section of experimental medicine and therapeutics memorial lecture
TRANSCRIPT

Volume 58 June 1965
Section of ExperimentalMedicine and TherapeuticsPresident Professor Eleanor Zaimis mD
Meeting November 241964
Walter Ernest DixonMemorial Lecture
A New Concept of TemperatureControl in the Hypothalamusby ProfessorW Feldberg CBE MA MD FRS(National Institutefor Medical Research,Mill Hill, London)
Walter Ernest Dixon was one of the first to con-ceive the idea of chemical transmission of nerveeffects, that nerves act through the release of adrug-like substance. At the beginning of thecentury (1906) he suggested that the vagus nervemay act on the heart through the release of aninhibitory muscarine-like substance, and he per-formed experiments to test his theory. What gavehim this idea? He was a pharmacologist andknew that a characteristic feature ofan innervatedstructure like the heart is its sensitivity to chemicalsubstances, that is, to drugs. I think it must havebeen his intuition, as a pharmacologist, whichmade him realize that a nerve impulse reachinga chemically sensitive structure like the heartwould be a most inefficient stimulus for the pur-pose for which it was sent out, if it were notendowed with the power to release a drug-likesubstance. It is worth pointing out in this con-nexion that all the early contributions to ourknowledge of how a nerve impulse is transmittedto an effector organ were made by pharmacolo-gists.The central synapse is also a chemically sensi-
tive structure. Nerve cells are as sensitive todrugs as are the heart, smooth muscles or glandcells, and to approach the central synapse fromthe pharmacological point of view may give usinformation about the functions of the brainwhich purely electrophysiological methods cannotprovide.To use this approach, however, we need special
methods. It is not sufficient to inject drugs intra-venously or subcutaneously; we must introducethem into the brain so as to avoid peripheraleffects, and we must be able to localize the site ofaction within the brain. This aim is largely
Fig 1 Diagram offrontal section through the cat's brainwith implanted Collison cannula for intraventricularinjections
achieved by allowing the drugs to penetrate thebrain from its inner surface, that is, from thecerebral ventricles. Before I come to the maintopic of my lecture I shall discuss briefly ourmethods and, for one drug, how they work.
All our experiments were done on cats, andFig 1 shows the specially constructed cannula weuse to inject drugs into the cerebral ventricles,In an aseptic operation, the cannula is screwedinto the skull in such a way that its tip rests in alateral ventricle. The butt outside the skull isclosed by a cap with a rubber diaphragm.Through this diaphragm the drugs are painlesslyinjected in the following weeks or monthswithout the cat being anesthetized.
Fig 2 is an X-ray photograph after an intra-ventricular injection of iophendylate (Myodil).It indicates the sites reached by drugs injectedthrough such an indwelling cannula, and italsooutlines the shape of the ventricular system inthe cat. This outline is used for the diagrams ofFigs 4 and 5. The contrast medium has filled theanterior and inferior horns of the lateral ven-
9 395

396 Proceedings ofthe Royal Society ofMedicine
Fig 2 X-rayphotograph ofthe ventricular systemof the cat's brain after an injection of Myodilthrough the implanted Collison cannula shown inFig 1. (From afilm by Feldberg& Sherwood 1963)
tricles as well as the third ventricle around themassa intermedia, except for a small part in frontof the massa. The aqueduct stands out as a thinline, and the fourth ventricle is seen as a triangle.The Myodil has also entered the subarachnoidspace around the brain stem through the for-amina of Luschka so the whole ventricular systemand parts of the subarachnoid space are flooded.Drugs injected through such a cannula maytherefore act on any structure situated near theventricular lumen and near the surface of thebrain stem. How can we distinguish betweenthese sites of action?
This has become possible for those effectswhich are not abolished by anLesthesia, becausewe can perfuse the ventricles under anasthesia.With the methods now available we can excludethe subarachnoid space from the perfusion, andwe can limit the action of the perfused drug to agiven part of the ventricular system.
Methodsfor Perfusing Given Parts oftheCerebral Ventricles with DrugsTo exclude the subarachnoid space we perfusefrom a lateral ventricle and collect the outflowfrom a cannula passed through the openedcisterna along the floor of the fourth ventricle intothe middle of the aqueduct (Bhattacharya &Feldberg 1958). With this method, which isillustrated diagrammatically in Fig 3, we excludefrom the perfusion not only the subarachnoidspace, but also the fourth ventricle.The passage of a drug can be further limited
to a given part of the perfused area by perfusingthe other parts with artificial cerebrospinal fluid(CSF); the area perfused remains unchanged. Toexclude a drug from one of the two lateral
ventricles, both are cannulated and then per-fused through the two cannulh, but with separateinjectors, one delivering the drug, the otherartificial CSF. The outflow is again from theaqueduct. Drugs perfused in this way thus passonly through one lateral ventricle, the thirdventricle, and the rostral part of the aqueduct.
