seladin-1 expression in rat adrenal gland: effect of
TRANSCRIPT

53
Seladin-1 expression in rat adrenal
gland: effect of adrenocorticotropichormone treatmentMarie-Claude Battista, Claude Roberge, Melissa Otis and Nicole Gallo-Payet
Service of Endocrinology, Department of Medicine, Faculte de medecine et des sciences de la sante, Universite de Sherbrooke, 3001, 12th Avenue North,Sherbrooke, Quebec, Canada J1H 5N4
(Requests for offprints should be addressed to N Gallo-Payet; Email: [email protected])
Abstract
Seladin-1 (KIAA0018) gene is the seventhmost highlyexpressed
gene in the adult adrenal gland, along with genes coding for
steroidogenic enzymes. The aim of the present study was to
investigate the localization of the Seladin-1 protein in control
and ACTH-treated rat adrenal glands and to verify whether
Seladin-1 is involved in secretion. Immunofluorescence studies
revealed that Seladin-1 was localized principally in the zona
fasciculata, cytoplasm, and nucleus. Expression of Seladin-1 was
increased byACTH treatment, in vivo and in culture conditions.
Subcellular fractionation of fasciculata cells showed that Seladin-
1 was mainly present in the nucleus, membrane, and
cytoskeleton fractions and, to a lesser extent, in the cytosol.
ACTH treatment decreased Seladin-1 expression in the cytosol,
with a concomitant increase in the nuclear fraction. In the
glomerulosa and fasciculata cells in culture, ACTH induced a
Journal of Endocrinology (2007) 192, 53–660022–0795/07/0192–053 q 2007 Society for Endocrinology Printed in Great
relocalization of Seladin-1 into specific nuclear regions. This
ACTH-induced relocalization was abrogated by the pre-
treatment of cells with 75 nM U18666A (an inhibitor of
Seladin-1). In addition, fasciculata cells exhibited an increase in
the basal level of steroid secretionwhen cultured in the presence
of U18666A (25 and 75 nM), although ACTH-induced
secretion was decreased. In summary, the present study
demonstrates that the protein expression of Seladin-1 is more
abundant in fasciculata cells than in glomerulosa cells and that
the ACTH treatment increases both expression and nuclear
localization of the protein. Results also suggest that depending
on its cellular localization, the D24-reductase activity of
Seladin-1 may play a major role in steroid secretion in the
adrenal gland.
Journal of Endocrinology (2007) 192, 53–66
Introduction
The adult adrenal cortex is composed of three concentric
layers: the zona glomerulosa, the zona fasciculata, and the zona
reticularis, all of which present different morphological and
functional properties. Zona glomerulosa is specialized in the
production of aldosterone while zona fasciculata/reticularis
synthesize cortisol in humans and bovine, and corticosterone
in rodents (for review, see Rainey 1999). The overall
production of aldosterone, however, is in the order of
picomolar, compared with the micromolar range for
cortisol/corticosterone (Gallo-Payet & Payet 1989, Rainey
1999, Sewer &Waterman 2003). The adrenal gland undergoes
constant dynamic structural changes and is generally well
acknowledged that cellular proliferation is preferentially
observed at the periphery of the gland (zona glomerulosa)
while cell death is increased in zona reticularis. The regulated
balance between proliferation and apoptosis is a prerequisite for
the integrative functionality of the gland (for review, see
Wolkersdorfer & Bornstein 1998, Vinson 2003, 2004).
Aside from angiotensin II (Ang II), adrenocorticotropin
hormone (ACTH) is the most potent stimulus of aldosterone
secretion by glomerulosa cells and of corticosterone by
fasciculata cells (Gallo-Payet & Payet 2003). ACTH acts not
only on the immediate, transcription-independent stimu-
lation of adrenal steroid synthesis and release, but also
increases the expression of a number of genes including
those involved in steroidogenesis (Sewer & Waterman 2003).
Despite several studies using both animal and human
glomerulosa and fasciculata cells, the precise molecular
mechanisms by which ACTH stimulates growth and
secretory activities are complex and poorly understood.
Over the past few years, studies on gene profiling and
regulation have provided key elements in our comprehension
as to how cell functions are regulated at the molecular level.
For instance, Seladin-1 (K1AA0018) gene is the seventh most
highly expressed known gene in the human adult adrenal
gland, along with genes coding for steroidogenic enzymes
(Hu et al. 2000). This gene is also expressed in the human fetal
adrenal gland with a fourfold increase in the expression when
compared with the adult (Rainey et al. 2001), thus being the
highest expressed known gene in human fetal adrenal gland
(Rainey et al. 2002). This protein has also been identified in
the brain and termed Seladin-1, for SELective Alzheimer
Disease INdicator 1, due to its propensity to protect neurons
from b-amyloid peptide-induced toxicity, thus promoting cell
DOI: 10.1677/joe.1.07062Britain Online version via http://www.endocrinology-journals.org

M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands54
survival (Greeve et al. 2000). Seladin-1 is a human homolog of
the Diminuto/Dwarf1 gene described in plants where it is
involved in growth and steroid synthesis (Takahashi et al.
1995, Klahre et al. 1998). In normal human adrenal cortex,
Seladin-1 mRNA expression is present throughout the gland
although more intense in the zona fasciculata (Sarkar et al.
2001). In adrenal glands from dexamethasone-treated rats
after ACTH treatment as well as in active human
adrenocortical adenomas, Seladin-1/hDiminuto mRNA is
overexpressed conversely to a reduced expression in adrenal
carcinomas (Sarkar et al. 2001, Luciani et al. 2004). In the
human adrenocortical cancer cell line H295R as well as in the
cultured human primary adrenocortical cells, Seladin-1
mRNA is regulated by the ACTH/cAMP pathway (Sarkar
et al. 2001, Luciani et al. 2004).
Seladin-1, a member of the flavin adenine dinucleotide-
dependent oxidoreductases family, is also named 3-b-hydroxysterol D-24-reductase (DHCR24; Greeve et al.
2000). This enzyme is known to catalyze the conversion of
desmosterol to cholesterol (Waterham et al. 2001), the
preferential pathway of cholesterol synthesis in the human
and the rat fetal brain (Fumagalli & Paoletti 1963). In humans,
mutations of the DHCR24 gene result in a rare and severe
recessive autosomic disorder called desmosterolosis. This
pathology is characterized by desmosterol accumulation in
plasma and tissues, by multiple congenital anomalies, and by
severe mental retardation (Waterham et al. 2001). Recently,
Seladin-1 was also described as a key regulator of Ras-induced
senescence and responses to oncogenic and oxidative stress
(Wu et al. 2004).
There is limited data available relative to Seladin-1 at the
protein level in the adrenal gland, the tissue with the highest
transcription level of Seladin-1 (Greeve et al. 2000). There-
fore, the first aim of the present study was to generate a
specific antibody to investigate the expression and cellular/
intracellular distribution of Seladin-1 protein in the adrenal
gland of both control and ACTH-treated rats. The second
aim was to compare this distribution in primary cultures of
adult rat glomerulosa and fasciculata cells, before and after
ACTH treatment. Using this model, we thereby explored
whether Seladin-1 is possibly involved in selected adreno-
cortical cell functions.
Materials and Methods
Chemicals
The chemicals used in the present study were obtained from
the following sources: Tissue-Tek from Miles (Elkhart, IN,
USA); Minimum Essential Medium (MEM Eagle’s medium)
and OPTI-MEMw from Life Technologies; collagenase from
NewEnglandNuclear (Boston,MA,USA);DNase fromSigma
Chemical Co.; ACTH 1–24 peptide (Cortrosyn) from
Organon (Toronto, Ont., Canada); ACTH 1–39 peptide
(Synacthen Depot 100 IU/ml) from Ciba Pharmaceuticals
Journal of Endocrinology (2007) 192, 53–66
(Caldwell, NJ, USA); angiotensin II from Bachem (Marina
Delphen, CA, USA). BrdU (5-bromo-2-deoxyuridine), anti-
BrdU, the secondary antibodies (mouse and rabbits) coupled
to Alexa-Fluor-488, Alexa-Fluor-594, the lectin wheat germ
agglutinin (WGA), and 4 0, 6 0-diamino-2-phenylindole
(DAPI) were from Molecular Probes (Eugene, OR, USA).
The anti-actin was from Chemicon International, Inc.
(Temecula, CA, USA); Vectamount from Vector Laboratories
(Burlingame, CA, USA); horseradish peroxidase-conjugated
anti-rabbit and anti-mouse antibodies and enhanced chemi-
luminescence (ECL) system from Amersham. The ProteoEx-
tract Subcellular Proteome Extraction Kit and the Seladin-1
inhibitor, U18666A (3b-(2-diethylaminoethoxy)androst-5-
en-17-one), were purchased from Calbiochem (San Diego,
CA, USA) and the RNAaqueous-4PCR Kit from Ambion
(Austin, TX, USA). [3H]aldosterone (76 Ci/mmol) and
[3H]corticosterone (70 Ci/mmol) were obtained from New
England Nuclear, Dupont (Boston, MA, USA) and the anti-
corticosterone from Medicorp (Montreal, Quebec, Canada).
