br/OlalaCMS/uploads/arquivos/14_04_14_11_56_28_boletim_graos_a
bril_2014.pdf> Acesso em: 12 set. 2014.

92
COMSTOCK, R.E.; ROBINSON, H.F.; HARVEY, P.H. A breeding procedure designed to
make maximum use of both general and specific combining ability. Agronomy Journal,
Madison, v.41, p.360-367, 1949.
COSTA, E. F. N.; SANTOS, M. F.; MORO, G. V.; ALVES, G. F.; JÚNIOR, C. L. S. Herança
da senescência retardada em milho. Pesquisa Agropecuária Brasileira, Brasília, v. 43, n. 2,
p. 207-213, 2008.
CRITCHLEY, W.; KLAUS, S. Water harvesting: a manual for the design and
construction of Water Harvesting Schemes for Plant Production. Rome: FAO 1991. 154
p.
CRUZ, C.D.; REGAZZI, A.J.; CARNEIRO, P.C.S. Modelos biométricos aplicados ao
melhoramento de plantas. Viçosa: UFV, v. 1, 2004. 480 p.
CRUZ, C.D. Programa Genes: Biometria. Editora UFV. Viçosa (MG). 382p. 2006.
DE LEON, N.; COORS, J.G. Twenty-four cycles of mass selection for prolificacy in the
Golden Glow maize population. Crop Science, Madison, v.42, p.325-333, 2002.
DOW, E.W. Resistance to drought and density stress in Canadian and European maize (Zea
mays L.) hybrids. 1981. MS thesis, University of Guelph, Ontario, Canada.
DOW SEMENTES – GUIA DE SEMENTES. 2014/2015. Dow Agrosciences. Annual.
DUDLEY, J.W.; MOLL, R.H. Interpretation and use of estimates of heritability and genetic
variances in plant breeding. Crop Sci. Madison, v. 9, p. 257-261, 1969.
DURÃES, F. O. M.; SANTOS, M. X. dos; GAMA, E. E. G; MAGALHÃES, P. C.;
ALBUQUERQUE, P. E. P.; GUIMARÃES, C. T. Fenotipagem Associada à Tolerância a
Seca em Milho para Uso em Melhoramento, Estudos Genômicos e Seleção Assistida por
Marcadores. Sete Lagoas: Embrapa-CNPMS. 2004. 18 p. (Circular Técnica 39).
DUVICK, D.N. Biodiversity, carrying capacity and people. In: ANNUAL MEETING
AMERICAN ASSOCIATION FOR ADVANCEMENT IN SCIENCE, 1995, Atlanta.
(paper)…
DWIVEDI, S.; UPAHYAYA, H.; SUBUDHI, P.; GEHRING, C.; BAJIC, V.; ORTIZ, R.
Enhancing abiotic stress tolerance in cereals through breeding and transgenic interventions.
In: JANICK, J. (Ed.). Plant breeding reviews. v.33. New Jersey: John Wiley & Sons/ Inc.
Hoboken, 2010. p. 31-93.
EDMEADES, G.O.; CHAPMAN, S.C.; BOLAÑOS, J.; BÄNZIGER, M.; LAFITTE, H.R.
Recent evaluations of progress in selection for drought tolerance in tropical maize. In:
PROCEEDINGS OF THE FOURTH EASTERN AND SOUTHEM AFRICA MAIZE
CONFERENCE, 4, 1994, Harare. Maize research for stress environments: proceedings…
Mexico: CIMMYT, 1995. p. 94-100.
EDMEADES, G.O.; BÄNZIGER, M.; BECK, D.L.; BOLAÑOS, J; ORTEGA, A.
Development and per se performance of CIMMYT maize populations as drought tolerant

93
sources. In: PROCEEDING OF A SYMPOSIUM, 1996, El Batán. Developing Drought and
Low N-Tolerant Maize: proceedings… Mexico,DF.: CIMMYT, 1997a. p. 254-262.
EDMEADES, G.O.; BOLAÑOS, J.; BÄNZIGER, M.; CHAPMAN, S.C.; ORTEGA,A.;
LAFITTE, H.R.; FISCHER, K.S.; PANDY,S. Recurrent selection under managed drought
stress improve grain yields in tropical maize. In: PROCEEDING OF A SYMPOSIUM,
1996, El Batán. Developing Drought and Low N-Tolerant Maize: proceedings… Mexico,
DF.: CIMMYT, 1997b. p. 415-425.
EDMEADES, G.O.; BOLAÑOS, J.; CHAPMAN, S.C.; LAFITTE, H.R..; BÄNZIGER, M.
Selection improves drought tolerance in tropical maize populations: I. gains in biomass, grain
yield, and harvest index. Crop Science, Madison, v. 39, n. 5, p.1306-1315, 1999.
EDMEADES, G.O.; BÄNZIGER, M.; CAMPOS, H.; SCHUSSLER, J. Improving tolerance
to abiotic stress in staple crops: A Random or Planned Process? In: ARNEL R.
HALLAEUER INTERNATIONAL SYMPOSIUM ON PLANT BREEDING, 2006, Mexico.
Book of abstracts…Mexico: CIMMYT, 2006.
ETA-NDU, J.T.; OPENSHAW, S.J. Selection criteria for grain yield and moisture in maize
yield trials. Crop Science, Madison, v. 32, p.332-335, 1992.
FALCONER, D.S. Introduction to Quantitative Genetics. 3.ed. New York:Longman, 1989.
FALKE, K.C. Molecular genetic analysis of modified recurrent full-sib selection in two
European F2 Flint maize populations. Dissertation of Doctor. 2007. 72p. Institut für
Pflanzenzüchtung, Saatgutforschung und Populationsgenetik der Universität Hohenheim.
FALUBA, J.S. Potencial genético da população de milho UFV 7. 2008. 70f. Dissertação
(Mestrado em Genética e Melhoramento de Plantas), Universidade Federal de Viçosa, Viçosa.
FAO: ORGANIZAÇÃO DAS NAÇÕES UNIDAS PARA A AGRICULTURA E
ALIMENTAÇÃO. Food and Agricultural commodities production. 2012. Disponível em:
<http://www.faostat.fav.org/site/339/default.aspx> Acesso em: 12 set. 2014.
FARHAD, M.S.; BABAK, A.M.; REZA, Z.M.; HASSAN, R.S.N.; AFSHIN, T. Response of
proline, soluble sugars, photosynthetic pigments and antioxidant enzymes in potato (Solanum
tuberosum L.) to different irrigation regimes in greenhouse condition. Australian Journal of
Crop Sciencie, Brisbane, v. 5, p. 55-60, 2011.
FAROOQ, M.; WAHID, A.; KOKAYASHI, N.; FUJITA, D.; BASRA, S.M.A. Plant drought
stress: effects mechanisms and management. Agronomy for Sustainable Development, v.
29, p. 185-212. 2009.
FEDERER, W.T. Experimental design. New York: MacMillan, 1955. 544p.
FISCHER, K.S.; JOHNSON, E.C.; EDMEADES, G.O. Breeding and selection for drought
resistance in tropical maize. In: Drought resistance in crops with emphasis on rice. Los
Nanõs: International Rice Research Institute, 1982. p. 377-400.

94
FISCHER, K.S.; EDMEADES, G.O.; JOHNSON, E.C. Recurrent selection for reduced tassel
branch number and reduced leaf-area density above the ear in tropical maize populations.
Crop Science, Madison, v. 27, n. 6, p. 1150-1156, 1987.
FOOLAD, M.R.; LIN, G.Y. Relationship between cold tolerance during seed germination and
vegetative growth in tomato: analysis of response and correlated response to selection. J. Am.
Soc. Hort. Sci., Geneva, v. 126, n. 2, p. 216–220, 2001.
FRAHM, M.A.; ROSAS, J.C.; MAYEK-PÉREZ, N.; LÓPEZ-SALINAS, E.; ACOSTA-
GALLEGOS, J.A.; KELLY, J.D. Breeding beans for resistance to terminal drought in the
lowland tropics. Euphytica, Netherlands, v.136, p.223–232, 2004.
GAMA, E.E.G.; PARENTONI, S.N.; DURÃES, F.O.M.; LEITE, C.E.P.; SANTOS, M.X.;
PACHECO, C.A.P.; OLIVEIRA, A.C. (2001). Tropical maize synthetics improvement for
moisture-stress tolerance for small-scale farmers. Seventh Eastern and Southern Africa
Regional Maize Conference. 11th
-15th
February, 2001, p.288-291.
GAN, S.; AMASINO, R.M. Inhibition of leaf senescence by autoregulated production of
cytokinin. Science, Washington, v. 270, n. 5244, p. 1986–1988, 1995.
GEBRE, G.B. Genetic variability and inheritance of drought and plant density adaptive traits
in maize. 2005. 189p. Thesis of Philosophiae Doctor – University of free State, Faculty of
Natural and Agricultural Sciences, Bloemfontein, South Africa.
GERALDI, I.O.; MIRANDA FILHO, J.B.; VENCOVSKY, R. Estimates of genetic
parameters for tassel characters in maize and breeding persceptives. Maydica, Bergano, v.30,
n.1, p.1-14, 1985.
GOWDA, C.L.L.; SERAJ, R.; SRINIVASON, G.; CHAUHHAN, Y.S.; REDDY, B.V.S.;
RAI, K.N. Opportunities for improving crop water productivity through genetic enhancement
of dryland genetic. In: WANI, di S.P.; ROCKSTRÖM, J.; OWIS, T.Y. (Eds). Rainfed
agriculture. Cambridge: CAWMA, 2009. p.133-163.
GRANIER, C.; TARDIEU, F. Water deficit and spactial pattern of leaf development.
Variability in response can be simulated using a simple model of leaf simulated development.
Plant Physiology, Bethesda, v.119, n.2, p.609-619, 1999.
GRANT, R.F. Interaction between carbon dioxide and water deficits affecting canopy
photosynthesis: simulation and testing. Crop Science, Madison. v. 32, n.6, p.1322-1328,
1992.
GOMES E GAMA, E.E.; PARENTONI, S.N.; DURÃES, F.O.M.; LEITE, C.E.P.; SANTOS,
M.X.DOS; PACHECO, C.A.P.; OLIVEIRA, A.C.DE. Tropical maize synthetics breeding in
moisture-stress tolerance for small-scale farmers in Brazilian semi arid regions. Revista
Milho e Sorgo, Sete Lagoas, v.3, n.1, p.69-76, 2004.
GU, W. L., SHEN, X. Y., DAI, J. Y., HU, A. C., SU, Z. S.; CHEN, J. The physiological
response of maize to drought at different growth stages. J. Shenyang Agril. Univ., Ontario,
v. 21, n.3, p. 186-190, 1990.