This still leaves large areas of grey matterexposed to the perfused drugs. As illustrated inFig 4, the walls of the anterior horn contain theolfactory grey matter, the septum and caudatenucleus, the walls of the inferior horn, the hippo-campus and amygdala. From the third ventriclethe drugs may penetrate the hypothalamus andthe thalamus, particularly its massa intermediaand habenula, and from the aqueduct they mayenter the central grey matter.The final stage, the regional perfusion with
drugs through a given part of this perfused areaso that a drug irrigates either only a lateralventricle, or only its anterior or inferior horn, orthe third ventricle, or even only that part of it
Fig 3 Diagram ofmedian sagittal section of cat's brainto illustrateperfusionfrom a cannulated lateral ventricleto aqueduct. (From Bhattacharya & Feldberg 1958;drawing modified by Fleischhauer)
10

Section ofExperimental Medicine and Therapeutics 397
Fig 4 Diagram of the ventricular system of the cat'sbrain to illustrate regions of grey matter borderinglateral and third ventricles, as well as aqueduct. Onlythe left lateral ventricle is shown. (Reproduced fromCarmichael et al. 1964a by kindpermission)
lying dorsal or ventral to the massa intermedia,is carried out by multiple cannulation of theventricular system (Carmichael et al. 1964a). Thevarious modifications are illustrated in Fig 5.Four or five cannulk are used, each inserted intoa different part. One cannula acts as an outflow;the others attached to separate injectors serve as
inflow. Outflow and inflow through the cannuleare marked by correspondingly directed arrows.
One, at most two, deliver the drug, the othersartificial CSF. The cannulk delivering the drugs
are solid black, and the parts of the ventricularsystem exposed to the drugs are shaded. Thismethod prevents access of the drug to any partperfused with artificial CSF.A drug is always prevented from entering the
right lateral ventricle by perfusing it withartificial CSF delivered through an implantedcannula. This cannula is not shown in the dia-grams which outline only the left lateral ventricle,third ventricle and aqueduct, with the cannul;inserted into these cavities.To perfuse drugs through the left lateral
ventricle, three cannule are inserted into thisventricle, one into the anterior horn, one into theinferior horn, and a third, which serves as outflow,into the body. Both the inferior and anterior horncannulk deliver the drugs. A fourth cannula isinserted into the aqueduct; it serves as inflow,but of artificial CSF, which thus perfuses thethird ventricle in retrograde direction, and pre-vents the drug from entering it.To perfuse drugs through either the inferior or
anterior horn alone the arrangement remains thesame, but the drugs are delivered through onlyone of the two cannule; the other deliversartificial CSF. These three methods of perfusionare illustrated in Fig 5A, B, C.As shown in Fig 5D, the same arrangement
provides a method for perfusing drugs throughthe third ventricle. For this purpose the drug is
Fig 5 The diagrams give the outline ofthe left lateral ventricle, third ventricle
f ~~~~into these cavities,, to illustrate ther ~~~~regional perluision with drugs through
given parts of the perfused area. Fordetails see text. (Reproduced fromCarmichael et al. 1964a by kindpermission)
11

398 Proceedings ofthe Royal Society ofMedicine
delivered through the aqueductal cannula andperfuses the third ventricle in retrograde direc-tion, whilst its entrance into the lateral ventricleis prevented by the perfusion of artificial CSFthrough the anterior and inferior horn cannulk.With this method it is, however, unavoidable thata small area of the lateral ventricle between theforamen of Monro and the outflow cannula isexposed to the drug. This does not happen whenthe third ventricle is cannulated and the drug isdelivered through this cannula whilst artificialCSF is perfused through the lateral ventricles andthe outflow collected from the aqueductalcannula. This method is illustrated in Fig 5E.Fig 5F shows the method for perfusing drugsthrough either the ventral or dorsal half of thethird ventricle. A double-bore cannula is insertedinto the third ventricle in such a way that theopening of the inner tube lies ventral, that of theouter tube dorsal to the massa intermedia. Thediagram shows the arrangement for the per-fusion of drugs through the ventral half. Thedrug is delivered through the inner, and artificialCSF through the outer tube. For the perfusion ofdrugs through the dorsal half the procedure isreversed.At the end of each experiment the perfusion is
checked by substituting the dye bromphenol bluefor the drug to ascertain which regions are stained,i.e., reached by the drug.
