secondary olfactory projections and pallial topography in the pacific hagfish, eptatretus stouti
TRANSCRIPT

THE JOURNAL OF COMPARATIVE NEUROLOGY 337:529-542 (1993)
Secondary Olfactory Projections and Pallial Topography in the Pacific
Hagfish, Eptatretus stouti
HELMUT WICHT AND R. GLENN NORTHCUTT Klinikum der Johann Wolfgang Goethe-Universitat, Zentrum der Morphologie, Abteilung
Neurobiologie, 60590 Frankfurt, Germany (H.W.); Neurobiology Unit, Scripps Institution of Oceanography, and Department of Neuroscience, School of Medicine, University of California,
San Diego, La Jolla, California 92093 (R.G.N.)
ABSTRACT The extent of the secondary olfactory projections shows great variation among different
groups of craniates. Gnathostomes typically display restricted secondary olfactory projections, whereas lampreys have more extensive projections. Any attempt to determine the phylogenetic polarity of these characters, that is, to decide which is primitive and which is derived, requires an investigation of the secondary olfactory system in the sister group of lampreys and gnathostomes, the haghhes. Therefore the secondary olfactory projections of the Pacific hagfish, Eptatretus stouti, were traced with the use of horseradish peroxidase and the lipophilic fluorescent tracing compound DiI. The projections are bilateral and massive to all pallial areas and the septum, moderate to the striatum, and relatively weak to the preoptic and infundibular regions of the hypothalamus, reaching caudally to the diencephalic-mesencephalic boundary. Merents to the olfactory bulb arise from the pallium, the preoptic area, and the ventral thalamus. We compare the secondary olfactory projections in hagfishes with those in lampreys and in gnathostomes, and we conclude that the presence of extensive secondary olfactory projections is a primitive character of craniate brains. c 1993 Wiley-Liss, Inc.
Key words: Myxinoidea, forebrain, olfactory system, pallium, phylogeny
In all craniates there is an intimate embryological, mor- phological, and functional connection between the telen- cephalon and the olfactory system; any study of telence- phalic evolution must therefore take into account the olfactory system in general and the distribution of second- ary olfactory projections in particular. I t was long thought that the telencephalon of anamniotic craniates was domi- nated by secondary olfactory projections and was therefore chiefly concerned with the processing of olfactory informa- tion. The extensive nonolfactory areas that characterize the telencephalon of amniotes were thought to have arisen relatively late in phylogeny, together with the appearance of the first tetrapods ("smell brain"-theory; Edinger, '04; Ariens Kappers et al., '36; Herrick, '48; Girgis, '70; see Nieuwenhuys, '67, for a review of the classical literature). This scenario, which can still be found in many medical textbooks, is most probably wrong. Over the past three decades numerous experimental studies on the secondary olfactory projections in gnathostomes (for reviews see Heimer, '69; Northcutt, '81; Neary, '90; Nieuwenhuys and Meek, '90a,b; Smeets, '90; Ulinski, '90) have shown that secondary olfactory projections are relatively restricted in all gnathostomes that have been investigated with modern
tracing techniques. The lateral pallium, the septum, and portions of the medial and dorsal pallia have been identified as major olfactorecipient areas in the telencephalon, but gnathostomes also display large telencephalic areas devoid of secondary olfactory input.
It was therefore surprising when Northcutt and Puzd- rowski ('88) discovered relatively extensive secondary olfac- tory projections in an agnathan, the Silver lamprey, Icchthyo- myzon unicuspis. In comparing lampreys and gnathostomes, they could not determine which is the plesiomorphic (primi- tive) condition: restricted or extensive secondary olfactory projections. This question can only be answered by study- ing the relevant outgroup, the hagfishes (Myxinoidea). If they display a pattern of secondary olfactory projections similar to that observed in lampreys, then cladistic theory (Hennig, '66; Eldredge and Cracraft, '80; Wiley, '81; North- cutt, '84) would indicate that extensive secondary olfactory projections are the plesiomorphic pattern for craniates.
Accepted July 2, 1993. Address reprint requests to Helmut Wicht, Klinikum der Johann Wolf-
gang Goethe-Universitat, Zentrum der Morphologie, Abteilung Neurobiolo- pie, Theodor-Stern-Kai 7,60590 Frankfurt, FRG.
D 1993 WILEY-LISS, INC.

530 H. WICHT AND R.G. NORTHCUTT
Alternatively, if hagfishes possess restricted secondary olfactory projections, then this pattern would be considered plesiomorphic and the pattern observed in lampreys an apomorphic (derived) trait.
Until recently, attempts to study and compare forebrain organization in hagfishes were impeded by a century-old controversy regarding forebrain homologies in these ani- mals. The forebrain of myxinoids is actually unique with respect to the organization of the ventricular system (which is greatly reduced and lacks a lumen in most areas of the forebrain) and with respect to the organization of the pallium (which displays five alternating cellular and fibrous layers). We recently completed a study of the forebrain of myxinoids (Wicht and Northcutt, '92a), which resulted in a reinterpretation of many of the cell masses in the forebrain. In particular, we have argued that one area that has traditionally been regarded as part of the pallium (the central prosencephalic nucleus) is actually not a pallial structure. Although many questions remain unanswered, we believe we now have a sound basis for describing the secondary olfactory projections in hagfishes. Some of our data have appeared in abstract form (Northcutt, '85).
MATERIALS AND METHODS Adult Pacific hagfish, Eptatretus stouti (body length
approximately 400 mm) were trapped off the coast of La Jolla, California at depths of approximately 200 m. The connections of the olfactory bulb were determined by using either horseradish peroxidase (HRP) or the fluorescent compound 1,1'-dioctadecyl-3,3,3',3-tetramethylindocarbo- cyanine perchlorate (DiI) as a tracer.