95
GUIMARÃES, P.S. Mecanismos fisiológicos e avaliação de progênies de irmãos germanos
interpopulacionais de milho com ênfase em tolerância à seca. 2013. 135f. Tese (Doutorado
em Genética, Melhoramento Vegetal e Biotecnologia) - Instituto Agronômico de Campinas,
Campinas.
GUIMARÃES, P.S.; BERNINI, C.S.; PEDROSO, F.K.J.; PATERNIANI, M.E.A.G.Z.
Characterizing corn hybrids (Zea mays L.) for water shortage by principle components
analysis. Maydica, Bergamo, v.59, n.1, p. 72-79, 2014.
GUIMARÃES, P.S.; BERNINI, C. S.; PATERNIANI, M.E.A.G.Z.; GALLO, P.B.;
LODOVICO, F.A. Seleção de Progênies de Irmãos Germanos Interpopulacionais Visando a
Discriminação Quanto à Tolerância à Seca em Milho. In: XXIX CONGRESSO NACIONAL
DE MILHO E SORGO, 29, 2012, Águas de Lindóia. Diversidade e inovações na era dos
transgênicos: resumos...Campinas: Instituto Agronômico / Associação Brasileira de Milho e
Sorgo, v.1, 2012. p. 3116-3121.
GUTIERREZ, R.M.; SAN MIGUEL, Ch.R.; LARQUÉ-SAAVEDRA, A. Stomatal
conductance in successive selection cycles of the drought tolerance maize population
‘Tuxpeño Sequía’. In: PROCEEDING OF A SYMPOSIUM, 1996, El Batán. Developing
Drought and Low N-Tolerant Maize: proceedings… Mexico,DF.: CIMMYT, 1997. p.
212-214
GUTIERREZ, R.M.;SAN-MIGUEL-CHAVES, R.;LARQUE-SAAVEDRA, A. Physiological
aspects in Tuxpeno maize with improved drought tolerance. Maydica, Bergamo, v. 43, p.
137-141, 1998.
HALLAUER, A.R. Compendium on recurrent selection methods and their application. Crit.
Rev. Plant. Sci., n. 3, n. 1, p.1-33, 1985.
HALLAUER, A.R; MIRANDA FILHO, J.B. de. Quantitative genetics in maize breeding.
2 ed. Ames: Iowa State University Press, 1988. 468p.
HAYANO-KANASHIRO, C.; CALDERÓN-VÁSQUEZ, C.; LACLETTE-IBARRA, E.;
ESTRELLA-HERRERA, L.; SIMPSON, J. Analysis of gene expression and physiological
responses in three Mexican maize landraces under drought stress and recovery irrigation.
Public Library of Science. San Francisco, v.4, n.10, 2009.
HIRON, R. W. P; WRIGHT, S. T. C. The role of endogenous abscisic acid in the response of
plants to stress. J. Expt. Bot., London, v. 24, p. 769-781, 1973.
HÖRTENSTEINER, S. Stay-green regulates chlorophyll and chlorophyll-binding protein
degradation during senesce. Trends in plant science, v. 14, n.3, p. 155-162, 2009.
HSIAO, T.C. Plant responses to water stress. Annual Review of Plant Physiology, Palo
Alto, n.24, p.519-570, 1973.
HUSSAIN, I. Genetics of drought tolerance in maize (Zea mays L.). 2009. 201f. Thesis
(Doctor of philosophy in plant breeding and genetics) - Department of plant breeding &
genetics - University of Agriculture, Faisalabad, Pakistan.

96
IZGE, A.U.; DUGJE, I.Y. Performance of drought tolerant three-way and top cross maize
hybrids in Sudan Savanna of North Eastern Nigeria. Journal of Plant Breeding and Crop
Science. v.3, n.11, p.269-695, 2011.
JAMIESON, P.D.; MARTIN, R.J.; FRANCIS, G.S. Drought influence on grain yield of
barley, wheat and maize. . New Zealand J. Crop and Hort. Sci., Wellington, v. 23, n. 1, p.
55-66, 1995.
JONES, H.G.; LAKSO, A.N.; SYVERTSEN, J.P. Physiological control of water status in
temperature and subtropical fruit trees. Horticultural Reviews, Portland, v.7, p.301-344,
1985.
JONES, H.G.; SERRAJ, R.; LOVEYS, B.R.; XIONG, L.; WHEATON, A.; PRICE, A.
Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of
plant responses to water stress in the field. Funct. Plant Biol., Collingwood, v. 36, p. 978-
989, 2009.
KANG, M.S. Using genotype by environment interaction for crop cultivar development.
Advances in Agronomy, Newark, v.62, p.199-246, 1998.
KRAMER, P.J.; BOYER, J.S. Water relations of plants and soils. San Diego: Academic,
1995. 495p.
KUCHANUR, P.H. Identification of drought tolerant germoplasm in maize (Zea mays L.).
2010. 247 f. Thesis (Doctor of philosophy in genetic and plant breeding) - University of
Agricultural Sciences, Dharwad, India.
LARCHER, W. Ecofisiologia vegetal. São Carlos: RiMa, 2004. 531p.
LARQUE-SAAVEDRA, A.; WAIN, R.L. Abscisic acid levels in relation to drought tolerance
in varieties of Zea Mays L. Nature. London, v. 251, p. 716-717. 1974.
LEVITT, J. Response of plants to environmental stress water, radiation salt and other
stresses. Academic, New York. 1980. 607p.
LIEDGENS, M.; SOLDATI, A.; STAMP, P.; RICHNER, W. Root development of maize
(Zea mays L.). Crop Sci., Madison, v. 40, p. 1665-1672, 2000.
LIZASO, J.I.; WESTGATE, M.E.; BATCHELOR, W.D.; FONSECA, A. Predicting potential
kernel set in maize from singe flowering characteristics. Crop Science, Madison, v. 43, p.
892-903, 2003.
LOPES, M.S.; ARAUS, J.L.; VAN HEERDEN, P.D.R.; FOYER, C.H. Enhancing drought
tolerance in C4 crops. J. Exp. Bot., London, v. 62, p. 3135-3153, 2011.
MAGALHÃES, P.C.; DURÃES, F.O.M.; Fisiologia da Produção de Milho. Embrapa Milho e
Sorgo, Circular Técnica, 76, 10p, 2006.

97
MAGALHÃES, P.C.; ALBUQUERQUE, P.E.P.de; KARAM, D.; CANTÃO, F.R.O.
Caracterização de plantas de milho sob estresse hídrico. Embrapa Milho e Sorgo, Circular
Técnico, 116, 6p, 2009.
MAGALHÃES, P.C.; SOUZA, T.C. Mecanismos fisiológicos de tolerância e avaliação do
estresse hídrico em milho. In: XXIX CONGRESSO NACIONAL DE MILHO E SORGO, 29,
2012, Águas de Lindóia. Diversidade e inovações na era dos transgênicos:
resumos...Campinas: Instituto Agronômico / Associação Brasileira de Milho e Sorgo, v.1,
2012. 780 p.
MASCLAUX-DAUBRESSE, C.; PURDY, S.; LEMAITRE, T.; POURTAU, N.;
TACONNAT, L.; RENOU, J-P.; WINGLER, A. Genetic variation suggests interaction
between cold acclimation and metabolic regulation of leaf senescence. Plant physiology,
Bethesda, v. 143, p. 434-446, 2007.
MCCULLY, M.E. Roots in soil: unearthing the complexities of roots and their rhizospheres.
Annual Review of Plant Physiology and Plant Molecular Biology, v.50, p.695-718, 1999.
MEENA-KUMARI, D.S.; VILAMA, Y.; ARORA, B. Pysiological parameters governing
drought tolerance in maize. Indian Journal Plant Physiology, New Delhi, v.9, n.2, p.203-
204, 2004.
MENGEL, B. B.; BARBER, S. A., Development and distribution of the corn root system
under field conditions. Agron. J., Madison, v. 66, p. 341-344, 1974.
MESSMER, R.; STAMP, P. Trends in drought research. Kasetsart J. (Nat. Sci.), Zurich, v.
44, n.4, p. 507-516, 2010.
MICKELSON, S.M.; STUBER, C.S.; SENIOR, L.; KAEPPLER, S.M. Quantitative trait loci
controlling leaf and tassel traits in a B73 x MO17 population of maize. Crop Science,
Madison. v.42, n.6, p.1902-1909, 2002.
MIRANDA, L.T. de. A característica “latência” do milho (Zea mays L.) e suas possibilidades
no melhoramento. 1972. 93f. Tese (Doutoramento) - Escola Superior de Agricultura “Luiz de
Queiroz”- USP, Piracicaba.
MIRANDA, L.T. de; MIRANDA, L.E.C. de; SAWAZAKI, E.; SCHMIDT, N. C. Maize
Genetics Cooperation Newsletter, v. 56, p. 32-33. 1982. Disponível em:
<www.agron.missouri.edu > Acesso em: 22/07/2013.
MIRANDA, L.T.; MIRANDA, L.E.C. Milho: Genética Ecológica. In: FURLANI, A.M.C.;
VIÉGAS, G.P. O melhoramento de plantas no Instituto Agronômico. Campinas: Instituto
Agronômico, v.1, 1993. p. 363-409.
MONNEVEUX, P.; RIBAUT, J.M. Secundary traits for drought tolerance improvement in
cereals. In: RIBAUT, J.M. (Ed.) Drought adaptation in cereals. N.Y.: The Haworth Press,
2006. p. 97-144.

98
MONNEVEUX, P.; SANCHEZ, C.; TIESSEN, A. Future progress in drought tolerance in
maize needs new secondary traits and cross combinations. The journal of agricultural
science, Toronto, v. 146, p. 287-300, 2008.
MUCHOW, R.C.; SINCLAIR, T.R. Nitrogen response of leaf photosynthesis and canopy
radiation use efficiency in field grown maize and sorghum. Crop Science, Madison, n.34,
p.721-727, 1994.
MUÑOZ, O. A. Relaciones agua-planta bajo sequia, en varios sinteticos de maiz resistentes a
sequia y heladas. 1975. f. Tesis (Doctor en Ciências) - Colegio de Postgraduados – ENA,
Chapingo, Mexico.
MUTAVA, R.N.; PRASAD, P.V.V.; TUINSTRA, M.R.; KOFOID, K.D.; YU, J.
Characterization of sorghum genotypes to water stress. Aust. J. Crop. Sci., Brisbane, v.1,
p.31-36, 2008.
NASS, L.L.; MIRANDA FILHO, J.B. Synthesis of new composites of maize (Zea mays L.)
for population improvement in Brazil. In: Memorias de la XVIII Reunión Latinoamericana
del Maíz. Embrapa/CNPMS, CIMMYT, Sete Lagoas, p.309-317, 1999.
PAIVA, A.S.; FERNANDES, E.J.; RODRIGUES, T.J.D.; TURCO, J.E.P. Condutância
estomática em folhas de feijoeiro submetido a diferentes regimes de irrigação. Engenharia
Agrícola, Jaboticabal, v. 25, n.1, p.161-169, 2005.
PALACIOS DE LA ROSA, G. Variedades e híbridos de maiz latente y tolerantes a La sequia
y a las heladas. Agricultura Mexicana, México, v.107, p. 38-39, 1959.
PANDEY, S.; GARDNER, C.O. Recurrent selection for population, variety, and hybrid
improvement in tropical maize. Advances in Agronomy, v.48, p. 1-87, 1992.
PARK, S.Y.; YU, J.W.; PARK, J.S.; LI, J.; YOO, S.C.; LEE, N.Y.; LEE, S.K; JEONG, S.W.;
SEO, H.S.; KOH, H.J.; JEON, J.S; PARK, Y.II.; PAEK, N.C. The senescence-induced stay
green protein regulates chlorophyll degradation. The plant cell, Norwich, v.19, p.1641-1664,
2007.
PATERNIANI, E. Selection among and within half-sib families in a Brazilian population of
maize (Zea mays L.). Crop. Scien., Madison, v. 7, p. 212-216. 1967.
PATERNIANI, E. Avaliação de métodos de seleção entre e dentro de famílias de meios-
irmãos no melhoramento de milho (Zea mays L.). 1968. 92f. Dissertação (Mestrado) – Escola
Superior de Agricultura Luiz de Queiroz, Piracicaba.
PATERNIANI, E.; VENCOVSKY, R. Reciprocal recurrent selection in maize based on
testcrosses of half-sib families. Maydica, Bergamo, v.22, p.141-152, 1977.
PATERNIANI, E. VENCOVSKY, R. Reciprocal recurrent selection in maize based on half-
sib progênies and prolific plants in maize. Maydica, Bergamo, v.23, p.209-219, 1978.