Sites ofAction ofIntraventricular TubocurarineWith the methods described it is possible tolocalize the site where a drug acts within thecerebral ventricles when producing its variouseffects. This will be shown for tubocurarine.Injected intravenously, it produces motor par-alysis, a peripheral effect which is used inanasthesia to bring about muscular relaxation.Tubocurarine, however, is also a potent centralstimulant, although none of its central effectsoccurs on intravenous injection. The reason isthat it cannot reach the nervous structures whenpassing through the brain via the blood stream,because for tubocurarine, as for certain othersubstances, a barrier exists between blood andbrain: the blood-brain barrier. With our methodsof perfusing the cerebral ventricles we circumventthis barrier.Why do I choose tubocurarine, a substance
which can have no physiological function, sinceit does not occur naturally in the brain, andwhich cannot even reach the nervous structures inthe brain when injected into the blood stream?There are three reasons: (1) For the purpose ofillustrating what the methods can do, tubo-curarine is an ideal substance because it has awide spectrum of central effects which originatenot from one but from several sites bordering
Table IEffects of tubocurarine on perfusionthrough the cerebral ventricles
Autonomic effectsMydriasis. Hippus. Withdrawal ofnictitating membranes.Widening ofpalpebral fissures. Pilo-erection. Rise inarterial blood pressure. Salivation. Lacrimation
Motor effects and changes in respirationShivering or tremor. Increased reflex muscularexcitability. Myoclonic jerks. Increase in depth andrate ofrespiration. Swallowing movements
Abnormal activity in the EEGRhythmic discharge of spikes interrupted by periods ofast activity. Rhythmic discharge of 'slow waves'
different parts of the cerebral ventricles. (2) Ananalysis of the central actions of tubocurarinemay lead to a better understanding of somepatterns of abnormal brain activity which occurin certain clinical disorders. (3) Tubocurarine isthe only substance so far fully analysed with themethods of regional perfusion. So there wasreally no choice.
In Table 1 the effects produced by tubocurarinewhen perfused through the cerebral ventricles aresummarized under three headings. The auto-nomic effects, the motor effects and the changesin respiration occurred only when the tubocur-arine passed through the third ventricle; formost of these effects the analysis was carriedfurther and the site could be restricted to the walls,of the ventral half of the third ventricle which areformed by the hypothalamus. These actions thusall resulted from an action on the hypothalamus(Carmichael et al. 1964b).
In contrast, the abnormal activity recorded inthe electrocorticogram resulted from an action onstructures lining the lateral ventricle and thusoccurred only when tubocurarine was perfusedthrough this part of the ventricular system. Butthe two components of this abnormal discharge,the spikes with intermittent periods of fastactivity on the one hand, and the slow waves onthe other, were found to have different sites oforigin. To produce the spike discharge withepisodes of fast activity the tubocurarine had topass through the inferior horn, to produce theslow waves it had to pass through the anteriorhornofa lateral ventricle (Feldberg&Fleischhauer1962, 1963, Carmichael et al. 1964b).An illustration of the abnormal electrical
activity recorded in the electrocorticogram isgiven in Fig 6. The upper two records were takenafter a few minutes' perfusion of tubocurarinethrough the inferior horn; they show the onset ofthe spike discharge with an episode of fastactivity followed by a period of electrical silence,that is, a period in which no abnormal activity isrecorded. The middle records are obtained fromanother experiment and show the slow waves on
12

Section ofExperimental Medicine and Therapeutics 399
A
LO~~~~LB
C
LF~WJJJJ4~14J, LF
O~4 LO
Fig 6 Monopolar records of electrical activity of theleft frontal (LF) and lekf occipital (LO) cortex inanwsthetized cats during perfusion with tubocurarine ofthe left inferior horn (A), the left anterior horn (B) andthe whole left lateral ventricle. Calibration 600 , V.Time in seconds. (Reproduced from Carmichael et al.1964b by kindpermission)
perfusion of tubocurarine through the anteriorhorn, and the lowest records show what happenswhen tubocurarine is perfused through both theanterior and inferior horn. Spikes are super-
imposed on the slow waves and may actuallyarise from them; the slow waves continue duringthe episode and during the period of electricalsilence which follows it.
Three structures from which the slow waves
could originate line the anterior horn: theolfactory grey matter, the caudate nucleus andthe septum (Fig 4). The first has been excludedbecause the slow waves also appear when theolfactory grey matter has been destroyed electro-lytically. No experiments have yet been performedto distinguish between the other two structures,but the septum appears to be the more likely siteof origin because it contains structures whichmay act as a pace-maker for electrical events inthe hippocampus (Petsche & Stumpf 1962).