For HRP tracing, the animals (12) were anesthetized in tricaine-methanesulfonate (MS 222) dissolved in seawater (1:10,000) and packed in crushed ice, and the olfactory bulbs were surgically exposed. The HRP (Sigma, type VI) was dried on the tip of an insect pin and introduced into one olfactory bulb. The wound was then closed, sutured and glued (Vetbond), and the animals were allowed to survive for 7-15 days in seawater tanks at 4°C. Afterwards, they were reanesthetized and perfused transcardially with 0.1 M
phosphate buffer at pH 7.4, followed by a solution of 2% glutaraldehyde in the same buffer. The brains were dis- sected from the skulls and postfixed for 2 hours in cold buffered perfusate with 20% sucrose added for cryoprotec- tion. They were then embedded in 15% gelatin, postfixed overnight, and cut on a freezing microtome at 40 km in the transverse plane. The HRP was visualized by either the protocol of Hanker et al. ('77) or the method of Adams ('811, and the sections were counterstained with either methylene green, cresyl violet, or neutral red.
For DiI tracing, the animals (8) were deeply anesthetized and perfused with 0.1 M phosphate buffer at pH 7.4, fol- lowed by a solution of 4% paraformaldehyde in phosphate buffer. The fixed brains were dissected from the skulls, and crystals of DiI (Molecular Probes, Eugene, Oregon) were introduced into one olfactory bulb with an insect pin or fine forceps. Brains were then stored in 4% paraformaldehyde in phosphate buffer for 1-4 weeks at 40°C. During this storage, the lipophilic DiI spreads by passive diffusion along the plasma membranes of cells that are in contact with the injection site, thus revealing afferent and efferent connec- tions (see Godement et al., '87). After this period, the brains were embedded in egg yolk, and the blocks were postfixed in 4% paraformaldehyde in phosphate buffer overnight and cut on a vibratome at 50 km. Sections were collected in phosphate buffer, counterstained with m-phenylene di- amine, and mounted on glass slides. See Wicht and North- cutt ('90) for technical details of counterstaining. Sections were inspected and photographed on a microscope equipped with epifluorescence.
Throughout this paper we use our own terminology for forebrain areas in myxinoids. This terminology was estab- lished in a previous study (Wicht and Northcutt, '92a) and related to the classical terminology originated by Jansen ('30).
RESULTS The results obtained with HRP and those obtained with
DiI were consistent but not identical. Typically, the HRP injection sites were confined to the olfactory bulbs and
BO BOci Bog BOm BOn BOP coh coib cop0 cotp Di dp frf Gnatho. HA HYinf Inp IP mP Myx. NC NCd NCvl Pl-P5 P2mc P41 P4m
olfactory bulb internal cellular layer of BO glomerular layer of BO mitral cell layer of BO olfactory fiber layer of BO periglomerular layer of BO habenular commissure interbulbar commissure postoptic commissure commissure of the posterior tuberculum diencephalon dorsal pallium retroflex fascicle Gnathostomata habenula infundibular hypothalamus in terpeduiicular nucleus lateral pallium medial pallium Myxinoidea central prosencephalic nucleus dorsal subnucleus of NC ventrolateral subnucleus of NC pallial layers 1-5 magnocellular part of P2 lateral subdivision of P4 medial subdivision of P4
Abbreviations
Petro. PO Se SP St Tel Tha THdi THe THi Thsh Tl l t TM TP trolp trols trom trov vac vinf VJ vsh I V VII IX X
Petromyzon tidae preoptic region septum spinal nerves striatum telencephalon anterior nucleus of thalamus diffuse nucleus of the thalamus external nucleus of thalamus internal nucleus of thalamus subhabenular nucleus of thalamus triangular nucleus of thalamus mesencephalic tectum posterior tubercular area deep part of lateral olfactory tract superficial part of lateral olfactory tract medial olfactory tract ventral olfactory tract cerebral aqueduct infundibular recess of third ventricle preoptic recess of third ventricle subhabenular ventricle (part of third ventricle) roots of olfactory nerve trigeminal nerve and ganglion facial nerve glossopharyngeal nerve vagal nerve

HAGFISH SECONDARY OLFACTORY PROJECTIONS
2A 2B 3 A 3 B 4 A 4 B
53 1
Fig. 1. Dorsal view of the brain of the Pacific hagfish, Eptutretus stouti. Roman numerals indicate cranial nerves, and vertical lines indicate levels of individual sections in Figures 2-4. Scale bar: 1 mm.
excluded the most lateral and medial parts of the bulb (Fig. 2A). The DiI injection sites were much larger and often included the rostra1 striatal and pallial areas, which adjoin the olfactory bulb caudally. As might be expected, the DiI cases showed much more extensive projections than the HRP cases, including projections reaching as far as the mesencephalic tegmentum. Our descriptions are therefore based chiefly on the HRP material, and we report only those connections revealed by both HRP and DiI.
A dorsal view of the brain of the Pacific hagfish is shown in Figure 1, along with the levels of the individual sections of Figures 2 ,3 , and 4, which are chartings of the secondary olfactory projections. As the organization of the secondary olfactory tracts is complex, there is a stylized three- dimensional reconstruction of the projections in Figure 6.
Caudal to the injection site, secondary olfactory fibers collect in the internal cellular layer of the olfactory bulb (Fig. 2B). There is, however, no distinct secondary olfactory fiber layer; the fibers intermingle with the cell bodies of the internal cellular layer. At the same level, the secondary olfactory fibers separate into three major fiber tracts: the medial, ventral, and lateral olfactory tracts, the last of which comprises superficial and deep subdivisions. Each of
and terminates in the septal area, which is wedged between the left and right olfactory bulbs (Figs. 2B, 5C). By way of this tract some fibers also reach the contralateral olfactory bulb, and a few single labelled cells were seen in the internal cellular layer of the contralateral bulb (Fig. 2B).
Ventral olfactory tract The ventral olfactory tract leaves the bulb ventrolaterally
(Fig. 2B) joins the lateral forebrain bundle in the preoptic region (Figs. 3A,B), and continues caudally through the diencephalon (Fig. 4A). I t gives off a few fibers into the postoptic commissure (Fig. 3B, 5D), but the course of these fibers could not be followed beyond their decussation. The ventral olfactory tract ends at the mesodiencephalic border (Fig. 4B), at the level of the commissure of the posterior tuberculum. Occasionally, individual fibers, which appar- ently stem from the ventral olfactory tract, were observed in the infundibular nucleus of the hypothalamus, the dorsal thalamus, and the triangular nucleus of the thalamus (Fig. 4A,B). There was no distinct terminal field associated with this tract. Throughout its extent in the forebrain, labelled perikarya (Fig. 5D) were seen in association with this tract.
these tracts will be described separately, as well as the relationships of the secondary olfactory fiber system, the central prosencephalic nucleus, and a particular subdivi- sion of the pallium (medial part of pallial layer 4).
