scientific annual report 2009 annual report... · scientific annual report 2013 . cerm centro di...
TRANSCRIPT

Scientific Annual Report 2013
CERM Centro di Risonanze
Magnetiche Università di
Firenze
C.I.R.M.M.P. Consorzio
Interuniversitario Risonanze
Magnetiche di MetalloProteine

0

Table of Contents Table of Contents................................................................................................................................... 1 Financial Support ................................................................................................................................... 2 Acknowledgements ............................................................................................................................... 3 Introduction ........................................................................................................................................... 4
Who we are ........................................................................................................................................ 4 Highlights of Research at CERM ............................................................................................................ 5
What CERM does ................................................................................................................................ 5 Where We Are ....................................................................................................................................... 6 CERM - CIRMMP Organization .............................................................................................................. 7
Staff .................................................................................................................................................... 7 Research Areas ...................................................................................................................................... 9
Structural Biology ............................................................................................................................... 9 In-cell NMR and Systems Biology .................................................................................................... 14 Metabolomics by NMR .................................................................................................................... 15 NMR Methodological Advancements .............................................................................................. 17 Bioinformatics and Databases ......................................................................................................... 20 Structure-based Vaccine and Drug Design ....................................................................................... 21 Fast Field Cycling Relaxometry ........................................................................................................ 23
Research with spin-off Institutions ..................................................................................................... 24 FiorGen Onlus ................................................................................................................................... 24 Giotto Biotech Srl ............................................................................................................................. 24
Instrumentation................................................................................................................................... 26 Solution and Solid State NMR spectrometers .................................................................................. 26 Fast Field Cycling Relaxometry ......................................................................................................... 29 X-ray Crystallography ....................................................................................................................... 30 Biological and Biophysical Equipment and Facilities ........................................................................ 30
EPR ......................................................................................................................................... 30 Multi Angle/Dynamic Light Scattering .................................................................................... 31 Surface Plasmon Resonance .................................................................................................... 31 Isothermal Calorimetry (ITC) ................................................................................................... 31 Circular Dichroism ................................................................................................................... 31 Fluorimetry .............................................................................................................................. 32 Stopped-flow ........................................................................................................................... 32
Biobank ............................................................................................................................................ 33 Bioinformatics and computational structural biology tools ............................................................. 33
Grid ......................................................................................................................................... 33 NMR Infrastructure ............................................................................................................................. 35
European NMR Infrastructure .......................................................................................................... 35 NMR National Access ....................................................................................................................... 36 International Doctorate in Structural Biology ................................................................................... 37
List of 2013 Publications ............................................................................................................. 38 Books and book chapters…………………………………………………………………............................................41 Highlights and Acknowledgements of CERM Research………………….……………………………………………..42 Meetings Organized by CERM ..................................................................................................... 42 Seminars and Research Seminars (RSS) Held at CERM .................................................................. 42
1

Financial Support
Tuscany Regional Government
European Commission
2

Acknowledgements
We thank the following institutions for stimulating interactions: Bracco SpA
Bruker BioSpin Srl
Fiorgen Onlus
Giotto Biotech Srl
Menarini Srl
Novartis Vaccines
and Diagnostics Srl
Siena Biotech SpA
Gruppo Sapio Srl
3

W H O
W E
A R E
Introduction CERM, Centre for Magnetic Resonance, is a scientific institution for research, transfer and higher education at the University of Florence. It is also the national laboratory of the Interuniversity Consortium for Magnetic Resonance of MetalloProteins (CIRMMP) which embraces three Italian Universities: Florence, Siena and Bologna. CERM is an infrastructure for Life Sciences with a particular focus on structural biology and specializations in bioinformatics, molecular biology, structural biology, novel drug design and development, and metabolomics.
Being a leading laboratory at both national and international level, CERM receives project funding from the Tuscan Regional Government, the Italian Ministry of Higher Education and Research (MIUR) and the
European Commission. Since 1994 CERM has been recognized as a transnational research infrastructure for its expertise and state-of-the-art instrumentation for NMR in Life Sciences.
The core technology at CERM is NMR spectroscopy, and its onsite instrumentation is among the most advanced in the world. A European transnational access service, financed by the EC since 1994 in addition to a national service operating since 1990, places CERM at the top of the list in the NMR field. Its two-fold status as a public-private laboratory fosters interactions between private industry and public research institutions such as Universities, CNR Institutes and European counterparts, promoting synergistic activities such as collaborations and services to SMEs.
CERM is involved in the ESFRI (European Strategy Forum on Research Infrastructures) and in the Italian Infrastructure roadmaps, and is one of the Core Centres of the European Integrated Structural Biology Infrastructure (INSTRUCT). CERM is also an e-infrastructure, managing a GRID-based platform, together with other laboratories in Europe, for biomolecular structure calculations, funded by the European
Commission (WeNMR http://www.wenmr.eu). CERM also promoted the creation of a “biobank of biological samples and biomolecular resources”, the da Vinci European Biobank (https://www.davincieuropeanbiobank.org) and of a center for research technology transfer, the CERM-TT, funded by the Tuscany Region.
4

W H A T
C E R M
D OE S
Highlights of Research at CERM The research activity of CERM can be summarized in the scheme below. During 2013 a number of projects have been developed, either as an extension of the activities of previous years or as new projects. Most of these projects receive specific funding from national and/or European organizations. Investments and costs in 2013 amounted to € 3.660.000,00: € 540.000,00 towards training and education, € 2.200.000,00 for new equipment and € 620.000,00 towards research activities. An additional € 300.000,00 covered operational costs. The actual replacement
value of the instrumentation at CERM is close to € 40.000.000,00. Our 2013 student body included 14 PhD students, 16 postdoctoral scientists and 10 undergraduate students. CERM has provided instrumentation access to several research groups. The interdisciplinary character of CERM research projects combined with the excellence of its instrumentation constitutes a point of reference for the scientific community and for cultural growth in the country, as demonstrated by the increased usage of the infrastructure by national scientists.
5

W H E R E
W E
A R E
Where We Are
CERM is located in the Scientific Campus (“Polo Scientifico”) of the University of Florence in Sesto Fiorentino, an area just west of the city of Florence. The main building has over 3000 square meters of usable space and contiguous laboratories measure more than 500 square meters. The campus borders
Florence International Airport and is a mere 15 minutes from the world-renowned centre of renaissance art and culture, Firenze.
6

ORGANIZATION
CERM-CIRMMP Organization
SCIENTIFIC BOARD OF CERM Florence Prof. Lucia Banci (Director) Dr. Marco Fragai Prof. Claudio Luchinat Prof. Roberta Pierattelli Prof. Antonio Rosato Prof. Paola Turano
CIRMMP BOARD OF DIRECTORS Florence Prof. Claudio Luchinat (President) Prof. Lucia Banci Bologna Prof. Stefano Ciurli Prof. Francesco Capozzi Siena Prof. Elena Gaggelli Prof. Stefano Mangani CIRMMP SCIENTIFIC BOARD Florence Prof. Lucia Banci Prof. Claudio Luchinat Siena Prof. Gianni Valensin Utrecht Prof. Rolf Boelens Frankfurt Prof. Harald Schwalbe
FACULTY MEMBERS University of Florence Dr. Claudia Andreini Prof. Lucia Banci Dr. Vito Calderone Dr. Francesca Cantini Prof. Simone Ciofi-Baffoni Prof. Isabella Felli Dr. Marco Fragai Prof. Claudio Luchinat Prof. Giacomo Parigi Prof. Mario Piccioli Prof. Roberta Pierattelli Prof. Antonio Rosato Prof. Paola Turano
ASSOCIATE FACULTY MEMBERS
University of Florence Prof. Cristina Nativi
University of Siena Prof. Stefano Mangani Prof. Elena Gaggelli Prof. Gianni Valensin
University of Bologna Prof. Stefano Ciurli Dr. Barbara Zambelli Prof. Francesco Capozzi Dr. Elena Babini
7

ORGANIZATION
POST DOCTORAL FELLOWS Caterina Bernacchioni Gabriele Cavallaro Chiara Cefaro ** Talita Duarte Pagani Karolina Gajda Angelo Gallo Ewa Gralka ** Vasantha Kumar Daniela Lalli Enrico Luchinat Tommaso Martelli Riccardo Peruzzini Enrico Ravera Jeffrey T. Rubino Maria Rosaria Saviello Julia Winkelmann
PHD STUDENTS Letizia Barbieri Eduardo Calçada Lucio Ferella Carmelo Fuccio * Gianluca Gallo Tomas Hosek Magdalena Korsak * Serena Lorenzini Marcela Oliveira Nogueira Alessandro Piai Mauro Rinaldelli Claudio Santucci * Erica Secci Yana Valasatava
UNDERGRADUATE STUDENTS Witold Andralojc Lorenzo Baronti Azzurra Carlon Silvia Ciambellotti Veronica Ghini Alexandra Louka * Riccardo Muzzioli Michele Salvi Veronica Nasta
VISITING STUDENTS AND POSTODOCS Diego Brancaccio
FIORGEN SCIENTISTS Leonardo Tenori Xiaoyu Hu
GIOTTO SCIENTISTS Valentina Borsi Linda Cerofolini Tatiana Kozyreva Sara Neri Tobias Schubeis
SCIENTIFIC MANAGER Francesca Morelli
TECHNICIANS Marco Allegrozzi Fabio Calogiuri Rebecca Del Conte Andrea Giachetti Leonardo Gonnelli Massimo Lucci Cristina Mescalchin Enrico Morelli
ADMINISTRATIVE SUPPORT Sara Damasceni Francesca Di Gloria Simona Fedi Milena Moazzi Laura Norfini Lisa Orlando
* Giotto Biotech Srl ** Fiorgen Onlus
8

Research Areas
Structural Biology Structural biology seeks to provide a complete and coherent picture of biological phenomena at the molecular and atomic level. The goals of structural biology include developing a comprehensive understanding of the molecular shapes and forms embraced by biological macromolecules and extending this knowledge to understand how different molecular architectures are used to perform the chemical reactions that are central to life at the cellular level. In addition, structural biologists are interested in understanding related processes such as protein folding, protein dynamics, molecular modeling, drug design, and computational biology. Central tools used in this research include X-ray diffraction, NMR, electron microscopy, other spectroscopies and biophysical methods, protein expression, biophysical and bio-organic chemistry, computer science and bioengineering. In particular, the synergy between NMR, X-ray crystallography, small angle X-ray scattering (SAXS), X-ray absorption spectroscopy (XAS), mass spectrometry, fast field cycling relaxometry and bioinformatics enabled at CERM to visualize protein structures at the atomic level thus enhancing the understanding of protein functions from several different and complementary perspectives. Specifically it is possible to study how proteins interact with other molecules, how they undergo conformational changes, and how enzymes perform catalysis. A combination of the above mentioned techniques is used routinely to understand the role played by a protein in the frame of cellular metabolism, or to rationally engineer an enzyme for a specific industrial process, or to determine how to design novel drugs that target a particular protein, or to understand what changes might improve them. All the information obtained at the atomic and molecular level can then be assembled to yield a more comprehensive and dynamical view of cellular processes. This is an approach at the systems biology level, which allows to understand how function is modulated by
interactions among biomolecules, thus addressing the missing links between molecules and cellular physiology. CERM has adopted an innovative strategy of integrated systems biology approach using several structural tools for addressing more and more challenging questions. CERM is currently applying this approach to different classes of metalloproteins involved in iron-sulfur protein biogenesis and maturation of metalloenzymes. Each organism possesses mechanisms that allow the uptake of the quantity of metal necessary for the cell and insertion of the metal in the final protein targets. We have characterized the structure, dynamics and protein-protein interactions of some of these systems in eukaryotes. In particular, an intense research activity was devoted to investigate molecular aspects of iron sulfur protein biogenesis processes. Biogenesis of iron–sulfur cluster proteins is a highly regulated process that requires complex protein machineries. In most iron-sulfur final target proteins, the cluster(s) acts as electron-transfer group in mediating one-electron redox processes. It is now evident that several rare and apparently dissimilar human diseases are attributable to defects in the basic process of Fe-S cluster biogenesis. Although these diseases (which include Friedreich’s ataxia, ISCU myopathy, a rare form of sideroblastic anemia, an encephalomyopathy caused by dysfunction of respiratory chain complex I and multiple mitochondrial dysfunctions syndrome) affect different tissues, a feature common to many of them is that mitochondrial iron overload develops as a secondary consequence of a defective Fe-S cluster biogenesis. Therefore the understanding of the iron-sulfur cluster biogenesis is of key relevance. The major achievements obtained in 2013 adopting an integrate approach involved the investigation of the electron transfer process which is responsible for assembling cytosolic iron-sulfur cluster proteins. In humans this process involves two proteins, i.e. anamorsin, an iron sulfur protein, and Ndor1, an NADPH-
RESEARCH
AREAS
9

