scavenging in antarctica: intense variation between sites and seasons in shallow benthic necrophagy
TRANSCRIPT

y and Ecology 349 (2007) 405–417www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolog
Scavenging in Antarctica: Intense variation between sites andseasons in shallow benthic necrophagy
Dan A. Smale ⁎, David K.A. Barnes, Keiron P.P. Fraser, Paul J. Mann, Matt P. Brown
British Antarctic Survey, Natural Environment Research Council, High Cross, Madingley Road, Cambridge, CB3 OET, UK
Received 17 November 2006; received in revised form 7 February 2007; accepted 7 June 2007
Abstract
The response of scavengers to a feeding cue at Adelaide Island, West Antarctic Peninsula was investigated using a baited videocamera system. Fourteen experimental deployments, each lasting 72 h were conducted at two contrasting sites during the winterand summer of 2005. The rate of bait consumption varied between sites but not between seasons, and was low in comparison withstudies at lower latitudes and greater depths. At the Hangar Cove site, the nemertean Parborlasia corrugatus was out-competed atthe bait and displaced by the lysianassid amphipod Cheirmedon femoratus during winter. However, C. femoratus did not feed onthe bait during summer, allowing P. corrugatus to monopolise the feeding opportunity. At the South Cove site the asteroidOdontaster validus dominated the bait in both seasons but sporadic feeding by the nototheniid fish Notothenia coriicepsconsiderably affected consumption rates during two of the six deployments. Scavengers were attracted to the bait in very highnumbers and opportunistic necrophagy seems to be a successful strategy in an environment that is intensely disturbed by ice.© 2007 Elsevier B.V. All rights reserved.
Keywords: Baited camera; Consumption rates; Necrophagy; Scavengers; Southern Ocean
1. Introduction
Primary productivity is extremely seasonal in Ant-arctic waters (El-Sayed, 1988; Whitaker, 1982). Proba-bly as a result, a wide range of primary consumers showstrong seasonality in feeding activity (Barnes and Clarke,1995). A reduced period of activity during winter hasbeen observed in both suspension feeders (includingbryozoans, bivalve molluscs, hydroids and holothurians)and grazers (Barnes and Clarke, 1995; Brockington,2001; Fraser et al., 2002). However, some taxa such asbrachiopods appear aseasonal with respect to feeding by
⁎ Corresponding author. Tel.: +44 1223 221494; fax: +44 1223221259.
E-mail address: [email protected] (D.A. Smale).
0022-0981/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jembe.2007.06.002
making use of resuspended material in winter (Pecket al., 2005). Although some scavenging groups havebeen fairly well described (De Broyer et al., 2001 andreferences therein), there have been few year-roundbehavioural studies on scavengers, even though a highdiversity of Antarctic fauna exhibit necrophagoustendencies (Arntz et al., 1994; Presler, 1986).
Scavenging is clearly a successful strategy in Antarcticshallow waters, perhaps due to a high frequency of icescouring disturbance, which results in high faunalmortality (Brown et al., 2004). Dense aggregations ofscavenging fauna feeding in newly formed iceberg scourshave been widely reported (Dayton et al., 1974; Heineet al., 1991; Slattery and Oliver, 1986; Zamorano et al.,1986). In fact, some of the most widely spread andconspicuous members of nearshore Antarctic communi-ties (e.g. the echinoderms Ophionotus victoriae and

406 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
Odontaster validus) have an unspecialised catholic dietand demonstrate a range of feeding strategies, includingopportunistic scavenging (Dayton et al., 1974; Dearborn,1977; Kellogg et al., 1983;McClintock et al., 1988). It hasrecently been established that the frequency of icescouring is highly seasonal (Smale et al., 2006) and thatdisturbance events may cease entirely during winterwhilst icebergs are ‘locked in’ by seasonal fast ice. Thisreduction in ice-mediated disturbance (and thereforefaunal mortality) during winter is likely to result in areduction in feeding opportunities for scavengers. Thisprediction is unlikely to hold for high latitude locationsinfluenced by anchor ice, as the intensity of anchor icedisturbance peaks during the winter months (Daytonet al., 1969). Although a large number of secondary andtertiary consumers have developed independence fromfluctuations in primary productivity (Arntz et al., 1994),others (including some opportunistic scavengers) dem-onstrate seasonal patterns of feeding strategies (Arnaud,1977). The influence of ice scour on biodiversity (Guttand Piepenburg, 2003; Gutt et al., 1996; Teixido et al.,2004) and recolonisation (Peck et al., 1999) has been amajor focus of Antarctic marine studies in recent years,but the responses of the scavenging guild to damaged ordecaying benthos are still poorly understood.