Medial olfactory tract The medial olfactory tract is the shortest of the three
major tracts. It leaves the olfactory bulb ventromedially
Lateral olfactory tract The lateral olfactory tract is the largest of all secondary
olfactory tracts. Its superficial subdivision leaves the olfac- tory bulb dorsolaterally (Fig. 2B) and maintains a subpial position throughout the hemispheres (Figs. 3A,B, 4A), thus forming the outermost of the five pallid layers (Pl) . Along its course it gives off fibers to the underlying cellular pallid

532 H. WICHT AND R.G. NORTHCUTT
A
B Fig. 2. Drawings of transverse sections midway through the olfac-
tory bulb (A) and through the rostra1 hemisphere (B), charting the secondary olfactory projections after an injection of tracer into the right olfactory bulb. The blackened areas represent the tracks of the needle
used to inject the tracer; the stippled area represents the approximate extent of a typical horseradish peroxidase (HRP) injection site. Labelled fibers are symbolized by thin lines, labelled perikarya by triangles.
layer, P2 (Fig. 5B). In most of the HRP preparations, there are conspicuous gaps in the labelling of the superficial olfactory tract (Fig. 3B, 5A). Since none of the injection
sites involved the entire olfactory bulb (see above), these gaps may indicate a topographical organization of the bulbo-pallid projection.

HAGFISH SECONDARY OLFACTORY PROJECTIONS 533
B Fig. 3. Drawings of transverse sections midway through the hemisphere (A) and at the level of the inter-
bulbar commissure (B), charting the secondary olfactory projections (continuation of Fig. 2). Arrow points to a gap in the labelling of the superficial part of the lateral olfactory tract. Other conventions as for Figure 2.

534 H. WICHT AND R.G. NORTHCUTT
B
Fig. 4. Drawings of transverse sections at the diencephalo-telencephalic boundary (A) and midway through the diencephalon (B) charting the secondary olfactory projections (continuation of Fig. 3). Conven- tions as for Figure 2.
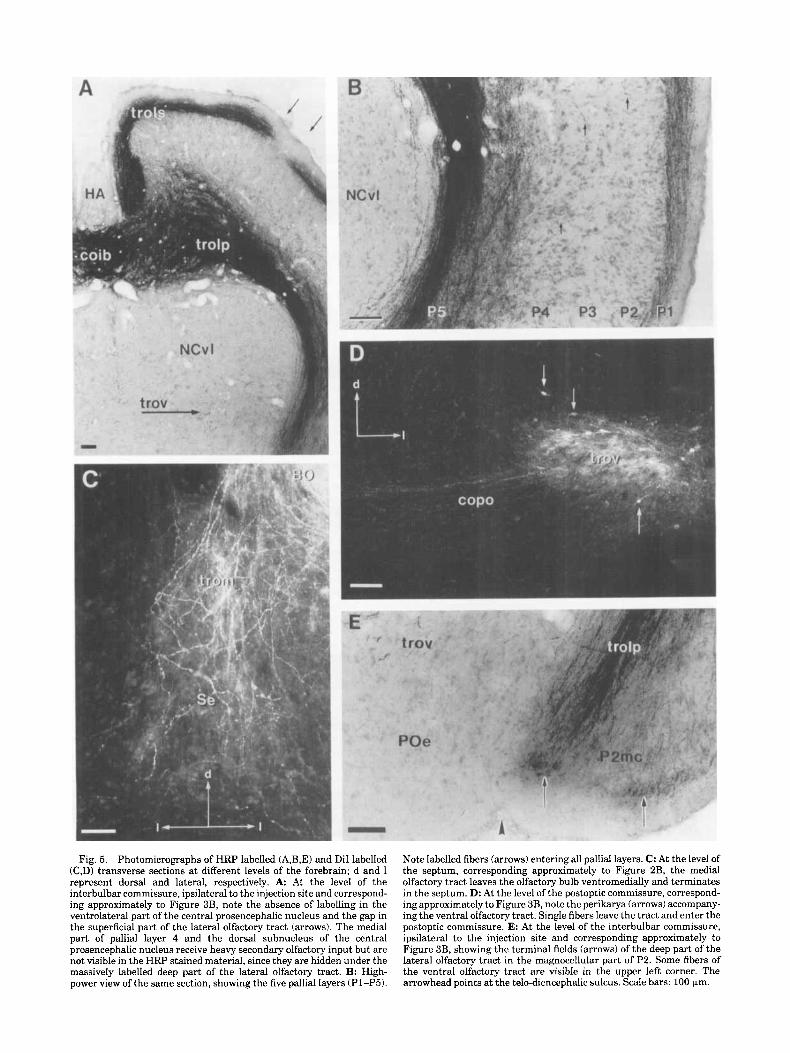
Fig. 5. Photomicrographs of HRP labelled (A,B,E) and DiI labelled (C,D) transverse sections at different levels of the forebrain; d and I represent dorsal and lateral, respectively. A At the level of the interbulbar cornmisswe, ipsilateral to the injection site and correspond- ing approximately to Figure 3B, note the absence of labelling in the ventrolateral part of the central prosencephalic nucleus and the gap in the superficial part of the lateral olfactory tract (arrows). The medial part of pallid layer 4 and the dorsal subnucleus of the central prosencephalic nucleus receive heavy secondary olfactory input but are not visible in the HRP stained material, since they are hidden under the massively labelled deep part of the lateral olfactory tract. B: High- power view of the same section, showing the five pallid layers (Pl-P5).
Note labelled fibers (arrows) entering all pallid layers. C: At the level of the septum, corresponding approximately to Figure 2B, the medial olfactory tract leaves the olfactory bulb ventromedially and terminates in the septum. D: At the level of the postoptic commissure, correspond- ing approximately to Figure 3B, note the perikarya (arrows) accompany- ing the ventral olfactory tract. Single fibers leave the tract and enter the postoptic commissure. E: At the level of the interbulbar commissure, ipsilateral to the injection site and corresponding approximately to Figure 3B, showing the terminal fields (arrows) of the deep part of the lateral olfactory tract in the magnocellular part of P2. Some fibers of the ventral olfactory tract are visible in the upper left corner. The arrowhead points at the telo-diencephalic sulcus. Scale bars: 100 km.