Aldosterone antiserum was purchased from ICN Bio-
chemicals (Costa Mesa, CA, USA). All the other chemicals
were of A-grade purity.
Animals
Long–Evans female rats weighing 200–250 g were purchased
from Charles River, St Constant, Quebec, Canada. Upon
arrival, the rats were kept under controlled light and
temperature in our institution’s animal care facilities for 2
weeks prior to the start of the experimental procedure to
allow the animals to adapt to their environment, thereby
reducing stress. Rats were then randomly distributed in
control or in ACTH-treated groups. Animals were treated
once a day (1500 h) for 7 days either with saline (control) or
with Synacthen Depot subcutaneously (8 IU/250 g body
weight per day) and killed by decapitation on the morning of
the eighth day. All the protocols were approved by the Animal
Care and Ethics Committee of our faculty. A number of three
to five animals were used for each experiment and three
different experiments were performed.
Generation and characterization of Seladin-1 antiserum
The Seladin-1 antiserum was produced by Affinity Bior-
eagents (Golden CO, USA): rabbits were immunized with the
same peptide as the one used by Greeve et al. (2000),
consisting of amino acid residues 203–218 of Seladin-1 H3N-
TPSENSDLFYAVPWSC-COOH, maleimide-conjugated
to keyhole-limpet hemocyanin. This sequence is identical
in human, rat, and mouse. Specificity of the antibody was
confirmed by western blotting. Briefly, following electro-
phoresis and transfer, polyvinylidene difluoride membranes
were incubated as described below with Seladin-1 antiserum
(dilution 1:1000), with the pre-immune serum (dilution
1:1000) or with the Seladin-1 antiserum neutralized by the
peptide raised against at 175-fold molar excess. The
www.endocrinology-journals.org

Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 55
expression of Seladin-1 was compared with total rat adrenal
gland, brain, and cerebellum for equal protein loading
(20 mg). Seladin-1 antiserum detected two bands (77 and
47 kDa) in the adrenal gland (lane 2), and three bands (77, 60,
and 47 kDa) in the brain (lane 3) and cerebellum (lane 4;
Fig. 1A). Lane 5 shows a negative control (Leammli buffer).
Since the pre-immune serum (Fig. 1B) detected one band in
the adrenal gland (47 kDa) and two bands in the brain and
A1 2 3 4 5
1 2 3 4 5
1 2 3 4 5
125
38
85
7760
47
B
125
31
38
85
7760
47
C
125
38
857760
47
Figure 1 Representative western blot analysis of Seladin-1expression. (A) Seladin-1 antiserum detection in rat adrenal gland(lane 2), rat brain (lane 3), and rat cerebellum (lane 4) for equalprotein loading (20 mg). Lane 1 shows kaleidoscope markers andlane 5 serves as a negative control (Laemmli Buffer). In the adrenalgland, both the 77 and the 47 kDa bands are present while in thebrain and cerebellum; there is an additional band at 60 kDa. (B)Immunoblot in the presence of pre-immune serum. In the adrenalgland, only the 77 kDa band is specific since the 60 and 47 kDabands are also detected with the pre-immune serum. (C) Peptideneutralization assay confirming the specificity of the 77 kDa band,since the latter disappears in all three tissues. All the assays wereperformed at identical dilutions and exposure times.
www.endocrinology-journals.org
cerebellum (60 and 47 kDa), these were deemed non-specific
bands. Therefore, the 77 kDa band observed in Fig. 1A
appeared as specific for Seladin-1. With the peptide
neutralized-Seladin-1 antiserum, the 77 kDa band was no
longer detected in the three tissues (Fig. 1C), indicating
specificity of this band. Similar observation was also made in
the human fetal adrenal gland (data not shown).
Preparation of glomerulosa and fasciculata cell cultures
Glomerulosa and fasciculata cells were obtained from adrenal
glands of female Long–Evans rats and isolated according to the
method previously described in detail (Gallo-Payet & Payet
1989). Isolation and cell dissociation of glomerulosa and
fasciculata zones were performed in MEM (supplemented
with 2% fetal bovine serum (FBS), 100 U/ml penicillin, and
100 mg/ml streptomycin). After a 20-min incubation at 37 8C
with collagenase (2 mg/ml) and DNase (25 g/ml), cells were
disrupted by gentle aspiration with a sterile 10 ml pipette,
filtered, and centrifuged for 10 min at 100 g. For western blot
analyses (Fig. 6) from freshly isolated cells, 1!106 glomerulosa
and fasciculata cells were immediately processed for subcellular
protein extraction, and frozen until western blotting. For cell
cultures, cell pellets were resuspended in OPTI-MEMw
supplemented with 2% fetal bovine serum, 100 U/ml
penicillin, and 100 g/ml streptomycin. The glomerulosa and
fasciculata cells were plated in triplicate for immunofluores-
cence experiments (5!104 cells per dish). The cells were left
to adhere to the Petri dishes for 45 min before adding the
culture medium. The cells were cultured at 37 8C in a
humidified atmosphere composed of 95% air–5% CO2. The
culture medium was changed 6 h after initial addition of the
medium, and the cells were used after 3 days of culture. The
glomerulosa cells were stimulated with either Ang II (100 nM)
or ACTH (10 nM) for 3-day treatments, twice a day and
fasciculata cellswere stimulatedwithACTH(10 nM) for 3-day
treatments, also twice a day.
Fasciculata cells (5!104 cells)were also treated for 3dayswith
orwithoutACTH (10 nM, twice a day) alone or in the presence
of the Seladin-1 inhibitor, U18666A (25–75 nM, once a day).
U18666A, a cell-permeable, amphiphilic amino-steroid, is a
high-affinity binding inhibitor of the D24-reductase activity ofSeladin-1 (IC50: 0.15 mM), resulting in desmosterol accumu-
lation. At low concentrations (nanomolar range), U18666A
inhibits the D24-reductase activity of Seladin-1, with no effect
on cell viability (Bae & Paik 1997, Fliesler et al. 2000, Cenedella
et al. 2004). It inhibits the cholesterol metabolism activity of
Seladin-1. The cells were examined daily, and phase-contrast
images were taken using a microscope (Leica Corp., Deerfield,
IL, USA) equipped with a!32 objective.
Immunofluorescence studies in whole glands and cultured cells
Control and 7-day ACTH-treated female Long–Evans rats
were killed and adrenal glands dissected and retrieved. Glands
were either embedded in paraffin for hematoxylin–eosin
Journal of Endocrinology (2007) 192, 53–66

M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands56
counterstaining in order to identify morphological structures
or frozen and embedded in a cryoprotectant optimal cutting
temperature (OCT Tissue-Tek) for immunofluorescence
studies. Tissue sections (5 mm) or glomerulosa and fasciculata
cells grown in culture for immunofluorescence were fixed in
3.7% formaldehyde, permeabilized in PBS–0.2% Triton
X-100, and blocked in PBS–0.5% BSA, 0.2% Triton. Tissue
sections and cells were then incubated with the primary anti-
Seladin-1 antiserum or with the pre-immune serum (both
1:1000) followed by incubation with a secondary conjugated
anti-rabbit antibody coupled with Alexa-Fluor-594 nm
(1:500; red, frozen tissue) or Alexa-Fluor-488 nm (1:500;
green, cultured cells) for 1 h at room temperature. The cells
were also incubated with the lectin WGA, known as a Golgi
apparatus marker (5 mg/ml; red) coupled to Alexa-594 (1:500,
red; Kovacs et al. 2004). Tissues and cells were also stained with
DAPI (1:1000; blue) for visualization of nuclei. Slides were
mountedwithVectashieldmountingmedium and imageswere
acquired with a Hamamatsu, ORCA-ER digital camera and
examined under a fluorescence Nikon Eclipse 2000 inverted
microscope (Nikon, Mississauga, ON, Canada) equipped for
epi-illumination. Tissue images were acquired using a!10 or
!20 objectives, whereas cell images were acquired using a
!100 objective. Images were processed with Metamorph
(version 4.6r10) software (Universal Imaging Corporation,
Downingtown, PA, USA). In all the cases, no specific staining
was observed when primary antiserum was replaced by pre-
immune serum at the same dilution and exposure time. Images
were acquired using identical camera settings for contrast and
brightness. To better evaluate Seladin-1 nuclear expression
within the zona glomerulosa (under the capsule, four to five
cell layers), zona fasciculata, and zona reticularis, results were
expressed as the percentage of Seladin-1-stained nuclei found
within each cortical zone when compared with the total
number of nuclei present in that zone. Seladin-1 staining
within the nuclei was defined as the resulting pink color
obtained by the superposition of DAPI and anti-Seladin-1,
while total number included blue-colored nuclei.