Fig 4 also shows two structures lining theinferior horn from which the spike discharge withthe intermittent periods of fast activity couldoriginate, amygdala and hippocampus. To estab-lish the origin of the spike discharge, needleelectrodes were inserted into the amygdala andhippocampus, and records taken simultaneously
with the electrocorticogram to find out where thedischarge appears first. This would indicate theirorigin. The result was unequivocal: the spikesappear first in the hippocampus, then in theoccipital cortex and last in the amygdala (Fig 7).We are thus dealing with an action of tubocur-arine on the hippocampus, i.e. with a hippo-campal discharge.Although intraventricular tubocurarine acts at
at least three separate-sites which lie in the wallsof the anterior horn, of the inferior horn and ofthe third ventricle, the effects originating fromthese sites interact.For instance, the abnormal discharge interacts
with the myoclonus and with the autonomicresponses. This becomes evident when both thelateral and the third ventricle are perfused withtubocurarine. Then both myoclonus and auto-nomic effects come under the influence of theabnormal discharge and are reinforced by it.Then each slow wave or spike is seen to coincidewith a jerk, with a small abrupt widening of thepupils and with a small wave of pilo-erection.The slow waves and spikes trigger off theseevents.The interaction of the abnormal discharge on
the diencephalic effects of tubocurarine mayperhaps help us to obtain a clearer understandingof some features of epilepsy encountered in man.In many forms of human epilepsy, includingtemporal epilepsy, we can distinguish betweentwo phenomena: an abnormal electrical activityrecorded in the electroencephalogram, and thefit, the overt seizure. The abnormal electricalactivity may continue for long periods withoutspread to other regions, that is, without loss ofconsciousness and without clonic or tonic con-vulsions. This abnormal electrical activity, or thisfocal discharge, has features in common with theabnormal discharge produced experimentally incats when tubocurarine irrigates the lateral ven-tricle, and it may in fact originate in the hippo-campus. In itself, however, it does not producethe fit. Apparently a change in the excitable stateof other structures, possibly lying in the dien-cephalon, is required before the epileptic focaldischarge can spread to precipitate the overtseizure, in the same way as the abnormal dis-charge produced by tubocurarine will trigger offmotor effects only when the hypothalamus isstimulated by tubocurarine.
Epilepsy, especially temporal lobe epilepsy, istreated in two ways: by surgical eradication of thedeep focal discharging lesion, and by reducing theexcitability of the central nervous system bygiving a barbiturate. In the experiments on catswith intraventricular tubocurarine, pentobarbi-tone sodium was found to depress myoclonuswithout greatly affecting the abnormal discharge.
13

400 Proceedings ofthe Royal Society ofMedicine
A
D
B C
E
In epilepsy, too, the main action of the barbit-urates may therefore not be on the focal dis-charge itself, but on the excitability of those partsof the brain to which the discharge spreads.
A New Concept ofTemperature ControlThe latest development in our studies on theaction of drugs introduced into the cerebral ven-tricles has led to a new concept of temperaturecontrol in the hypothalamus.Whenever a substance, artificially administered,
acts in minute amounts on structures in which itis normally- present, the problem arises ofwhether the pharmacological effects observedmimic a physiological function. Through thework of Vogt (1954) and of Amin et al. (1954),we know that noradrenaline, adrenaline and5-hydroxytryptamine (5-HT) occur naturally inthe walls of the ventral half of the third ventricle,that is, in the hypothalamus: noradrenalineoccurs in a concentration of about 1 1lg/g tissue,the others in much weaker concentrations.
Fig 8, taken from Vogt's paper, shows thedistribution of noradrenaline in the dog's brain.The highest concentration is shown by the blacktriangles. The same distribution applies foradrenaline and 5-HT. When these amines are
Fig 7 Monopolar records of electricalactivity of the left and right occipitalcortex (LO and RO), of the leftamygdala (LA) and the left hippo-campus (LH) during perfusion of thecerebral ventricles. Record A takenbefore, records B, C, D and E, 16, 25, 28and35 min after beginning ofperfusionwith tubocurarine through the leftlateral ventricle. Calibration I m V.Time in seconds. (Reproduced fromFeldberg & Fleischhauer 1963 by kindpermission)
injected into the cerebral ventricles they penetratethe hypothalamus and affect body temperature.Hence the question, do these effects mimic aphysiological function?Body temperature is under the control of the
hypothalamus. But we do not know what goes onin this structure to bring about this control. Is itthe release of noradrenaline, adrenaline and5-HT as suggested by their pharmacologicaleffects when injected intraventricularly?How did it happen- that Dr Myers and I
investigated the effect of these amines on tem-
Fig 8 Medial sagittal section ofa dog's brain. Distribu-tion of noradrenalie. A, 10 f.glg; +, >0 4 <1 0i&glg; o, >0-3 <0.4 pg/g; -, >0-2 <0-3 zg/g freshtissue. (Reproducedfrom Vogt 1954 by kindpermission)
~~~~~~~~~~~~~T T r -1
14
L .-.j --.J.L- -A .-..- i -1.1 I.---LHI
W-1-Immilli I III--d' I"llil iiii -14RO -, -,r-- -T .1--- ITII 'I . .., -1 lrlw -71 m --O.