dependent diflavin oxidoreductase. These two human proteins form a stable complex in vivo that was proposed to provide electrons for assembling cytosolic iron–sulfur cluster proteins. The Ndor1–anamorsin interaction was also suggested to be implicated in the regulation of cell survival/death mechanisms. In our study we unraveled the molecular basis of recognition between the FMN-binding domain of Ndor1 and anamorsin as well as the electron transfer process between them. This is based on a combined NMR and X-ray structural characterization of Ndor1 and anamorsin and on the identification of those protein regions involved in complex formation and those involved in electron transfer. We found that an unstructured region of anamorsin is essential for the formation of a specific and stable protein complex with Ndor1, whereas the C-terminal region of anamorsin (CIAPIN1 domain), containing [2Fe-2S] redox centers, transiently interacts through complementary
charged residues with the FMN-binding site region of Ndor1 to perform electron transfer. The molecular model of the proposed electron transfer process (Figure 1) provides significant information on the functional processes in which Ndor1-anamorsin interaction is implicated, i.e., the assembly of iron-sulfur clusters, of diferric proteins and the regulation of cell survival/death mechanisms. Indeed, the disruption of the stable interaction between anamorsin and Ndor1 might provoke the interruption of the electron flow between the two proteins within the cell and, as a result of that, its essential function for cellular survival is abolished and consequently cell death mechanisms might be activated (Banci L, Bertini I, Calderone V, Ciofi-Baffoni S, Giachetti A, Jaiswal D, Mikolajczyk M, Piccioli M, Winkelmann J. (2013) Molecular view of an electron transfer process essential for iron-sulfur protein biogenesis. Proc Natl Acad Sci U S A 110(18):7136-7141). Figure 1. Model of the electron transfer process between Ndor1 and anamorsin. Anamorsin (in pink) can be tightly bound to both closed and open conformational states of Ndor1 (in gray) due to the specific recognition between an unstructured region of anamorsin and a region of the FMN-binding domain that is solvent exposed in both open and closed conformations (in yellow). The N-terminal domain of anamorsin is not involved in the protein-protein recognition process. Upon NADPH (in blue) binding, electrons can efficiently be transferred in the closed conformation of Ndor1 to FAD (in red) and then to FMN (in green) to produce the hydroquinone state of FMN. Interflavin electron transfer in Ndor1 significantly populates the open conformation, which exposes to the solvent the FMN moiety and allows the formation of a transient interaction with the [2Fe-2S] cluster region of anamorsin located in the CIAPIN1 domain. The latter interaction allows an efficient transfer of one electron from the hydroquinone state of FMN to the [2Fe-2S] cluster (in black).
RESEARCH
AREAS 10

An integrated approach exploiting spectroscopic techniques (paraNMR, EPR, Mössbauer) was developed to investigate [2Fe-2S] binding properties and the electronic properties of anamorsin (Figure 2). It showed that i) each cysteine-rich motifs (CX8CX2CXC and CX2CX7CX2C) in anamorsin is able to bind independently a [2Fe-2S] cluster through its four cysteine residues, the binding of one cluster mutually excluding the binding of the second; ii) the [2Fe-2S] centers have unique electronic properties which can be interpreted by considering that both cysteine-rich motifs are located in a highly unstructured and flexible protein region, whose local conformational heterogeneity can induce anisotropy in metal coordination. This study contributes to the understanding of the functional role of the CIAPIN1 domain in the anamorsin family, suggesting that only the [2Fe-2S] cluster bound to the CX8CX2CXC motif is indispensable in the electron transfer chain assembling cytosolic Fe/S proteins
(Banci L, Ciofi-Baffoni S, Mikolajczyk M, Winkelmann J, Bill E, Pandelia ME (2013) Human anamorsin binds [2Fe-2S] clusters with unique electronic properties. J Biol Inorg Chem. 18(8):883-93)
Figure 2. EPR and Mössbauer spectra of [2Fe-2S]-anamorsin.
In another study we investigated the superoxide dismutase 1 (SOD1) maturation. This is mainly accomplished with the SOD1-specific chaperone, CCS, a dimeric protein with three distinct domains in each monomer. We showed that the first domain of human CCS (hCCSD1) is responsible for copper transfer to its protein partner, human SOD1 (hSOD1). The NMR solution structure (Figure 3) of the copper(I)-loaded form of hCCSD1 contributed to characterize the copper-transfer mechanism to hSOD1. NMR spectroscopy also allow to examine the hSOD1 mutants C57A, C146A, and C57A/C146A, which are unable to form the structurally conserved disulfide bond in SOD1, in order to investigate the role of these cysteines during hSOD1 copper acquisition. Together, the information on both hCCS and hSOD1, along with a sequence analysis of eukaryotic CCSD1, allows us to propose important mechanistic aspects regarding the copper-transfer process from hCCS to hSOD1 (Banci L, Cantini F, Kozyreva T, Rubino JT (2013) Mechanistic aspects of hSOD1 maturation from the solution structure of Cu(I) -loaded hCCS domain 1 and analysis of disulfide-free hSOD1 mutants. Chembiochem. 14(14):1839-44).
Figure 3. A) Overlay of 20 conformers of the CuI-hCCSD1 structure (PDB ID: 2RSQ). The side chains of the CuI-binding cysteines and Arg71 are shown. The CuI atom for each conformer is shown as a sphere. The unstructured N- and C-terminal regions are omitted for clarity. B) Superimposition of the mean structures of CuI-hCCSD1, CuI-Hah1 (PDB ID: 1L4), and CuI-Atx1 (PDB ID: 1FD8). The side chains of Cys22, Cys25, and Arg71 (CuI-hCCSD1 numbering) are shown.
The functional role of unstructured protein domains is an emerging field in the frame of intrinsically disordered proteins. The involvement of intrinsically disordered domains (IDDs) in protein targeting and biogenesis processes in mitochondria is so far not known. The structural/dynamic and functional properties of an IDD of the sulfhydryl oxidase
RESEARCH
AREAS
11

ALR (augmenter of liver regeneration) located in the intermembrane space of mitochondria showed that this unstructured domain performs a dual function in two cellular compartments: it acts (i) as a mitochondrial targeting signal in the cytosol and (ii) as a crucial recognition site in the disulfide relay system of intermembrane space (Figure 4). This domain provides an exciting new paradigm for IDDs ensuring two distinct functions that are linked to intracellular organelle targeting (Banci L, Bertini I, Cefaro C, Ciofi-Baffoni S, Gajda K, Felli IC, Gallo A, Pavelkova A, Kallergi E, Andreadaki M, Katrakili N, Pozidis C, Tokatlidis K. (2013) An intrinsically disordered domain has a dual function coupled to compartment-dependent redox control. J Mol Biol. 425(3):594-608).
Figure 4. The functional role of the unstructured N-terminal domain of ALR in ALR protein import and electron transfer process with Mia40.
The characterization of the functional states of proteins in solution is another field of research at CERM. In this respect, the interdomain flexibility of MMP-1 in the absence of collagen peptides is being characterized in more detail in order to map the most accessed conformations within the ensemble of all sterically possible conformations in solution. Paramagnetic ions (Tb3+, Dy3+, Tm3+) were placed in the CAT domain of full-length MMP-1 using the CLaNP-5 tag. PCSs, RDCs and SAXS are thus used to rank a number of randomly generated conformations according to their MO value. The MO of a given conformation is defined, and numerically calculated, as the maximum weight that this conformation can have in any suitable ensemble while still maintaining the ensemble’s ability to reproduce the experimental data. Ranking the conformations according to their MO values thus reflects their possible sampling. The conformations with the highest MO are clustered in a well defined region, and correspond to relatively elongated structures.
To select the MMP-1 conformations that may be more relevant in solution for collagen binding, the structures with the highest MO values have been compared after superimposing their HPX domains. These results (Figure 5) are in nice agreement with a previous analysis performed using relaxation and SAXS data, indicating that the compact arrangement of the two domains observed in the crystal structure cannot be fully representative of the conformations sampled by the protein in solution, where it must exist in a more extended arrangement for at least one third of the time. This suggests the existence of a relatively well defined, high MO, reciprocal orientation of the two domains in solution that is not distant from the MMP-1 structure reported in the first step of the collagenolytic mechanism (Cerofolini L, Fields GB, Fragai M, Geraldes CF, Luchinat C, Parigi G, Ravera E, Svergun DI, Teixeira JM. Examination of matrix metalloproteinase-1 in solution: a preference for the pre-collagenolysis state. J. Biol. Chem. 2013, 288, 30659-30671).
RESEARCH
AREAS 12

Figure 5. Interaction of FL-MMP-1 with the substrate. In the panel, from the top to the bottom: i) structure with the highest MO, two morphing intermediate steps, the experimentally-driven docked complex where the hemopexin-like domain and the catalytic domain bind the triple-helical collagen. The structure with the highest MO e morphing structures were aligned to the hemopexin-like domain of the docked complex. FL-MMP-1 and THP are represented as white and yellow surfaces, respectively. In blue is the MMP consensus sequence HEXXHXXGXXH and the cleavage site (Gly-Ile) in the first chain of THP. The catalytic zinc ion is represented as an orange sphere. To facilitate visualizing the movement of the catalytic domain with respect to the hemopexin-like domain, the blue and the red arrows indicates the direction of helices hA and hC of the catalytic domain defined by residues 130-141 and 250-258, respectively.
In a research activity aimed to reveal new DNA structural motifs provides the molecular bases to elucidate novel functional mechanisms of cell and the way to interact with them. By combining advanced state-of-the-art computations and experiments, we have identified a new DNA structural motif, named “G-triplex”. G-triplex (Figure 6) can be formed in guanine-rich regions of the genome and is characterized by the formation of G:G:G triad planes stabilized by an array of Hoogsteen-like hydrogen-bonds. This discovery further expands the structural complexity of the
genome highlighting once more the high polymorphism of the DNA polymers. This is the first time that DNA is found to assume this kind of topology and the abundant presence of guanine-rich regions in the genome makes imperative to investigate its biological role in the near future (Limongelli V, De Tito S, Cerofolini L, Fragai M, Pagano B, Trotta R, Cosconati S, Marinelli L, Novellino E, Bertini I, Randazzo A, Luchinat C, Parrinello M. The G-triplex DNA. Angew Chem Int Ed Engl. 2013, 52, 2269-2273).
.
Figure 6. The thrombin binding aptamer (TBA) has been shown to adopt a stable G-triplex structural motif, in addition to the usual G-quadruplex.
RESEARCH
AREAS 13

In-cell NMR and systems biology At CERM, in-cell NMR is being further developed and applied to study protein folding and maturation events. The technique was previously established in E. coli cells, and was then extended to human cells. NMR on human cells allows studying human proteins in their real environment, together with their specific partners. The gene of interest is transiently transfected into cultured human cells, using a mammalian vector optimized for high expression levels. With this technique, two or more proteins – such as specific chaperones of the protein of interest – can be simultaneously overexpressed and isotopically labelled. With this approach, the complete sequence of maturation events leading to mature, active copper,zinc-superoxide dismutase 1 (Cu,Zn-SOD1) enzyme was followed in the cytoplasm of human cells. Intracellular zinc binding, homodimer formation and copper uptake events were followed, and copper chaperone for SOD1 (CCS) oxidation of the SOD1 intrasubunit disulfide bond was found to occur through both copper-dependent and independent mechanisms (Banci L, Barbieri L, Bertini I, Luchinat E, Secci E, Zhao Y, Aricescu AR. (2013) Atomic-resolution monitoring of protein maturation in live human cells by NMR. Nat Chem Biol 9(5):297-9). Several mutants of the SOD1 gene are known to cause the onset of the familial variant of amyotrophic lateral sclerosis (fALS), a neurodegenerative disease. With the in-human-cell NMR approach, several SOD1 mutants associated with the fALS disease are currently investigated. For a set of these mutants, the zinc binding step of SOD1
maturation is found to be impaired in the cell. For the first time, an intracellular unstructured species is observed by NMR, which likely is a precursor of toxic aggregation products of fALS SOD1 mutants. Coexpression of the same SOD1 mutants with CCS restores the zinc binding step of SOD1 maturation, indicating that CCS acts as a molecular chaperone, and is able to prevent the intracellular formation of the unstructured species. The folding of human Mia40, an oxidoreductase of the mitochondrial inter-membrane space (IMS), was characterized in the cytoplasm, prior to mitochondrial import. The oxidation state of cytoplasmic Mia40 was found to be dependent on glutaredoxin 1 (Grx1) and, to a lesser extent, on thioredoxin 1 (Trx1), thiol-disulfide regulating proteins of the cytoplasm. Grx1 and Trx1 are therefore part of the cell regulation mechanism which keeps newly synthesized Mia40 in the unfolded, import-competent state (Banci L, Barbieri L, Luchinat E, Secci E. (2013) Visualization of redox-controlled protein fold in living cells. Chem Biol 20(6):747-52). Several proteins, like Mia40, exert their biological function in specific cellular compartments, such as the IMS of mitochondria. A novel in-mitochondria NMR approach is currently being developed at CERM, which allows structural information to be obtained on proteins inside intact mitochondria isolated from human cells. Such technique can in principle be extended to several other organelles, being the first of a series of “in organello NMR” approaches for structural systems biology.
The folding state of the human mitochondrial protein Mia40 in the cytoplasm, prior to mitochondrial import, was monitored by in-cell NMR. Mia40 accumulates in the cytoplasm in the folded, oxidized state. Cytoplasmic Grx1 and Trx1 can regulate Mia40 redox state, and are able to keep most Mia40 in the unfolded, import-competent state.
RESEARCH
AREAS
14