Tissue protein extraction and subcellular fractionation
The glomerulosa and fasciculata zones were isolated from the
adrenal gland and separately snap-frozen in liquid nitrogen
and stored at K80 8C for tissue protein extraction, or cells
were immediately processed for subcellular protein fraction-
ation, as described below. The zona fasciculata fraction also
contains the zona reticularis, which is difficult to separate
from the medulla which we attempted to discard with two
small scapel cuts, but remnants are always possible. Since this
fraction contains predominantly fasciculata, it is referred to as
zona fasciculata throughout the text. Total tissue protein
extraction was performed after addition of boiling 2%
SDS/1% Triton/PBS solution to the zona glomerulosa and
zona fasciculata tissues (0.2 g tissue/ml). Tissues were
homogenized using a Teflon potter and heated at 100 8C
for 5 min. This procedure was repeated thrice. Subcellular
Journal of Endocrinology (2007) 192, 53–66
protein fractionation (cytosol, membranes, nucleus, and
cytoskeleton fractions) was done using the ProteoExtract
Subcellular Proteome Extraction Kit according to the
manufacturer’s instructions (Calbiochem). This kit contains
four extraction buffers prepared with ultra-pure chemicals
to ensure high reproducibility, protease inhibitor cocktail to
prevent protein degradation and benzonase nuclease to
remove contaminating nucleic acids. ProteoExtract Subcel-
lular Proteome Extraction Kit yields the total proteome
fractionated into four subproteomes of decreased complexity.
This kit has a high efficiency for subcellular fractionation
(Abdolzade-Bavil et al. 2004) and is widely used (Singh et al.
2004, Zhang & Insel 2004). All samples were assayed for
protein content before western blotting. Equivalent amounts
of proteins (15–30 mg for each fraction of subcellular
fractionation of isolated cells and 15 mg for total tissue proteinextraction) were compared in each experiment.
Western blotting
All the samples were separated on 12% SDS-polyacrylamide
gels and proteins transferred electrophoretically onto poly-
vinylidene difluoride membranes. After two washes with
Tris-buffered saline (pH 7.5)–Tween 20 (0.05%; TBS-T),
membranes were blocked with 5% milk/TBS-T. Membranes
were incubated with Seladin-1 antiserum (dilution 1:1000).
Detection was performed by reaction with horseradish
peroxidase-conjugated anti-rabbit secondary antibody and
visualized by enhanced chemiluminescence (ECL system)
according to the manufacturer’s instructions. Immunoreactive
bands were scanned by laser densitometry and expressed in
arbitrary units.
For whole zona glomerulosa and zona fasciculata protein
extracts, since the cell volume of glomerulosa cells is lower
than that of fasciculata cells, quantification of blots were
normalized to actin. For whole gland lysate immunoblotting,
after Seladin-1 incubation, membranes were stripped for 2 h
with glycine 0.2 M (pH 2.5) at 70 8C, then washed, blocked
in 1% gelatin, and further incubated with the anti-actin
antibody for 16 h (dilution 1:1000). In the experiments
involving subcellular fractionation, the results are expressed
as the percentage of Seladin-1 protein within each
subcellular fraction relative to total Seladin-1 content within
the cell.
RNA extraction and quantitative real-time PCR
Total RNA from glomerulosa and fasciculata zones, obtained
from control and ACTH-treated rats, was extracted and
treated with DNase I (to digest contaminating genomic
DNA) using RNAaqueous-4PCR Kit according to the
manufacturer’s recommendations. RNA content was
measured spectrophotometrically while RNA quality was
assessed by electrophoresis on denaturing 1% agarose gel.
Total RNA (0.5 mg) was denatured (65 8C, 5 min) in the
presence of 0.5 mg oligo(deoxythymidine)12–18 (Invitrogen)
www.endocrinology-journals.org

Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 57
and 0.5 mM dNTP (Amersham Pharmacia Biotech) and
reverse transcribed at 42 8C for 60 min in 20 ml of 1! first
strand buffer (250 mM Tris–HCl (pH 8.3), 375 mM KCl,
15 mM MgCl2) containing 10 mM dithiothreitol, 40 U
RNAse OUT inhibitor (Invitrogen), and 200 U SuperScript
II Reverse Transcriptase (Invitrogen). Inactivation of the
enzyme (70 8C, 15 min) was followed by glyceraldehyde-3-
phosphate dehydrogenase (GAPDH) PCR to asses the quality
of the template cDNA. Real-time PCR primers for rat
Seladin-1 and GAPDH (Invitrogen) were designed with
Beacon Designer 2.0 software (PREMIER, Biosoft Inter-
national, Palo Alto, CA, USA). Primer pairs are located on
both sides of an intron or on an exon–exon junction.
GAPDH: sense, TGGTGCCAAAAGGGTCATC; antisense,
CTTCCACGATGCCAAAGTTG; and Seladin-1: sense,
GGGTGTTTGTGTGCCTCTTCC; antisense, GCTCCT-
TCCACTCCCGTACC. Real-time PCR was performed
with an iCycler iQ Detection System using iQ SYBR Green
Supermix (Bio-Rad Laboratories) according to the manu-
facturer’s instructions and as described previously (Salzmann
et al. 2004). Annealing temperature was 60.4 8C. Real-time
PCR products were analyzed on 1! Tris/acetate/EDTA-
buffered 2.5% agarose gel and visualized by ethidium bromide
staining. To ensure the specificity of each real-time PCR,
prior melting curve analyses were performed for all detected
products with only a single peak recorded corresponding to a
unique melting temperature specific to each product. Sizes of
the amplicons obtained were 176 pb for GAPDH and 156 pb
for Seladin-1.
Incubation and measurement of aldosterone and corticosteronesecretion
Fasciculata and glomerulosa cells were plated in 24-well
plates (50!104 cells/well) and were treated daily for 3 days
with or without ACTH (10 nM, twice a day) alone or in the
presence of the Seladin-1 inhibitor, U18666A (25–75 nM,
once a day). After 3 days, the incubation medium was
removed by aspiration and stored at K20 8C for RIAs. For
acute stimulations, on the third day of cell culture, cells were
stimulated with or without ACTH (10 nM) alone or in the
presence of U18666A (25–75 nM), during a 2-h period.
Prior to each experiment, the medium from cultured cells
was aspirated, and the cells washed twice with cold Hanks’
buffered saline (HBS; 130 mM NaCl, 3.5 mM KCl, 1.8 mM
CaCl2, 0.5 mM MgCl2, 2.5 mM NaHCO3, and 5 mM
HEPES) supplemented with 1 g/l glucose and 0.5% BSA.
The cells were incubated in a 1 ml solution consisting of
0.9 ml HBS-glucose supplemented with 0.5% BSA–
0.1 mg/ml bacitracin and 0.1 ml stimuli. After a 2-h
incubation at 37 8C in a 95% air–5% CO2 environment,
the incubation medium was removed by aspiration and
stored at K20 8C until assayed for steroid content
determined by RIA, using specific antiserum and tritiated
steroid as tracer.
www.endocrinology-journals.org
Proliferation assays
The cell proliferation was measured using fluorescence BrdU
incorporation as previously described (Otis et al. 2005). The
glomerulosa and fasciculata cells were plated in 24-well plates
(30!104 cells/well). The cells were treated daily for 3 days
with or without ACTH (10 nM twice a day) alone or in the
presence of the inhibitor, U18666A (25–75 nM, once a day).
On the third day, cells were fixed with 3.7% (v/v)
formaldehyde in HBS for 10 min at room temperature and
permeabilized for 10 min with 0.2% Triton X-100 in HBS.
The cells were then incubated with anti-BrdU Alexa-Fluor-
594 (1:500). Fluorescence intensity was determined using a
Microplate Fluorescence Reader FL600 (Bio-Tek; excitation:
560/40 nm; emission: 645/40 nm).
Data analysis
The data are presented as meansGS.E.M. of the number of
experiments indicated in parentheses. Statistical analyses of
the data were performed using one-way ANOVA, followed
by a test of simple effects when appropriate. Homogeneity of
variance was assessed by Bartlett’s test and P values were
obtained by Tukey honestly significant differences. For simple
comparisons between two groups, Student’s t-test was
performed.