M La J A ,-AIU-- i1j.1al,m hol iki' -I INI -1LO ..-.11-1 r -11 -Irrr--nlq, T-rrwl- -T-1--T I FfW -T- WT 7-"
ff [III Alla A ,Ili I 1.1 ii..k I ji I.ALA 41.1-11- W
Section of Experimental Medicine and Thlerapeutics 401
oc
3910 '*-I.~~~~~~~~~~~I[v-vo*/*-*@\ /-\~~~/I
I I\
\,
\ I./F SOA "
DU!N2 3 4 5 6 7 8 9 10
HOURS
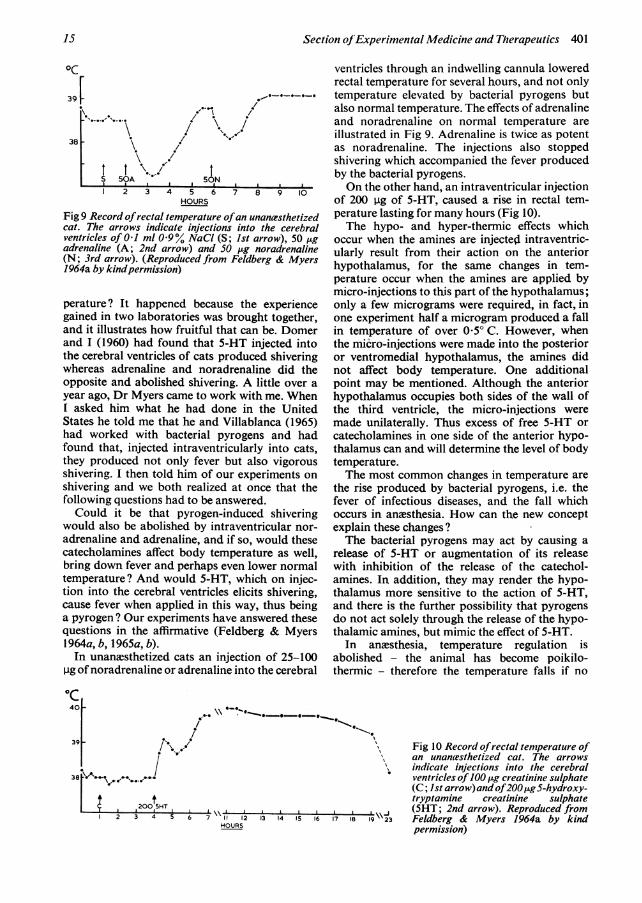
Fig 9 Record ofrectal temperature ofan unanxsthetizedcat. The arrows indicate injections into the cerebralventricles of 0 1 ml 09% NaCI (S; Ist arrow), 50 ,ugadrenaline (A; 2nd arrow) and S0 ug noradrenaline(N; 3rd arrow). (Reproduced from Feldberg & Myers1964a by kindpermission)
perature? It happened because the experiencegained in two laboratories was brought together,and it illustrates how fruitful that can be. Domerand I (1960) had found that 5-HT injected intothe cerebral ventricles of cats produced shiveringwhereas adrenaline and noradrenaline did theopposite and abolished shivering. A little over ayear ago, Dr Myers came to work with me. WhenI asked him what he had done in the UnitedStates he told me that he and Villablanca (1965)had worked with bacterial pyrogens and hadfound that, injected intraventricularly into cats,they produced not only fever but also vigorousshivering. I then told him of our experiments onshivering and we both realized at once that thefollowing questions had to be answered.Could it be that pyrogen-induced shivering
would also be abolished by intraventricular nor-adrenaline and adrenaline, and if so, would thesecatecholamines affect body temperature as well,bring down fever and perhaps even lower normaltemperature? And would 5-HT, which on injec-tion into the cerebral ventricles elicits shivering,cause fever when applied in this way, thus beinga pyrogen ? Our experiments have answered thesequestions in the affirmative (Feldberg & Myers1964a, b, 1965a, b).In unanesthetized cats an injection of 25-100
pg ofnoradrenaline or adrenaline into the cerebral
oc40 -
39 / ;
38 \A
200 5HT2 3 4 5 6 7 \\ 11 12 3 14 IS 16 17 IB 19\23
HOURS
ventricles through an indwelling cannula loweredrectal temperature for several hours, and not onlytemperature elevated by bacterial pyrogens butalso normal temperature. The effects of adrenalineand noradrenaline on normal temperature areillustrated in Fig 9. Adrenaline is twice as potentas noradrenaline. The injections also stoppedshivering which accompanied the fever producedby the bacterial pyrogens.On the other hand, an intraventricular injection
of 200 pg of 5-HT, caused a rise in rectal tem-perature lasting for many hours (Fig 10).The hypo- and hyper-thermic effects which
occur when the amines are injected intraventric-ularly result from their action on the anteriorhypothalamus, for the same changes in tem-perature occur when the amines are applied bymicro-injections to this part of the hypothalamus;only a few micrograms were required, in fact, inone experiment half a microgram produced a fallin temperature of over 0.50 C. However, whenthe micro-injections were made into the posterioror ventromedial hypothalamus, the amines didnot affect body temperature. One additionalpoint may be mentioned. Although the anteriorhypothalamus occupies both sides of the wall ofthe third ventricle, the micro-injections weremade unilaterally. Thus excess of free 5-HT orcatecholamines in one side of the anterior hypo-thalamus can and will determine the level of bodytemperature.The most common changes in temperature are
the rise produced by bacterial pyrogens, i.e. thefever of infectious diseases, and the fall whichoccurs in anesthesia. How can the new conceptexplain these changes?The bacterial pyrogens may act by causing a
release of 5-HT or augmentation of its releasewith inhibition of the release of the catechol-amines. In addition, they may render the hypo-thalamus more sensitive to the action of 5-HT,and there is the further possibility that pyrogensdo not act solely through the release of the hypo-thalamic amines, but mimic the effect of 5-HT.