Metabolomics by NMR Metabolomics is a further “omic” science that is now emerging with the purpose of elaborating a comprehensive analysis of the metabolome, which is the complete set of metabolites (i.e. small molecules intermediates) in an organism, tissue, cell or biofluid. While genomics tells you what could happen, metabolomics tells you
what is really happening. There are close to hundred thousand proteins encoded in the genome, but only a few thousand metabolites. The projects involve varied areas of interest, in particular the biomedical field. The primary current metabolomic projects, are:
• Gluten sensitivity
• Broncho-pulmonary obstructive diseases and lung cancer using exhaled breath condensate
• Study on metabolic fingerprint of heart failure
• New statistical methods for data analysis and clustering
• Prognostic and diagnostic studies on patients with metastatic colorectal cancer
• Prognostic and diagnostic studies on patients with pancreatic cancer
• Discovery of new biomarkers for diagnosis and prognosis of melanoma
• Metabolomics application to urogenital cancers (prostate, bladder, kidney)
• Metabolomics of intestinal ischemia
• Applications of metabolomics in nutritional studies (EU-funded projects Chance and Pathway-27, collaborations with the “Assessorato regionale all’agricoltura”)
• Metabolomics for the assessment of samples quality and impact of preanalytical procedures.
The study of biofluids is performed through nuclear magnetic resonance (NMR) spectroscopy. Data obtained from NMR spectra are analysed with advanced computational techniques such as supervised learning methods (Support Vector Machines, Neural Networks) and with standard techniques such as primary components analyses or multilinear regression methods. The goal of metabolomics is to integrate the individualization of specific metabolomic markers with the set of cellular products to explain the inner workings of the onset of a disease. During 2013 seven papers were published in the field. Four of them deals with applications of metabolomics to diseases fingerprinting: samples taken from infants of gestational diabetic mothers (Dani C, Bresci C, Berti E, Ottanelli S, Mello G, Mecacci F, Breschi R, Hu X, Tenori L, Luchinat C. (2013) Metabolomic
profile of term infants of gestational diabetic mothers J Matern Fetal Neonatal Med), from individuals with heart failure (Metabolomic fingerprint of heart failure in humans: A nuclear magnetic resonance spectroscopy analysis. Tenori L, Hu X, Pantaleo P, Alterini B, Castelli G, Olivotto I, Bertini I, Luchinat C, Gensini GF. (2013). Int J Cardiol, 168 (4), 113-115; Padeletti L, Modesti PA, Cartei S, Checchi L, Ricciardi G, Pieragnoli P, Alterini B, Padeletti M, Hu X, Tenori L, Luchinat C (2013). Metabolomic does not predict response to cardiac resynchronization therapy in patients with heart failure. J Cardiovascular Med,) and from patients affected by COPD (Bertini I, Luchinat C, Miniati M, Monti S, Tenori L. (2013) Phenotyping COPD by 1H NMR metabolomics of exhaled breath condensate. Metabolomics) were analyzed by NMR metabolomics. Metabolomics provided an useful tool for disease investigation giving new hint on the investigated diseases. For example, an unexpected and counterintuitive results was obtained studying heart failure:
RESEARCH
AREAS
15

although healthy controls and patients are extremely well discriminated by the metabolic profile, the disease itself cannot be stratified, i.e. the metabolic alterations are present even in patient with very low or moderate severity. Another paper consists of a review on the metabolic changes found in celiac patients, especially in the light of the know modification affecting the gut microbiota consortium (Calabro A, Gralka E, Luchinat C, Saccenti E, and Tenori L, Metabolomic perspective on coelic disease. Autoimmune disease). One is a methodological work devoted to establish the best pre-analytical procedures for the analysis
of bacterial samples (Bertini I, Hu X, Luchinat C 2013 Global metabolomics characterization of bacteria: pre-analytical treatments and profiling Metabolomics). Finally, in the last paper the effects of intra- and post-operative ischemia on the metabolic profile of clinical liver tissue was monitored by NMR metabolomics (Cacciatore S, Hu X, Viertler C, Kap M, Bernhardt GA, Mischinger HJ, Riegman P, Zatloukal K, Luchinat C, Turano P (2013) Effects of Intra- and Post-Operative Ischemia on the Metabolic Profile of Clinical Liver Tissue Specimens Monitored by NMR. J Proteome Res.).
OPLS score plots. The plot depicts the discriminatory power of metabolomics. The graphical separation of HF subjects (solid black dots) from healthy subjects (solid red squares) corresponds to the capability of the statistical model to discriminate the two populations, using 1D-NOESY spectra.
RESEARCH
AREAS 16

NMR Methodological Advancements The development of new NMR methods is essential for broadening the range of NMR spectroscopy applications to the study of a wider range of biological systems. Recently, focus has been placed on 13C direct detection NMR spectroscopy both in solution and the solid state. A suite of multidimensional NMR experiments based on 13C detection has been developed and offers an additional tool
for the study of biological macromolecules. It can allow spectral assignment and it can provide more, and in some cases unique, information on the conformational and dynamical properties particularly for those systems which cannot be studied by other NMR techniques such as disordered or unstructured proteins.
The most recent developments focus on the implementation of the so called “fast methods” for the study of these proteins directly in-cell. A new experiment, the HN-BESTCON (Gil S, Hošek T, Solyom Z, Kümmerle R, Brutscher B, Pierattelli R, Felli IC (2013). NMR Spectroscopic Studies of Intrinsically Disordered Proteins at Near-Physiological Conditions, Angew. Chemie, 52(45) 11808-11812) was designed, enabling to obtain a 2D map in a couple of minutes. Together with the other 13C detected CON variants it permits to study intrinsically disordered proteins approaching physiological conditions. The reduction in experimental time also enabled the design of a suite of multidimensional NMR experiments with more than three dimensions (4D, 5D, etc.) (Bermel W, Felli IC, Gonnelli L, Koźmiński W, Piai A, Pierattelli R, Zawadzka-
Kazimierczuk A. (2013) High-dimensionality 13C direct-detected NMR experiments for the automatic assignment of intrinsically disordered proteins J. Biomol. NMR, 57(4), 353-361). These experiments are particularly useful to study complex proteins characterized by significant signal overlap. Indeed, the increasing number of frequencies associated to each cross peak in a multidimensional spectrum alleviates the problem of extensive overlap, contributing to an increased resolution which allows us to focus on more complex systems. The figure shows how one can easily visualize the information content of multidimensional NMR experiments. As the experiments based on 13C detection in solution share some common aspects to experiments in the solid state, some of the innovations introduced in solution NMR were
RESEARCH
AREAS
17

exploited to optimize the performance of several solid state NMR experiments. The key development in solid state NMR pursued lately, however, resides in the possibility to use 1H NMR experiments, enabled by the use of high-MAS probeheads at high magnetic field. Solid-state NMR (SS-NMR) of proteins requires that those molecules be immobilized, usually by crystallization, freezing, or lyophilization. However, self-crowding can also slow molecular rotation sufficiently to prevent the nuclear interactions from averaging. To achieve self-crowding, ultracentrifugation can be used. We have shown that sedimented solute NMR (SedNMR) provides a simple method to prepare biological samples for SS-NMR experiments with minimal perturbation. SedNMR might be a suitable methodology to selectively entrap and observe transient complexes or oligomeric states that fail to crystallize and/or are too big to be observed by routine methods of solution state NMR (Bertini I, Luchinat C, Parigi G, Ravera E. (2013) SedNMR: on the edge between solution and solid-state NMR. Acc Chem Res. 46(9):2059-69) Ex situ sedimentation may provide spectra of higher intensity (and resolution) than crystalline samples, if larger protein concentrations or increased packing efficiency
are achieved in sediments than in crystals. We have also shown that a small protein of 32 kDa can provide highly resolved in situ sediment spectra (Fig.1). In the case of even smaller proteins, the molecular packing in the sediment might not be enough to completely prevent rotational diffusion. Cooling the sediment just below the freezing of bulk solvent may yield again well resolved spectra, suitable for spectroscopic investigation. (Fragai M, Luchinat C, Parigi G, Ravera E. (2013) Practical considerations over spectral quality in solid state NMR spectroscopy of soluble proteins J Biomol NMR. 57(2) 155-66). The sedimented state is also amenable for Dynamic Nuclear Polarization (Ravera E, Corzilius B, Michaelis VK, Rosa C, Griffin RG, Luchinat C, Bertini I. (2013) Dynamic nuclear polarization of sedimented solutes. J Am Chem Soc. 135(5):1641-1644). We have shown that sedimentation of the protein enables significant DNP enhancements without the addition of a glass-forming material such as glycerol.
Fig.1 DARR spectrum of hSOD sedimented in situ, recorded at 20.0 T, 14 kHz MAS, 290.8 K effective sample temperature
RESEARCH
AREAS 18

Summary of the approaches to achieve sedimentation for SS-NMR sample preparation: (left) in situ sedimentation; (right) ex situ sedimentation using ultracentrifugal devices.
The protein MMP-1 is composed of two rigid domains connected by a flexible linker. Its conformational heterogeneity has been investigated through NMR measurements based on the exploitation of pseudocontact shifts (pcs), self orientation residual dipolar couplings (rdc), which become available after addition of a paramagnetic probe to one protein domain, and SAXS data (Cerofolini L, Fields GB, Fragai M, Geraldes CF, Luchinat C, Parigi G, Ravera E, Svergun DI, Teixeira JM. (2013). Examination of matrix metalloproteinase-1 in solution: a preference for the pre-collagenolysis state J. Biol. Chem., 288, 30659). The approach is based on the calculation for the sterically allowed protein conformations of the maximum occurrence (MO) consistent with the collected data (Fragai M, Luchinat C, Parigi G, Ravera E (2013) Conformational freedom of metalloproteins revealed by paramagnetism-assisted NMR. Coord. Chem. Rev. 257, 2652). The MO of a given conformation is defined, and numerically calculated, as the maximum weight that this conformation can have in any suitable ensemble while still maintaining the ensemble’s ability to reproduce the experimental data. Ranking the conformations according to their MO values thus reflects their possible sampling. A strategy proposed for monitoring and possibly improving the accuracy of a protein structure in solution is that of taking a crystal
structure as a starting model and to validate or to refine it by applying few, highly informative, long range NMR restraints. The program REFMAC5 from CCP4 was modified to allow for the simultaneous use of X-ray crystallographic data and paramagnetic NMR data (pseudocontact shifts and self-orientation residual dipolar couplings) and/or diamagnetic residual dipolar couplings (Rinaldelli et al. Acta Crystallographica D, 2013 in press). Incorporation of these long range NMR restraints in REFMAC5 can reveal differences between solid state and solution conformations of molecules or, in their absence, be used together with X-ray crystallographic data for structural refinement. Since NMR and X-ray data are complementary, when a single structure is consistent with both sets of data and still maintains reasonably "ideal" geometries, the reliability of the derived atomic model is expected to increase. The program was tested on five different proteins: the catalytic domain of matrix metalloproteinase-1, GB3, ubiquitin, free calmodulin and calmodulin complexed with a peptide: in some cases the joint refinement produced a single model consistent with both sets of observations, in other cases it indicated, outside the experimental uncertainty, the presence of different protein conformations in solution and in the solid state.
RESEARCH
AREAS 19