99
PATERNIANI, M.E.A.G.Z.; SAWAZAKI, E.; GALLO, P.B.; LUDERS, R.R.; SILVA,
R.M.da. Estimatives of genetic parameters in a maize composite and potential for recurrent
selection. Crop Breeding and Applied Biotechnology, n°4, p.81-85, 2004.
PATERNIANI, M.E.A.G.Z.; BERNINI, C.S.; GUIMARÃES, P.S.; GALLO, P.B. Seleção de
Progênies de Meios-Irmãos e Estimativas de Parâmetros Genéticos da População Tolerante à
Seca de Milho IAC Tol 1. In: XXIX CONGRESSO NACIONAL DE MILHO E SORGO, 29,
2012, Águas de Lindóia. Diversidade e inovações na era dos transgênicos:
resumos...Campinas: Instituto Agronômico / Associação Brasileira de Milho e Sorgo, v.1,
2012. p. 3129-3134.
PHOELMAN, J.M.; SLEPER, D.A. Breeding field crops. Iowa: Iowa State University
Press, 4°Ed., 1995, 494p.
RAMALHO, M.A.P. Eficiência relativa de alguns processos de seleção intrapopulacional no
milho baseados em famílias não endógamas. 1977. 122f. Tese (Doutorado em Genética e
Melhoramento de Plantas) – Escola Superior de Agricultura “Luiz de Queiroz”, Piracicaba.
REIS, M.C.dos; PADUA, J.M.V.; ABREU, G.B.; GUEDES, F.L.; BALBI, R.V.; SOUZA,
J.C. Estimates for genetic variance components in reciprocal recurrent selection in
populations derived from maize single-cross hybrids. The Scientific World Journal. 2014.
RIBAULT, J.M.; HOISINGTON, D.H.; DEUTSCH, J.A.; JIANG, C.; GONZALES-de-
LEON, D. Identification of quantitative trait loci under drought condition in tropical maize. 2.
Yield components and marker-assisted selection strategies. Theor. Appl. Genet., n.94, p.887-
896, 1997.
RIBAUT, J.M.; BÉTRAN, J.; MONNEVEUX, P.; SETTER, T. Drought tolerance in maize.
In: BENNETZEN, J.L.; HAKE, S.C. (Eds.). Handbook of maize: Its biology. Netherlands:
Springer, 2009. p. 311-344.
RICHARDS, R.A.Variation between and within species of rape seed (B. Campestris and B.
napus) in response to drought stress. III. Physiological and physiochemical characters. Aust.
J. Agric. Res., v. 29, p. 491-501, 1978.
RIVERO, R.M., SHULAEV, V., BLUMWALD, E. Cytokinin-dependent photorespiration
and the protection of photosynthesis during water deficit. Plant Physiol. Bethesda, v. 150, p.
1530–1540, 2009.
RITCHIE, S.W.; HANWAY, J.J.; BENSON, G.O. How the corn plant develops. Special
Report n°. 48. Iowa State University of Science and Technology Cooperative Extension
Service, Ames. 1993.
SALAH, H.B.H.; TARDIEU, F. Control of leaf expansion rate of droughted maize plants
under fluctuating evaporative demand. A superposition of hydraulic and chemical messages?
Plant Physiol., Bethesda, v.114, p. 893-900, 1997.
SAWAZAKI, E. Melhoramento da cultivar de milho IAC Maya. Campinas: Fundação
Cargill, 1980. 49p.

100
SAWAZAKI, E. FERREIRA, S.; LEVI, J.L.; NETO, E.V.; INADA, I.; VECHIO, W.A.de;
MIRANDA, L.T.de; MAEDA, J.A. Teste regional de cultivares de milho em parcelas com e
sem bordadura. In: XV CONGRESSO NACIONAL DE MILHO E SORGO, 15, 1984,
MACEIÓ. Anais...Brasília: Embrapa-DDT, 430p.
SACCARDY, K.; CORNIC, G., BRULFERT, J.; REYSS, A. Effect of drought stress on net
CO2 uptake by Zea leaves. Planta, Monterrey, v.199, p.589–595, 1996.
SCAPIM, C.A.; CARVALHO, C.G.P.; CRUZ, C.D. Uma proposta de classificação dos
coeficientes de variação para cultura do milho. Pesquisa Agropecuária Brasileira, Brasília,
v.30, p.683-686, 1995.
SCHACHTMAN, D.P.; GOODGER, J.Q. Chemical root to shoot signaling under drought.
Trends Plant Science, Cambridge, v.13, p.754-760, 2008.
SCHULZE, E-D.; BECK. E.; MÜLLER-HOHENSTEIN,K. Plant Ecology. Berlin: Springer,
2005. 692 p.
SETIMELA, P.; MACROBERT, J.; ATLIN, G.N.; MAGOROKOSHO, C.; TAREKEGNE,
A.; MAKUMBI, D.; TAYE, G. Performance of elite maize varieties tested on-farm trials in
eastern and southern Africa. Presented at the ASA, CSSA, and SSSA International Annual
Meetings, October 2012, Cincinnati, Ohio, USA.
SILVA, H.D.; REGAZZI, A.J.; CRUZ, C.D.; VIANA, J.M.S. Análise de experimentos em
látice quadrado com ênfase em componentes de variância. i. Análises individuais. Pesquisa
Agropecuária Brasileira, Brasília, v. 34, n. 10, p .1811-1822, 1999.
SILVA, R.G.; MIRANDA, G.V.; CRUZ, C.D.; GALVÃO, J.C.C.; SILVA, D.G. Estimativas
de parâmetros genéticos da produção de grãos das populações de milho UFVM 100 e UFVM
200, submetidas a déficit hídrico. Revista Brasileira de Milho e Sorgo, Sete Lagoas. v.7, n.
1, p. 57-67, 2008.
SILVA, M.de.A.; SANTOS, C.M.dos; LABATE, C.A.; GUIDETTI-GONZALEZ, S.;
BORGES, J.de.S.; FERREIRA, L.C.; DeLIMA, R.O.; FRITSCHE-NETO, R. Melhoramento
para eficiência no uso de água. In: FRITSCHE-NETO, R.; BORÉM, A. (Ed.). Melhoramento
de plantas para condições de estresses abióticos. Viçosa: UFV, 2011. 250p.
SOUZA Jr., C.L.; GERALDI, I.O.; ZINSLY, J.R. Influence of tassel size on the expression of
prolificacy in maize. Maydica, Bergamo, v. 30, n. 3, p. 321-328, 1985.
SOUZA Jr., C.L.; SANTOS, M.X.dos; MAGNAVACA, R.; GOMES e GAMA, E.E.
Estimativas de parâmetros genéticos na interpopulação de milho BR-105 x BR-106 e suas
implicações no melhoramento. Pesquisa Agropecuária Brasileira, Brasília, v.28, n.4, p.473-
479, 1993.
SOUZA, T. C.; MAGALHÃES, P. C.; PEREIRA, F. J.; CASTRO, E. M.; PARENTONI, S.N.
Morpho-physiology and maize grain yield under periodic soil flooding in sucessive selection
cycles. Acta Physiol. Plant., Poznan, v. 33, n. 5, p. 1877-1885, 2011.

101
SOUZA, T.C.de. Aplicação de Ácido Abscísico (ABA) e Caracterização Fisiológica e
Anatômica em Dois Híbridos de Milho Contrastantes ao Estresse de Seca. 2012. 134f. Tese
(Doutorado em Fisiologia de Plantas) – Universidade Federal de Lavras (UFLA), Lavras.
SRINIVASAN, G. BÄNZIGER, M.; EDMEADES, G.O.; LOTHROP, J.E.; TORRES, J.L.
Identification of drought tolerance in elite tropical highland maize germplasm. In: In:
PROCEEDING OF A SYMPOSIUM, 1996, El Batán. Developing Drought and Low N-
Tolerant Maize: proceedings… Mexico,DF.: CIMMYT, 1997. p. 309-312.
STREIT, N.M.; CANTERLE, L.P.; CANTO, M.W. do; HECKTHEUER, L.H.H. As
clorofilas. Ciência Rural, Santa Maria, v. 35, n. 3, p. 748-755, 2005.
TAIZ, L.; ZEIGER, E. Fisiologia vegetal. 3 ed. Porto Alegre: Artmed, 2006. 722p.
TEARE, I.D.; SCHIMMELPFENNING, H.; WALDREN, R.P. Rainout shelter and drainage
lysimeters to quantitatively measure drought stress. Agronomy Journal, Madison. v.65, n.4,
p.544-457, 1973.
TORRES NETTO, A.; CAMPOSTRINI, E.; OLIVEIRA, J. G.; SMITH, R. E. B.
Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502 readings in
coffee leaves. Scientia Horticulturae, Agassiz, v. 104, n. 2, p. 199-209, 2005.
THOMAS, H.; HOWARTH, C.J. Five ways to stay-green. Journal of experimental Botany,
London, v.51, n. 1, p. 329-337, 2000.
THOMAS, H.; SMART, C.M. Crops that stay-green. Ann. Appl. Biol., Warwick, v.123, p.
193-219, 1993.
VENCOVSKY, R.; BARRIGA, P. Genética biométrica no fitomelhoramento. Ribeirão
Preto: Revista Brasileira de Genética. 1992, 486p.
VENUPRASAD, R.; LAFITTE, H.R.; ATLIN, G.N. Response to direct selection for grain
yield under drought stress in rice. Crop Science, Madison, v.47, p.285-293, 2007.
WANG, B.; LI, Z.; ENEJI, A.E.; TIAN, X.; ZHAI, Z.; LI, J.; DUAN, L. Effects of coronatine
on growth, gas exchange traits, chlorophyll content, antioxidant enzymes and lipid
peroxidation in maize (Zea mays L.) seedlings under simulated drought stress. Plant
Production Science, Tokyo, n.11, p.283-290. 2008.
WESTGATE, M.E. Physiology of flowering in maize: identifying avenues to improve kernel
set during drought. In: PROCEEDING OF A SYMPOSIUM, 1996, El Batán. Developing
Drought and Low N-Tolerant Maize: proceedings… Mexico,DF.: CIMMYT, 1997. p.
136-141.
WESTGATE, M.E.; OTEGUI, M.E.; ANDRADE, F.H.; Physiology of the corn plant. In:
SMITH, C.W.; BÉTRAN, J.; RUNGE, E.C.A. (Eds). Corn: origin, history, technology and
production. New Jersey: John Wiley & Sons, Inc., 2004. p.235-271.
WILLMAN, M.R.; BELOW, F.E.; LAMBERT, R.J.; HOWEY, A.E.; MIES, D.W. Plants
traits related to productivity of maize. I. Genetic variability, environmental variation, and

102
correlation with grain yield and stalk lodging. Crop Science, Madison, v.27, p.1116-1121,
1987.
YANG, J.C.; ZHANG, J.H.; WANG, Z.Q.; ZHU, Q.S.; LIU, L.J. Involvement of abscisic
acid and cytokinins in the senescence and remobilization of carbon reserves in wheat
subjected to water stress during grain filling. Plant Cell Environ., Logan, v. 26, n.10, p.
1621–1631, 2003.
ZAIDI, P. H.; YADAV, M.; SINGH, D. K.; SINGH, R. P. Relationship between drought and
excess moisture tolerance in tropical maize (Zea mays L.). Australian Journal of Crop
Science, Brisbane, v. 1, n. 3, p. 78-96, 2008.
ZHOU, Q.; YU, B. Changes in content of free, conjugated and bound polyamines and osmotic
adjustment in adaptation of vetiver grass to water deficit. Plant physiology and
Biochemistry, v.48, p.417-425, 2010.
ZHU, J.K. Salt and drought stress signal tranduction in plants. Annual Review of Plant
Biology, Palo Alto, v.53, n.1, p.247-273, 2002.
ZINSELMEIER, C.; LAUER, M.J.; BOYER, J.S. Reversing drought-induced losses in grain
yield: sucrose maintains embryo growth in maize. Crop Science, Madison. v.35, p.1390-
1400, 1995.
ZIMOYO, C.; BERNARDO, R. Drought tolerance in maize: indirect selection through
secondary traits versus genomewide selection. Crop Science, Madison, v.53, p. 1269-1275,
2013.