536 H. WICIIT AND R.G. NORTHCUTT
The deep subdivision the lateral olfactory tract leaves the olfactory bulb at its caudal pole (Fig. 3A). I t issues fibers to the striatal region, then curves along the dorsal and lateral margins of the central prosencephalic nucleus (Figs. 3A,B). It thus forms a major part of the deepest pallial layer (P5), which is a mixed layer of fibers and cells. Along its course the deep subdivision of the lateral olfactory tract issues numerous fibers laterally into the overlaying pallial layers P4 and P3 (Figs. 3B, 5B). The deep subdivision of the lateral olfactory tract finally ends with a distinct terminal field in the magnocellular part of the pallial layer P2 (Fig. 3B, 5E).
Both subdivisions of the lateral olfactory tract were also labelled contralaterally. Many decussating secondary olfac- tory fibers were observed in the interbulbar commisure (Fig. 3B), and fewer in the habenular commissure (Fig. 4A). After decussation, the fibers on the contralateral side rearrange to form superficial and deep lateral tracts, which follow the same course to the same targets as their ipsilat- eral counterparts. Fewer labelled fibers were seen on the contralateral side, however. Ipsilateral to the injection site, single lightly labelled perikarya were observed in all pallial layers (Fig. 3A,B).
nucleus, together with the medial part of pallial layer 4, form the above-mentioned arch of cells around the interbul- bar commissure. This arch of cells receives dense innerva- tion from commissural fibers of the deep subdivision of the lateral olfactory tract, which decussate in the interbulbar and habenular commissures (Fig. 3B). Again, only single labelled fibers were seen in the ventrolateral subdivision of the central prosencephalic nucleus at these levels (Fig. 3B).
DISCUSSION As noted above, it was our goal to compare the relative
extent of secondary olfactory projections to the telencepha- lon in myxinoids with that in lampreys and gnathostomes. Obviously, how one defines the telencephalon is critical to such a comparison. Although there is a consensus regarding what constitutes the telencephalon in gnathostomes, appli- cation of the term is much less clear in hagfishes and lampreys. We will therefore review some of the problems related to defining the telencephalon and some of its components in hagfishes before comparing the telence- Dhalic secondarv olfactorv afferents. For a more detailed kiscussion of th;! telenceihalon in agnathans, the reader is referred to Northcutt and Puzdrowski ('88) and Wicht and prosencephalic the
part of pallial layer 4, and their relation to the system of secondary olfactory fibers
Northcutt ( ' 9 ~ ~ ) .
Homology of the pallial areas and the central prosencephalic nucleus of hagfishes The general topography of the central prosencephalic
nucleus and the medial tmrt of Dallial laver 4 is evident in Figure 7A. The central prosencephalic iucleus consists of three subnuclei: dorsal, ventrolateral, and medial (Kusu- noki et al., '81; Wicht and Northcutt, '92a). The medial subnucleus is relatively small and not illustrated. The ventrolateral subnucleus is the largest of these subdivisions and forms an ovoid mass of gray matter in the center of the prosencephalon (Fig. 7A). The rostral pole of this sub- nucleus appears in the core of the forebrain, immediately caudal to the olfactory bulb (Fig. 3A), and the caudal pole is close to the lateral surface of 'the brain in the diencephalon (Fig. 4A). The dorsal subnucleus of the central prosence- phalic nucleus appears a t levels slightly rostral to the interbulbar commissure and, together with the medial part of layer 4 of the pallium, forms a conspicuous arch of cells around the posterior margin of the interbulbar commissure (Figs. 3B, 7A).
It is important to note that, even though the central prosencephalic nucleus borders the olfactory bulb with its rostral pole, it is separated from the bulb rostrally and the pallium laterally by the bowl-shaped vestiges of the lateral ventricles (Fig. 7A). Fibers of the deep subdivision of the lateral olfactory tract surround the central prosencephalic nucleus at its rostral pole (Fig. 3A), but the vestiges of the lateral ventricles apparently form a barrier that prevents the fibers from entering the nucleus. Instead, the deep sub- division of the lateral olfactory tract issues fibers laterally into the overlaying pallial area (Fig. 5B). We occasionally saw single labelled fibers in the rostral part of the ventrolat- eral subnucleus of the central prosencephalic nucleus (Fig. 3A), but their origin remains unclear.
The situation is different more caudally, that is, between the caudal margin of the interbulbar commissure ,and the rostral margin of the habenular commissure (Fig. 7A). There are no vestiges of the lateral ventricles at these levels, and the subnuclei of the central prosencephalic nucleus are in close contact with their surroundings. In this area, the dorsal subnucleus of the central prosencephalic
Based on the presence of a massive secondary olfactory input, there is little doubt that the laminated pallium of hagfishes is a homologue of the lateral pallium of other craniates (Wicht and Northcutt, '92a). I t is not clear, however, whether hagfishes also possess forebrain compo- nents comparable to the medial and dorsal pallia of other craniates.
The central prosencephalic nucleus of hagfishes has traditionally been assigned to the telencephalon and has been homologized with the medial pallium of other crani- ates (Holmgren, '19; Jansen, '30). We recently reviewed the available morphological, embryological, hodological, and histochemical data (Wicht and Northcutt, '92a) and con- cluded that the central prosencephalic nucleus, as a whole, is not part of the telencephalon and is not the homologue of the medial pallium; in all probability it is a rostral dience- phalic structure, although its exact homology has remained unclear. Our present data, as well as some other informa- tion on the connections and immunohistochemistry of the central prosencephalic nucleus, which has become available in the meantime, necessitate a critical reevaluation of this hypothesis, particularly with respect to the individual subnuclei of the central prosencephalic nucleus. The cen- tral prosencephalic nucleus consists of three subnuclei (Kusunoki et al., '81; Wicht and Northcutt, '92a): a very large ventrolateral subnucleus, a small medial subnucleus (not illustrated herein), and an intermediate-sized dorsal subnucleus.