Results
Localization and protein expression of Seladin-1 in adrenal tissueand cells from control and ACTH-treated rats
Using the Seladin-1 antibody characterized in Fig. 1, the
distribution and cellular localization of Seladin-1 were first
studied by immunofluorescence on frozen adult rat adrenal
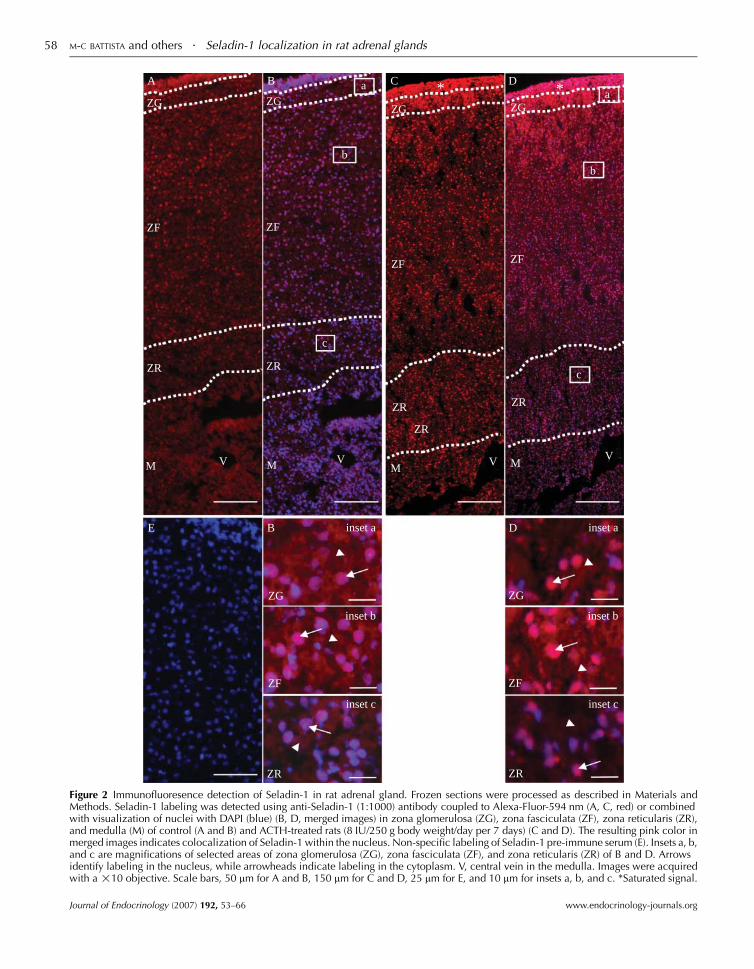
glands. As shown in Fig. 2A, the highest expression of
Seladin-1 was found at the periphery of the adrenal gland,
in the zona glomerulosa and zona fasciculata, while the
lowest expression was found in the zona reticularis and
medulla. To better differentiate intracellular expression,
colocalization with nuclear labeling (Fig. 2B) highlighted
that, in zona glomerulosa (Fig. 2B, inset a) and zona
fasciculata (Fig. 2B, inset b), labeling was observed in both
the nucleus (arrow) and the cytoplasm (arrowhead).
However, labeling in the nucleus was greater and stronger
in zona fasciculata than in zona glomerulosa. In zona
reticularis (Fig. 2B, inset c), nuclear labeling was present but
less in the other zones while cytoplasmic labeling was very
weak. In the medulla, Seladin-1 labeling was practically
absent. A 7-day treatment with ACTH increased overall
expression of Seladin-1 throughout the gland (Fig. 2C and
D). As shown in corresponding insets (Fig. 2D, insets a, b
and c), labeling was increased in the nuclei in the three
zones. Labeling was completely abrogated when Seladin-1
Journal of Endocrinology (2007) 192, 53–66
E
B
ZG
ZF
ZR
M V
aA
ZG
ZF
ZR
M V
D
ZG
ZF
M
ZR
*
V
ZG
ZF
VM
ZR
C
ZR
*
b
c
b
c
a
B
ZG
ZF
ZR
D inset a
inset c
inset b
inset a
inset c
inset b
ZG
ZF
ZR
Figure 2 Immunofluoresence detection of Seladin-1 in rat adrenal gland. Frozen sections were processed as described in Materials andMethods. Seladin-1 labeling was detected using anti-Seladin-1 (1:1000) antibody coupled to Alexa-Fluor-594 nm (A, C, red) or combinedwith visualization of nuclei with DAPI (blue) (B, D, merged images) in zona glomerulosa (ZG), zona fasciculata (ZF), zona reticularis (ZR),and medulla (M) of control (A and B) and ACTH-treated rats (8 IU/250 g body weight/day per 7 days) (C and D). The resulting pink color inmerged images indicates colocalization of Seladin-1 within the nucleus. Non-specific labeling of Seladin-1 pre-immune serum (E). Insets a, b,and c are magnifications of selected areas of zona glomerulosa (ZG), zona fasciculata (ZF), and zona reticularis (ZR) of B and D. Arrowsidentify labeling in the nucleus, while arrowheads indicate labeling in the cytoplasm. V, central vein in the medulla. Images were acquiredwith a !10 objective. Scale bars, 50 mm for A and B, 150 mm for C and D, 25 mm for E, and 10 mm for insets a, b, and c. *Saturated signal.
M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands58
Journal of Endocrinology (2007) 192, 53–66 www.endocrinology-journals.org

A
B
ZG
ZF
ZG
ZF
Figure 3 Hematoxylin–eosin staining of control (A) and ACTH-treated (8 IU/250 g body weight/day per 7 days) (B) rat adrenalglands from the capsule to the mid-fasciculata zone. Lines delineatezona glomerulosa (ZG) from zona fasciculata (ZF).
Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 59
antiserum was replaced by its pre-immune serum at
matching dilution and similar exposure time (Fig. 2E).
For quantification of Seladin-1 expression in the various
adrenal zones, histological comparisons with hematoxylin–
eosin staining was used to clearly delineate zones in frozen
sections, which were then identified by dashed lines in Fig. 2.
As shown in Fig. 3A and B, in adrenal glands from control
animals, the zona glomerulosa and zona fasciculata were
0
20
40
60
80
100
Sel
adin
-1 in
nuc
leus
(% o
f tot
al e
xpre
ssio
n)
ZG ZF ZR
ControlACTH*
*
Figure 4 Quantification of Seladin-1 immunofluorescence labelingin whole frozen adrenal gland sections. Results represent thepercentage of nuclei labeled with Seladin-1 protein relative to thetotal number of nuclei within each adrenocortical section of zonaglomerulosa (ZG), zona fasciculata (ZF), and zona reticularis (ZR)from control (nZ3) and ACTH-treated rats (8 IU/250 g bodyweight/day per 7 days; nZ3). Results are expressed as meansGS.E.M. Statistical significance when compared with respectivecontrol conditions: *P!0.05.
www.endocrinology-journals.org
clearly distinct, while after ACTH treatment, zonation was
not as clearly delineated. A progressive change from small to
large cells was observed, which rendered the separation from
zona glomerulosa to zona fasciculata difficult to identify.
Results from Fig. 4 confirm that the basal level of expression
in the nucleus was similar between zona glomerulosa and
fasciculata and lower in zona reticularis. ACTH treatment did
not modify the number of Seladin-1-labeled nuclei in zona
glomerulosa, while increasing the number of labeled nuclei
from 67.5G1.8 to 84.6G4.4% (P!0.05) in zona fasciculata
and from 32.1G7.6 to 58.0G5.2% (P!0.05) in the zona
reticularis. Western blotting further confirmed the morpho-
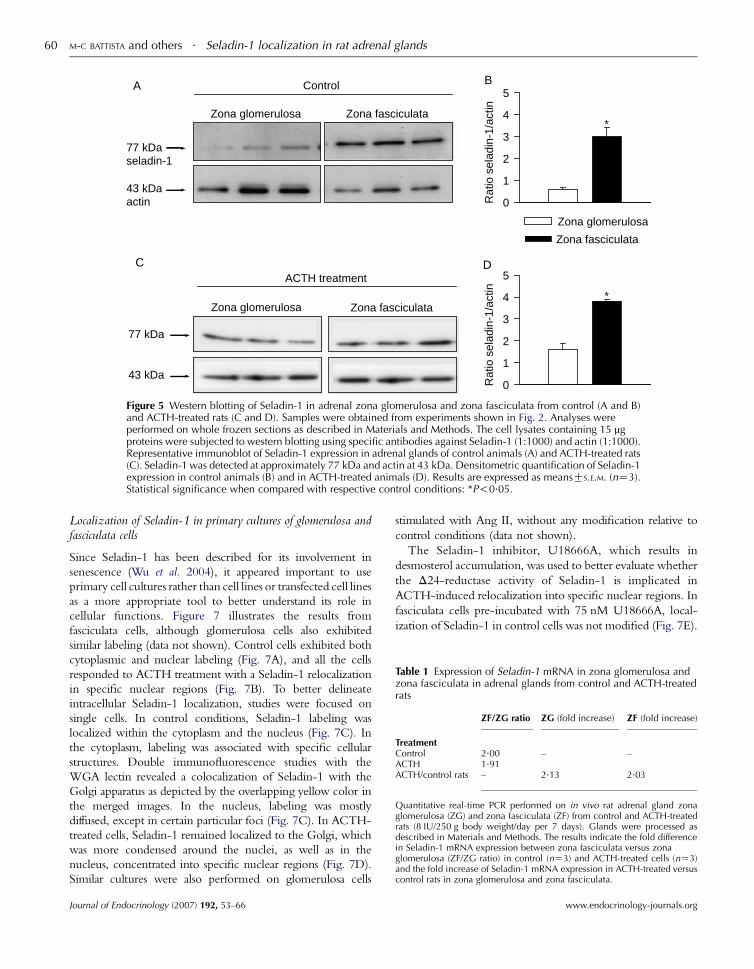
logical observations that the overall expression of Seladin-1
was higher in the zona fasciculata than in the glomerulosa.