103
ANEXOS

104
Anexo 1. Médias de massa de grãos (MG), estimativas de heterose em relação à média dos
pais ( H ), heterose relativa ( %H ) de 20% das progênies interpopulacionais (PI(1,2))
selecionadas, no ciclo 0 e testemunhas em Campinas e Mococa, na safrinha de 2012.
Progênies
interpopulacionais
MG
(g planta-1
)
MG
(kg ha-1
)
MG(1)
Maya Latente
(%)
MG(2)
DAS2B707
(%)
PI(1,2)-17 139,33 7.431 129,92 92,04
PI(1,2)-18 124,48 6.639 116,08 82,23
PI(1,2)-23 147,23 7.853 137,31 97,27
PI(1,2)-24 134,34 7.165 125,28 88,75
PI(1,2)-27 144,71 7.718 134,95 95,60
PI(1,2)-40 135,90 7.248 126,73 89,78
PI(1,2)-44 130,95 6.985 122,12 86,51
PI(1,2)-49 129,98 6.932 121,21 85,86
PI(1,2)-52 142,27 7.588 132,68 93,99
PI(1,2)-59 141,62 7.553 132,06 93,55
PI(1,2)-68 144,37 7.700 134,63 95,37
PI(1,2)-71 139,55 7.443 130,14 92,19
PI(1,2)-72 143,13 7.634 133,47 94,55
PI(1,2)-73 150,42 8.022 140,26 99,36
PI(1,2)-81 124,48 6.639 116,07 82,23
PI(1,2)-100 144,30 7.696 134,56 95,32
PI(1,2)-102 124,27 6.628 115,89 82,10
PI(1,2)-106 137,55 7.336 128,26 90,86
PI(1,2)-107 143,82 7.671 134,12 95,01
PI(1,2)-109 140,49 7.493 131,01 92,80
PI(1,2)-110 144,90 7.729 135,13 95,73
PI(1,2)-117 148,08 7.898 138,10 97,83
PI(1,2)-124 141,80 7.563 132,23 93,67
PI(1,2)-129 148,47 7.918 138,45 98,08
PI(1,2)-136 127,12 6.780 118,55 83,98
PI(1,2)-138 152,75 8.147 142,45 100,91
PI(1,2)-141 129,61 6.912 120,86 85,61
PI(1,2)-147 133,13 7.100 124,14 87,94
PI(1,2)-155 137,10 7.312 127,84 90,56
PI(1,2)-163 143,53 7.654 133,82 94,80
PI(1,2)-174 132,56 7.070 123,62 87,57
PI(1,2)-181 142,78 7.615 133,15 94,32
PI(1,2)-186 141,96 7.572 132,39 93,78
PI(1,2)-188 130,60 6.965 121,78 86,27
PI(1,2)-189 141,44 7.544 131,90 93,44
PI(1,2)-191 141,67 7.556 132,12 93,59
PI(1,2)-193 136,23 7.266 127,05 90,00
PI(1,2)-194 127,75 6.814 119,13 84,39
PI(1,2)-195 141,88 7.567 132,31 93,73
PI(1,2)-196 134,57 7.175 125,45 88,87
Média Prog.
(selecionadas) 138,53 7.388 129,2 91,52
AV (124,27-152,75) (6.628-8.147) (115,89-142,45) (82,10-100,92)

105
Média Geral 127,35 6.780
Testemunhas
Pop. Tol. 1 115,59 6.049 105,77 74,92
Pop. Sensível 92,79 5.346 93,47 66,22
DAS2B707 149,56 8.073 141,16 100
IAC Maya Lat. 101,16 5.719 100 70,83
H (40 PI(1,2)) 30,15
%H (40 PI(1,2)) 27,82
H (196 PI(1,2)) 18,98
%H (196 PI(1,2)) 17,51
CV(%) 10,6 13,7
(1,2): porcentagem em relação às testemunhas IAC Maya Latente e DAS2B707, respectivamente (em kg ha-1
).
Anexo 2. Médias de altura de planta (AP), altura de espiga (AE), florescimento masculino
(FM), florescimento feminino (FF), intervalo entre florescimentos (IF), estimativas de
heterose em relação à média dos pais ( H ) e heterose relativa ( %H ) de 20% das progênies
interpopulacionais (PI(1,2)) selecionadas, no ciclo 0 e testemunhas em Campinas e Mococa, na
safrinha de 2012.
Progênies
interpopulacionais
AP
(cm)
AE
(cm)
FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
PI(1,2)-17 251 148 65,6 69,3 3,7
PI(1,2)-18 232 152 65,9 69,3 3,4
PI(1,2)-23 246 146 66,2 69,7 3,5
PI(1,2)-24 243 144 67,2 70,1 2,9
PI(1,2)-27 254 146 67,7 70,6 2,9
PI(1,2)-40 238 156 70,6 73,0 2,5
PI(1,2)-44 252 147 69,6 72,0 2,4
PI(1,2)-49 254 156 68,7 70,0 1,3
PI(1,2)-52 251 157 69,9 71,1 1,2
PI(1,2)-59 239 146 68,0 71,2 3,2
PI(1,2)-68 227 153 71,1 73,5 2,5
PI(1,2)-71 257 151 67,5 70,3 2,8
PI(1,2)-72 252 154 69,3 70,2 0,9
PI(1,2)-73 228 139 68,5 71,9 3,5
PI(1,2)-81 253 154 70,4 72,0 1,6
PI(1,2)-100 247 126 69,4 71,5 2,0
PI(1,2)-102 224 144 69,5 74,3 4,8
PI(1,2)-106 253 142 70,2 71,7 1,5
PI(1,2)-107 222 134 68,8 72,7 3,9
PI(1,2)-109 230 127 70,6 71,3 0,8
PI(1,2)-110 227 138 69,5 72,3 2,8
PI(1,2)-117 241 139 70,1 72,8 2,8
PI(1,2)-124 235 150 70,3 73,1 2,8
PI(1,2)-129 245 148 68,1 70,5 2,4
PI(1,2)-136 228 130 70,7 73,6 2,9
Anexo 1. (Continuação)

106
PI(1,2)-138 244 143 70,6 72,8 2,2
PI(1,2)-141 247 145 68,7 70,4 1,7
PI(1,2)-147 240 154 70,4 71,6 1,2
PI(1,2)-155 249 130 68,8 72,6 3,7
PI(1,2)-163 239 133 69,9 74,4 4,5
PI(1,2)-174 244 141 69,0 70,4 1,4
PI(1,2)-181 238 126 70,9 72,6 1,8
PI(1,2)-186 220 145 67,9 71,0 3,1
PI(1,2)-188 226 130 66,7 69,8 3,0
PI(1,2)-189 240 148 71,2 74,9 3,8
PI(1,2)-191 233 139 68,2 70,6 2,5
PI(1,2)-193 256 145 69,7 72,3 2,6
PI(1,2)-194 231 153 71,3 74,9 3,7
PI(1,2)-195 230 131 70,6 72,9 2,3
PI(1,2)-196 232 131 69,3 71,7 2,4
Média Prog.
(selecionadas) 240 143 69,2 71,8 2,6
AV (220-257) (126-157) (65,6-71,3) (69,3-74,9) (0,8-4,8)
Média geral 244 142 68,9 71,2 2,2
Testemunhas
Pop.Tol.1 215 121 68,6 70,6 2,0
Pop. Sensível 203 124 68,9 71,7 2,7
DAS2B707 219 114 69,5 72,0 2,5
Maya Lat. 261 145 68,8 71,4 2,6
H (40 PI(1,2)) 2 0,29
%H (40 PI(1,2)) 0,84 12,45
H (196 PI(1,2)) 6 -0,1
%H (196 PI(1,2)) 2,52 -4,34
CV(%) 3,87 5,08 1,68 2,38 55,25
Anexo 3. Médias de plantas acamadas e quebradas (A+Q), número de ramificações do pendão
(NRP), stay-green (SG), prolificidade (PROL), comprimento de espiga (CE), número de
fileiras de grãos na espiga (NFE), estimativas de heterose em relação à média dos pais ( H ) e
heterose relativa ( %H ) de 20% das progênies interpopulacionais (PI(1,2)) selecionadas, no
ciclo 0 e testemunhas em Campinas e Mococa, na safrinha de 2012.
Progênies
interpopulacionais
A+Q(1)
(arcoseno)
NRP
(ramif.
pendão-1
)
SG
(nota)
PROL
(espigas
planta-1
)
CE
(cm)
NFE
(f.p.e.)
PI(1,2)-17 13,4 15,5 1,6 1,05 14,9 16,0
PI(1,2)-18 12,4 15,5 1,8 1,03 15,7 12,9
PI(1,2)-23 14,9 14,0 1,5 0,76 14,0 15,1
PI(1,2)-24 11,1 15,0 2,0 0,97 15,7 15,0
PI(1,2)-27 13,8 11,5 1,7 1,00 15,8 14,6
PI(1,2)-40 4,3 19,5 1,6 0,93 14,8 15,0
PI(1,2)-44 13,1 13,5 1,2 0,85 15,1 15,7
PI(1,2)-49 10,3 14,0 1,6 0,91 15,4 13,4
PI(1,2)-52 10,9 13,5 2,3 1,19 15,7 13,6
Anexo 2. (Continuação)

107
PI(1,2)-59 12,4 13,5 1,0 1,01 16,1 14,0
PI(1,2)-68 7,7 12,0 1,1 0,79 15,3 14,3
PI(1,2)-71 5,3 12,5 1,9 1,01 15,7 13,5
PI(1,2)-72 11,4 14,5 1,3 0,82 14,8 12,8
PI(1,2)-73 8,7 14,5 1,3 1,14 15,7 12,9
PI(1,2)-81 17,0 15,0 1,4 0,86 15,9 14,6
PI(1,2)-100 12,4 17,0 1,4 1,07 13,4 14,0
PI(1,2)-102 12,2 19,0 1,2 1,31 13,8 14,6
PI(1,2)-106 3,1 15,0 1,6 0,91 15,6 13,9
PI(1,2)-107 9,5 15,5 1,8 1,11 15,5 14,6
PI(1,2)-109 11,4 16,5 1,5 0,91 16,1 15,6
PI(1,2)-110 6,6 19,0 1,7 0,78 15,0 14,0
PI(1,2)-117 12,0 16,0 2,1 0,89 14,5 15,7
PI(1,2)-124 6,7 14,5 1,9 0,83 15,2 16,5
PI(1,2)-129 10,8 15,0 1,6 1,13 14,6 15,0
PI(1,2)-136 12,5 13,5 2,0 0,91 15,4 15,8
PI(1,2)-138 8,2 16,0 1,6 0,91 16,3 17,2
PI(1,2)-141 9,3 17,5 2,2 0,96 16,5 14,8
PI(1,2)-147 11,9 17,0 2,3 0,84 16,7 13,3
PI(1,2)-155 10,5 15,5 1,8 0,91 17,5 13,5
PI(1,2)-163 12,3 13,0 1,2 0,83 15,1 14,2
PI(1,2)-174 11,8 17,0 1,1 0,94 16,0 13,1
PI(1,2)-181 14,2 16,5 1,2 0,83 16,4 14,2
PI(1,2)-186 7,7 15,5 1,4 1,05 15,3 15,3
PI(1,2)-188 8,4 18,0 1,8 0,85 15,6 15,4
PI(1,2)-189 11,4 12,5 1,6 0,89 15,0 16,2
PI(1,2)-191 10,5 14,0 2,0 0,95 16,0 16,2
PI(1,2)-193 12,0 13,5 1,5 0,93 17,4 16,8
PI(1,2)-194 6,6 13,5 2,1 0,98 15,8 13,2
PI(1,2)-195 8,6 12,0 1,6 0,86 17,3 14,4
PI(1,2)-196 8,9 15,0 1,6 0,99 14,4 15,7
Média Prog.
(selecionadas) 10,4 15,0 1,6 0,95 15,5 14,6
AV (3,1-17) (11,5-19,5) (1-2,3) (0,76-1,31) (13,4-17,5) (12,7-17,1)
Média geral 11,27 15,4 1,57 0,93 15,45 14,5
Testemunhas
Pop. Tol. 1 9,1 12,5 1,2 1,06 15,9 16,0
Pop. Sensível 7,9 13,0 1,2 1,18 15,8 14,0
DAS2B707 8,0 13,0 1,0 0,76 15,8 16,0
IAC Maya Lat. 12,2 15,5 1,3 1,34 15,7 16,0
H (40 PI(1,2)) 1,0 0,35
%H (40 PI(1,2)) 7,14 28
H (196 PI(1,2)) 1,4 0,32
%H (196 PI(1,2)) 10 25,6
CV (%) 27,65 2,18 8,0 12,72 6,33 4,49
(1)
: dados transformados para arcoseno 100)5,0( x ; f.p.e: fileiras por espiga.
Anexo 3. (Continuação)