There are strong reciprocal connections between the medial and ventrolateral subnuclei and the preoptic and infundibular hypothalamus, respectively, but barely any connections with pallial areas (F. Amemyia and R.G. North- cutt, unpublished observations). The hypothalamic connec- tion, if interpreted as a fornix, might appear to indicate a homology of the ventrolateral subdivision of the central prosencephalic nucleus and the medial pallium, but the hypothalamic connection is reciprocal in hagfishes, whereas

HAGFISH SECONDARY OLFACTORY PROJECTIONS 537

Figure 7

HAGFISH SECONDARY OLFACTORY PROJECTIONS 539
the fornix of gnathostomes consists primarily of pallio- hypothalamic projections (Northcutt and Ronan, '92). Fur- thermore, if these subdivisions of the central prosence- phalic nucleus and the medial pallium were homologous, one would also expect a prominent input from the olfactore- cipient lateral pallium and another bilateral projection from the dorsal thalamus, as is typical for the medial pallium of other craniates (Scalia and Colman, '75; Kicliter, '79; Northcutt and Kicliter, '80; Neary, '84; Wicht and Himstedt, '88; Northcutt and Ronan, '92).
The medial and ventrolateral subnuclei of the central prosencephalic nucleus of hagfishes, however, lack these connections. In a recent immunohistochemical study, Don- ald et al. ('92) have demonstrated natriuretic peptide- positive cells in the ventrolateral subnucleus of the central prosencephalic nucleus of the Atlantic hagfish, Myxine glutinosa. In elasmobranchs (Donald et al., '92) and tel- eosts (Donald and Evans, '921, however, perikarya positive for this substance do not occur in the medial pallium but, preferentially, in ventral thalamic and preoptic hypotha- lamic areas. Thus, with respect to the interpretation of the ventrolateral subnucleus of the central prosencephalic nucleus as a diencephalic structure, the new data are compatible with the conclusions we drew in our previous paper (Wicht and Northcutt, '92a).
Our interpretation may need revision, however, with respect to the interpretation of the dorsal subnucleus of the central prosencephalic nucleus. The dorsal subnucleus, which is located ventral and posterior to the interbulbar commissure, and rostral to the habenular commissure, does receive significant secondary olfactory input, mainly from fibers related to the interbulbar and habenular commis- sures. Together with an area described as the medial part of pallial layer 4 (Wicht and Northcutt, '92a), which also receives a distinct secondary olfactory input from these
Fig. 7. Comparison of the topography of major telencephalic areas in hagfishes (A) and lampreys (B). Note that the drawing is intended to illustrate the relative position of individual cell groups rather than their accurate shape and size. In the hagfish (A) the lateral pallium is shown on the right side only, and a part of it has been cut away to expose the deeper structures. Color coding: orange, lateral pallium; purple, dorsal pallium (in the lamprey), medial part of pallial layer 4 (in the hagfish); red, medial pallium (in the lamprey), dorsal subnucleus of the central prosencephalic nucleus (in the hagfish); green, striatum; blue, preoptic nuclei; yellow, commissures (interbulbar commissure, rostrally, and habenular commissure caudally); light brown, ventrolateral sub- nucleus of the central prosencephalic nucleus (in the hagfish). The commissures depicted are the interbulbar commissure, rostrally, and the habenular commissure, caudally. Both contain secondary olfactory fibers in lampreys and hagfishes. The medial and dorsal pallia of lampreys (B) are located between these commissures and receive an olfactory input from fibers related to these commissures (Northcutt and Puzdrowski, '88). The arch of cells around the posterior aspect of the interbulbar commissure in hagfishes (P4m and NCd) shows a similar topography, and it also receives secondary olfactory input from fibers related to these commissures, that is, decussating fibers of the deep part of the lateral olfactory tract (see text and Fig. 3B). This arch may therefore constitute a homologue of the dorsal and medial pallia of lampreys. Also note the topographical relationship between the central prosencephalic nucleus and the ventricular system in hagfishes. The (uncolored) structure with the ventrally directed "panhandle" is the diencephalic part of the reduced ventricular system of hagfishes; the panhandle represents the infundibular recess. The vestigial lateral ventricles (also uncolored) connect with the diencephalic ventricular system. They cap the central prosencephalic nucleus rostrally and form a sharp barrier between this nucleus, the olfactory bulb, the striatum, and the laterodorsally adjoining lateral pallium. See Wicht and North- cutt ('92a) for a more detailed description of the ventricular system.
fibers, the dorsal subnucleus forms an arch of cells around the posterior aspect of the interbulbar commissure, bor- dered caudally by the habenular commissure (Figs. 3B, 7A). The medial and dorsal pallia of lampreys, as described by Northcutt and Puzdrowski ('881, display similar topographi- cal and connectional relationships. They are also located between the two commissures (Fig. 7B), and they do receive an input from decussating fibers of the secondary olfactory system. It is therefore possible that the dorsal subnucleus of the central prosencephalic nucleus, and the medial part of pallial layer 4, in hagfishes are homologous to the medial and dorsal pallia of other craniates.
Do a terminal nerve and an anterior olfactory nucleus exist in hagfishes?
After injections of tracers into the olfactory bulb of hag- fishes, a number of retrogradely labelled were found in the laminated (lateral) pallium. A second group of labelled cells was observed in association with the ventral olfactory tract in the preoptic area and in the ventral thalamus. Very few cells were labelled in the internal cellular layer of the contralateral olfactory bulb. Bulbar afferents arise from similar locations in lampreys (Northcutt and Puzdrowski, '88).
It is tempting to interpret the ventral olfactory tract and the labelled cells that accompany it as the terminal nerve, as described in lampreys and other craniates. The course of this tract is similar to that described as the central projec- tion of the terminal nerve in lampreys (von Bartheld et al., '871, and the labelled cells that accompany it might be centrally displaced ganglionic cells of the terminal nerve, which give rise to a retinopetal projection in many other craniates (e.g., Miinz et al., '82; Stell et al., '84, Northcutt and Butler, '91). However, this interpretation is mislead- ing. The ventral olfactory tract of the present study and the cells that accompany it show no immunoreactivity for gonadotropin-releasing hormones (GnRH; Nozaki, '85) or for molluscan cardioexcitatory peptide (FMRFamide), both of which are reliable markers for the terminal nerve (Stell et al., '84). A large number of FMRFamide- and GnRH- positive perikarya occur in the infundibular nucleus of the hypothalamus (Wicht and Northcutt, 1992b; R.G. North- cutt, unpublished observations), but they are not identical to the cells seen in conjunction with the ventral olfactory tract in the present study. Furthermore, although hag- fishes do possess a retinopetal projection, it does not arise from the cells accompanying the ventral olfactory tract but from a cell group in the rostral midbrain tegmentum (Wicht and Northcutt, '90). Thus it seems likely that the ventral olfactory tract and the cells that accompany it actually are a component of the secondary olfactory system rather than a homologue of the terminal nerve.