Seladin-1 protein appeared as a single band of 77 kDa, with
expression levels fivefold higher in the zona fasciculata than in
the zona glomerulosa (Fig. 5A and B; P!0.05). ACTH
treatment increased overall expression in both zona glomer-
ulosa and zona fasciculata (Fig. 5C and D).
The above results on protein expression were also
corroborated with real-time PCR analysis of Seladin-1
mRNA expression, which indicated that the expression in
zona fasciculata was indeed twofold greater than in zona
glomerulosa and that ACTH increased Seladin-1 expression
by twofold, in both zona glomerulosa and zona fasciculata
(Table 1). Together, these results indicate that in vivo, Seladin-
1 is largely expressed in the zona fasciculata and to a lesser
extent in the zona glomerulosa. Following ACTH treatment,
a preferential localization of Seladin-1 was observed in the
nucleus of fasciculata cells.
Since specific functions are associated with specific
intracellular structures, subcellular fractionation followed by
western blotting was performed on isolated zona glomerulosa
and zona fasciculata, obtained fromcontrol andACTH-treated
rats. As shown in Fig. 6A, in glomerulosa cells, Seladin-1 was
mainly present in themembrane and nuclear fractions and, to a
lesser extent in the cytosol and cytoskeleton. ACTH treatment
decreased Seladin-1 expression in the cytosol (from 19.4G1.8to 1.7G0.5%; P!0.001) and membrane fraction (from
30.5G1.9 to 18.6G1.8%; P!0.01), with a concomitant
increase in the cytoskeletal fraction (from 18.6G1.2 to 38.8G4.4%; P!0.001). Although Seladin-1 was also increased in thenuclear fraction, it did not reach statistical significance. In
fasciculata cells (Fig. 6B), Seladin-1 was mainly present in the
nucleus, and to a lesser extent in the cytoskeleton andmembrane
fractions. ACTH treatment decreased cytosolic content and
increasednuclear localizationof Seladin-1 (37.5G0.3 to 48.0G5.0%; P!0.05). Therefore, in control conditions, the
cytoplasmic distribution was higher in zona glomerulosa than
in zona fasciculata, while the association with cytoskeleton was
higher in zona fasciculata when compared with zona
glomerulosa. On the other hand, ACTH-treatment favored
cytoskeletal localization within glomerulosa cells and as
demonstrated previously in vivo, nuclear localization within
fasciculata cells. Such observations could be associated with
differential effects for each respective zone.
Journal of Endocrinology (2007) 192, 53–66
Control
43 kDaactin
A
77 kDaseladin-1
Zona glomerulosa Zona fasciculata
0
1
2
3
4
5
Rat
io s
elad
in-1
/act
in
B
*
Zona glomerulosa
Zona fasciculata
C
Zona glomerulosa Zona fasciculata
ACTH treatment
43 kDa
77 kDa
0
1
2
3
4
5
Rat
io s
elad
in-1
/act
in
*
D
Figure 5 Western blotting of Seladin-1 in adrenal zona glomerulosa and zona fasciculata from control (A and B)and ACTH-treated rats (C and D). Samples were obtained from experiments shown in Fig. 2. Analyses wereperformed on whole frozen sections as described in Materials and Methods. The cell lysates containing 15 mgproteins were subjected to western blotting using specific antibodies against Seladin-1 (1:1000) and actin (1:1000).Representative immunoblot of Seladin-1 expression in adrenal glands of control animals (A) and ACTH-treated rats(C). Seladin-1 was detected at approximately 77 kDa and actin at 43 kDa. Densitometric quantification of Seladin-1expression in control animals (B) and in ACTH-treated animals (D). Results are expressed as meansGS.E.M. (nZ3).Statistical significance when compared with respective control conditions: *P!0.05.
Table 1 Expression of Seladin-1 mRNA in zona glomerulosa andzona fasciculata in adrenal glands from control and ACTH-treatedrats
ZF/ZG ratio ZG (fold increase) ZF (fold increase)
TreatmentControl 2.00 – –ACTH 1.91ACTH/control rats – 2.13 2.03
Quantitative real-time PCR performed on in vivo rat adrenal gland zonaglomerulosa (ZG) and zona fasciculata (ZF) from control and ACTH-treatedrats (8 IU/250 g body weight/day per 7 days). Glands were processed asdescribed in Materials and Methods. The results indicate the fold differencein Seladin-1 mRNA expression between zona fasciculata versus zonaglomerulosa (ZF/ZG ratio) in control (nZ3) and ACTH-treated cells (nZ3)and the fold increase of Seladin-1 mRNA expression in ACTH-treated versuscontrol rats in zona glomerulosa and zona fasciculata.
M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands60
Localization of Seladin-1 in primary cultures of glomerulosa andfasciculata cells
Since Seladin-1 has been described for its involvement in
senescence (Wu et al. 2004), it appeared important to use
primary cell cultures rather than cell lines or transfected cell lines
as a more appropriate tool to better understand its role in
cellular functions. Figure 7 illustrates the results from
fasciculata cells, although glomerulosa cells also exhibited
similar labeling (data not shown). Control cells exhibited both
cytoplasmic and nuclear labeling (Fig. 7A), and all the cells
responded to ACTH treatment with a Seladin-1 relocalization
in specific nuclear regions (Fig. 7B). To better delineate
intracellular Seladin-1 localization, studies were focused on
single cells. In control conditions, Seladin-1 labeling was
localized within the cytoplasm and the nucleus (Fig. 7C). In
the cytoplasm, labeling was associated with specific cellular
structures. Double immunofluorescence studies with the
WGA lectin revealed a colocalization of Seladin-1 with the
Golgi apparatus as depicted by the overlapping yellow color in
the merged images. In the nucleus, labeling was mostly
diffused, except in certain particular foci (Fig. 7C). In ACTH-
treated cells, Seladin-1 remained localized to the Golgi, which
was more condensed around the nuclei, as well as in the
nucleus, concentrated into specific nuclear regions (Fig. 7D).
Similar cultures were also performed on glomerulosa cells
Journal of Endocrinology (2007) 192, 53–66
stimulated with Ang II, without any modification relative to
control conditions (data not shown).
The Seladin-1 inhibitor, U18666A, which results in
desmosterol accumulation, was used to better evaluate whether
the D24-reductase activity of Seladin-1 is implicated in
ACTH-induced relocalization into specific nuclear regions. In
fasciculata cells pre-incubated with 75 nM U18666A, local-
ization of Seladin-1 in control cells was not modified (Fig. 7E).
www.endocrinology-journals.org

†
*
†
% o
f Sel
adin
-1 p
rote
inin
sub
cellu
lar
com
part
men
ts
0
10
20
30
40
50
60
cytosol
cytoskeletonmembrane
nucleus
cytosol
cytoskeletonmembrane
nucleus
A%
of S
elad
in-1
pro
tein
in s
ubce
llula
r co
mpa
rtm
ents
0
10
20
30
40
50
60
*
*
B
Control
ACTH
Figure 6 Western blot analysis of Seladin-1 performed insubcellular fractions of adrenocortical cells. Subcellular fraction-ation was performed from isolated zona glomerulosa cells (A) andzona fasciculata cells (B). Results are expressed as the percentage ofSeladin-1 protein expression within each subcellular fractionrelative to total intracellular expression of Seladin-1 in control andACTH-treated rats (8 IU/250 g body weight/day per 7 days). Resultsare expressed as meansGS.E.M. of three different experiments.Statistical significance when compared with respective controlconditions: *P!0.05; †P!0.01.
Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 61
However, in ACTH-stimulated cells, Seladin-1 was retained in
the Golgi apparatus (Fig. 7F), indicating that pre-incubation
with U18666A abrogated the ACTH-induced relocalization of
Seladin-1 into specific nuclear regions.