Bioinformatics and Databases The bioinformatics research activity in 2013 at CERM was carried out along the lines followed in previous years, focusing on the development of methods and tools specifically targeted to the study of metalloproteins, and ultimately aimed at facilitating knowledge discovery processes and thereby advancing our understanding of the interaction among metal ions and biological macromolecules. In more detail, we worked in parallel on (i) the expansion and improvement of the MetalPDB database (http://metalweb.cerm.unifi.it/), and (ii) the creation of novel web resources for the analysis of metalloprotein structures. There exists a strong synergy between these two topics as the data contained in MetalPDB, which represents our central platform to collect information on metalloproteins, are used as the basis for the design of new analysis tools, whose results are, in turn, integrated into the database. This virtuous circle is made possible by the fact that all of our resources share the same unifying conceptual framework, provided by the description of metal sites in proteins and nucleic acids as Minimal Functional Sites (MFSs). MFSs are 3D templates that describe the local environment around the metal(s) independently of the larger context of the macromolecular structure. Such local environment has a determinant role in tuning the chemical reactivity of the metal, ultimately contributing to the functional properties of the whole system. As regards point (i), we worked on the process of functional annotation of the MFSs stored in MetalPDB, which involved (a) the creation of interfaces to assist human annotators, and (b) the execution of the complex and time-consuming annotation task. This process, which is still ongoing, is speeded up thanks to the grouping of MFSs into equivalent sites (broadly defined as sites found in corresponding positions in similar structures and containing the same metals), so that we expect it to be completed soon. The paper describing MetalPDB was published at the beginning of this year, in the 2013 database issue of Nucleic Acids Research (Andreini C, Cavallaro G, Lorenzini S, Rosato A. MetalPDB: a database of metal sites in biological macromolecular structures. Nucleic Acids Res. 2013 41(Database issue):D312-9). As regards point (ii), we first developed the MetalS2 (Metal Sites Superposition) tool for the structural alignment of MFSs in any pair of metal-binding
biological macromolecules. On our example data sets, MetalS2 unveiled structural similarities that other programs for protein structure comparison do not consistently point out, and overall identified a larger number of structurally similar MFSs. MetalS2 supports the comparison of MFSs harbouring different metals and/or with different nuclearity, and is available both as a stand-alone program and a web tool at http://metalweb.cerm.unifi.it/tools/metals2MetalS2 (Andreini C, Cavallaro G, Rosato A, Valasatava Y. A tool for the structural alignment of minimal functional sites in metal-binding proteins and nucleic acids. J Chem Inf Model. 2013 Nov 25;53(11):3064-75). With MetalS2 available, we were able to develop MetalS3 (Metal Sites Similarity Search), a database search tool to identify metal sites having structural similarity to a query metal site structure within the MetalPDB database. MetalS3 uses a suitably adapted version of the algorithm implemented in MetalS2, and can be accessed through a web interface at http://metalweb.cerm.unifi.it/tools/metals3/. It systematically compares the structure of the query metal site to each MFS in MetalPDB, and keeps the best superposition for each MFS. All these superpositions are then ranked according to the MetalS3 scoring function and presented to the user in tabular form. The user can interact with the output web page to visualize the structural alignment or the sequence alignment derived from it. Options to filter the results are also available. Test calculations showed that the MetalS3 output correlates well with expectations from protein homology considerations. A practical application of MetalS3 is to detect MFS structural similarities that are not associated with a homology relationship among the proteins containing the MFSs. These situations may be indicative of the occurrence of common functional properties that are endowed by the MFS itself. Such observations can provide useful hints for experimental work. The paper describing MetalS3 was accepted for publication in the Journal of Biological Inorganic Chemistry. We are now working mainly on MetalPDB annotation, as well as on further tools exploiting the concept of MFS.
RESEARCH
AREAS 20

Structure-based Vaccine and Drug Design CERM is a leading center in the development of an innovative approach in vaccine development. It is based on the knowledge of the antigen structure and of its interactions with antibodies. Researchers at CERM, in collaboration with Novartis Vaccines and Diagnostics, have applied this approach on a meningococcal antigen for engineering a chimeric molecule that elicits a broad protective immunity against Neisseria meningitides serogroup B. In particular in Neisseria meningitidis, the protective antigen factor H binding protein (fHBP) (recently discover by Novartis as a novel and powerful protective antigen able to induce antibodies that kill the bacteria) has more than 300 different sequences, classified in three distinct antigenic variants, which do not induce cross-protective immunity. In order to generate a single antigen able to induce immunity against all sequence variants, the solution structure of the variant one of fHBP has been firstly solved by CERM researchers. Subsequently, the surface of the carboxyl-terminal domain of variant one has been divided into overlapping areas of the size typical of conformational epitopes, and each area has been mutated to introduce residues specific to variants two and three. Thus, a lead chimeric antigen able to induce high level of cross-protective antibodies against all variant strains has been identified. This study has demonstrated that the structure-based design of multiple immunodominant antigenic surfaces on a single protein scaffold represents a novel way to create broadly protective vaccines and can be very successful once sequence variability of protective antigens is a major limit to the development of vaccines. The structure-based approach for generating more stable and efficient vaccines antigens is currently applied, in collaboration with Novartis, to the pathogen Clostridium difficile, with the final goal being that of obtaining a new candidate vaccine to use for the preliminary trials. The structural vaccinology strategy is therefore an effective general way to create broadly protective vaccines against several pathogens and it is particularly powerful in case of antigenic variation between closely related strains and species
Researchers at CERM are also involved in the development of new candidate drugs and biological probes. In particular a new bifunctional probe to target galectins for directing MMPI to specific sites has been designed and investigated. The formation of the ternary complex MMP-Probe-Lectin in situ can be used to exploit the lectin for attracting MMPI to the tumour and, as an added benefit, to block protumoural activities of lectin at the same time. The same strategy can be applied to other pathological states with concomitant upregulation of MMPs/galectins, such as rheumatoid arthritis, thus broadening the applicability of this concept (Bartoloni M, Domínguez BE, Dragoni E, Richichi B, Fragai M, André S, Gabius HJ, Ardá A, Luchinat C, Jiménez-Barbero J, Nativi C. Targeting matrix metalloproteinases: design of a bifunctional inhibitor for presentation by tumour-associated galectins. Chemistry. 2013, 19, 1896-1902). Branched amphiphilic molecules exhibiting specific inhibitory activity towards individual members of the MMP family have been designed in collaboration with scientists at the Weizmann Institute. These amphiphilic isomers share the same chemical nature, providing versatile nonspecific binding reactivity that allows to probe hidden regulatory residues on a given protein surface. Using the advantage provided by amphiphilic ligands, here we explore a new approach for determining hidden regulatory sites. This approach includes diverse experimental analysis, such as structural spectroscopic analyses, NMR, and protein crystallography combined with computational prediction of effector binding sites. We have demonstrated how our approach works by analyzing members of the MMP family that possess a unique set of such sites. Our work provides a proof of principle for using ligand effectors to unravel hidden regulatory sites specific to members of the structurally homologous MMP family (Melikian M, Toccafondi M, Berezovsky IN, Luchinat C, Sagi I. Unraveling ridde regulatory sites in structurally homologous metalloproteases. J Mol Biol. 2013, 425, 2330-2346). This approach may be exploited for the design of novel molecular effectors and therapeutic agents affecting protein catalytic
RESEARCH
AREAS
21

function via interactions with structure-specific regulatory sites. In addition, a new class of potent matrix metalloproteinase (MMP) inhibitors designed by structure-based optimization of the well-known arylsulfonamide scaffold have been designed at CERM . Molecules show an ethylene linker connecting the sulfonamide group with the P1′ aromatic portion and a d-proline residue bearing the zinc-binding group. The affinity improvement provided by these modifications led us to discover a nanomolar MMP inhibitor bearing a carboxylate moiety as zinc-binding group, which might be a promising lead molecule. Notably, a significant selectivity for MMP-8, MMP-12, and MMP-13 was observed with respect to MMP-1 and MMP-7. In order to clarify the structural and thermodynamic bases of the selectivity, we have investigated a phosphinic inhibitor that is one of the few potent and selective MMP-12 inhibitors reported so far. It is effective, when tested in mice with established atherosclerotic plaques, in blocking plaque growth and in preventing their rupture. Although other selective MMP-12 inhibitors without a phosphinic acid moiety have been reported subsequently by our group and by others, phosphinic peptide inhibitors retain a number of important features that justify pursuing their development as selective inhibitors toward other MMP members. Indeed, phosphinic
peptides are transition-state analogues, they are fairly stable in vivo, and display no metabolism and no toxicity in mice animal models under chronic intravenous administration for over a month. Isothermal titration calorimetry (ITC) experiments indicate that the binding of this inhibitor to MMP-12 is mostly entropy driven and involves a proton uptake from the buffer. Comparing phosphinic versus hydroxamate inhibitors reveals that the chelation of the zinc ion is slightly different, leading the inhibitor backbone to adopt a position in which the hydrogen bonding with the MMP-12 active site is less favorable in phosphinic inhibitor while maintaining high affinity (Czarny B, Stura EA, Devel L, Vera L, Cassar-Lajeunesse E, Beau F, Calderone V, Fragai M, Luchinat C, Dive V. Molecular determinants of a selective matrix metalloprotease-12 inhibitor: insights from crystallography and thermodynamic studies. J Med Chem. 2013, 56, 1149-1159). In 2013 the project FINDING (Farmaci innovativi per malattie neurodegenerative) funded by POR CReO FESR 2007–2013, has been completed with the production of soluble variants of the enzyme QPTC that are suitable of screening studies via solution NMR and X-ray crystallography and have led to the identification of a series of good ligands, to be further optimized by the industrial partner SienaBiotech.
RESEARCH
AREAS
22
Fast Field Cycling Relaxometry Fast field cycling relaxometry is a tool for measuring nuclear relaxation rates from very low magnetic fields (0.01 MHz proton Larmor frequency) up to 1 T (about 45 MHz proton Larmor frequency). The field dependence of the relaxation rates provides information on the structural and dynamic features of a molecule (and, in the case of paramagnetic systems, on the electron relaxation). In the context of the characterization of polarizing agents for solution dynamic nuclear polarization (DNP) experiments, we have collected NMRD (Nuclear Magnetic Relaxation Dispersion) profiles of solvent water protons in the presence of nitroxide radicals (TEMPONE) or in the presence of paramagnetic metal complexes (like manganese(II), gadolinium(III) and oxovanadium(IV)). The fit of the relaxation profiles using a model for dipolar inner-sphere and outer-sphere relaxation provided the structural and dynamic parameters needed for determining the coupling factor, on which the solvent DNP enhancement depends (Neugebauer P, Krummenacker JG, Denysenkov VP, Parigi G, Luchinat C, Prisner TF. (2013) Liquid state DNP of water at 9.2 T: an experimental access to saturation. Phys Chem Chem Phys. Apr 28;15(16):6049-56; Luchinat C, Parigi G, Ravera E (2013). J. Biomol. NMR DOI 10.1007/s10858-013-9728-8). NMRD profiles have been acquired and analyzed also for the characterization of water and protein dynamics in sedimented systems (Luchinat C, Parigi G, Ravera E. (2013) Water and protein dynamics in sedimented systems: a relaxometric investigation. Chem Phys
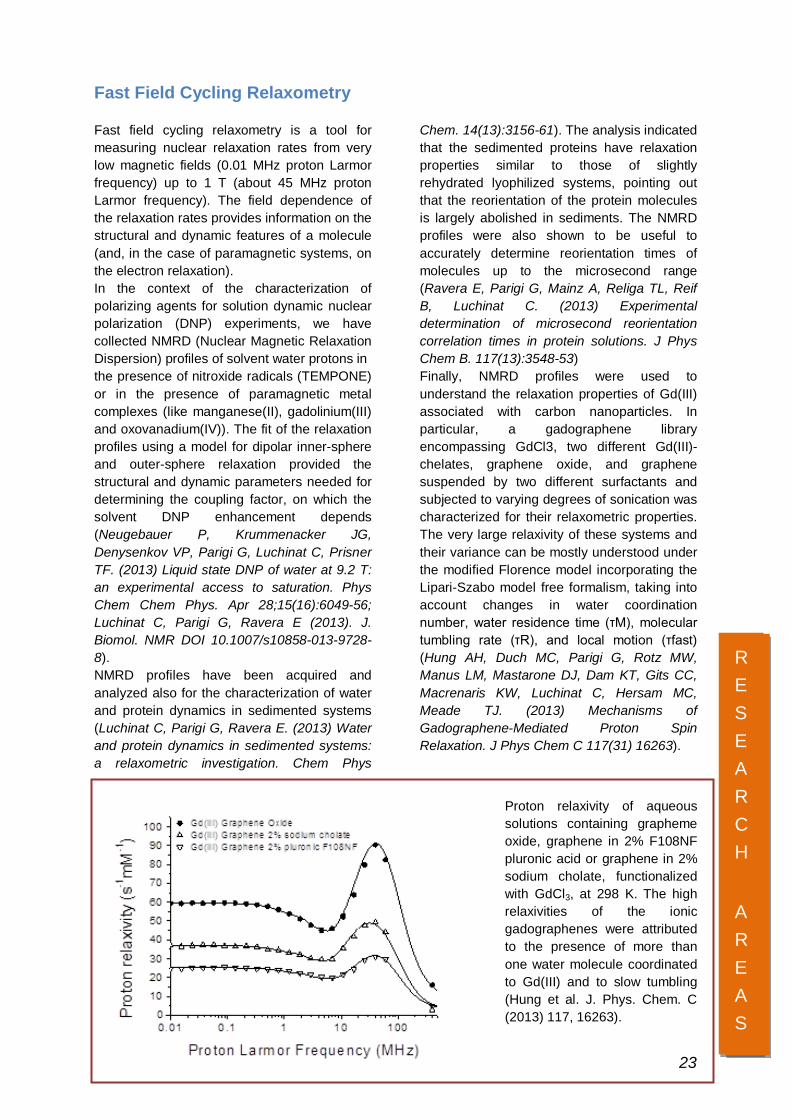
Chem. 14(13):3156-61). The analysis indicated that the sedimented proteins have relaxation properties similar to those of slightly rehydrated lyophilized systems, pointing out that the reorientation of the protein molecules is largely abolished in sediments. The NMRD profiles were also shown to be useful to accurately determine reorientation times of molecules up to the microsecond range (Ravera E, Parigi G, Mainz A, Religa TL, Reif B, Luchinat C. (2013) Experimental determination of microsecond reorientation correlation times in protein solutions. J Phys Chem B. 117(13):3548-53) Finally, NMRD profiles were used to understand the relaxation properties of Gd(III) associated with carbon nanoparticles. In particular, a gadographene library encompassing GdCl3, two different Gd(III)-chelates, graphene oxide, and graphene suspended by two different surfactants and subjected to varying degrees of sonication was characterized for their relaxometric properties. The very large relaxivity of these systems and their variance can be mostly understood under the modified Florence model incorporating the Lipari-Szabo model free formalism, taking into account changes in water coordination number, water residence time (τM), molecular tumbling rate (τR), and local motion (τfast) (Hung AH, Duch MC, Parigi G, Rotz MW, Manus LM, Mastarone DJ, Dam KT, Gits CC, Macrenaris KW, Luchinat C, Hersam MC, Meade TJ. (2013) Mechanisms of Gadographene-Mediated Proton Spin Relaxation. J Phys Chem C 117(31) 16263).
Proton relaxivity of aqueous solutions containing grapheme oxide, graphene in 2% F108NF pluronic acid or graphene in 2% sodium cholate, functionalized with GdCl3, at 298 K. The high relaxivities of the ionic gadographenes were attributed to the presence of more than one water molecule coordinated to Gd(III) and to slow tumbling (Hung et al. J. Phys. Chem. C (2013) 117, 16263).
RESEARCH
AREAS
RESEARCH
AREAS
23