Underwater camera technology has become a usefultool in the investigation of scavenger responses to afeeding cue. The vast majority of such studies have beenundertaken in a relatively deep water (Collins et al.,1999; Jones et al., 1998; Premke et al., 2006; Yau et al.,2002) or in areas of high fishing intensity where
Fig. 1. Recording equipment used to monitor scave
scavenging opportunities have been increased anthro-pogenically through fishery discards (Bozzano andSarda, 2002; Groenewold and Fonds, 2000; Ramsayet al., 1997). In the current study we utilised similartechnology to obtain the first recordings of shallowwater scavengers in Antarctica, both beneath winter fastice and during the ice free summer season. The studywas carried out to test two hypotheses; firstly, thatconsumption rates of bait differ between summer andwinter and secondly, that patterns of scavenger arrivaland displacement at the bait will be influenced by theseasonality of the Antarctic environment.
2. Methods
2.1. Experimental protocol
The response of shallow water scavengers to bait wasrecorded during the austral summer and winter of 2005at two contrasting study sites at Adelaide Island, WestAntarctic Peninsula (WAP). Scavenging activity wasrecorded using an underwater video camera (KM UKcolour zoom camera, model OE1366MKII) connectedto a power source and a time-lapse recorder (1 frame/s)positioned on the adjacent shore (Fig. 1). Experimentaldeployments were constantly illuminated with anunderwater lamp (KM UK model OE11-135) set tohalf power (∼150 W). SCUBA divers positioned thecamera at water depths of 12–14 m and recordingcontinued for 72 h following bait placement. The baitwas positioned centrally within a 0.25 m−2 quadrat to
nging activity at Adelaide Island, Antarctica.
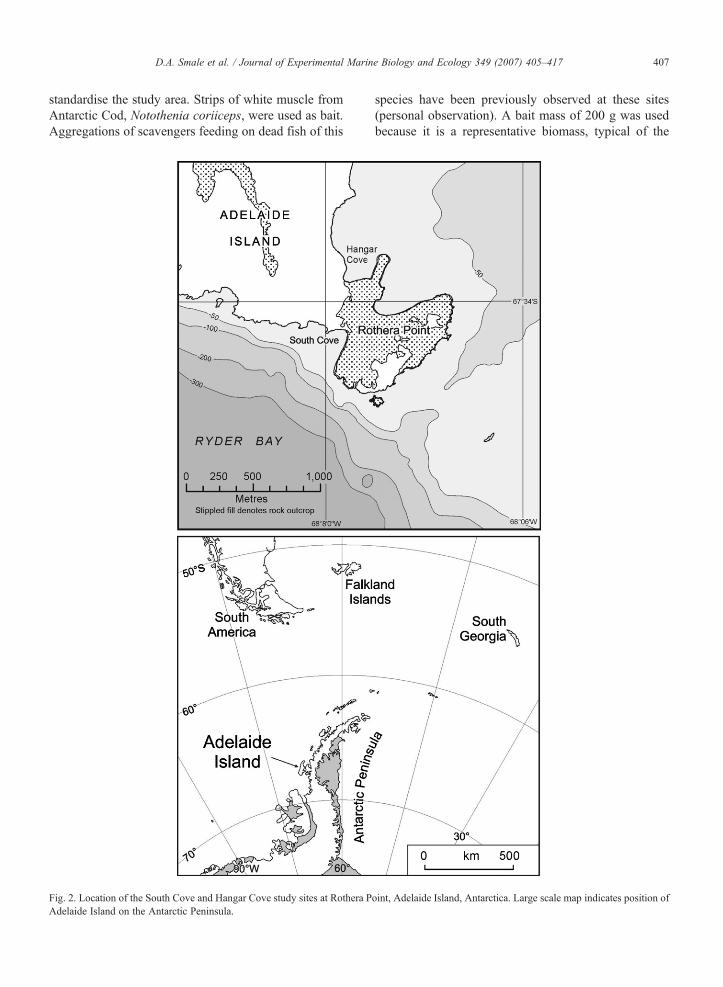
407D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
standardise the study area. Strips of white muscle fromAntarctic Cod, Notothenia coriiceps, were used as bait.Aggregations of scavengers feeding on dead fish of this
Fig. 2. Location of the South Cove and Hangar Cove study sites at Rothera PAdelaide Island on the Antarctic Peninsula.