108
Anexo 4. Médias de massa de grãos (MG), altura de planta (AP), altura de espiga (AE),
florescimento masculino (FM), florescimento feminino (FF), intervalo entre florescimentos
(IF), plantas acamadas e quebradas (A+Q), número de ramificações do pendão (NRP), stay-
green (SG), prolificidade (PROL), comprimento de espiga (CE), número de fileiras de grãos
na espiga (NFE), estimativas de heterose em relação à média dos pais ( H ) e heterose relativa
( %H ) de 8 progênies interpopulacionais selecionadas em Campinas e Mococa, na safrinha de
2012.
Progênies
Interpopulacionais
MG
(g planta-1
)
MG
(kg ha-1
)
AP
(cm)
AE
(cm)
FM
(d.a.s.)
FF
(d.a.s)
IF
(dias)
PI(1,2)-49 129,98 6.932 254 156 68,7 70,0 1,3
PI(1,2)-52 142,27 7.588 251 157 69,9 71,1 1,2
PI(1,2)-59 141,62 7.553 239 146 68,0 71,2 3,2
PI(1,2)-72 143,13 7.634 252 154 69,3 70,2 0,9
PI(1,2)-100 144,30 7.696 247 126 69,4 71,5 2,0
PI(1,2)-109 140,49 7.493 230 127 70,6 71,3 0,8
PI(1,2)-117 148,08 7.898 241 139 70,1 72,8 2,8
PI(1,2)-147 133,13 7.100 240 154 70,4 71,6 1,2
Média de 8 prog.
(selecionadas) 139,18 7.487 244 145 69,5 71,1 1,7
Média geral 127,35 6.780 244 142 68,9 71,2 2,2
H (8 PI(1,2)) 30,8 6 -0,65
%H (8 PI(1,2)) 28,42 2,52 -27,9
Progênies
Interpopulacionais
A+Q(1)
(%)
NRP
(ramif.
pendão-1
)
SG
(nota)
PROL
(espigas
planta-1
)
CE
(cm)
NFE
(f.p.e.)
PI(1,2)-49 10,3 14,0 1,6 0,91 15,4 13,4
PI(1,2)-52 10,9 13,5 2,3 1,19 15,7 13,6
PI(1,2)-59 12,4 13,5 1,0 1,01 16,1 14,0
PI(1,2)-72 11,4 14,5 1,3 0,82 14,8 12,8
PI(1,2)-100 12,4 17,0 1,4 1,07 13,4 14,0
PI(1,2)-109 11,4 16,5 1,5 0,91 16,1 15,6
PI(1,2)-117 12,0 16,0 2,1 0,89 14,5 15,7
PI(1,2)-147 11,9 17,0 2,3 0,84 16,7 13,3
Média de 8 prog.
(selecionadas) 11,3 15,5 1,7 0,96 15,5 14,1
Média geral 11,27 15,4 1,57 0,93 15,45 14,50
H (8 PI(1,2)) 1,5 0,45
%H (8 PI(1,2)) 10,7 36
(1): dados transformados para arcoseno 100)5,0( x ; f.p.e: fileiras por espiga.

109
Anexo 5. Médias de altura de planta (AP), altura de espiga (AE), plantas acamadas e
quebradas (A+Q), florescimento masculino (FM), florescimento feminino (FF), intervalo
entre florescimentos (IF), stay-green (SG), massa de grãos (MG) de 196 progênies
interpopulacionais (PI(1,2)) avaliadas em Campinas, na safrinha de 2012.
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-
1)
PI(1,2)-1 217,0 121,0 8,8 69,5 72,0 2,5 1,3 112,0
PI(1,2)-2 215,0 130,5 10,0 68,5 70,5 2,0 1,5 133,0
PI(1,2)-3 218,0 143,5 11,3 71,5 73,0 1,5 2,5 138,2
PI(1,2)-4 234,0 140,5 14,7 70,5 73,0 2,5 1,4 148,6
PI(1,2)-5 211,0 123,0 9,4 69,5 76,0 6,5 1,3 162,3
PI(1,2)-6 218,0 132,5 8,1 72,5 76,0 3,5 1,4 149,3
PI(1,2)-7 234,0 155,0 11,3 70,5 73,0 2,5 1,6 134,4
PI(1,2)-8 222,0 136,5 14,8 72,5 75,5 3,0 1,2 129,3
PI(1,2)-9 231,0 137,0 8,1 70,5 73,0 2,5 1,7 120,6
PI(1,2)-10 212,0 133,0 17,6 72,0 73,5 1,5 1,3 147,8
PI(1,2)-11 239,0 156,5 13,6 67,0 68,5 1,5 2,1 149,6
PI(1,2)-12 242,0 156,5 11,5 69,0 71,5 2,5 1,5 109,8
PI(1,2)-13 246,0 149,5 14,3 69,0 70,5 1,5 1,2 143,5
PI(1,2)-14 225,0 137,5 15,1 70,5 72,5 2,0 1,9 118,7
PI(1,2)-15 240,0 155,0 19,4 70,0 72,0 2,0 1,9 109,4
PI(1,2)-16 249,0 156,5 12,5 68,5 71,0 2,5 1,3 143,9
PI(1,2)-17 261,0 163,0 13,5 65,2 69,1 3,9 1,8 146,9
PI(1,2)-18 229,0 158,0 12,8 66,5 69,5 3,3 1,8 135,9
PI(1,2)-19 221,0 139,0 12,0 69,5 72,0 2,5 2,3 128,3
PI(1,2)-20 251,0 152,5 15,6 68,5 70,0 1,5 1,4 134,8
PI(1,2)-21 249,0 160,0 5,5 69,5 71,5 2,0 1,6 137,1
PI(1,2)-22 260,0 163,0 12,2 69,0 71,5 2,5 1,6 140,6
PI(1,2)-23 256,0 153,0 11,6 66,3 70,2 3,9 1,7 169,3
PI(1,2)-24 252,0 155,0 10,2 65,2 67,4 2,2 2,4 152,7
PI(1,2)-25 262,0 169,5 7,0 69,5 71,5 2,0 1,9 159,2
PI(1,2)-26 248,0 154,0 6,6 68,0 70,0 2,0 2,1 130,6
PI(1,2)-27 267,0 149,0 18,3 68,4 71,6 3,1 1,9 163,1
PI(1,2)-28 217,0 133,5 10,5 65,5 69,0 3,5 2,2 136,2
PI(1,2)-29 253,0 150,0 10,3 67,5 69,5 2,0 2,0 144,5
PI(1,2)-30 260,0 167,0 8,8 70,5 73,0 2,5 2,0 147,5
PI(1,2)-31 247,0 144,0 10,0 68,5 70,0 1,5 2,1 144,0
PI(1,2)-32 233,0 143,5 13,3 67,5 69,0 1,5 1,4 114,5
PI(1,2)-33 248,0 155,5 8,1 68,0 70,5 2,5 2,2 130,0
PI(1,2)-34 256,0 161,0 8,9 67,5 68,5 1,0 2,6 126,9
PI(1,2)-35 269,0 171,0 7,4 69,0 71,5 2,5 1,0 117,3
PI(1,2)-36 237,0 155,0 8,1 71,0 73,5 2,5 1,7 155,2
PI(1,2)-37 222,0 141,0 9,9 67,0 69,0 2,0 1,7 137,3
PI(1,2)-38 244,0 155,5 11,5 69,0 70,0 1,0 1,7 126,8
PI(1,2)-39 264,0 154,0 11,9 73,0 77,0 4,0 1,4 126,0

110
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-40 256,0 179,0 3,7 72,1 74,6 2,5 1,7 156,4
PI(1,2)-41 244,0 151,0 5,5 68,5 71,5 3,0 1,2 132,8
PI(1,2)-42 249,0 150,0 7,4 75,0 77,0 2,0 1,7 140,8
PI(1,2)-43 243,0 162,0 11,9 66,5 68,5 2,0 1,5 124,6
PI(1,2)-44 244,0 159,0 6,8 71,0 72,4 1,4 1,0 144,8
PI(1,2)-45 231,0 149,0 14,0 68,5 70,5 2,0 1,8 145,6
PI(1,2)-46 241,0 155,0 7,0 69,5 71,0 1,5 1,8 128,8
PI(1,2)-47 261,0 165,0 10,9 72,5 76,0 3,5 1,9 111,0
PI(1,2)-48 252,0 157,0 11,9 68,0 70,5 2,5 2,2 127,5
PI(1,2)-49 260,0 168,0 5,7 68,3 70,2 1,9 1,0 146,4
PI(1,2)-50 258,0 161,0 16,8 67,0 70,0 3,0 1,8 133,6
PI(1,2)-51 255,0 163,0 9,4 70,0 72,5 2,5 1,7 123,2
PI(1,2)-52 257,0 178,0 8,4 68,2 69,2 1,0 3,0 135,4
PI(1,2)-53 249,0 154,0 15,5 69,5 73,0 3,5 1,8 127,7
PI(1,2)-54 239,0 133,0 14,4 65,5 68,0 2,5 1,8 151,0
PI(1,2)-55 233,0 134,5 16,8 70,0 72,5 2,5 1,1 145,1
PI(1,2)-56 240,0 147,0 13,0 71,0 76,5 5,5 1,3 145,8
PI(1,2)-57 250,0 147,0 14,2 67,5 70,5 3,0 2,1 114,6
PI(1,2)-58 251,0 152,5 11,5 68,0 70,5 2,5 2,1 136,7
PI(1,2)-59 235,0 156,0 3,7 68,2 72,9 4,7 1,1 172,4
PI(1,2)-60 258,0 141,0 11,3 68,5 70,5 2,0 1,5 130,0
PI(1,2)-61 270,0 167,5 11,5 68,5 70,5 2,0 1,3 129,9
PI(1,2)-62 259,0 159,5 10,9 71,5 74,0 2,5 1,8 132,1
PI(1,2)-63 233,0 141,0 13,4 68,5 73,0 4,5 1,6 127,3
PI(1,2)-64 234,0 143,5 11,5 70,0 75,5 5,5 1,4 133,4
PI(1,2)-65 234,0 139,0 11,9 70,5 74,0 3,5 1,7 155,4
PI(1,2)-66 244,0 146,5 11,0 69,0 73,0 4,0 1,8 114,5
PI(1,2)-67 258,0 162,5 14,0 67,5 70,0 2,5 1,6 137,5
PI(1,2)-68 236,0 158,0 1,2 71,9 74,5 2,5 1,1 167,4
PI(1,2)-69 257,0 152,0 18,8 70,0 72,0 2,0 2,0 129,7
PI(1,2)-70 266,0 168,0 13,2 69,0 69,5 0,5 1,5 148,5
PI(1,2)-71 261,0 163,0 3,8 66,8 69,4 2,6 2,1 150,2
PI(1,2)-72 243,0 161,0 6,8 67,0 68,9 1,9 1,1 168,1
PI(1,2)-73 219,0 146,0 5,7 65,6 69,9 4,3 1,3 177,7
PI(1,2)-74 244,0 144,5 10,7 68,5 72,0 3,5 1,3 143,4
PI(1,2)-75 214,0 120,5 12,4 68,5 71,0 2,5 1,1 145,0
PI(1,2)-76 248,0 156,0 14,1 67,5 69,5 2,0 1,7 125,8
PI(1,2)-77 248,0 146,0 12,5 69,5 72,0 2,5 1,7 148,0
PI(1,2)-78 241,0 129,5 13,0 69,5 70,5 1,0 1,6 122,1
PI(1,2)-79 253,0 161,5 13,0 69,5 73,0 3,5 2,2 135,0
PI(1,2)-80 271,0 155,0 10,5 68,0 69,5 1,5 1,5 123,6
Anexo 5. (Continuação)