Research with spin-off Institutions Fiorgen Onlus The Pharmacogenomics FiorGen Onlus Foundation (www.fiorgen.net) is a non-profit foundation that promotes studies in predictive and personalized medicine. The Foundation conducts research in the application of genomic and molecular data to help find new products, and to help determine the susceptibility to a particular disease or disorder. CERM and FiorGen work together on the main lines of structural biology and metabolomics. The study of diseases through the holistic approach of metabolomics can be very important to obtain new information on their mechanisms and their impact on human health. During 2013 Fiorgen Foundation continued its research activity in this field, publishing a total of seven paper. At the same time, new emerging applications of metabolomics in the field of food analysis and human nutrition were explored. A fruitful collaboration with the Regional Department of Agriculture was developed, particularly aimed to the valorization of local agricultural products trough the analysis of the molecular fingerprint. Moreover a project devoted to study the metabolic effects of selected probiotics in healthy volunteers is going to start at the beginning of 2014. All the papers published with the contribution of the FiorGen Foundation are significant for the development of the respective scientific fields and are published on high impact factor journals. CERM houses the data processing center and the main biorepository of the da Vinci European BioBank (daVEB;
https://www.davincieuropeanbiobank.org), owned by FiorGen. The interaction between daVEB and CERM is strategic and synergistic. Scientific collaborations in the metabolomic field contribute to the development of SOPs validated by NMR and to the enrichment of the biobank in terms of type and number of samples. daVEB currently houses a collection of unique samples (biofluids, tissues and DNA) of growing importance by number in the following areas: melanoma, rare skin diseases, diseases of the genital-urinary cancer, diseases of the cardio-circulatory, digestive diseases, breast cancer, non-Hodgkin's lymphoma, diseases associated with aging bone. On the other hand, the biobank acts as a support to the metabolomics research via NMR carried out at CERM by providing a conservation service of samples and associated data following protocols in accordance with international standards. As a member of CEN/TC 140, FiorGen is involved in the production of European guidelines for the collection, handling, transport and storage of urine, serum and plasma samples for metabolomic studies. The daVEB biobank is a partner of the project RISE (Competence center-RISE Network infrastructure for industrial research and incubation for advanced services to innovative companies, coordinated CSAVRI) ; PAR-FAS 2007-2013 funding of Regione Toscana (approved May, 21 2013). It will operate as an infrastructure to support experimental development activities and provision of services, with open access to private companies.
Giotto Biotech Srl Giotto Biotech S.r.l. is a new and rapidly growing biotech SME founded in 2011 as a spin-off of CERM which aims at contributing to the biomedical sciences by providing enabling products and services, with a particular focus on complementary technologies to the field of
nuclear magnetic resonance (NMR). Giotto Biotech provides a full range of basic raw materials needed for research in the field of biomedical sciences in custom manufacturing, consulting and services. The company is active in various fields including protein
RESEARCH
AREAS 24

production and isotope labeling, organic synthesis, the development of algae as cell factories, services NMR, and information technologies. The services include NMR metabolomics and statistical analysis. Research projects at Giotto Biotech include: pNMR - It is a project financed by the European Commission that combines 9 academic research groups and 4 collaborating industrial companies to train the next generation of PhD students and post-doctoral researchers, in developing and applying novel experimental and theoretical methods in the NMR spectroscopy of systems containing paramagnetic metals. The team will investigate a variety of important problems in chemistry and biology including catalysts, battery materials, metalloproteins and large protein-protein assemblies. Pathway-27 "Pivotal assesment of the effects of bioactives on health and wellbeing. From human genome to food industry" - a research project carried out by a pan-European interdisciplinary team of 16 life/social scientists and 9 high tech/food processing SMEs that will evaluate the effectiveness of 3 bioactives (docosahexaenoic acid, β-glucan, anthocyanins) as ingredients of enriched foods, evaluating both the bioactive-food matrix interactions and the extent of synergism between the three active molecules. The project will determine the impact of the bioactive enriched foods on physiologically relevant endpoints related to Metabolic Syndrome risk and deliver a better understanding of the role and mechanisms of action of the three bioactives. MAPI-INT –Is a project funded by the Tuscany Region whose goal is to identify early biological markers of intestinal ischemia in
patients who come to the emergency department with abdominal pain. This aim will be achieved mainly through metabolomics analysis of biological samples taken from such patients, in order to identify one or more metabolites as early markers of the disease under study, in order to develop a suitable diagnostic kit for early diagnosis of the disease. ACTILA –Is a new technology platform for multifunctional immunotherapy of cancer: synergy between adoptive allogenic cell therapy, monoclonal antibodies and immunostimulatory molecules; the project is financed by the Tuscany Region. Giotto Biotech is a partner in the following European projects together with CERM: IDPbyNMR –Is a project that is part of the Marie Curie Action for Initial Training Network (ITN) in the 7th Framework Programme of the European Commission, the purlose of which is to establish a framework to train a new generation of young researchers in the emerging area of the intrinsically disordered proteins, proteins that gain functional advantages by remaining natively unstructured, either completely or partially, thus challenging well-established concepts in structural biology. BIOLABEL –Is a project financed by MIUR, whose ultimate goal is the marketing of products / biomolecules isotopically enriched, technologically advanced and at a lower cost than currently available. The achievement of these objectives is to maximize the growth of isotopically enriched microalgae on an industrial scale and the development, adaptation and optimization techniques for the production of biomolecules variously marked for scientific use.
RESEARCH
AREAS
25

INSTRUMENTATION
Instrumentation Solution and solid state NMR spectrometers All NMR instruments are state-of-the-art, digital spectrometers equipped with a variety of cryoprobes as well as of probes over a broad range of frequency range and a number of nuclei. In addition to all the standard pulse sequences for spectroscopic, structural, dynamical, and functional characterization, tailored pulse sequences for structural determination of high molecular weight
proteins and paramagnetic systems are implemented, as well as 13C direct detection protocols for “protonless” NMR experiments and structural characterization of biomolecules, including unfolded or partially unfolded ones. Special protocols and devices are available for solid state experiments. Set-up and pulse sequences for in-cell NMR experiments are also implemented.
26

Field (T) 22.3 1H Frequency (MHz) 950 Probe types Cryo probes
Probes TCI Cryo (1H/13C/15N/D with 2H decoupling)
SampleFormat 5 mm (tubes) Bore 54 mm SampleType Solution, cells, micelles
PulseLibrary
Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition, 13C direct detection biomolecular NMR experiments
Field (T) 21.1 1H Frequency (MHz) 900 Probe types Cryo and RT probes
Probes
TCI Cryo (1H/13C/15N with 2H decoupling) TXI RT (1H/13C/15N with 2H decoupling)
SampleFormat 5 mm (tubes) Bore 54 mm SampleType Solution, cells, micelles
PulseLibrary
Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition, 13C direct detection biomolecular NMR experiments
INSTRUMENTATION
27
950 MHz •three-channel and 2H
decoupling instrument, equipped with a TCI cryoprobe
900 MHz •three-channel and 2H
decoupling instrument, equipped with a TCI cryoprobe
850 MHz wide bore •with MAS accessories for
solid-state studies, with 1.3 mm and 3.2 mm triple resonance probes
800 MHz •four-channel instrument,
equipped with a TXI cryoprobe and a high-power probe for the study of fast-relaxing systems
700 MHz wide bore •with MAS accessories for
solid-state studies, with 3.2 mm and 4 mm double and triple resonance probes
700 MHz •four-channel instrument,
equipped with a TXO cryoprobe for 13C direct detection with 1H, 2H, 13C and 15N decoupling
700 MHz •three-channel and 2H
decoupling instrument, with a TCI cryoprobe and automatic sample changer
600 MHz •three-channel and 2H
decoupling instrument, equip-ped with a TCI 1H/13C/31P cryoprobe, as well as an automatic sample changer
600 MHz •with HR-MAS accessories and
a number of BB probes which cover a frequency range from 600 to 20 MHz
500 MHz •three-channel instrument
equipped with TCI cryoprobe 400 MHz Fast Field Cycling Relaxometer
Field (T) 20.0 1H Frequency (MHz) 850 Probe types MAS
Probes 3.2 mm MAS DVT 15N/13C/1H 1.3 mm MAS 1H-19F/BB/15N
SampleFormat 1.3 mm , 3.2 mm (rotors) Bore 89 mm
SampleType Powder, microcrystals, precipitated, fibrils, membranes, solution
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
Field (T) 18.8 1H Frequency (MHz) 800 Probe types Cryo and RT probes
Probes
TXI Cryo (1H/13C/15N with 2H decoupling) TXI RT (1H/13C/15N with 2H decoupling) QXI RT (1H/13C/15N/31P with 2H decoupling) 1H-SEL High Power (prototype)
SampleFormat 5 mm , 2.5 mm for 1H-SEL probe (tubes)
Bore 54 mm SampleType Solution, cells, micelles
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
Field (T) 16.4 1H Frequency (MHz) 700 Probe types Cryo and RT probes
Probes
TXO Cryo (13C/15N/1H with 2H decoupling) TXI RT (1H/13C/15N with 2H decoupling)
SampleFormat 5 mm (tubes) Bore 54 mm SampleType Solution, cells, micelles
PulseLibrary
Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition, 13C direct detection biomolecular NMR experiments
Field (T) 16.4 1H Frequency (MHz) 700
Probe types Cryo and RT probes
Probes
TCI Cryo (13C/15N/1H with 2H decoupling)
2 x TXI RT (1H/13C/15N with 2H decoupling)
SampleFormat 5 mm (tubes) Bore 54 mm SampleType Solution, cells, micelles SpecialAccessories Sample Changer (120 positions)
PulseLibrary
Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition, 13C direct detection biomolecular NMR experiments
Field (T) 16.4 1H Frequency (MHz) 700 Probe types MAS
Probes 3.2 mm MAS 15N/13C/1H 4 mm MAS 15N/13C/1H
SampleFormat 4 mm , 3.2 mm (rotors) Bore 89 mm
SampleType Powder, microcrystals, precipitated, fibrils, membranes, solution
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
INSTRUMENTATION
28
Field (T) 14.1 1H Frequency (MHz) 600
Probe types RT probes, MAS probes
Probes
TXI RT (1H/13C/15N with 2H decoupling) HR-MAS (1H/13C/15N with 2H decoupling) MAS-1H selective 1H-SEL High Power RT 5 mm BBO RT 10 mm BBO RT 10 mm BB (LowFreq) RT 5 mm BBI RT
SampleFormat 5 mm, 10 mm (tubes), 4 mm (rotors)
Bore 54 mm
SampleType Solution, cells, micelles, tissues
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
Field (T) 14.1 1H Frequency (MHz) 600
Probe types Cryo and RT probes
Probes
TCI Cryo (1H/13C/31P with 2H decoupling) TXI RT (1H/13C/15N with 2H decoupling)
SampleFormat 5 mm
Bore 54 mm
SampleType Solution, cells, micelles
SpecialAccessories Sample Changer (120 positions), Bruker liquid handler system
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
Field (T) 11.7 1H Frequency (MHz) 500
Probe types Cryo and RT probes
Probes TCI Cryo (1H/13C/15N) TXI RT (1H/13C/15N) BBI RT
SampleFormat 5 mm
Bore 54 mm
SampleType Solution, cells, micelles
PulseLibrary
Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition, 13C direct detection biomolecular NMR experiments
Field (T) 9.4 1H Frequency (MHz) 400 Probe types RT probes
Probes BBO RT 1H-Selective
SampleFormat 5 mm
Bore 54 mm
SampleType Solution, cells, micelles
PulseLibrary Standard Bruker Libraries, Home-built libraries for 1D to nD data acquisition
Fast Field Cycling Relaxometry
Field (T) 0.00024-1
1H Frequency (MHz) 0.01-42
SampleFormat 10 mm (tubes)
SampleType Solution
INSTRUMENTATION
29