From our immunocytochemical description of the FMRF- amide-positive cell group in the infundibular nucleus of the hypothalamus (Wicht and Northcutt, '92b), it appears as if these cells also project to the olfactory bulb, since numerous FMRFamide-positive fibers which probably stem from this cell group were seen in the olfactory bulb. However, injections of tracers into the olfactory bulb failed to label these cells retrogradely. In part this discrepancy might be explained by the fact that the FMRFamide-positive cells show a very high degree of collateralization. Fibers positive for FMRFamide were seen in most parts of the forebrain, but perikarya were restricted to the infundibular hypotha- lamic nucleus and its immediate vicinity (Wicht and North-

540 H. WICHT AND R.G. NORTHCUTT
"/
Gnatho. Petro.
\
Myx .
Fig. 8. A cladogram showing the genealogical interrelationships of the three major Recent craniate taxa (bottom), and a diagrammatic representation of the extent of secondary olfactory projections to their telencephala (top). The cladogram at the bottom is in accordance with the most widely accepted interpretation of craniate interrelationships, based on a large number of morphological, physiological, and paleonto- logical characters (L@vtrup, '77; Janvier, '81; Jefferies, '86; Bardack, '91). This interpretation has recently been challenged based on a se- quence analysis of ribosomal RNA in various chordate taxa, and Stock and Whitt ('92) have proposed that lampreys and hagfishes actually are monophyletic. According to their sequence data, however, the ribo- somal RNA sequence of hagfishes has changed about 30% faster than the sequence of any other Recent chordate, including lampreys, and it therefore appears questionable whether this character is of any use in
cutt, '92b). It is therefore possible that application of tracers to only some of the collaterals, as in the case of olfactory bulb injections, is not sufficient to achieve label- ling of the perikarya.
A small number of retrogradely labelled cells and some labelled fibers were observed in the internal cellular layer of the olfactory bulb, contralateral to the injection site. These cells and fibers, as well as the bilaterality of the projections to the hemispheres, may be due to the presence of an anterior olfactory nucleus in hagfkhes. In mammals this nucleus, which is located in the transitional zone between the olfactory bulb and the telencephalon proper, shows strong reciprocal connections with the main olfactory bulb. In addition, it projects to a number of telencephalic targets that also receive an input from the olfactory bulb itself. However, unlike the olfactory bulb, whose projections are strictly ipsilateral, the anterior olfactory nucleus of mam- mals gives rise to projections that are not restricted to ipsilateral targets. Projections from the anterior olfactory
comparing hagfishes with other craniates. Furthermore, a cladistic analysis of Recent craniates in relation to recently discovered Silurian and Devonian fossil jawless fishes supports the notion that hagfishes actually are the sister group of all other craniates, as suggested by the above cladogram (see Forey and Janvier, '93, for a review). Telence- phalic cell groups receiving secondary olfactory projections are symbol- ized by hatched areas in the drawings. As a model for the gnathostome pattern (left) we choose an amphibian hemisphere. The pattern observed in lampreys is shown in the middle, that in hagfishes on the right. Note that all the diagrams are somewhat idealized, and not all of the cell groups depicted are actually at the same transverse level. This is particularly true for the position of the septum in lampreys and hag- fishes. See text for details.
nucleus also reach contralateral structures, such as the contralateral lateral pallium, olfactory bulb, and anterior olfactory nucleus. Furthermore, the projections from the anterior olfactory nucleus to the hemispheres are more widespread than the projections from the olfactory bulb (Switzer et al., '85). In principle, a similar situation might exist in hagfishes and other anamniotes that display bilat- eral higher order olfactory projections (see Northcutt and Puzdrowski, '88). The bilaterality might be the result of an uncrossed secondary olfactory projection arising from the olfactory bulb and a bilateral tertiary olfactory projection arising from an anterior olfactory nucleus, which may be hidden or dispersed among the cells of the internal layers of the olfactory bulb. Thus, assuming the presence of a hidden anterior olfactory nucleus, the pattern of olfactory projec- tions observed in anamniotes could be explained in terms of mammalian anatomy.
There may also be genuine differences in the organization of the higher order olfactory connections in mammals and

HAGFISH SECONDARY OLFACTORY PROJECTIONS 54 1
nonmammals, however. Thus Kemali and Guglielmotti (’871, working on the olfactory system of a frog, Rana esculenta, recognized an anterior olfactory nucleus (corre- sponding to the internal granular layer of the olfactory bulb of other authors) and compared its connections to the connections of the olfactory bulb. Unlike in mammals, both the olfactory bulb and the “anterior olfactory nucleus” displayed bilateral projections to the hemispheres, but neither projected to the contralateral bulb or contralateral “anterior olfactory nucleus.” Also in contrast to the situa- tion observed in mammals (see above), projections from the “anterior olfactory nucleus” were more restricted than those from the olfactory bulb.
Given the scarcity of available data and the obvious differences in mammalian and nonmammalian connectiv- ity, it is currently not possible to come to a meaningful conclusion regarding the presence or absence of an anterior olfactory nucleus in hagfishes. However, for the purpose of the present investigation, that is, determining of the rela- tive extent of olfactory projections onto the telencephalon, it seems reasonable to treat the entire system linking the olfactory bulb to the telencephalon as a unit. This unit is most appropriately termed the system of secondary olfac- tory projections, even though it is possible that some higher order olfactory projections might be included, particularly in anamniotes.