In anesthesia, temperature regulation isabolished - the animal has become poikilo-thermic - therefore the temperature falls if no
Fig 10 Record ofrectal temperature ofan unanmsthetized cat. The arrowsindicate injections into the cerebralventricles of100 ,ig creatinine sulphate(C; Ist arrow)andof200HgS-hydroxy-tryptamine creatinine sulphate(5HT; 2nd arrow). Reproduced fromFeldberg & Myers 1964a by kindpermission)
15
38

402 Proceedings ofthe Royal Society ofMedicine
- 36Z.,
S 35
0. 34EI-
6 7 8 9 1011 1213141516171819Time (hr)
Fig 11 Effect of intravenousinjections at zero hour, of 45, 50and 60 mg/kg chloralose on rectaltemperature of cats. The recordsfor 45 and 50 mg/kg were obtainedfrom the same cat at an interval ofone month. (Reproduced fromFeldberg & Myers 1964b by kindpermission)
external heat is applied. In cats given intravenousinjections of 45, 50 and 60 mg/kg of the anms-thetic chloralose the temperature falls (Fig 11).A similar fall is obtained with intraperitonealinjection of pentobarbitone sodium, but thetemperature usually recovers more steeply, andthe animal shivers vigorously.The fall in temperature produced by anms-
thetics can be attributed to their action on thehypothalamus because they also lower bodytemperature when given by the intraventricularroute in doses too small to be effective onsystemic application. In the unanesthetized cat afall in temperature can be obtained with intra-ventricular injections of less than 1 mg chloraloseor with 3 mg pentobarbitone sodium.
Intraventricular pentobarbitone has a doubleeffect; a fall followed by a rise beyond the pre-injection level. With successive injections eachfall is followed by a rise, so that finally fever isreached, and the onset of each rise is associatedwith vigorous shivering. Thus pentobarbitoneclearly has a pyrogenic action.
Shivering after intraperitoneal pentobarbitonehas been known for a long time, has been usedfor studying the mechanism of shivering (Lippoldet al. 1959), and is abolished by small amounts ofadrenaline or noradrenaline given intraventric-ularly (Domer & Feldberg 1960).With chloralose the analysis was carried a stage
further. To restrict its action to the walls of theventral half of the third ventricle, we proceededas follows: In cats anmsthesia was induced bypentobarbitone sodium and all parts of the ven-tricles were perfused with artificial CSF. Theanmsthesia naturally produced a fall in temper-ature, but we waited until anxsthesia lightenedand temperature began to rise, and then perfusedchloralose through the third ventricle (Fig 12).
. ...