X-ray Crystallography CERM is equipped with standard crystallization facilities and with an automated nano-dispensing device (Mosquito, TTP Labtech).
Furthemore it has full access to the Interdepartmental Crystallography Centre of the University of Florence (CRIST), equipped,
among other instruments, with a sealed-tube diffractometer bearing a CCD detector (AgilentTechnologies) for routinely in-house data collections, and regular access to synchrotron beam time slots in Europe.
Biological and Biophysical Equipment and Facilities CERM is equipped with state-of-the-art facilities for large scale protein cloning, expression (in prokaryotes through the use of fermenters) and also for expression in
eukaryotic cells. Fully equipped facilities for protein purification are also available. A wide array of equipment is available for the biochemical and biophysical characterization of biomolecules. They include:
EPR CERM has access to 9.43 GHz (X-Band, ElexSys E500) and 94 GHz (W-Band, ElexSys E600) instruments.
Frequency 9.43 GHz (X-Band) Model ElexSys E500 Cavity SHQ, ST102 Cryogenic apparatus Helium, nitrogen SampleHolder Tube, quartz tube, quartz rod SampleType Solution, frozen solution, lyophilized powder, single crystal samples.
EQUI PMENT
&
FACILITIES 30
Multi Angle/Dynamic Light Scattering CERM has DLS instrument for measurements on batch samples or on in-flow samples (FPLC coupling).
DetectorType Refractive Index
StdWavelength 658 nm
Resolution 1 nm
Surface Plasmon Resonance CERM is equipped with an SPR device for studying protein-ligand or protein-protein interactions.
Model ProteOn XPR36 NumberOfInteractionSpots 36 UniformityOfResponse > 0.98 DynamicRange 1-40000 RU ExperimentTemperatureRange 15-40° C MassDetectionLimit 201 Da ConcentrationDetectionLimit subnanomolar SpecialAccessories Autosampler BaselineNoise < 1.5 RU
Isothermal Calorimetry (ITC) CERM is equipped with an ITC device to study the thermodynamics of protein-ligand of protein-protein interactions
Model VP-ITC MicroCalorimeter Noise level (3 minutes RMS windows averaged over 1 hour) 1 nanocal/sec (4 nanowatts)
Baseline constancy (average one hour time intervals: 310 rpm stirring)
± 5 nanocal/sec (± 20 nanowatts)
Minimum response time 15 seconds Operating temperature range 2° to 80° C Cell volume 1.4301 mL Syringe volume 293 µL
Circular Dichroism CERM is equipped with a CD device to gain information on the secondary structure of proteins
Frequency 94 GHz (W-Band) Model ElexSys E600 Cavity Low-Q, High-Q Cryogenic apparatus Helium, nitrogen SampleHolder Tube, quartz tube, quartz rod SampleType Solution, frozen solution, lyophilized powder, single crystal samples.
EQUI PMENT
&
FACILITIES
31

Model Jasco J 810 Light source 150W air cooled xenon lamp Detector Head-on photomultiplier tube Measurement wavelength 163 to 900 nm Spectral bandwidth 0.01 to 15 nm Response 0.5 msec to 32 sec Scanning system Continuos scan; step scan Scanning speed to 10000 nm/min (continuos scan)
Data interval: 0.025 to 10 nm (step scan) 0.1 to 100 nm (step scan) 0.5 msec to 60 min (time change)
Cd full scale ± 10,200,2000 mdeg
CD resolution 0.0005 mdeg (at ± 10 mdeg full scale) 0.01 mdeg (at ± 200 mdeg full scale) 0.1 mdeg (at ± 2000 mdeg scale)
Nitrogen gas displacement
Atmosphere in the light source unit, monochromator unit, and sample chamber is displaced using dry nitrogen gas
Fluorimetry CERM is equipped with a Fluorescence Spectrophotometer
Brand and Model Varian Cary Eclipse
Measurement modes Fluorescence, phosphorescence, chemiluminescence and bioluminescence
Source Xe pulsed lamp
Wavelength range (nm)
Mechanical Excitation: 190–1100 nm. Zero order selectable. Emission: 190–1100 nm. Zero order selectable. Operational Excitation: 200–900 nm with standard PM tube. Zero order selectable. Emission: 200–900 nm with standard PM tube. Zero order selectable.
Wavelength accuracy (nm) ± 1.5 nm Wavelength reproducibility (nm) ± 0.2 nm
Detectors High performance R928 photomultiplier detectors. Separate R928 PMT for reference signal
Sensitivity (Raman Band of Water)
>500:1 RMS, 500 nm excitation, excitation and emission slits 10 nm, 1 sec Signal Averaging time. >750:1 RMS, 350 nm excitation, excitation and emission slits 10 nm, 1 sec Signal Averaging time
Minimum sample volume 0.5 mL (using standard 10 mm fluorescence cell)
Accessories Single peltier cell Holder Microplate reader (not thermostatized)
Stopped-flow Spectrophotometer CERM is equipped with a stopped-flow Spectrophotometer
Brand and Model Applied Photophysics SX.18MV Light source
150W air-cooled Xe arc lamp Power controlled Xenon lamp power supply – Arc lamp ignite
Cell block volume 20 μL
EQUI PMENT
&
FACILITIES
32

Pathlength 10 mm and 2 mm Dead time 1 ms Drive volume 100μL per syringe Mixing options Single mixing stopped-flow Monochromator 05-109 Spectra Kinetic Monochromator
Useful operating range of 200 to beyond 700 nm Detector Applied Photophysic Absorption and Emission
Photomultiplier Hardware and Software Acorn 5000 computer equipped with RISC OS 3
operating system SX.18MV Kinetic Spectrometer Workstation Software Number of points in each kinetic trace: 400 to 4000
Biobank CERM hosts the main repository and the IT center of the da Vinci European BioBank (https://www.davincieuropeanbiobank.org/) for biological (both human and non-human) sample storage and retrieval. Through this biobank CERM
has established connections with the ESFRI European Biobank Infrastructure BBMRI. The connection with BBMRI is also enforced by the metabolomics research activities of CERM and its spin-off FiorGen foundation.
Sample type Human biospecimens (plasma, serum, urine, tissues, cells),
bacterial expression vectors.
Sample storage
Mechanical freezers for storage at -80° C, equipped with auxiliary LN2 cooling system. Tanks for cryopreservation in nitrogen vapour phase (-150° C), with automatic nitrogen supply
Storage supervision and control system
Sample storage and withdrawal handled by the software Easy Track 2D. Continuous monitoring of storage temperature.
Database
Information system based on a set of server clusters equipped with Linux Red Hat Enterprise operating systems and Oracle RAC databases. Completely automatized and high speed robotic backup systems. Online browsing of daVEB inventory to obtain samples and/or associated data.
Bioinformatics and computational structural biology tools CERM provides integrated databases and software for genome browsing, metal binding analysis, structure calculation with/without paramagnetic constraints, sequence validation, domain organization, evolution, protein complex analysis.
Access to programs for NMR data processing and structural calculations is also provided via web, and through the Grid computing platforms.
Grid computing CERM is a node of the electronic infrastructure within the EC- project WeNMR (http://www.wenmr.eu), which in November 2013 reached its conclusion. . The overall goal has been to enable analysis of NMR data, alone or in combination with other techniques for structural investigation of biomacromolecules (mainly SAXS), in a highly
automated manner using standardized protocols through web portals. Nevertheless, the WeNMR e-Infrastructure remains operational, to continue to serve its vibrant and steadily growing users community. The overall appreciation of WeNMR at its final review meeting in Brussels was very high: indeed, the report by the reviewing committee described
EQUI PMENT
&
FACILITIES
33

WeNMR results as “exceeding expectations”. The user community now counts more than 1,000 researchers registered within the WeNMR Virtual Research Community and about 600 members of the Virtual Organization, i.e. researchers allowed to make use of the European grid Infrastructure via the WeNMR portals and software tools. More than 90 publications in peer-reviewed journals acknowledged use of the WeNMR resources. Within this e-infrastructure, CERM has implemented and maintains the following programs: AnisoFit for the fitting of orientation and paramagnetic tensors; Antechamber for the calculation of force field parameters for organic ligands, aimed at the subsequent use with AMBER; XPLOR-NIH for the calculation of 3D structures of biomacromolecules from NMR data; AMBER for structural refinement as well as long molecular dynamics simulations;
MaxOcc for the calculation of the maximum possible occurrence of protein conformations in multi-domain proteins from a combination of paramagnetic NMR data and SAXS; sedNMR for the calculation of the properties of sedimented solutes that are relevant to the outcome of solid-state NMR experiments. CERM is a member of the Italian Grid Initiative, through which is involved in the European Grid Initiative. Finally, CERM is a partner of the EC funded BioMedBridges project which involves all the various BioMedical ESFRI Research Infrastructures. It aims at enhancing the excellence of the individual biomedical sciences infrastructures and at ensuring good coordination across the different domains by building concrete e-bridges linking them and thus providing a complex knowledge environment comprising standards, ontologies, data and services.
EQUI PMENT
&
FACILITIES
34

NMR Infrastructure European NMR Infrastructure CERM/CIRMMP provides an ideal environment for research in the field of NMR spectroscopy applied to biological systems. The uniqueness of the infrastructure lies in the wide number of available instruments. Of the various types and in the unique expertise. This allows the optimal use of the instrumentation also in a combined way, when needed. All of the high-resolution NMR instruments are part
of the digital Bruker Avance series. Most of them are equipped with cryoprobes to improve the sensitivity of 1H and/or 13C, including a prototype 13C cryoprobehead. Several probeheads (heads measuring single, double and triple resonance, low and high frequency, high power) are also available to meet specific needs. At the 600 MHz NMR spectrometer a card to remove radiation damping is available. The 500, a 600 MHz, two 700 MHz, the 800 MHz, and the 900 MHz spectrometers are equipped with a CryoProbe. One of the two 700 MHz spectrometers is equipped with a prototype CryoProbe for 13C direct detection, developed with the scientific input and contribution of CERM. Thanks to promising results, Bruker also developed an optimised CryoProbe not only for proton detection, but also for carbon (TCI) on the market; the 950,
900 and 700 MHz spectrometers are equipped with this device. The highest magnetic field available for solution NMR is the 950 MHz equipped with a TCI CryoProbe. A 700 MHz WB and 850 MHz WB are dedicated to solid-state NMR spectroscopy. The 700 MHz is equipped with a 4 mm and 3.2 mm MAS double and triple resonance probes. The 850 MHz, Ultra Shielded Ultra Stabilized is
equipped with 1.3 mm and 3.2 mm MAS probes for 13C, 15N and 1H detection. With regards to low-field instruments, CERM is a unique example for the efforts made towards the development of relaxometry. The most sensitive instrument for low-field relaxation (High Sensitivity Fast Field Cycling NMR) developed during RTD projects for the EC and patented by Stelar (Italy), is available, and covers the 0.01-45 MHz range. The computational infrastructure of CERM includes a cluster for the more intensive calculations, with 16 blades harbouring a total of 80 CPU cores. Ten servers are use to host services from web pages to databases and to enable access to the European Grid. A number of graphic stations are available for interactive NMR data analysis.
INFRASTRUCTURE 35

A molecular biology and a cellular biology lab are available to users, together with other biophysical equipment for EPR, CD, UV-vis, stopped-flow measurements, manual and automated crystallization facilities and X-ray diffractometry. Users can also access other university infrastructures, such as those of mass spectrometry, Raman resonance, and non-linear spectroscopies.
Access to European users is funded by the European Commission since 1994. In 2013 this funding was provided through two different programs: EastNMR (www.east-nmr.eu) and BioNMR (www.bio-nmr.net). Overall, under the two programs, 360 days of machine time were granted to European users for 19 different research projects.
NMR National Access In 2013, several National Service users from public and private laboratories used CERM instrumentation. Although the MIUR – FIRB Project, that provided Access to the Users, has been completed in June 2010, this center decided to maintain its instrumentation available to all National Users, provided their research project matched quality criteria in terms of scientific interest, excellence and feasibility, as assessed by an external Evaluation Commetee. The NMR National Service protocol provides access to all including Technical Assistance for both spectra acquisition and elaboration. Users can use CERM’s chemistry and biological labs to complete their sample preparation or carry out necessary chemical or biochemical procedures. National Service users can access the instruments and labs of several Interdepartmental Centres in strict collaboration with CERM, such as the Genomics and Proteomics Centre GeneExpress, the Interdepartmental Centre of Mass Spectrometry, and all instruments
available at the University of Florence’s Department of Chemistry. All users who used the NMR National Service applied for access through an on-line portal submitting the research project and the needed instrument access. Applications were considered and accepted by a local panel, composed of members of the CERM Faculty and another panel made up of national experts. Both panels gave favourable opinions, respectively, based on technical feasibility of the scientific relevance and the appropriateness of the application with the presented project. Overall, national academic visitors access to the Facility accounted for a total of 55 days. The access of users from private entities substantially increased with respect to 2012. The days of machine time that CERM made available to national members (both from public research laboratories and private entities) amounted to 218 spectrometer days without taking into account CIRMMP and CERM Associates (scientists from the University of Bologna or from Fiorgen).
INFRASTRUCTURE
36