Phylogenetic considerations Several relevant aspects of the evolution of the secondary
olfactory system have already been discussed by Northcutt and Puzdrowski (’88). In comparing hagfishes, lampreys, and gnathostomes, it is evident that the following charac- ters are plesiomorphic for the secondary olfactory system of craniates: 1) bilaterality, 2) the presence of a medial tract innervating the septum, 3) the presence of a lateral tract innervating the pallium, and 4) the presence of a caudally directed tract (our ventral olfactory tract) innervating the basal diencephalon and reaching back to the mesodience- phalic boundary. We will limit or discussion to the relative extent of secondary olfactory projections and the tentative conclusions that can be drawn from this.
Although there is considerable variation in secondary olfactory projections, some features are common to all gnathostomes (Fig. 8): the lateral pallium and (at least in amphibians and birds-Scalia et al., ’68; Northcutt and Royce, ’75; Reiner and Karten, ’85) the lateral part of the dorsal pallium receive secondary olfactory afferents. With the exception of elasmobranchs (Smeets, ’831, the septum and the most rostral and ventral parts of the medial pallium also receive secondary olfactory input. In some species, particularly among amphibians (Northcutt and Kicliter, ’80), there also is a moderately strong input to the striatum.
In lampreys (Northcutt and Puzdrowski, ’88; Fig. 8) the pattern is similar, but there is, in addition, a strong projection to the entire dorsal pallium and a moderately strong projection to the medial pallium. In hagfishes there is a strong input to all pallid fields and the septum, and there is a moderate input to the striatum (Fig. 8).
Regardless of whether the extent of the secondary olfac- tory projections is measured by the percent of telencephalic area covered by these projections, or by the number of cell groups they reach, hagfishes have more extensive second- ary olfactory projections than lampreys, and lampreys have more extensive projections than gnathostomes. The cladis- tic relation of these three taxa (Fig. 8) thus indicates that
extensive secondary olfactory systems are a plesiomorphic character for craniate brains and, further, that extensive nonolfactory telencephalic centers arose with the origin of gnathostomes.
Studies on the nonolfactory connections of the pallia in hagfishes and lampreys are now needed in order to deter- mine how the extensive nonolfactory areas of the forebrain in gnathostomes evolved. If there is an ascending dience- phalic or other, presumably sensory, input to the pallium in hagfishes and lampreys, and if this input overlaps the secondary olfactory input, the nonolfactory centers of the telencephalon in gnathostomes may have evolved by a process of parcellation (Ebbesson, ’80). Alternatively, if there are no such projections and no such overlap in lampreys and hagfishes, it is likely that the nonolfactory centers in the telencephalon of gnathostomes evolved by a different mechanism, for example, the differentiation of new pallial fields, or the invasion of diencephalic projections with a concomitant retreat of the secondary olfactory fibers.
ACKNOWLEDGMENTS We appreciate the graphic skills of Inge Szasz, who
prepared the final versions of Figures 6 and 7. Mary Sue Northcutt assisted throughout all phases of the research and preparation of the manuscript. This work was sup- ported by a stipend and a travel grant from the Deutsche Forschungsgemeinschaft to H.W. (Wi 909/1-2 and Wi 909/2-1) and grants from the National Institutes of Health to R.G.N. (NS 24869 and NS 24669).
LITERATURE CITED Adams, J.C. (1981) Heavy metal intensification of DAB-based HRP reaction
product. J . Histochem . C ytochem .29: 7 75. Ariens Kappers, C.U., G.C. Huber, and E.C. Crosby (1936) The Comparative
Anatomy of the Nervous System of Vertebrates. Including Man. New York: Hafner (reprinted 1960).
Bardack, D. 11991) First fossil hagfish (Myxinoidea): A record from the Pennsylvanian of Illinois. Science 254~701-703.
Donald, J.A., and D.H. Evans (19921 Immunohistochemical localization of natriuretic peptides in the heart and brain of the gulf toadfish, Opsanus beta. Cell Tissue Res. 269:151-158.
Donald, J.A., A.J. Vomachka, and D.H. Evans (19921 Immunohistochemical localization of natriuretic peptides in the brains and hearts of the spiny dogfish Squalus acanthias and the Atlantic hagfish Myrine glutinosa. Cell Tissue Res. 270:535-545.
Ebbesson, S.O.E. 11980) The parcellation theory and its relation to interspe- cific variability in brain organization, evolutionary and ontogenetic development and neuronal plasticity. Cell Tissue Res. 213:179-212.
Edinger, L. (1904) Bau der nervosen Zentralorgane des Menschen und der Tiere. Leipzig: Vogel.
Eldredge, N., and J. Cracraft (1980) Phylogenetic Patterns and the Evolution- ary Process. New York Columbia University Press.
Forey, P., and P. Janvier I19931 Agnathans and the origin of jawed vertebrates. Nature 361t129-134.
Girgis, M. (1970) The rhinencephalon. Acta Anat. 76:157-199. Godement, P., J. Vanselow, S. Thanos, and F. Bonhoeffer (1987) A study in
developing visual systems with a new method of staining neurons and their processes in fixed tissue. Development 101:697-713.
Hanker, J.S., P.E. Yates, C.B. Metz, and A. Rustioni (1977) A new specific, sensitive and noncarcinogenic reagent for the demonstration of horserad- ish peroxidase. Histochem. J. 9:789-792.
Heimer. L. (1969) The secondary olfactory connections in mammals, reptiles and sharks. Ann. N.Y. Acad. Sci. 167:129-146.
Hennig, W. ( 1966) Phylogenetic Systematics. Urbana, University of Illinois Press.
Herrick, C.J. (1948) The Brain of the Tiger Salamander. Chicago: The University of Chicago Press.

542 H. WICHT AND R.G. NORTHCUTT
Holmgren, N. (1919) Zur Anatomie des Gehirns von Myxine. Kungliga Svenska Vetenskapsakademiens Handlingar 60t1-96.
Jansen, J. (1930) The brain of Myxine glutinosa. J. Comp. Neurol. 49.359- 507.
Janvier, P. (1981) The phylogeny of craniata, with particular reference to fossil “agnathans.” J. Vert. Paleontol. lt121-159.
Jefferies, R.P.S. (1986) The Ancestry of Vertebrates. Cambridge: British Museum of Natural History & Press Syndicate University Cambridge.
Kemali, M., and V. Guglielmotti (1987) A horseradish peroxidase study of the olfactory system of the frog, R a m esculenta. J. Comp. Neurol. 263r400-417.