111
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-81 255,0 162,0 13,8 72,6 74,5 1,9 1,3 142,7
PI(1,2)-82 222,0 138,0 7,4 68,0 70,5 2,5 1,2 124,3
PI(1,2)-83 221,0 144,0 11,5 69,0 72,5 3,5 1,5 114,4
PI(1,2)-84 226,0 130,5 4,1 71,0 72,5 1,5 2,1 150,6
PI(1,2)-85 238,0 152,0 7,4 69,0 71,5 2,5 2,6 140,8
PI(1,2)-86 241,0 152,5 11,3 72,0 74,0 2,0 1,7 131,4
PI(1,2)-87 240,0 154,5 11,0 69,0 71,0 2,0 1,3 114,1
PI(1,2)-88 262,0 167,0 11,3 67,5 69,0 1,5 1,7 136,2
PI(1,2)-89 242,0 159,0 8,9 67,5 70,5 3,0 1,2 121,7
PI(1,2)-90 245,0 157,5 10,8 69,0 70,0 1,0 1,2 126,0
PI(1,2)-91 231,0 130,0 15,3 66,5 69,0 2,5 1,7 134,8
PI(1,2)-92 250,0 151,5 18,9 70,5 77,0 6,5 1,3 128,9
PI(1,2)-93 248,0 145,0 16,4 71,5 73,5 2,0 1,3 134,6
PI(1,2)-94 252,0 142,0 8,1 69,5 70,5 1,0 1,4 165,0
PI(1,2)-95 241,0 146,5 14,0 70,0 72,5 2,5 1,5 124,8
PI(1,2)-96 260,0 156,0 17,4 70,5 72,0 1,5 1,8 135,1
PI(1,2)-97 254,0 150,0 11,5 68,5 70,0 1,5 1,4 113,1
PI(1,2)-98 256,0 158,0 16,4 69,5 71,0 1,5 1,7 129,2
PI(1,2)-99 237,5 148,0 10,5 67,5 69,0 1,5 1,1 132,9
PI(1,2)-100 249,0 125,0 6,60 67,6 70,7 3,1 1,5 181,1
PI(1,2)-101 237,0 145,0 8,8 68,0 70,0 2,0 1,1 118,9
PI(1,2)-102 225,0 167,0 8,7 70,7 77,1 6,4 1,0 150,1
PI(1,2)-103 240,0 139,0 14,9 70,5 73,0 2,5 1,1 133,2
PI(1,2)-104 257,5 147,0 18,6 69,0 72,0 3,0 1,5 133,4
PI(1,2)-105 282,0 183,5 8,9 69,5 70,5 1,0 2,6 108,6
PI(1,2)-106 272,0 156,0 2,9 69,4 70,3 0,9 2,0 161,3
PI(1,2)-107 228,0 150,0 2,5 69,8 74,3 4,5 1,7 163,4
PI(1,2)-108 244,0 152,0 8,1 70,0 72,5 2,5 1,6 119,0
PI(1,2)-109 241,0 145,0 8,5 72,6 73,3 0,6 1,7 166,2
PI(1,2)-110 246,0 147,0 5,6 70,4 72,7 2,3 2,1 167,5
PI(1,2)-111 246,5 143,0 8,8 68,0 71,0 3,0 1,3 149,0
PI(1,2)-112 265,5 165,5 6,6 67,0 69,0 2,0 1,4 123,9
PI(1,2)-113 277,0 170,0 8,1 68,5 70,5 2,0 1,4 129,5
PI(1,2)-114 263,5 163,0 10,9 67,0 68,0 1,0 2,0 152,4
PI(1,2)-115 262,0 154,0 10,0 68,0 69,5 1,5 1,3 135,8
PI(1,2)-116 261,5 164,0 12,5 67,0 68,5 1,5 1,2 119,7
PI(1,2)-117 237,0 147,0 5,4 70,9 75,4 4,5 2,1 163,7
PI(1,2)-118 224,5 134,5 8,8 69,0 71,0 2,0 1,4 122,0
PI(1,2)-119 242,5 151,5 9,1 68,5 71,0 2,5 2,0 107,8
PI(1,2)-120 248,5 151,0 5,5 69,0 74,0 5,0 1,9 118,0
PI(1,2)-121 243,5 128,0 24,7 70,0 71,5 1,5 1,6 143,3
Anexo 5. (Continuação)

112
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-122 267,0 159,0 16,9 69,5 70,5 1,0 2,4 165,0
PI(1,2)-123 251,5 154,5 13,6 70,5 72,0 1,5 2,0 148,0
PI(1,2)-124 229,0 166,0 3,9 72,6 73,9 1,4 2,2 158,2
PI(1,2)-125 279,0 175,5 7,4 68,5 70,5 2,0 1,9 120,4
PI(1,2)-126 245,5 139,0 15,9 68,5 69,5 1,0 1,5 126,7
PI(1,2)-127 239,5 131,5 13,2 68,5 70,0 1,5 1,3 139,9
PI(1,2)-128 261,0 153,0 12,4 68,5 69,0 1,0 2,2 151,7
PI(1,2)-129 241,0 157,0 4,9 67,8 69,6 1,8 1,6 178,3
PI(1,2)-130 250,0 149,0 5,5 66,5 68,5 2,0 1,3 137,7
PI(1,2)-131 257,0 139,0 15,7 70,5 74,0 3,5 1,4 119,4
PI(1,2)-132 245,5 152,5 9,6 70,5 75,5 5,0 2,0 128,9
PI(1,2)-133 264,5 162,5 5,5 69,0 71,5 2,5 2,0 148,5
PI(1,2)-134 255,0 163,5 12,8 69,5 76,5 7,0 2,5 126,9
PI(1,2)-135 237,5 146,0 12,9 68,0 70,5 2,5 1,4 129,0
PI(1,2)-136 223,0 121,0 11,7 70,9 72,5 1,6 2,3 145,8
PI(1,2)-137 248,0 143,5 15,5 67,5 69,5 2,0 1,8 148,4
PI(1,2)-138 260,0 154,0 4,9 72,6 74,6 2,1 2,1 175,1
PI(1,2)-139 256,5 153,5 12,4 68,5 70,5 2,0 1,4 137,4
PI(1,2)-140 255,5 155,5 10,0 71,5 74,5 3,0 1,8 112,0
PI(1,2)-141 236,0 159,0 5,0 66,0 68,8 2,7 2,3 136,3
PI(1,2)-142 253,0 144,5 10,5 66,5 68,5 2,0 1,2 132,7
PI(1,2)-143 257,0 164,5 6,6 68,0 69,5 1,5 2,2 112,7
PI(1,2)-144 243,5 135,0 17,9 69,0 70,0 1,0 2,4 118,3
PI(1,2)-145 224,5 135,0 8,1 69,0 73,0 4,0 1,6 148,8
PI(1,2)-146 266,0 157,5 8,1 68,0 69,5 1,5 1,2 132,5
PI(1,2)-147 249,0 182,0 6,0 66,1 68,6 2,6 2,1 156,5
PI(1,2)-148 249,0 148,5 17,3 65,5 68,0 2,5 1,9 111,4
PI(1,2)-149 243,0 148,0 15,3 69,0 71,0 2,0 2,0 117,3
PI(1,2)-150 247,0 151,5 13,5 68,5 70,0 1,5 1,7 135,6
PI(1,2)-151 255,5 156,5 12,2 69,5 71,5 2,0 2,0 111,5
PI(1,2)-152 248,5 138,5 11,9 66,5 67,5 1,0 1,9 143,9
PI(1,2)-153 230,5 139,0 11,0 68,0 71,0 3,0 1,5 140,4
PI(1,2)-154 201,5 112,0 12,8 69,5 72,5 3,0 1,3 115,0
PI(1,2)-155 254,0 139,0 7,9 69,6 73,4 3,7 1,7 171,1
PI(1,2)-156 259,0 153,0 14,7 68,0 71,0 3,0 1,6 124,1
PI(1,2)-157 266,0 151,5 13,4 68,0 70,0 2,0 1,2 134,9
PI(1,2)-158 269,0 166,5 11,9 71,0 72,5 1,5 1,6 123,3
PI(1,2)-159 261,5 157,0 16,2 69,5 71,0 1,5 1,6 156,1
PI(1,2)-160 252,0 151,5 15,5 70,5 71,5 1,0 1,4 135,1
PI(1,2)-161 258,0 163,0 8,8 68,0 70,0 2,0 1,6 156,4
PI(1,2)-162 247,0 144,0 8,9 68,5 70,5 2,0 1,3 131,8
PI(1,2)-163 242,0 127,0 4,8 71,5 74,9 3,4 1,0 162,2
Anexo 5. (Continuação)