International Doctorate in Structural Biology The International PhD course in Structural Biology is a research doctorate running in collaboration with the University of Frankfurt and Utrecht University. It is hosted by the Magnetic Resonance Center (CERM) of the University of Florence. The scientific fields cover most of the molecular aspects of life sciences. The main objective of the International PhD course in Structural Biology is the training of research doctors at the forefront of the knowledge in modern methodologies in molecular and structural biology, biotechnology and systems biology. It provides both theoretical and hands-on training in structural techniques applied to biological macromolecules in solution and in the crystalline state, as well as in non crystalline materials such as fibrils or proteins in membranes, and to biological macromolecules in their cellular environment. It also provides state-of-the-art training in molecular biology for the expression of isotope-enriched recombinant proteins for NMR studies. Finally, it offers top level ICT training through being a central node of WeNMR, the second largest European Grid Virtual Organization in Life Sciences. Bioinformatics and biostatistics training is secured through WeNMR and through a very active program of NMR-metabolomics studies. The scientific themes tackled by the course are: 1. nuclear magnetic resonance (in solution and in the solid state) and X-ray crystallography aimed at studying structure, function and dynamics in biological macromolecules and protein-protein adducts; 2. molecular biology techniques for the production of proteins, DNA and bacterial and prokaryotic cell growth; 3. development of new drugs, through drug design techniques; 4. bioinformatics as a basic strategy for the study of kinetics and thermodynamics of folding, protein dynamics, and for the exploitation and the development of methodologies for the use of genomic data banks in order to identify new molecular targets for the development of novel drugs;
5. In cell NMR studies, by which molecular pathways and cell import-export mechanisms are investigated 6. metabolomics studies, in which the individual metabolic fingerprints are related to disease states and fingerprints are utilized to provide early diagnosis or even identification of pre-disease states. The added value of this course is in the development of a transnational educational
project, able to create PhD doctors at the forefront regarding the scientific formation, the knowledge and development of research and technology, capable to consider multi-disciplinarity, transnational cooperation and mobility as primary needs, to evaluate collaborative projects as a requirement for high quality research. The doctoral program also relies on Faculty members who, in addition to scientists from CERM, include professors from other departments of the University of Florence and from the Universities of Frankfurt, Utrecht, Oxford and Lyon, all top places for Structural Biology. Full-time attendance is mandatory as is commitment to research activities. In addition to seminars and courses, students are asked to provide research seminars as a basic tool for their own training. Every PhD student is encouraged to liaise with foreign universities and take part in teaching and research training as well as internships abroad.
INFRASTRUCTURE 37

List of 2013 Publications
1. Bertini, I., Luchinat, C., Parigi, G., and Ravera, E., SedNMR: on the edge between solution and solid state NMR, Acc.Chem.Res., 46, 2059-2069, 2013 (IF 20.833).
2. Knight, M. J., Felli, I. C., Pierattelli, R., Emsley, L., and Pintacuda, G., Magic angle spinning NMR of paramagnetic proteins, Acc.Chem.Res., 46, 2108-2116, 2013 (IF 20.833).
3. Lalli, D. and Turano, P., Solution and solid state NMR approaches to draw iron pathways in the ferritin nanocage., Acc.Chem.Res., 46, 2676-2685, 2013 (IF 20.833).
4. Banci, L., Barbieri, L., Bertini, I., Luchinat, E., Secci, E., Zhao, Y., and Aricescu, A. R., Atomic-resolution monitoring of protein maturation in live human cells by NMR, Nat.Chem.Biol., 9, 297-299, 2013 (IF 12.948).
5. Theil, E. C. and Turano, P., Metalloenzymes: cage redesign explains assembly, Nat.Chem.Biol., 9, 143-144, 2013 (IF 12.948).
6. Limongelli, V., De Tito, S., Cerofolini, L., Fragai, M., Pagano, B., Trotta, R., Cosconati, S., Marinelli, L., Novellino, E., Bertini, I., Randazzo, A., Luchinat, C., and Parrinello, M., The G-triplex DNA, Angew.Chem.Int.Ed., 52, 2269-2273, 2013 (IF 13.734).
7. Gil, S., Hosek, T., Solyom, Z., Kümmerle, R., Brutscher, B., Pierattelli, R., and Felli, I. C., NMR studies of intrinsically disordered proteins near physiological conditions, Angew.Chem.Int.Ed., 52, 11808-11812, 2013 (IF 13.734).
8. Fragai, M., Luchinat, C., Parigi, G., and Ravera, E., Conformational freedom of metalloproteins revealed by paramagnetism-assisted NMR, Coord.Chem.Rev., 257, 2652-2667, 2013 (IF 11.016).
9. Knight, M. J., Felli, I. C., Pierattelli, R., Bertini, I., Emsley, L., Hermann, T., and Pintacuda, G., Rapid measurement of pseudocontact shifts in metalloproteins by proton detected solid - state NMR spectroscopy, J.Am.Chem.Soc. 13, 4, 14730-14733, 2013 (IF 10.677).
10. Ravera, E., Corzilius, B., Michaelis, V. K., Rosa, C., Griffin, R. G., Luchinat, C., and Bertini, I., Dynamic nuclear polarization of sedimented solutes, J.Am.Chem.Soc., 135, 1641-1644, 2013 (IF 10.677).
11. Banci, L., Bertini, I., Calderone, V., Ciofi-Baffoni, S., Giachetti, A., Jaiswal, D., Mikolajczyk, M., Piccioli, M., and Winkelmann, J., Molecular view of an electron transfer process essential for iron-sulfur protein biogenesis, Proc.Natl.Acad.Sci.U.S.A, 110, 7136-7141, 2013 (IF 9.737).
12. Rosato, A., Tejero, R., and Montelione, G. T., Quality assessment of protein NMR structures, Curr Opin Struct Biol, 23, 715-724, 2013 (IF 8.738).
13. Varadi, M., Kosol, S., Lebrun, P., Valentini, E., Blackledge, M., Dunker, A. K., Felli, I. C., Forman-Kay, J., Kriwacki, R. W., Pierattelli, R., Sussman, J. L., Svergun, D. I., Uversky, V. N., Vendruscolo, M., Wishart, D. S., Wright, P. E., and Tompa, P., pE-DB: a database of structural ensembles of intrinsically disordered and of unfolded proteins, Nucleic acids res., 2013, epub ahead of print (IF 8.278).
14. Andreini, C., Cavallaro, G., Lorenzini, S., and Rosato, A., MetalPDB: a database of metal sites in biological macromolecular structures, Nucleic Acids Res., 41, D312-D319, 2013 (IF 8.278).
15. Fragai, M., Luchinat, C., Martelli, T., Ravera, E., Sagi, I., Solomonov, I., and Udi, Y., SSNMR of biosilica-entrapped enzymes, Chemical Communication, 50, 421-423, 2013 (IF 6.378).
16. Banci, L., Barbieri, L., Luchinat, E., and Secci, E., Visualization of redox-controlled protein fold in living cells, Chem.Biol., 20, 747-752, 2013 (IF 6.157).
17. Bartoloni, M., Dominiquez, B. E., Dragoni, E., Richichi, B., Fragai, M., Andre', S., Gabius, H.-J., Ardà, A., Luchinat, C., Jimenez-Barbero, J., and Nativi, C., Targenting matrix
PUBLICATIONS
38

metalloproteinases:design of a bifunctional inhibitor of presentation by tumor-associated galectins, Chem.Eur.J., 19, 1896-1902, 2013 (IF 5.831).
18. Czarny, B., Stura, E. A., Devel, L., Vera, L., Cassar-Lajeunesse, E., Beau, F., Calderone, V., Fragai, M., Luchinat, C., and Dive, V., Molecular determinants of a selective matrix metalloprotease-12 inhibitor: insights from crystallography and thermodynamic studies, J.Med.Chem., 56, 1149-1159, 2-14-2013 (IF 5.614).
19. Tenori, L., Hu, X., Pantaleo, P., Alterini, B., Castelli, G., Olivotto, I., Bertini, I., Luchinat, C., and Gensini, G., Metabolomic fingerprint of heart failure in humans: a nuclear magnetic resonance spectroscopy analysis, In.J.Cardiol., 164, e113-115, 2013 (IF 5.509).
20. Cacciatore, S., Hu, X., Viertler, C., Kap, M., Bernhardt, G. A., Mischinger, H. J., Riegman, P., Zatloukal, K., Luchinat, C., and Turano, P., The effects of intra- and postoperative ischemia on the metabolic profile of clinical tissue specimens monitored by NMR, J.Proteome Res., 12, 5723-5729, 2013 (IF 5.056).
21. Hung, A. H., Duch, M. C., Parigi, G., Rotz, M. W., Manus, L. M., Mastarone, D. J., Dam, K. T., Gits, C. C., MacRenaris, K. W., Luchinat, C., Hersam, M. C., and Meade, T. J., Mechanisms of gadographene-mediated proton spin relaxation, J.Phys.Chem.C, 117, 16263-16273, 2013 (IF 4.814).
22. Cerofolini, L., Fields, G. B., Fragai, M., Geraldes, C. F. G. C., Luchinat, C., Parigi, G., Ravera, E., Svergun, D. I., and Teixeira, J. M. C., Examination of Matrix Metalloproteinase-1 (MMP-1) in solution: a preference for the pre-collagenolysis state, J.Biol.Chem., 288, 30659-30671, 2013 (IF 4.651).
23. Bertini, I., Luchinat, C., Miniati, M., Monti, S., and Tenori, L., Phenotyping COPD by 1H-NMR metabolomics of exhaled breath condensate, Metabolomics, 2013, epub ahead of print, (IF 4.433).
24. Andreini, C., Cavallaro, G., Rosato, A., and Valasatava, Y., MetalS2: a tool for the structural alignment of minimal functional sites in metal-binding proteins and nucleic acids, J.Chem.Inf.Model., 53, 3064-3075, 2013, (IF 4.304).
25. Banci, L., Bertini, I., Cefaro, C., Ciofi-Baffoni, S., Gajda, K., Felli, I. C., Gallo, A., Pavelkova, A., Kallergi, E., Andreadaki, M., Katrakili, N., Pozidis, C., and Tokatlidis, K., An intrinsically disordered domain has a dual function coupled to comportment-dependent redox control, J.Mol.Biol., 425, 594-608, 2013 (IF 3.905).
26. Udi, Y., Fragai, M., Grossman, M., Mitternacht, S., Arad-Yellin, R., Calderone, V., Melikian, M., Toccafondi, M., Berezovsky, I. N., Luchinat, C., and Sagi, I., Unraveling hidden regulatory sites in structurally homologous metalloproteases, J.Mol.Biol., 425, 2330-2346, 2013 (IF 3.905).
27. Neugebauer, P., Krummenacker, J. G., Denysenkov, V. P., Parigi, G., Luchinat, C., and Prisner, T. F., Liquid state DNP of water at 9.2 T: an experimental access to saturation, Phys.Chem.Chem.Phys., 15, 6049-6056, 2013 (IF 3.829).
28. Almeida, R. M., Turano, P., Moura, I., Moura, G. L. C., and Pauleta, S. R., Superoxide reductase: different interaction modes with its two redox partners, ChemBioChem, 14, 1858-1866, 2013 (IF 3.740).
29. Banci, L., Cantini, F., Kozyreva, T., and Rubino, J. T., Mechanistic aspects of hSOD1 maturation from the solution structure of Cu1-loaded hCCS domain 1 and analysis of disulfide-free hSOD1 mutants, ChemBioChem, 14, 1839-1844, 2013 (IF 3.740).
30. Calcada, E. O., Felli, I. C., Hosek, T., and Pierattelli, R., The heterogeneous structural behaviour of E7 from HPV16 revealed by NMR spectroscopy, ChemBioChem, 14, 1876-1882, 2013 (IF 3.740.
31. Bertini, I., Gallo, G., Korsak, M., Luchinat, C., Mao, J., and Ravera, E., Formation Kinetics and Structural Features of Beta-Amyloid Aggregates by Sedimented Solute NMR, ChemBioChem, 14, 1891-1897, 2013 (IF 3.740).
32. Bhaumik, A., Luchinat, C., Parigi, G., Ravera, E., and Rinaldelli, M., NMR crystallography on paramagnetic systems: solved and open issues, Cryst.Eng.Comm., 15, 8639-8656, 2013 (IF 3.879).
PUBLICATIONS
39