Kicliter, E. (1979) Some telencephalic connections in the frog, Runupipiens. J. Comp. Neurol. 185r75-86.
Kusunoki, T., T. Katoda, and R. Kishida (1981) Chemoarchitectonics of the forebrain of the hagfish, Eptatretus burgeri. J. Hirnforsch. 22.285-298.
Lflvtrup, S. (1977) The Phylogeny of Vertebrata. London: Wiley. Munz, H., B. Claas, W.E. Stumpf, and L. Jennes (1982) Centrifugal
innervation of the retina by luteinizing hormone releasing hormone (LHRH)-immunoreactive telencephalic neurons in teleostean fishes. Cell Tissue Res. 222.313-323.
Neary, T.J. (1984) Anterior thalamic nucleus projections to dorsal pallium in ranid frogs. Neurosci. Lett. 51t213-218.
Neary, T.J. (1990) The pallium of anuran amphibians. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. BAD, Comparative Structure and Evolution of the Cerebral Cortex. New York: Plenum Press, pp. 107-138.
Nieuwenhuys, R. (1967) Comparative anatomy of olfactory centers and tracts. Prog. Brain Res. 23:l-64.
Nieuwenhuys, R., and J. Meek (1990a) The telencephalon of actinopterygian fishes. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. 8A/I, Comparative Structure and Evolution of the Cerebral Cortex. New York Plenum Press, pp. 31-73.
Nieuwenhuys, R., and J. Meek (1990b) The telencephalon of sarcopterygian fishes. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. 8AI1, Comparative Structure and Evolution of the Cerebral Cortex. New York: Plenum Press, pp. 75-106.
Northcutt, R.G. (1981) Evolution of the telencephalon in nonmammals. Ann. Rev. Neurosci. 4:301-350.
Northcutt, R.G. (1984) Evolution of the vertebrate central nervous system: Patterns and processes. Am Zool. 24t701-716.
Northcutt, R.G. (1985) Efferent projections of the olfactory bulb in the Pacific hagfish. Am. Zool. 25:106A (abstract).
Northcutt, R.G., and A.B. Butler (1991) Retinofugal and retinopetal projec- tions in the Green Sunfish, Lepomis cyanellus. Brain Behav. Evol. 37.333-354.
Northcutt, R.G., and E. Kicliter (1980) Organization of the amphibian telencephalon. In S.O.E. Ebbesson (ed): Comparative Neurology of the Telencephalon. New York: Plenum Press, pp. 203-255.
Northcutt, R.G., and R.L. Puzdrowski (1988) Projections of the olfactory bulb and nervus terminalis in the Silver lamprey. Brain Behav. Evol. 3296-107.
Northcutt, R.G., and M. Ronan (1992) Merent and efferent connections of the bullfrog medial pallium. Brain Behav. Evol. 4O:l-16.
Northcutt, R.G., and G.J. Royce (1975) Olfactory bulb projections in the bullfrog, Rana catesbeiana. J. Morphol. 145251-268.
Nozaki, M. (1985) Tissue distribution of hormonal peptides in primitive fishes. In R.E. Foreman, A. Gorbman, J.M. Dodd, and R. Olson (eds): Evolutionary Biology of Primitive Fishes. New York: Plenum Press, pp. 433-454.
Reiner, A,, and H.J. Karten (1985) Comparison of olfactory bulb projections in pigeons and turtles. Brain Behav. Evol. 27:ll-27.
Scalia, F., and D.R. Colman (1975) Identification of telencephalic afferent thalamic nuclei associated with the visual system of the frog. Neurosci. Abstr. lr46 (abstract).
Scalia, F., M. Halpern, H. Knapp, and W. Riss (1968) The efferent connex- ions of the olfactory bulb in the frog: A study of degenerating unmyelin- ated fibres. J. Anat. 103.245-262.
Smeets, W.J.A.J. (1983) The secondary olfactory connections in two chon- drichthyans, the shark Scyliorhinus canicula and the ray Raja clauata. J. Comp. Neurol. 218t334-344.
Smeets, W.J.A.J. (1990) The telencephalon of cartilaginous fishes. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. 8AI1, Comparative Structure and Evolution of the Cerebral Cortex. New York: Plenum Press, pp. 3-30.
Stell, W.K., S.E. Walker, K.S. Chohan, and A.K. Ball (1984) The goldfish nervus terminalis: An LHRH- and FMRF-amide-immunoreactive olfacto- retinal pathway. Proc. Natl. Acad. Sci. USA 81.940-944.
Stock, D.W., and G.S. Whitt (1992) Evidence from 18s ribosomal RNA sequences that lampreys and hagfishes form a natural group. Science 257:787-789.
Switzer, R.C., J. de Olmos, and L. Heimer (1985) Olfactory system. In G. Paxinos (ed): The Rat Nervous System. Orlando: Academic Press, pp, 1-36.
Ulinski, P.S. (1990) The cerebral cortex of reptiles. In E.G. Jones and A. Peters (eds): Cerebral Cortex, Vol. SAII, Comparative Structure and Evolution of the Cerebral Cortex. New York: Plenum Press, pp. 139-215.
von Bartheld, C.S., H.W. Lindorfer, and D.L. Meyer (1987) The nervus terminalis also exists in cyclostomes and birds. Cell Tissue Res. 250t431- 434.
Wicht, H., and W. Himstedt (1988) Topologic and connectional analysis of the dorsal thalamus of Triturus alpestris (Amphibia, Urodela, Salaman- dridae). J. Comp. Neurol. 267:545-561.
Wicht, H., and R.G. Northcutt (1990) Retinofugal and retinopetal projec- tions in the Pacific hagfish, Eptutretus stouti. Brain Behav. Evol. 36.315-328,
Wicht, H., andR.G. Northcutt (1992a) The forebrain of the Pacific hagfish: A cladistic reconstruction of the ancestral craniate forebrain. Brain Behav. Evol. 40.2544.
Wicht, H., and R.G. Northcutt (1992b) FMRFamide-like immunoreactivity in the brain of the Pacific hagfish, Eptatretus stouti. Cell Tissue Res. 270.443-449,
Wiley, E.O. (1981) Phylogenetics. New York: Wiley.