113
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-164 250,0 152,0 16,2 69,0 70,0 1,0 1,4 125,2
PI(1,2)-165 254,5 145,5 14,9 69,5 71,5 2,0 1,3 125,2
PI(1,2)-166 249,0 142,5 12,5 68,0 69,5 1,5 1,3 137,2
PI(1,2)-167 260,0 159,5 14,4 68,0 70,0 2,0 2,3 117,5
PI(1,2)-168 274,5 168,0 12,8 68,0 71,0 3,0 1,2 127,6
PI(1,2)-169 263,5 161,5 8,8 67,0 69,5 2,5 1,4 134,6
PI(1,2)-170 254,0 155,5 12,0 68,0 70,0 2,0 1,9 106,5
PI(1,2)-171 239,5 155,5 15,3 67,0 70,0 3,0 2,2 122,7
PI(1,2)-172 237,0 144,5 13,4 67,0 69,5 2,5 1,5 129,0
PI(1,2)-173 249,5 159,5 13,9 68,0 69,0 1,0 1,7 132,4
PI(1,2)-174 254,0 142,0 2,6 70,3 73,1 2,9 1,2 167,7
PI(1,2)-175 251,0 153,0 6,6 67,0 69,5 2,5 1,5 167,6
PI(1,2)-176 255,5 153,0 13,5 68,0 71,0 3,0 1,6 148,6
PI(1,2)-177 233,5 136,0 13,5 67,0 68,0 1,0 1,7 125,6
PI(1,2)-178 243,0 144,0 13,6 67,5 69,5 2,0 1,4 155,8
PI(1,2)-179 236,0 155,0 10,7 68,5 71,0 2,5 1,7 132,0
PI(1,2)-180 246,0 146,5 12,0 67,0 70,0 3,0 1,9 151,8
PI(1,2)-181 239,0 123,0 12,5 70,9 75,6 4,7 1,4 164,2
PI(1,2)-182 241,0 147,5 13,2 66,0 70,0 4,0 1,7 164,2
PI(1,2)-183 236,5 144,0 12,0 70,0 74,0 4,0 1,6 144,5
PI(1,2)-184 234,0 135,5 15,7 66,5 68,5 2,0 1,6 141,9
PI(1,2)-185 260,5 167,0 11,0 67,0 69,0 2,0 1,3 136,4
PI(1,2)-186 217,0 167,0 7,3 66,3 69,5 3,2 1,3 155,3
PI(1,2)-187 253,0 156,5 13,0 69,0 71,0 2,0 1,6 155,3
PI(1,2)-188 217,0 142,0 6,9 60,9 64,7 3,7 2,0 159,7
PI(1,2)-189 232,0 150,0 10,4 70,0 75,5 5,5 1,6 179,1
PI(1,2)-190 214,0 120,5 14,1 66,5 69,5 3,0 1,6 158,2
PI(1,2)-191 223,0 145,0 4,6 67,4 70,2 2,8 2,3 172,6
PI(1,2)-192 240,5 150,0 13,4 69,0 74,0 5,0 1,4 172,6
PI(1,2)-193 256,0 160,0 9,22 69,7 71,7 1,9 1,7 162,3
PI(1,2)-194 223,0 159,0 1,44 70,6 73,7 3,1 2,7 141,5
PI(1,2)-195 210,0 144,0 7,9 67,6 69,6 2,0 1,8 165,5
PI(1,2)-196 225,0 140,0 1,7 68,9 70,9 2,1 1,8 158,2
Média 245,2 150,4 10,9 68,8 71,3 2,4 1,6 139,1
AV 201,5-
282
112-
183,5
1,2-
24,7
60,9-
75
64,7-
77,1
0,5-
7
1-
3,0
106,5-
181,1
CV(%) 4,36 4,19 31,4 1,61 2,6 50,5 8,9 11,64
(1) Dados transformados para arcoseno 100)5,0( x .
Anexo 5. (Continuação)

114
Anexo 6. Médias de altura de planta (AP), altura de espiga (AE), plantas acamadas e
quebradas (A+Q), florescimento masculino (FM), florescimento feminino (FF), intervalo
entre florescimentos (IF), stay-green (SG), massa de grãos (MG) de 196 progênies
interpopulacioanis (PI(1,2)) avaliadas em Mococa, na safrinha de 2012.
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-1 254,0 135,0 10,7 70,0 71,0 1,0 1,6 129,4
PI(1,2)-2 237,5 125,0 11,9 68,0 71,0 3,0 1,6 119,0
PI(1,2)-3 242,5 125,0 13,9 69,0 69,5 0,5 1,4 106,8
PI(1,2)-4 260,0 155,0 10,7 69,0 69,0 0,0 1,6 108,6
PI(1,2)-5 235,0 125,0 13,5 68,0 71,0 3,0 1,1 102,7
PI(1,2)-6 265,0 155,0 10,3 68,0 69,5 1,5 2,0 116,9
PI(1,2)-7 240,0 140,0 8,9 69,0 70,5 1,5 1,5 118,2
PI(1,2)-8 250,0 145,0 11,5 68,0 69,5 1,5 1,2 122,2
PI(1,2)-9 255,0 135,0 10,3 68,0 71,0 3,0 1,3 131,0
PI(1,2)-10 262,5 155,0 7,4 68,0 69,0 1,0 1,3 109,2
PI(1,2)-11 250,0 132,5 11,3 68,0 68,5 0,5 1,4 136,1
PI(1,2)-12 232,5 137,5 11,5 68,0 71,5 3,5 1,4 109,4
PI(1,2)-13 247,5 145,0 11,3 68,0 71,0 3,0 1,4 92,1
PI(1,2)-14 260,0 130,0 8,1 68,0 69,5 1,5 1,5 112,5
PI(1,2)-15 237,5 145,0 12,2 68,0 69,0 1,0 1,1 107,9
PI(1,2)-16 262,5 147,5 12,9 68,0 71,0 3,0 1,5 112,8
PI(1,2)-17 240,0 132,0 13,1 65,9 69,5 3,5 1,3 131,7
PI(1,2)-18 235,0 145,0 11,9 65,7 69,2 3,5 1,8 113,0
PI(1,2)-19 230,0 145,0 5,5 68,0 71,0 3,0 1,4 121,0
PI(1,2)-20 245,0 135,0 10,3 69,0 70,0 1,0 1,8 129,0
PI(1,2)-21 245,0 135,0 15,3 69,0 71,5 2,5 1,8 122,2
PI(1,2)-22 242,5 140,0 8,1 70,0 72,5 2,5 1,2 126,2
PI(1,2)-23 236,0 139,0 18,1 66,1 69,3 3,1 1,2 125,1
PI(1,2)-24 233,0 133,0 12,0 69,1 72,7 3,6 1,6 115,9
PI(1,2)-25 237,5 125,0 9,9 69,0 71,5 2,5 2,0 122,6
PI(1,2)-26 235,0 125,0 12,2 69,0 70,0 1,0 2,3 134,5
PI(1,2)-27 241,0 142,0 9,2 66,9 69,5 2,6 1,4 126,3
PI(1,2)-28 225,0 122,5 11,0 69,0 71,5 2,5 1,6 118,6
PI(1,2)-29 255,0 135,0 8,1 68,0 69,0 1,0 1,3 121,7
PI(1,2)-30 242,5 132,5 11,5 70,0 71,5 1,5 1,2 127,9
PI(1,2)-31 242,5 137,5 8,1 68,0 71,0 3,0 1,3 114,3
PI(1,2)-32 255,0 152,5 10,3 69,0 73,5 4,5 1,9 119,8
PI(1,2)-33 262,5 140,0 9,6 68,0 70,0 2,0 1,6 123,5
PI(1,2)-34 230,0 125,0 12,8 68,0 69,0 1,0 1,5 117,7
PI(1,2)-35 240,0 137,5 8,8 70,0 71,5 1,5 1,6 103,2
PI(1,2)-36 250,0 140,0 10,8 71,0 73,5 2,5 1,3 112,4
PI(1,2)-37 247,5 135,0 7,4 70,0 71,5 1,5 1,4 121,5
PI(1,2)-38 245,0 135,0 11,0 71,0 73,5 2,5 1,3 121,1

115
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-39 242,5 135,0 11,5 70,0 70,0 0,0 1,4 121,9
PI(1,2)-40 219,0 132,0 4,9 69,0 71,5 2,5 1,5 115,4
PI(1,2)-41 242,5 145,0 8,9 68,0 69,0 1,0 1,9 105,4
PI(1,2)-42 235,0 130,0 10,5 68,0 71,0 3,0 1,4 121,7
PI(1,2)-43 257,5 142,5 8,8 70,0 71,0 1,0 1,4 117,2
PI(1,2)-44 259,0 135,0 19,3 68,2 71,7 3,5 1,3 117,1
PI(1,2)-45 232,5 127,5 17,4 69,0 70,0 1,0 1,3 111,9
PI(1,2)-46 237,5 145,0 4,1 68,0 69,5 1,5 1,8 106,9
PI(1,2)-47 245,0 120,0 11,5 68,0 70,5 2,5 1,5 112,3
PI(1,2)-48 227,5 110,0 9,4 68,0 71,0 3,0 1,6 126,9
PI(1,2)-49 248,0 144,0 14,8 69,1 69,8 0,7 2,2 113,5
PI(1,2)-50 265,0 140,0 7,4 69,0 71,0 2,0 1,8 128,0
PI(1,2)-51 240,0 132,5 9,6 68,0 71,0 3,0 2,3 105,4
PI(1,2)-52 244,0 136,0 13,5 71,6 72,9 1,4 1,5 149,1
PI(1,2)-53 252,5 145,0 10,5 70,0 71,5 1,5 1,4 116,1
PI(1,2)-54 235,0 135,0 12,2 68,0 71,5 3,5 1,5 118,0
PI(1,2)-55 252,5 137,5 10,3 69,0 71,0 2,0 1,2 103,8
PI(1,2)-56 225,0 135,0 14,9 70,0 69,0 1,0 1,4 111,9
PI(1,2)-57 237,5 155,0 7,0 68,0 70,0 2,0 1,6 113,6
PI(1,2)-58 252,5 155,0 9,9 70,0 71,0 1,0 1,5 119,4
PI(1,2)-59 243,0 135,0 21,0 67,9 69,5 1,6 0,9 110,8
PI(1,2)-60 239,0 125,0 7,4 68,0 70,0 2,0 1,3 107,8
PI(1,2)-61 235,0 135,0 6,6 68,0 69,0 1,0 1,3 115,2
PI(1,2)-62 242,5 152,5 13,2 70,0 71,0 1,0 1,5 110,3
PI(1,2)-63 240,0 122,5 10,0 68,0 71,5 3,5 1,5 112,0
PI(1,2)-64 262,5 152,5 12,0 68,0 70,5 2,5 1,3 103,8
PI(1,2)-65 250,0 145,0 12,8 68,0 70,0 2,0 1,3 118,5
PI(1,2)-66 230,0 130,0 8,8 68,0 71,0 3,0 2,0 109,2
PI(1,2)-67 237,5 130,0 13,5 69,0 71,5 2,5 1,5 119,6
PI(1,2)-68 218,0 147,0 14,3 70,2 72,6 2,4 1,1 121,3
PI(1,2)-69 255,0 150,0 11,5 70,0 72,5 2,5 1,2 115,6
PI(1,2)-70 237,5 135,0 10,0 70,0 72,5 2,5 1,5 130,9
PI(1,2)-71 252,0 139,0 6,8 68,3 71,2 2,9 1,7 128,9
PI(1,2)-72 261,0 147,0 16,1 71,6 71,4 -0,2 1,5 118,1
PI(1,2)-73 237,0 132,0 11,6 71,3 73,9 2,6 1,3 123,1
PI(1,2)-74 245,0 150,0 10,5 70,0 70,0 0,0 1,4 122,6
PI(1,2)-75 245,0 137,5 10,3 69,0 71,0 2,0 1,5 110,8
PI(1,2)-76 250,0 135,0 9,1 68,0 69,0 1,0 1,4 109,6
PI(1,2)-77 257,5 152,5 9,6 69,0 71,0 2,0 1,7 119,3
PI(1,2)-78 245,0 144,0 10,3 68,0 72,0 4,0 1,6 114,3
PI(1,2)-79 260,0 145,0 7,4 71,0 71,5 0,5 2,0 122,8
Anexo 6. (Continuação)