33. Breydo, L., Reddy, K.D., Piai, A., Felli, I.C., Pierattelli, R., Uversky, V.N., The crowd you're in with: effects of different types of crowding agents on protein aggregation, Biochim Biophys Acta, 2013, epub ahead of print (IF 3,733)
34. Castaldo, C., Ciambellotti, S., Pablo-Latorre, R., Lalli, D., Porcari, V., and Turano, P., Soluble variants of human recombinant glutaminyl cyclase, Plos ONE, 8, e71657-, 2013 (IF 3.730).
35. Ravera, E., Parigi, G., Mainz, A., Religa, T. L., Reif, B., and Luchinat, C., Experimental determination of microsecond reorientation correlation times in protein solutions, J.Phys.Chem.B, 117, 3548-3553, 2013 (IF 3.607).
36. Felli, I. C., Putting the right spin on it, Chem.Phys.Chem., 14, 2998-2999, 2013 (IF 3.412).
37. Musiani, F., Ippoliti, E., Micheletti, C., Carloni, P., and Ciurli, S., Conformational fluctuations of UreG, an intrinsically disordered enzyme, Biochemistry, 52, 2949-2954, 2013 (IF 3.377).
38. Veggi, D., Gentile, M. A., Cantini, F., Lo Surdo, P., Nardi-Dei, V., Seib, K. L., Pizza, M., Rappuoli, R., Banci, L., Savino, S., and Scarselli, M., The Factor H Binding Protein of Neisseria meningitidis Interacts with Xenosiderophores in vitro, Biochemistry, 51, 9384-9393, 2013 (IF 3.377).
39. Gomez-Mingot, M., Alcaraz, L. A., Heptistall, J., Donaire, A., Piccioli, M., Montiel, V., and Iniesta, J., Electrochemical nitration of myoglobin at tyrosine103: structure and stability, Arch.Biochem.Biophys., 529, 26-33, 2013 (IF 3.370)
40. Banci, L., Ciofi-Baffoni, S., Mikolajczyk, M., Winkelmann, J., Bill, E., and Eirini Pandelia, M., Human anamorsin binds (2Fe.2S) clusters with unique electronic properties, J.Biol.Inorg.Chem., 18, 883-893, 2013 (IF 3.353).
41. Banaszak, K., Merloni, A., Zambelli, B., Kiliszek, A., Rypniewski, W. R., and Ciurli, S., Selectivity of Ni(II) and Zn(II) binding to sporosarcina pasteurii UreE, a metallo-chaperone in the urease assembly: a calorimetric and crystallographic study, J.Biol.Inorg.Chem., 18, 1005-1017, 2013 (IF 3.353).
42. Benini, S., Kosikowska, P., Cianci, M., Mazzei, L., Berlicki, L., and Ciurli, S., The crystal structure of sporacina pasteurii urease in a complex with citrate provides new hints for inhibitor design, J.Biol.Inorg.Chem., 18, 391-399, 2013 (IF 3.353).
43. Bertini, I., Borsi, V., Cerofolini, L., Das Gupta, S., Fragai, M., and Luchinat, C., Solution structure and dynamics of human S100A14, J.Biol.Inorg.Chem., 18, 183-194, 2013 (IF 3.353).
44. Biagi, F., Musiani, F., and Ciurli, S., Structure of the UreD-UreF-UreG-UreE complex in helicobacter pylori: a model study, J.Biol.Inorg.Chem., 18, 571-577, 2013 (IF 3.353).
45. Zambelli, B., Berardi, A., Martin-Diaconescu, V., Mazzei, L., Maroney, M. J., and Ciurli, S., Metal binding properties of helicobacter pylori UreF, an accessory protein in the nickel-based activation of urease, J.Biol.Inorg.Chem., 2013, epub ahead of print (IF 3.353)
46. Wegerich, F., Giachetti, A., Allegrozzi, M., Lisdat, F., and Turano, P., Cytochrome c and superoxide: a reply, J.Biol.Inorg.Chem., 18, 867-869, 2013 (IF 3.353).
47. Wegerich, F., Giachetti, A., Allegrozzi, M., Lisdat, F., and Turano, P., Mechanistic insights into the superoxide-cytochrome c reaction by lysine surface scanning, J.Biol.Inorg.Chem., 18, 429-440, 2013 (IF 3.353).
48. Barbet-Massin, E., Pell, A. J., Knight, M. J., Webber, A. L., Felli, I. C., Pierattelli, R., Emsley, L., Lesage, A., and Pintacuda, G., 13C-detected through-bond correlation experiments for protein resonance assignment by ultra-fast MAS NMR, ChemPhysChem ,14, 3131-3137, 2013 (IF 3.349).
49. Luchinat, C., Parigi, G., and Ravera, E., Water and protein dynamics in sedimented systems: a relaxometric investigation, Chem.Phys.Chem. 14, 3156-3161, 2013 (IF 3.349).
50. Fragai, M., Luchinat, C., Parigi, G., and Ravera, E., Practical considerations over spectral quality in solid state NMR spectroscopy of soluble proteins, J.Biomol.NMR, 57, 155-166, 2013 (IF 2.845)
PUBLICATIONS 40

51. Bermel, W., Bruix, M., Felli, I. C., Kumar, V. M. V., Pierattelli, R., and Serrano, S., Improving the chemical shift dispersion of multidimensional NMR spectra of intrinsically disordered proteins, J.Biomol.NMR, 55, 231-237, 2013 (IF 2.845).
52. Bermel, W., Felli, I. C., Gonnelli, L., Kozminski, W., Piai, A., Pierattelli, R., and Zawadzka-Kazimierczuk, A., High-dimensionality 13C direct-detected NMR experiments for the automatic assignment of intrinsically disordered proteins, J.Biomol.NMR, 57: 353-361, 2013 (IF 2.845).
53. Luchinat, C., Parigi, G., and Ravera, E., Can metal ion complexes be used as polarizing agents for solution DNP? A theoretical discussion, J.Biomol.NMR, 2013, epub ahead of print (IF 2.845).
54. Mori, M., De Lorenzo, E., Torre, E., Fragai, M., Nativi, C., Luchinat, C., and Arcangeli, A., A highly soluble matrix metalloproteinase-9 inhibitor for potential treatment of dry eye syndrome, Basic.Clin.Pharmacol.Toxicol., 111, 289-295, 2013 (IF 2.124).
55. Dani, C., Bresci, C., Berti, E., Ottanelli, S., Mello, G., Mecacci, F., Breschi, R., Hu, X., Tenori, L., Luchinat, C., Metabolomic profile of term infants of gestational diabetic mothers, J. Matern. Fetal Neonatal Med. 2013, epub ahead of print (IF 1.518)
56. DeJong, J. T., Soga, K. S., Kavazanjian, E., Burns, S., van Passen, L., Fragaszy, R., Al Qabany, A., Aydilek, A., Bang, S. S., Burbank, M., Caslake, L., Chen, C. Y., Cheng, X., Chu, J., Ciurli, S., Fauriel, S., Filet, A. E., Hamdan, N., Hata, T., Inagaki, S., Jefferis, S., Kuo, M., Larrahondo, J., Manning, D., Martinez, B., Mortensen, B., Nelson, D., Palomino, A., Renforth, P., Santamarina, J. C., Seagren, E. A., Tanyu, B., Tsesarsky, M., and Weaver, T., Biogeochemical processes and geotechnical applications: progress, opportunities and challanges, Geotechnique, 63, 287-301, 2013 (IF 1.461).
57. Cacciatore, S., Piccioli, M., and Turano, P., Electron self-exchange of cytochrome c measured via 13C detected protonless NMR, J.Porphyr.Phthalocyanines, 17, 142-149, 2013 (IF 1.433).
58. Felli, I. C., Piai, A., and Pierattelli, R., Recent advances in solution NMR studies: 13C direct detection for biomolecular NMR applications, Ann.Rep.NMR Spectroscop., 359-418, 2013 (IF 1.424).
Books and book chapters
Banci, L., (Ed.) Metallomics and the Cell, Series: Metal Ions in Life Sciences, Vol. 12, Springer, 2013
Banci, L., Bertini, I.. Metallomics and the Cell: some definitions and general comments, Met Ions Life Sci. 12:1-13, 2013
Bertini, I. and Pierattelli, R., Plastocyanin, Encyclopedia of Metalloproteins, Springer version online (http://www.springer.com/life+sciences/biochemistry+%26+biophysics/book/978-1-4614-1532-9)
Del Conte, R.; Lalli, D.; Turano, P. NMR as a tool to target protein-protein interactions, in Disruption of protein-protein interfaces, Mangani, S., editor; Springer-Verlag: Berlin Heidelberg, 2013; Chapter 4, pp. 83-111. Lalli, D.; Turano, P. Nuclear Magnetic Resonance as a Tool to Characterize the Interactome of Heme Proteins, in Handbook of Porphyrin Science, Ferreira, G. C.; Kadish, K. M.; Smith, K. M.; Guilard, R., editors; World Scientific Publishing (USA): 2013; Vol. 30, part II, Chapter 27, pp. 180-220.
PUBLICATIONS
41

Highligths and acknowledgements of CERM research SOLEIL Synchrotron acknowledgement for research results obtained through data collection on beeline PROXIMA1: http://www.synchrotron-soleil.fr/Soleil/ToutesActualites/2013/PX1-FeS-PNAS Silverman S: F1000Prime Recommendation of [Limongelli V et al., Angew Chem Int Ed Engl 2013, 52(8):2269-73]. In F1000Prime, 09 Apr 2013; DOI: 10.3410/f.717972271.793473734. F1000Prime.com/717972271#eval793473734 http://onlinelibrary.wiley.com/doi/10.1002/cbic.201300379/full and http://f1000.com/prime/717997728: highlights on Banci, L., Barbieri, L., Bertini, I., Luchinat, E., Secci, E., Zhao, Y., and Aricescu, A. R., Atomic-resolution monitoring of protein maturation in live human cells by NMR, Nat.Chem.Biol. , 9, 297-299, 2013
Meetings organized by CERM
12th Edition of Premio Città di Firenze on Molecular Sciences Organized by the Sacconi Foundation. Florence, December 16, 2013 GIDRM - NMR Day on "In-cell NMR analysis of biomolecular structure and function", December 6, 2013 Instruct Training Course "In-cell NMR analysis of biomolecular structure and function", CERM, University of Florence - Italy, December 2-6, 2013 Science Europe, LEGS Committee Meeting, CERM, University of Florence - Italy, November 4-6, 2013 WeNMR and INSTRUCT Practical Course on "Advanced methods for the integration of other structural data with NMR data", CERM, University of Florence - Italy, January 21-25, 2013
Seminars and Reserch Seminars (RSS) held at CERM January 29th 2013 at 6:00 p.m. - Prof. Jesús Jiménez-Barbero (Chemical and Physical Biology - CIB-CSIC, Madrid, Spain) - "Carbohydrate-Protein Interactions. A 3D view by using NMR." January 31st 2013 at 12:00 p.m. - Prof. Maurizio Brunori (Department of Biochemical Sciences - University of Rome La Sapienza) - "Morphogenesis of a protein: folding pathways and energy landscapes"
MEETINGS
42

March 8th 2013 at 12:00 p.m. - Letizia Barbieri (RSS) “Molecular Biology Techniques for Protein Expression” March 15th 2013 at 12:00 p.m. – Lucio Ferella (RSS) “Computational Grid pipeline for the paramagnetic systems” March 22nd 2013 at 12:00 p.m. - Tomáš Hošek (RSS) “Viral IDPs & NMR methodology” March 29th 2013 at 12:00 p.m. – Mauro RInaldelli (RSS) “Software tools for protein structural characterization via paramagnetic NMR data” April 5th 2013 at 12:00 p.m. - Eduardo Oliveira Calçada (RSS) “The wonderful world of IDPs:E7, E1A and CBP/p300” April 11th 2013 at 12:00 p.m. - Magdalena Korsak (RSS) “Insight into beta amyloid protein” April 18th 2013 at 12:00 - p.m. Yana Valasatava (RSS) “MetalS3: discovering the structural neighborhoods of Minimal Functional Sites” April 24th 2013 at 6:00 p.m. - Dr. Hadas Amit (BioTalentum Ltd., Gödöllõ, Hungary ) - "From human stem cells to intrinsically disordered proteins" May 9th 2013 at 6:00 p.m. – Carmelo Fuccio (RSS) New approach for isotopic enrichment of proteins” May 16th 2013 at 6:00 p.m. – Alessandro Piai (RSS) “Multidimensional 13C direct-detected NMR experiments for the sequence specific assignment of IDPs” May 23rd 2013 at 6:00 p.m. – Claudio Santucci (RSS) “Metabolomics in nutrition research: CHANCE project” May 30th 2013 at 6:00 p.m. – Erica Secci (RSS) “Protein expression in mammalian cells for in-cell NMR” June 28th 2013 at 12:00 – Gianluca Gallo (RSS) “Beta amyloid peptides in Alzheimer’s disease” July 4th 2013 at 6:00 p.m. - Prof. Greg B. Fields (Torrey Pines Institute for Molecular Studies Port St. Lucie, FL, USA) - "Progress in Selective Matrix Metalloproteinase Inhibition" July 12th 2013 at 5:45 p.m. – Serena Lorenzini (RSS) “Loss and gain of metal binding in superfamilies of homologous proteins” October 2nd, 2013 at 12:00 p.m. - Prof. Tullio Pozzan (Dept. of Biological Sciences - University of Padua and Director of the Department of Biomedical Sciences of CNR) - "Spying live cells in action: a view from inside organelles" December 2nd 2013 6:00 p.m. - Prof. Jan-Eric Litton (Dept. of Medical Epidemiology and Biostatistics Karolinska Institutet Stockholm, Sweden) - "Biobanks and biological research in Europe" M
EETINGS
43