e.zi r{;5N *,;
Fig 12 Record of rectal temperature of a cat duringperfusion of the cerebral ventricles. The record beginsimmediately after induction of anasthesia with intra-peritoneal pentobarbitone sodium 33 mg/kg. Perfusionof chloralose 1/2,500 for 15 min through the dorsal(at A) and ventral (at B) halfofthe third ventricle. Thearrangement for perfusing separately the two halves ofthe third ventricle is indicated above. (Repro-ducedfrom Feldberg & Myers 1965b by kindpermission)
16
t7
17 Section ofExperimental Medicine and Therapeutics 40340 -
39 _-,
38 -
37-u
0
36
~35-
33-
320 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Time (hr)
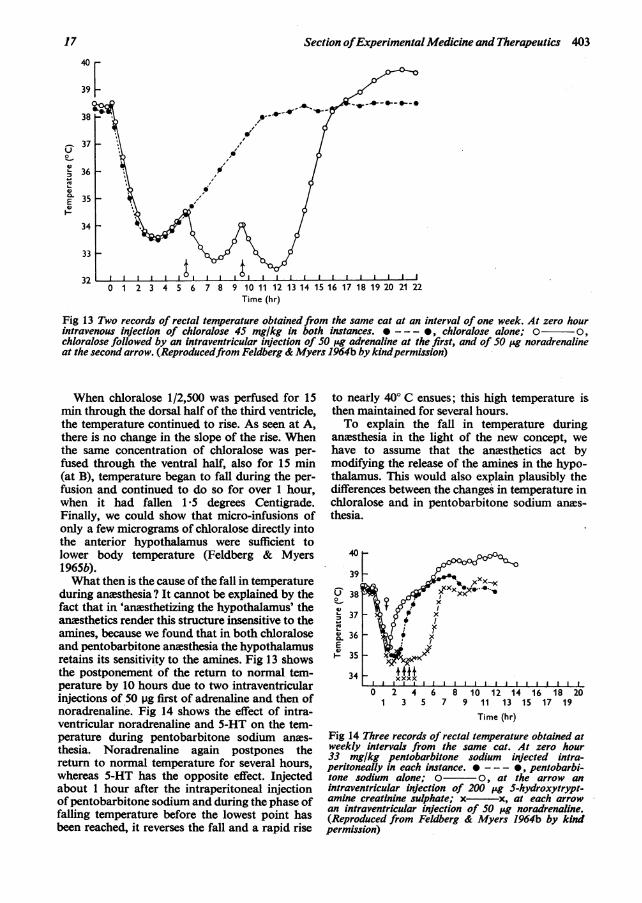
Fig 13 Two records of rectal temperature obtained from the same cat at an interval of one week. At zero hourintravenous injection of chloralose 45 mg/kg in both instances. *--- , chloralose alone; o o,chloralose followed by an intraventricular injection of 50 'ig adrenaline at the first, and of 50 LAg noradrenalineat the second arrow. (Reproducedfrom Feldberg & Myers 1964b by kindpermission)
When chloralose 1/2,500 was perfused for 15min through the dorsal half of the third ventricle,the temperature continued to rise. As seen at A,there is no change in the slope of the rise. Whenthe same concentration of chloralose was per-fused through the ventral half, also for 15 min(at B), temperature began to fall during the per-fusion and continued to do so for over 1 hour,when it had fallen 1'5 degrees Centigrade.Finally, we could show that micro-infusions ofonly a few micrograms of chloralose directly intothe anterior hypothalamus were sufficient tolower body temperature (Feldberg & Myers1965b).What then is the cause of the fall in temperature
during anesthesia? It cannot be explained by thefact that in 'anasthetizing the hypothalamus' theanesthetics render this structure insensitive to theamines, because we found that in both chloraloseand pentobarbitone anmsthesia the hypothalamusretains its sensitivity to the amines. Fig 13 showsthe postponement of the return to normal tem-perature by 10 hours due to two intraventricularinjections of 50 pg first of adrenaline and then ofnoradrenaline. Fig 14 shows the effect of intra-ventricular noradrenaline and 5-HT on the tem-perature during pentobarbitone sodium anms-thesia. Noradrenaline again postpones thereturn to normal temperature for several hours,whereas 5-HT has the opposite effect. Injectedabout 1 hour after the intraperitoneal injectionofpentobarbitone sodium and during the phase offalling temperature before the lowest point hasbeen reached, it reverses the fall and a rapid rise
to nearly 400 C ensues; this high temperature isthen maintained for several hours.To explain the fall in temperature during
anesthesia in the light of the new concept, wehave to assume that the anesthetics act bymodifying the release of the amines in the hypo-thalamus. This would also explain plausibly thedifferences between the changes in temperature inchloralose and in pentobarbitone sodium anes-thesia.