116
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-80 242,5 137,5 8,1 70,0 71,5 1,5 1,5 109,1
PI(1,2)-81 250,0 145,0 20,2 68,3 69,6 1,3 1,4 106,3
PI(1,2)-82 237,5 136,0 9,4 70,0 72,0 2,0 1,4 103,6
PI(1,2)-83 245,0 140,0 9,6 72,0 72,5 0,5 1,6 125,6
PI(1,2)-84 230,0 122,5 16,5 68,0 70,0 2,0 1,7 125,8
PI(1,2)-85 240,0 137,5 11,9 68,0 71,0 3,0 2,0 124,0
PI(1,2)-86 255,0 135,0 16,4 68,0 71,0 3,0 2,4 119,8
PI(1,2)-87 240,0 135,0 11,5 68,0 71,0 3,0 1,2 120,9
PI(1,2)-88 237,5 135,0 17,7 69,0 71,0 2,0 1,3 110,2
PI(1,2)-89 250,0 150,0 11,0 68,0 73,5 5,5 1,5 125,7
PI(1,2)-90 245,0 135,0 5,5 70,0 72,5 2,5 1,7 111,7
PI(1,2)-91 230,0 125,0 10,0 70,0 72,5 2,5 1,8 112,2
PI(1,2)-92 237,5 132,5 11,0 69,0 70,0 1,0 1,3 116,0
PI(1,2)-93 255,0 137,5 12,2 71,0 72,5 1,5 1,5 125,7
PI(1,2)-94 250,0 132,5 4,1 68,0 71,0 3,0 1,1 121,2
PI(1,2)-95 230,0 127,5 10,9 71,0 73,5 2,5 1,6 106,0
PI(1,2)-96 227,5 117,5 10,5 68,0 73,5 5,5 2,2 108,4
PI(1,2)-97 235,0 125,0 11,9 70,0 72,5 2,5 1,3 115,8
PI(1,2)-98 247,5 135,0 11,3 70,0 71,5 1,5 1,3 115,7
PI(1,2)-99 235,0 127,5 11,5 68,0 71,0 3,0 1,3 120,0
PI(1,2)-100 244,0 126,0 18,1 71,3 72,3 0,9 1,3 107,5
PI(1,2)-101 225,0 132,5 11,0 70,0 72,5 2,5 1,7 121,3
PI(1,2)-102 222,0 121,0 15,7 68,3 71,5 3,3 1,3 98,41
PI(1,2)-103 240,0 150,0 10,5 69,0 71,0 2,0 1,2 104,3
PI(1,2)-104 247,5 140,0 8,8 71,0 72,5 1,5 1,3 118,3
PI(1,2)-105 232,5 135,0 16,2 70,0 72,5 2,5 1,3 101,2
PI(1,2)-106 233,0 128,0 3,1 71,0 73,1 2,1 1,2 113,8
PI(1,2)-107 216,0 117,0 16,4 67,7 71,1 3,3 1,8 124,3
PI(1,2)-108 227,5 125,0 4,1 73,5 75,5 1,5 1,3 108,2
PI(1,2)-109 219,0 109,0 14,2 68,5 69,4 0,9 1,3 114,8
PI(1,2)-110 207,0 129,0 7,7 68,6 71,8 3,2 1,3 122,3
PI(1,2)-111 221,0 130,0 13,3 69,0 71,5 2,5 1,5 114,8
PI(1,2)-112 225,0 122,5 12,8 72,5 73,5 1,5 1,5 110,4
PI(1,2)-113 242,5 140,0 10,5 71,0 73,5 2,5 1,2 125,1
PI(1,2)-114 249,0 130,0 14,6 68,0 69,0 1,0 1,4 125,6
PI(1,2)-115 230,0 125,0 6,6 68,0 71,0 3,0 1,3 125,6
PI(1,2)-116 235,0 125,0 8,8 68,0 70,0 2,0 1,4 117,4
PI(1,2)-117 245,0 130,0 18,6 69,4 70,1 1,0 2,1 132,5
PI(1,2)-118 245,0 140,0 9,9 69,0 71,0 2,0 1,2 118,9
PI(1,2)-119 240,0 125,0 8,1 70,0 71,0 1,0 2,3 102,9
PI(1,2)-120 240,0 125,0 13,3 70,0 71,5 1,5 1,6 119,3
PI(1,2)-121 240,0 140,0 12,2 68,0 71,0 3,0 1,7 129,3
Anexo 6. (Continuação)

117
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-122 222,5 130,0 12,5 70,0 73,5 3,5 1,6 124,8
PI(1,2)-123 235,0 117,5 14,7 72,5 74,5 2,0 1,3 124,3
PI(1,2)-124 240,0 133,0 9,6 68,0 72,1 4,1 1,7 125,4
PI(1,2)-125 230,0 125,0 14,1 70,0 71,5 1,5 1,5 121,5
PI(1,2)-126 235,0 140,0 8,8 70,0 73,5 3,5 1,5 105,9
PI(1,2)-127 245,0 132,5 12,5 72,0 73,5 1,5 1,2 121,2
PI(1,2)-128 255,0 145,0 11,9 68,0 68,5 0,5 1,7 127,1
PI(1,2)-129 249,0 138,0 16,6 68,3 71,4 3,1 1,6 118,6
PI(1,2)-130 245,0 125,0 10,3 69,0 71,5 2,5 1,9 127,2
PI(1,2)-131 245,0 142,5 10,9 71,5 73,5 2,0 1,3 128,0
PI(1,2)-132 220,0 120,0 10,7 69,0 71,0 2,0 1,4 131,4
PI(1,2)-133 245,0 135,0 9,6 68,0 71,5 3,5 2,0 120,0
PI(1,2)-134 245,0 142,5 12,9 70,0 72,5 2,5 1,3 112,7
PI(1,2)-135 235,0 127,5 16,4 69,0 73,5 4,5 1,5 112,3
PI(1,2)-136 232,0 139,0 13,3 70,5 74,6 4,2 1,7 108,4
PI(1,2)-137 230,0 127,5 8,1 68,0 69,0 1,0 1,9 112,4
PI(1,2)-138 227,0 132,0 11,4 68,6 70,9 2,3 1,2 130,4
PI(1,2)-139 230,0 135,0 5,5 70,0 71,0 1,0 1,6 118,3
PI(1,2)-140 252,5 145,0 9,1 68,0 70,0 2,0 1,2 110,3
PI(1,2)-141 258,0 131,0 13,6 71,4 72,1 0,6 2,1 122,9
PI(1,2)-142 267,5 162,5 11,3 71,0 73,5 2,5 1,3 104,0
PI(1,2)-143 240,0 127,5 8,1 69,0 69,5 0,5 1,4 132,8
PI(1,2)-144 240,0 147,5 16,5 69,0 70,0 1,0 1,6 121,8
PI(1,2)-145 241,0 135,0 9,4 69,0 71,0 2,0 1,4 114,5
PI(1,2)-146 235,0 130,0 8,8 70,0 70,0 0,0 1,2 121,1
PI(1,2)-147 231,0 126,0 17,8 74,6 74,4 -0,2 2,4 109,7
PI(1,2)-148 245,0 127,5 9,9 69,0 71,5 2,5 1,3 106,6
PI(1,2)-149 245,0 135,0 14,0 70,0 73,5 3,5 1,4 100,9
PI(1,2)-150 247,5 135,0 15,9 68,0 71,0 3,0 1,2 95,2
PI(1,2)-151 235,0 135,0 7,4 71,5 71,5 0,0 1,3 110,1
PI(1,2)-152 245,0 137,5 18,3 68,0 68,5 0,5 1,3 120,9
PI(1,2)-153 255,0 142,5 15,7 70,0 73,5 3,5 2,2 116,0
PI(1,2)-154 245,0 135,0 9,6 71,0 71,0 0,0 1,1 127,0
PI(1,2)-155 243,0 121,0 12,9 68,1 71,8 3,7 1,8 103,1
PI(1,2)-156 260,0 135,0 16,4 70,0 72,5 2,5 1,6 121,6
PI(1,2)-157 252,5 140,0 11,3 71,5 73,5 2,0 1,4 116,5
PI(1,2)-158 250,0 147,5 10,0 69,0 72,5 3,5 1,2 111,5
PI(1,2)-159 265,0 155,0 8,8 68,0 70,5 2,5 1,7 121,3
PI(1,2)-160 262,5 135,0 8,9 68,0 69,0 1,0 1,2 111,0
PI(1,2)-161 235,0 135,0 12,9 71,0 72,5 1,5 2,2 109,8
PI(1,2)-162 255,0 135,0 9,9 70,0 72,5 2,5 1,5 124,9
PI(1,2)-163 235,0 139,0 19,8 68,3 73,8 5,5 1,3 124,9
Anexo 6. (Continuação)

118
Progênies AP
(cm)
AE
(cm)
A+Q(1)
(arcoseno) FM
(d.a.s.)
FF
(d.a.s.)
IF
(dias)
SG
(nota)
MG
(g planta-1
)
PI(1,2)-164 235,0 120,0 12,9 69,0 70,0 1,0 1,4 122,9
PI(1,2)-165 250,0 145,0 14,0 71,5 73,5 2,0 1,1 121,1
PI(1,2)-166 252,5 130,0 8,1 68,0 71,5 3,5 1,2 134,6
PI(1,2)-167 245,0 142,5 14,0 68,0 70,0 2,0 1,3 114,7
PI(1,2)-168 235,0 125,0 9,4 68,0 69,0 1,0 1,4 120,4
PI(1,2)-169 240,0 135,0 11,9 68,0 71,0 3,0 1,3 97,8
PI(1,2)-170 232,5 135,0 8,1 71,5 72,5 1,0 1,4 115,4
PI(1,2)-171 235,0 140,0 14,6 68,0 70,0 2,0 1,4 114,0
PI(1,2)-172 232,5 125,0 13,4 68,0 69,0 1,0 1,3 120,2
PI(1,2)-173 225,0 132,5 9,4 68,0 70,5 2,5 1,2 118,9
PI(1,2)-174 234,0 139,0 20,9 67,7 67,7 -0,01 1,0 97,44
PI(1,2)-175 235,0 117,5 12,0 70,0 71,5 1,5 1,4 114,0
PI(1,2)-176 235,0 125,0 5,5 70,0 71,5 1,5 1,9 117,8
PI(1,2)-177 245,0 132,5 18,3 68,0 69,0 1,0 1,5 125,1
PI(1,2)-178 267,5 155,0 9,6 68,0 70,0 2,0 1,2 120,6
PI(1,2)-179 242,5 135,0 11,5 71,5 72,5 1,0 1,3 117,6
PI(1,2)-180 245,0 140,0 12,5 68,0 70,0 2,0 1,4 131,3
PI(1,2)-181 236,0 129,0 15,9 70,8 69,7 -1,2 1,0 121,3
PI(1,2)-182 235,0 130,0 13,2 68,0 70,0 2,0 1,2 117,8
PI(1,2)-183 237,5 125,0 4,1 69,0 71,5 2,5 1,4 110,3
PI(1,2)-184 270,0 145,0 11,3 68,0 70,0 2,0 1,6 122,9
PI(1,2)-185 245,0 145,0 11,5 69,0 71,0 2,0 1,4 119,0
PI(1,2)-186 223,0 123,0 8,1 69,4 72,5 3,1 1,5 128,6
PI(1,2)-187 245,0 135,0 8,1 70,0 71,5 1,5 1,4 111,6
PI(1,2)-188 234,0 118,0 9,8 72,6 74,8 2,3 1,6 101,5
PI(1,2)-189 247,0 145,0 12,4 72,4 74,4 2,0 1,6 103,7
PI(1,2)-190 225,0 116,0 9,9 69,0 71,5 2,5 1,2 129,8
PI(1,2)-191 242,0 133,0 16,5 68,8 71,1 2,2 1,6 110,7
PI(1,2)-192 242,5 132,5 12,5 71,5 73,5 2,0 1,3 108,2
PI(1,2)-193 255,0 129,0 14,9 69,7 72,8 3,2 1,3 110,1
PI(1,2)-194 238,0 147,0 11,8 72,0 76,3 4,3 1,5 114,0
PI(1,2)-195 249,0 117,0 9,3 73,7 76,3 2,6 1,4 118,2
PI(1,2)-196 238,0 121,0 16,0 69,8 72,4 2,6 1,3 110,9 Médias 241,8 135,0 11,44 69,2 71,3 2,1 1,5 117,0
AV 207-
270
109-
162,5
3,1-
21
65,7-
74,6
67,7-
76,3
-1,2-
5,5
0,9-
2,4
92,1-
149,1
CV(%) 3,31 5,97 24,2 1,74 2,08 59,5 6,6 8,64
(1) Dados transformados para arcoseno 100)5,0( x .
Anexo 6. (Continuação)