Effect of Seladin-1 inhibitor (U18666A) on steroid secretion andproliferation
In order to investigate the possible involvement of Seladin-1
in steroid secretion, glomerulosa and fasciculata cells were
www.endocrinology-journals.org
treated with or without ACTH alone or with U18666A (25
and 75 nM), either for 2 h or 3 days. Results from Fig. 8
indicate that both 2-h and 3-day treatments with U18666A
increased basal steroid output in glomerulosa and fasciculata
cells, but decreased ACTH-induced aldosterone and corti-
costerone secretion. The inhibitory action of U18666A on
ACTH-induced steroid secretion was more pronounced
however after 3 days than after acute 2-h treatment, and
was more pronounced for aldosterone secretion in glomer-
ulosa cells than for corticosterone secretion in fasciculata cells
(63 versus 43% reduction respectively). These results suggest
that Seladin-1 is differentially implicated in basal and in
ACTH-induced steroid secretion. On the other hand, 3-day
treatment with U18666A did not modify active cell
proliferation observed in cultured cells (data not shown).
Discussion
This study characterizes for the first time the expression,
distribution, intracellular localization, and dynamics of
Seladin-1 protein in adrenal glands and in primary cultures
of adrenocortical cells, in control and in ACTH-treated rats.
The most notable findings of this study are that Seladin-1 (1)
is localized both in the cytoplasm and in the nucleus; (2)
subcellular localization is regulated by ACTH; and (3) is
differentially involved in basal and in ACTH-stimulated
steroid secretion.
In virtually all studies, Seladin-1 has been studied at the
mRNA level in transfected or retroviral-transducted cells,
under non-physiological stimulations (Greeve et al. 2000, Wu
et al. 2004) or in vivo in normal and pathological conditions in
human (Sarkar et al. 2001, Fuller et al. 2005, Luciani et al.
2005). Herein, the antibody generated to conduct the present
investigations revealed a single band with a molecular weight
(MW) of 77 kDa band for Seladin-1 in the rat adrenal gland,
brain, and cerebellum. This band was not detected when
Seladin-1 antiserum was replaced by the pre-immune serum
at same dilution and exposure time or when the Seladin-1
antiserum was neutralized by the peptide raised against
Seladin-1, indicating specificity of the antibody. A previous
study had reported a Seladin-1 MWof 60 kDa in normal and
Alzheimer’s disease brain tissues from 62- to 92-year-old
patients (Greeve et al. 2000). The differences between these
values may be related to differences in species (human versus
rat) or age. However, we cannot rule out the presence of the
60 kDa Seladin-1 band in the rat brain and cerebellum since it
could be masked by the strong non-specific 60 kDa band
observed in those rat tissues with the pre-immune serum and
the Seladin-1 antiserum. This observation is strengthened by
the fact that the intensity of 60 kDa band was reduced by the
antiserum neutralization assay. Since there is no stop codon
before the putative initiation codon for the 60 kDa Seladin-1
transcripts (human, rat, and mouse), the putative 5 0-
unstranslated Seladin-1 mRNA extremity may be a coding
sequence for the actual amino-terminal end of Seladin-1.
Journal of Endocrinology (2007) 192, 53–66

Golgi apparatusSeladin-1 Merge
Con
trol
D
C
AC
TH
U18
666A
E
U18
666A
+A
CT
H
Control ACTH
A
Low
er m
agni
ficat
ion
B
F
Figure 7 Immunofluorescence detection of Seladin-1 in 3-day zona fasciculata cultured cells. Impact of ACTH and U18666A(Seladin-1 inhibitor). The cells were cultured for 3-days in the absence (control, A, C) or in the presence of ACTH (10 nM) (B and D) andin the presence of 75 nM U18666A alone (E) or in combination with ACTH (F). After formaldehyde fixation and permeabilizationwith 0.1% Triton X-100, the cells were processed for immunofluorescence detection of Seladin-1 (1:1000) coupled to Alexa-488 (1:500,green), Golgi apparatus by the use of wheat germ agglutinin (WGA) lectin coupled to Alexa-594 (1:500, red) and nucleus (DAPI, blue).The resulting yellow color in merged images indicates colocalization of Seladin-1 with the WGA (Golgi apparatus) while the resultinglight blue color indicates colocalization of Seladin-1 with DAPI (nucleus). All images were acquired with a !100 objective. Scale bars,5.6 mm (magnification: !3500 for C, D and !4500 for E, F).
M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands62
Journal of Endocrinology (2007) 192, 53–66 www.endocrinology-journals.org

†*
*† ††
Ald
oste
rone
(Fol
d in
crea
se o
ver
cont
rol)
0
5
10
15
20
‡
§
none 25 nM 75 nM
U18666A
Cor
ticos
tero
ne(F
old
incr
ease
ove
r co
ntro
l)
0
5
10
15
20
none 25 nM 75 nM
U18666A
‡
2 h stimulation
A
2 h stimulationC
none 25 nm 75 nm
U18666A
Ald
oste
rone
(Fol
d in
crea
se o
ver
cont
rol)
0
5
10
15
20
§§
3-days stimulationB
Cor
ticos
tero
ne(F
old
incr
ease
ove
r co
ntro
l)
0
5
10
15
203-days stimulation
none 25 nM 75 nM
U18666A
‡
D
ControlACTH
Figure 8 Impact of Seladin-1 inhibitor, U18666A, on steroid secretion by zona glomerulosa cells(aldosterone) (A and B) and zona fasciculata cells (corticosterone) (C and D) cultured for 3 days. Steroids weremeasured by RIA as described in Materials and Methods. Secretion was measured after stimulation for 2 h(A and C) or after 3-day treatment (B and D) with ACTH (10 nM) or in the absence or in the presence of 25 and75 nM U18666A. Results are expressed as meansGS.E.M. (nZ4). Statistical significance when compared withrespective control conditions: *P!0.05; †P!0.01, comparison between control U18666A groups of cells;‡P!0.05; §P!0.01, comparison between ACTH-treated cells.
Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 63
There are currently a number of Seladin-1 mRNA sequences
and predictive sequences that are now associated with a
Seladin-1 protein larger than 60 kDa. For example, two
recent predictive rat Seladin-1 sequences were shown to have
a calculated MWof 86 kDa, hence clearly over 60 kDa.
At both the mRNA and the protein levels, Seladin-1 was
observed to be more abundant in the zona fasciculata of
control and ACTH-treated animals. This result is in
agreement with previous studies conducted on human
adrenal gland and human fasciculata cells in culture, at the
mRNA level (Sarkar et al. 2001, Luciani et al. 2004).
Moreover, the present results indicate that Seladin-1 is
localized both in the cytoplasm and in the nucleus, with a
differential gradient of expression from the outer to the inner
portion of the gland. Our hypothesis is that Seladin-1
localization is ubiquitous in the adrenal gland and that its
www.endocrinology-journals.org
presence is more important for adrenocortical than for adrenal
medullar functions.
In control glomerulosa cells, Seladin-1 was mostly
expressed in the membrane and nuclear fractions. Upon
ACTH-treatment, the protein was greatly reduced in the
cytosolic and membrane compartments, while conversely
increasing in the cytoskeletal fraction. In fasciculata cells, the
protein was localized mainly in nuclear, cytoskeletal, and
membrane fractions. ACTH treatment reduced expression in
the cytoplasm, with a concomitant relocalization into the
nuclear fraction. These results suggest that the nuclear
Seladin-1 function is more important in fasciculata than in
glomerulosa cells. Through the use ofWGA lectin as a marker
of the Golgi apparatus, immunofluorescence studies demon-
strate that, in the cytoplasm, the protein was mainly associated
with the Golgi apparatus. Such localization has also been
Journal of Endocrinology (2007) 192, 53–66

M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands64
reported by Greeve et al. (2000) where, in a transfected
human neuroglioma H4 cell line, Seladin-1–enhanced green
fluorescent protein was localized in the endoplasmic
reticulum and to the Golgi complex. Localization of
Seladin-1 in Golgi may be correlated with a role in
steroidogenesis since it is well known that in adrenocortical
cells, the Golgi apparatus plays important functional and
regulatory roles in corticosterone synthesis and that ACTH
stimulation often leads to Golgi hypertrophy (Magalhaes et al.
1991, Cheng & Kowal 1997). These data may explain why a
modification in Golgi structure is observed in cells stimulated
with ACTH in the present model. This may suggest that the
Seladin-1 localization to the Golgi apparatus is important for
steroid secretion.
Furthermore, when glomerulosa and fasciculata cells in
culture are stimulated with ACTH, there is a relocalization of
Seladin-1 in specific nuclear regions. Wu et al. (2004) have
recently described a massive translocation of Seladin-1 from
the cytoplasm to the nucleus in Seladin-1-transfected
fibroblasts, in cells exposed to 0.5 mM H2O2. The authors
attributed a major role of Seladin-1 in the cellular response to
oxidative stress, due to its ability to bind both the tumor
suppressor p53 and the E3-ubiquitin-ligase Mdm2 and to
displace Mdm2 from p53, thus protecting p53 from Mdm2-
induced degradation and enabling p53 accumulation (Wu
et al. 2004). It is well established that intense steroidogenesis in
zona fasciculata leads to oxidative stress due to the occurrence
of lipid peroxidation and the production of reactive oxygen
species and toxic derivatives, such as isocaproaldehyde
(Hornsby & Crivello 1983, Lefrancois-Martinez et al.