384
0
I I I I II I I
0 2 4 6 8 10 12 14 16 18 201 3 5 7 9 11 13 15 17 19
Time (hr)
Fig 14 Three records of rectal temperature obtained atweekly intervals from the same cat. At zero hour33 mg/kg pentobarbitone sodium injected intra-peritoneally in each instance. * - ---, pentobarbi-tone sodium alone; 0 o, at the arrow anintraventricular injection of 200 pg 5-hydroxytrypt-amine creatinine sulphate; x- x, at each arrowan intraventricular injection of 50 ,ug noradrenaline.(Reproduced from Feldberg & Myers 1964b by kindpermission)
17

404 Proceedings ofthe Royal Society ofMedicine 18
Whether an anesthetic produces a fall intemperature or a fall followed by a rise associatedwith shivering, the fundamental mechanismcould well be the same: the release of the amineswithin the hypothalamus. 'With both chloraloseand pentobarbitone sodium there may be fromthe beginning a release or increased release of allthree amines, but initially the effect of thecatecholamines would mask that of the released5-HT. This could be due to the fact that theamounts of noradrenaline available for release inthe hypothalamus are many times greater thanthose of 5-HT. The effect of 5-HT would becomeapparent some time later when the release orincreased release of the amines ceased. Thisappears plausible because on intraventricularinjection the hyperthermic effect of 5-HT lastslonger than the hypothermic effect ofthe catechol-amines. The difference in action between the twoanesthetics, chloralose and pentobarbitonesodium, might also occur only when their effectson the hypothalamus wear off, and there arevarious possible ways ofexplaining this difference.Our purely pharmacological experiments
neither prove nor disprove these ideas. To provethem we would have to demonstrate that theamines are actually released in the hypothalamusduring anaesthesia. In the last few months wehave begun to tackle this problem.We had to decide whether to look first for the
release of the catecholamines or of 5-HT, andwe decided on 5-HT. The isolated strip of therat's stomach fundus is a very sensitive prepara-tion for assaying 5-HT as shown by Vane (1957).When suspended in a 5 ml bath, it contracts tofess than a nanogram (a millionth of a milligram).When we added our fluid from the perfused
third ventricle to the bath containing such apreparation it contracted, and the contractionwas inhibited by small doses of 2-bromo-lysergicacid diethylamide, a specific inhibitor of 5-HT.This makes it pretty certain that the substance inthe perfusate is 5-HT (Feldberg & Myers 1965c.)Although a definite correlation between the
amounts of 5-HT released and temperature hasnot yet been established, there was often astriking coincidence: a rise in 5-HT activity of asample collected when the low temperature pro-duced by the anesthesia began to rise and the catstarted to shiver. One difficulty we encounteredwas that the samples of perfusate contained inter-fering substances which made the assay difficult.One further result may explain a phenomenon
which has long puzzled neurosurgeons: the brainfever and the shivering bouts that occur in certaincases of brain injury. We thought we must find
out how long it takes for the 5-HT activity todisappear from the perfusate when perfusion ofthe third ventricle was continued after thecat was killed, i.e. in the dead animal. To oursurprise, the 5-HT activity did not decrease, butincreased 3 to 24 times in the samples collectedduring the first 30-60 min after death, and onlythen diminished gradually. What is the meaningof this post-mortem effect, of this life after deathin the hypothalamus?
If we take the view which our pharmacologicalfindings impose upon us, that temperature isthe outcome of a fine balance in the release ofcatecholamines and of 5-HT in the hypothalamus,it would appear that the basal mechanism is therelease of 5-HT in order to keep up temperature.As soon as the blood supply to this part of thebrain is interfered with, 5-HT is released at anabnormally high rate. This happens after death,and if the animal were not dead its temperaturewould rise and it would shiver. The same mayhappen in brain injuries after accidents whensmall vessels in the hypothalamus may tear and,as a result of an abnormally high release of 5-HTin the regions which no longer receive theirnormal blood supply, temperature rises andshivering sets in. Supporting evidence for thisview may be the finding by.Sachs (1957) of 5-HTin the lumbar cerebrospinal fluid of a number ofpatients following head injury.The aim of my lecture was to demonstrate our
procedure of approaching the brain from its innersurface, that is, from the cerebral ventricles,because I think this approach may give us newand fundamental information about how thebrain works, and this information, we hope, mayultimately also help the clinician.
REFERENCESAmin A H, Crawford T B B & Gaddum JH(1954) J. Physiol. 126,596Bhattacharya B K & Feldberg W (1958) Brit. J. Pharmacol. 13, 156Carmichael E A, FeldbergW & Fileischhauer K(1964a) J. Physiol. 173, 354(1964b) J. Physiol. 175, 303DixonW (1906) Brit. med. J. ii, 1807Domer F R & FeldbergW (1960) Brit. J. Pharmacol. 15, 578FeldbergW & Fleischhauer K(1962) J. Physiol. 160, 258(1963) J. Physiol. 168,435FeldbergW & Myers RD(1964a) J. Physiol. 173, 226(1964b) J. Physiol. 175, 464(1965a) J. Physiol. 177, 239(1965b) J. Physiol. 178 (in press)(1965c) J. Physiol. (in press)Lippold 0 C J, Redfearn JW T & Vuco J (1959) J. Physiol. 146, 1Myers RD & Villablanca J (1965) Amer. J. Physiol. (in press)Petsche H & Stumpf C(1962) Colloq. int. Cent. nat. Rech. Sci. No. 107, p 121Sachs E jr (1957) J. Neurosurg. 14, 22Vane JR (1957) Brit. J. Pharmacol. 12, 344VogtM (1954) J. Physiol. 123, 451