1999). This may explain the important quantity of
endogenous anti-oxidant compounds (vitamin E,
b-carotene, and vitamin C; Hornsby & Crivello 1983) and
the presence of enzymes implicated in the endogenous
detoxification of steroidogenesis by-products (Martinez et al.
2001) in the adrenal gland. Since Seladin-1 is an oxido-
reductase enzyme (Mushegian & Koonin 1995), and in
agreement with the results of Wu et al. (2004), the present
results of a prominent Seladin-1 accumulation in specific
nuclear regions may thus reflect an ACTH-induced
regulation of Seladin-1 association with nuclear partners,
such as p53 and Mdm2. The ACTH-induced Seladin-1
relocalization into specific nuclear regions is inhibited by
U18666A suggesting an implication of the D24-reductaseactivity of Seladin-1. This hypothesis is supported by the
observation that ACTH has a protective effect against
oxidative stress (Pomeraniec et al. 2004). Together, the
observation that ACTH induces relocalization in specific
intranuclear compartments raises the hypothesis that Seladin-
1 may play a major role in protecting adrenocortical cells
against negative side effects due to intense steroidogenesis,
either as a new major participant in the p53-Mdm2 interplay
and/or through its own oxidoreductase activity. Such nuclear
relocalization was not observed with Ang II treatment in
glomerulosa cells, further supporting the notion that nuclear
relocalization is associated with intense steroidogenesis.
Journal of Endocrinology (2007) 192, 53–66
Indeed, Ang II stimulates aldosterone secretion by 2- to
5-fold, compared with the 30- to 60-fold increase observed
with ACTH (Gallo-Payet & Payet 1989, Campbell et al.
2003).
The role of Seladin-1 in steroid secretion was assessed
herein through the use of U18666A. At low concentrations
(nM–mM), U18666A is a high-affinity binding inhibitor of
the D24-reductase activity of Seladin-1, resulting in
desmosterol accumulation (Bae & Paik 1997, Fliesler et al.
2000, Cenedella et al. 2004). In the present study, U18666A
was used at concentrations ranging from 25 to 75 nM
because, at this concentration, it does not induce adrenal cell
toxicity and has little impact on cholesterol trafficking (Feng
et al. 2003). Our results indicate that U18666A increases basal
steroid secretion, but decreases ACTH-responsiveness for
both aldosterone and corticosterone. The observation that
U18666A effects on basal- and on ACTH-stimulated cells are
more pronounced on aldosterone production than on
corticosterone production may stem from a lower expression
level of Seladin-1 in zona glomerulosa cells, resulting in less
residual functional Seladin-1 following U18666A treatment.
Stimulation of basal secretion by this inhibitor may be due
to desmosterol accumulation produced by U18666A-induced
Seladin-1 inhibition. Desmosterol is a better substrate than
cholesterol for steroidogenesis. It is more efficiently esterified
than cholesterol, an essential step in sterol transport (Nordby
& Norum 1975), and is fourfold more efficient as a substrate
for the side chain cleavage reaction that produces precursors
for steroidogenesis (Arthur et al. 1976, Mason et al. 1978).
However, desmosterol has also been recently identified as a
major negative regulator of cholesterol metabolism, being a
strong inhibitor of sterol-regulatory element-binding protein
2 processing and a strong repressor of hydroxy-methyl-
glutaryl-coenzymeA (HMG-CoA) reductase and low-density
lipoprotein receptor expression (Yang et al. 2006). Therefore,
by controlling desmosterol levels, the D24-reductase activity
of Seladin-1 may play a major and dual role in regulating
cholesterol uptake and biosynthesis, thereby modulating
steroidogenesis. From these observations, low desmosterol
concentrations may have positive effects on steroidogenesis, as
a better substrate than cholesterol, which may explain the
observed U18666A-induced increase in basal secretion.
However, in ACTH-treated cells, the U18666A-induced
desmosterol accumulation may impair the larger ACTH-
induced increase in steroidogenic machinery. Seladin-1 may
therefore act as a negative regulator of basal steroidogenesis
when localized in the Golgi. Conversely, ACTH actions on
Seladin-1 expression and subcellular localization may have
two complementary effects: the first being the removal of the
inhibition of secretion in the cytoplasm, whereas the second
action would be associated with protection against ACTH-
induced oxidative stress.
Seladin-1 expression has also been reported to be altered in
adrenocortical adenomas and carcinomas, being overex-
pressed or repressed depending on tumor type (Sarkar et al.
2001, Luciani et al. 2004). However, Seladin-1 is also
www.endocrinology-journals.org

Seladin-1 localization in rat adrenal glands . M-C BATTISTA and others 65
abundantly expressed in mesenchymal stem cells (Benvenuti et
al. 2006). On the other hand, U18666A induces apoptosis in
cortical neurons in primary cultures (Koh & Cheung 2006).
These observations suggest that Seladin-1 is involved in the
balance between proliferation and apoptosis. However, such a
role was not observed in glomerulosa cell cultures, since
U18666A treatment had no effect on the high level of
proliferation.
In summary, the results obtained in the present study are
the first to assess Seladin-1 expression and distribution in the
adrenal gland in vivo and in the rat primary cultures of
adrenocortical cells. Using a multi-faceted approach, the
study demonstrates that the protein expression of Seladin-1 is
more abundant in fasciculata cells than in glomerulosa cells
and that ACTH treatment increases both expression and
nuclear localization of Seladin-1. Taken together, the results
obtained herein, combined with previous data, suggest that
depending on its cellular localization, Seladin-1 plays a major
role in steroid secretion in the adrenal gland. It is proposed
that the D24-reductase activity of Seladin-1 may be engaged
in maintaining low levels of steroidogenesis. ACTH treatment
induces a relocalization of Seladin-1 into the nucleus, thus
facilitating ACTH-induced stimulation of steroidogenesis in
the cytoplasm. Concomitantly, Seladin-1, in the nucleus,
could be involved (through possible association with p53 and
Mdm2 as partners), in protection against ACTH-induced
oxidative stress, an undesirable consequence of intense
steroidogenesis.
Acknowledgements
The authors thank Lucie Chouinard for her invaluable
experimental assistance and stimulating discussions. This
work was supported by grants from the Fondation pour la
Recherche sur les Maladies Infantiles and the Canadian
Research Chair Program to. N G-P is also a recipient of a
Canada Research Chair in Endocrinology of the Adrenal
Gland. The authors declare that there is no conflict of interest
that would prejudice the impartiality of this scientific work.
References
Abdolzade-Bavil A, Hayes S, Goretzki L, Kroger M, Anders J & Hendriks R
2004 Convenient and versatile subcellular extraction procedure, that
facilitates classical protein expression profiling and functional protein
analysis. Proteomics 4 1397–1405.
Arthur JR, Blair HA, Boyd GS, Mason JI & Suckling KE 1976 Oxidation of
cholesterol and cholesterol analogues by mitochondrial preparations of
steroid-hormone-producing tissue. Biochemical Journal 158 47–51.
Bae SH & Paik YK 1997 Cholesterol biosynthesis from lanosterol:
development of a novel assay method and characterization of rat liver
microsomal lanosterol delta 24-reductase. Biochemical Journal 326 609–616.
Benvenuti S, Saccardi R, Luciani P, Urbani S, Deledda C, Cellai I, Francini F,
Squecco R, Rosati F, Danza G et al. 2006 Neuronal differentiation of
human mesenchymal stem cells: changes in the expression of the
Alzheimer’s disease-related gene seladin-1. Experimental Cell Research 312
2592–2604.
www.endocrinology-journals.org
Campbell S, Otis M, Cote M, Gallo-Payet N & Payet M 2003 Connection
between integrins and cell activation in rat adrenal glomerulosa cells. A role
for RGD peptide in the activation of the p42/p44MAPK pathway and of
intracellular calcium. Endocrinology 144 486–495.
CenedellaRJ, JacobR,BorchmanD,TangD,Neely AR, Samadi A,MasonRP
& Sexton P 2004 Direct perturbation of lens membrane structure may
contribute to cataracts caused by U18666A, an oxidosqualene cyclase
inhibitor. Journal of Lipid Research 45 1232–1241.
Cheng B & Kowal J 1997 Role of the Golgi complex in adrenocortical
steroidogenesis. Microscopy Research and Technique 36 503–509.
Feng B, Yao PM, Li Y, Devlin CM, Zhang D, Harding HP, Sweeney M,
Rong JX, Kuriakose G, Fisher EA et al. 2003 The endoplasmic reticulum is
the site of cholesterol-induced cytotoxicity in macrophages. Nature Cell
Biology 5 781–792.
Fliesler SJ, Richards MJ, Miller C, Peachey NS & Cenedella RJ 2000 Retinal
structure and function in an animal model that replicates the biochemical
hallmarks of desmosterolosis. Neurochemical Research 25 685–694.
Fuller P, Alexiadis M, Jobling T & McNeilage J 2005 Seladin-1/DHCR24
expression in normal ovary, ovarian epithelial and granulosa tumours.
Clinical Endocrinology 63 111–115.
Fumagalli R & Paoletti R 1963 The identification and significance of
desmosterol in the developing human and animal brain. Life Sciences 5
291–295.
Gallo-Payet N & Payet MD 1989 Excitation–secretion coupling: involvement
of potassium channels in ACTH-stimulated rat adrenocortical cells.
Journal of Endocrinology 120 409–421.
Gallo-Payet N & Payet MD 2003 Mechanism of action of ACTH: beyond
cAMP. Microscopy Research and Technique 61 275–287.
Greeve I, Hermans-Borgmeyer I, Brellinger C, Kasper D, Gomez-Isla T,
Behl C, Levkau B & Nitsch RM 2000 The human DIMINUTO/
DWARF1 homolog seladin-1 confers resistance to Alzheimer’s disease-
associated neurodegeneration and oxidative stress. Journal of Neuroscience 20
7345–7352.
Hornsby P & Crivello J 1983 The role of lipid peroxidation and biological
antioxidants in the function of the adrenal cortex. Part 2. Molecular and
Cellular Endocrinology 30 123–147.
HuRM,HanZG,SongHD,PengYD,HuangQH,RenSX,GuYJ,HuangCH,
Li YB, Jiang CL et al. 2000 Gene expression profiling in the human
hypothalamus–pituitary–adrenal axis and full-length cDNAcloning.PNAS97
9543–9548.
Klahre U, Noguchi T, Fujioka S, Takatsuto S, Yokota T, Nomura T, Yoshida S
& Chua NH 1998 The Arabidopsis DIMINUTO/DWARF1 gene encodes
a protein involved in steroid synthesis. Plant Cell 10 1677–1690.
Koh CH & Cheung NS 2006 Cellular mechanism of U18666A-mediated
apoptosis in cultured murine cortical neurons: bridging Niemann-Pick
disease type C and Alzheimer’s disease. Cellular Signalling 18 1844–1853.
Kovacs WJ, Schrader M, Walter I & Stangl H 2004 The hypolipidemic
compound cetaben induces changes in Golgi morphology and vesicle
movement. Histochemistry and Cell Biology 122 95–109.
Lefrancois-Martinez AM, Tournaire C, Martinez A, Berger M, Daoudal S,
Tritsch D, Veyssiere G & Jean C 1999 Product of side-chain cleavage of
cholesterol, isocaproaldehyde, is an endogenous specific substrate of mouse
vas deferens protein, an aldose reductase-like protein in adrenocortical cells.
Journal of Biological Chemistry 274 32875–32880.
Luciani P, Ferruzzi P, Arnaldi G, Crescioli C, Benvenuti S, Nesi G, Valeri A,
Greeve I, Serio M, Mannelli M et al. 2004 Expression of the novel
adrenocorticotropin-responsive gene selective Alzheimer’s disease
indicator-1 in the normal adrenal cortex and in adrenocortical adenomas
and carcinomas. Journal of Clinical Endocrinology and Metabolism 89
1332–1339.
Luciani P, Gelmini S, Ferrante E, Lania A, Benvenuti S, Baglioni S,MantovaniG,
Cellai I,Ammannati F, SpadaA et al. 2005Expressionof the antiapoptotic gene
seladin-1 and octreotide-induced apoptosis in growth hormone-secreting and
nonfunctioning pituitary adenomas. Journal of Clinical Endocrinology and
Metabolism 90 6156–6161.
Magalhaes M, Serra T, Pinto P & Magalhaes M 1991 The effects of monensin
on Golgi complex of adrenal cortex and steroidogenesis. Tissue and Cell 23
209–215.
Journal of Endocrinology (2007) 192, 53–66

M-C BATTISTA and others . Seladin-1 localization in rat adrenal glands66
Martinez A, Aigueperse C, Val P, Dussault M, Tournaire C, Berger M,
Veyssiere G, Jean C & Lefrancois Martinez A 2001 Physiological functions
and hormonal regulation of mouse vas deferens protein (AKR1B7) in
steroidogenic tissues. Chemico-Biological Interactions 130–132 903–917.
Mason JI, Arthur JR & Boyd GS 1978 Control of sterol metabolism in rat
adrenal mitochondria. Biochemical Journal 174 1045–1051.
Mushegian A & Koonin E 1995 A putative FAD-binding domain in a distinct
group of oxidases including a protein involved in plant development. Protein
Science 4 1243–1244.
Nordby G & Norum KR 1975 Substrate specificity of lecithin:cholesterol
acyltransferase. Esterification of desmosterol, b-sitosterol, and cholecalci-
ferol in human plasma. Scandinavian Journal of Clinical and Laboratory
Investigation 35 677–682.
Otis M, Campbell S, Payet MD & Gallo-Payet N 2005 Angiotensin II
stimulates protein synthesis and inhibits proliferation in primary cultures of
rat adrenal glomerulosa cells. Endocrinology 146 633–642.
Pomeraniec Y, Grion N, Gadda L, Pannunzio V, Podesta EJ & Cymeryng CB
2004 Adrenocorticotropin induces heme oxygenase-1 expression in adrenal
cells. Journal of Endocrinology 180 113–124.
Rainey WE 1999 Adrenal zonation: clues from 11beta-hydroxylase and
aldosterone synthase. Molecular and Cellular Endocrinology 151 151–160.
Rainey WE, Carr BR, Wang ZN & Parker CR Jr 2001 Gene profiling of
human fetal and adult adrenals. Journal of Endocrinology 171 209–215.
Rainey WE, Parker CR Jr, Rehman K & Carr BR 2002 The adrenalgenetic
puzzle: how do the fetal and adult pieces differ?Endocrine Research 28 611–622.
Salzmann C, Otis M, Long H, Roberge C, Gallo-Payet N &Walker CD 2004
Inhibition of steroidogenic response to adrenocorticotropin by leptin:
implications for the adrenal response to maternal separation in neonatal rats.
Endocrinology 145 1810–1822.
Sarkar D, Imai T, Kambe F, Shibata A, Ohmori S, Siddiq A, Hayasaka S,
Funahashi H & Seo H 2001 The human homolog of Diminuto/Dwarf1
gene (hDiminuto): a novel ACTH-responsive gene overexpressed in benign
cortisol-producing adrenocortical adenomas. Journal of Clinical Endocrinology
and Metabolism 86 5130–5137.
Sewer M & Waterman M 2003 ACTH modulation of transcription factors
responsible for steroid hydroxylase gene expression in the adrenal cortex.
Microscopy Research and Technique 61 300–307.
Journal of Endocrinology (2007) 192, 53–66
Singh LP, Green K, Alexander M, Bassly S & Crook ED 2004 Hexosamines
and TGF-beta1 use similar signaling pathways to mediate matrix protein
synthesis in mesangial cells. American Journal of Physiology. Renal Physiology
286 F409–F416.
Takahashi T, Gasch A, Nishizawa N & Chua NH 1995 The DIMINUTO
gene of Arabidopsis is involved in regulating cell elongation. Genes and
Development 9 97–107.
Vinson GP 2003 Adrenocortical zonation and ACTH.Microscopy Research and
Technique 61 227–239.
Vinson GP 2004 Glomerulosa function and aldosterone synthesis in the rat.
Molecular and Cellular Endocrinology 217 59–65.
WaterhamHR,Koster J,RomeijnGJ,HennekamRC,VrekenP,AnderssonHC,
FitzPatrick DR, Kelley RI &Wanders RJ 2001 Mutations in the 3beta-
hydroxysterol Delta24-reductase gene cause desmosterolosis, an autosomal
recessive disorder of cholesterol biosynthesis.American Journal ofHumanGenetics
69 685–694.
Wolkersdorfer G & Bornstein S 1998 Tissue remodelling in the adrenal gland.
Biochemical Pharmacology 56 163–171.
Wu C, Miloslavskaya I, Demontis S, Maestro R & Galaktionov K 2004
Regulation of cellular response to oncogenic and oxidative stress by
Seladin-1. Nature 432 640–645.
Yang C, McDonald JG, Patel A, Zhang Y, Umetani M, Xu F, Mangelsdorf DJ,
Westover EJ, Covey DF, Cohen JC et al. 2006 Sterol intermediates from
cholesterol biosynthetic pathway as LXR ligands. Journal of Biological
Chemistry 281 27816–27826.
Zhang L & Insel PA 2004 The pro-apoptotic protein Bim is a convergence
point for cAMP/protein kinase A- and glucocorticoid-promoted apoptosis
of lymphoid cells. Journal of Biological Chemistry 279 20858–20865.
Received in final form 17 October 2006Accepted 20 October 2006Made available online as an Accepted Preprint30 October 2006
www.endocrinology-journals.org