species have been previously observed at these sites(personal observation). A bait mass of 200 g was usedbecause it is a representative biomass, typical of the
oint, Adelaide Island, Antarctica. Large scale map indicates position of
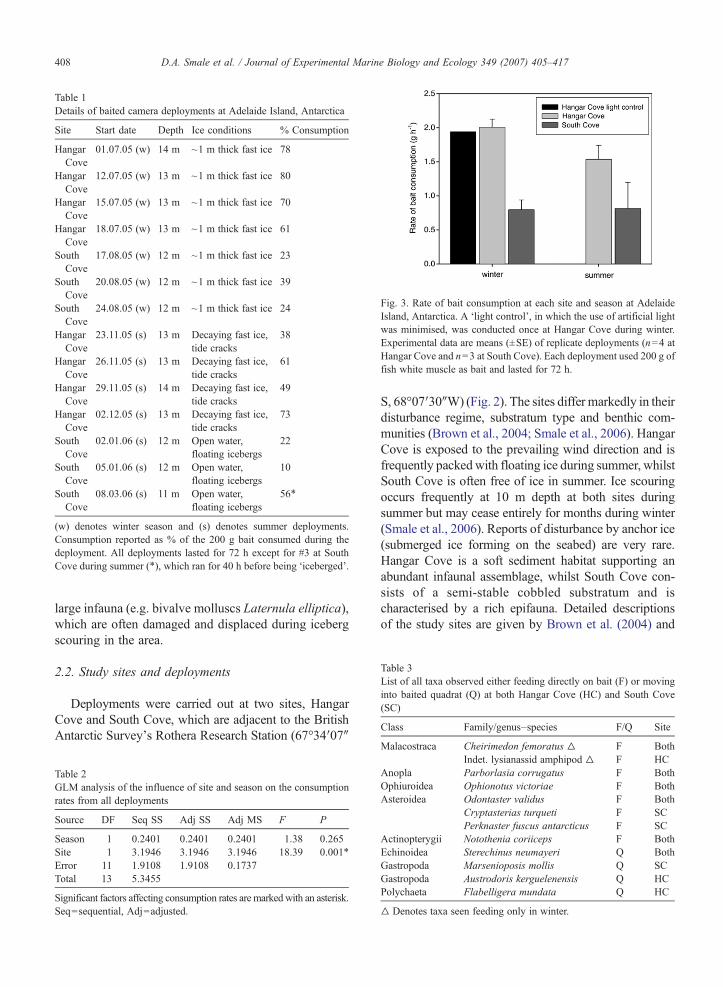
Table 1Details of baited camera deployments at Adelaide Island, Antarctica
Site Start date Depth Ice conditions % Consumption
HangarCove
01.07.05 (w) 14 m ~1 m thick fast ice 78
HangarCove
12.07.05 (w) 13 m ~1 m thick fast ice 80
HangarCove
15.07.05 (w) 13 m ~1 m thick fast ice 70
HangarCove
18.07.05 (w) 13 m ~1 m thick fast ice 61
SouthCove
17.08.05 (w) 12 m ~1 m thick fast ice 23
SouthCove
20.08.05 (w) 12 m ~1 m thick fast ice 39
SouthCove
24.08.05 (w) 12 m ~1 m thick fast ice 24
HangarCove
23.11.05 (s) 13 m Decaying fast ice,tide cracks
38
HangarCove
26.11.05 (s) 13 m Decaying fast ice,tide cracks
61
HangarCove
29.11.05 (s) 14 m Decaying fast ice,tide cracks
49
HangarCove
02.12.05 (s) 13 m Decaying fast ice,tide cracks
73
SouthCove
02.01.06 (s) 12 m Open water,floating icebergs
22
SouthCove
05.01.06 (s) 12 m Open water,floating icebergs
10
SouthCove
08.03.06 (s) 11 m Open water,floating icebergs
56⁎
(w) denotes winter season and (s) denotes summer deployments.Consumption reported as % of the 200 g bait consumed during thedeployment. All deployments lasted for 72 h except for #3 at SouthCove during summer (⁎), which ran for 40 h before being ‘iceberged’.
Fig. 3. Rate of bait consumption at each site and season at AdelaideIsland, Antarctica. A ‘light control’, in which the use of artificial lightwas minimised, was conducted once at Hangar Cove during winter.Experimental data are means (±SE) of replicate deployments (n=4 atHangar Cove and n=3 at South Cove). Each deployment used 200 g offish white muscle as bait and lasted for 72 h.
Table 3List of all taxa observed either feeding directly on bait (F) or movinginto baited quadrat (Q) at both Hangar Cove (HC) and South Cove(SC)
Class Family/genus–species F/Q Site
408 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
large infauna (e.g. bivalve molluscs Laternula elliptica),which are often damaged and displaced during icebergscouring in the area.
2.2. Study sites and deployments
Deployments were carried out at two sites, HangarCove and South Cove, which are adjacent to the BritishAntarctic Survey's Rothera Research Station (67°34′07″
Table 2GLM analysis of the influence of site and season on the consumptionrates from all deployments
Source DF Seq SS Adj SS Adj MS F P
Season 1 0.2401 0.2401 0.2401 1.38 0.265Site 1 3.1946 3.1946 3.1946 18.39 0.001⁎
Error 11 1.9108 1.9108 0.1737Total 13 5.3455
Significant factors affecting consumption rates are marked with an asterisk.Seq=sequential, Adj=adjusted.
S, 68°07′30″W) (Fig. 2). The sites differ markedly in theirdisturbance regime, substratum type and benthic com-munities (Brown et al., 2004; Smale et al., 2006). HangarCove is exposed to the prevailing wind direction and isfrequently packedwith floating ice during summer, whilstSouth Cove is often free of ice in summer. Ice scouringoccurs frequently at 10 m depth at both sites duringsummer but may cease entirely for months during winter(Smale et al., 2006). Reports of disturbance by anchor ice(submerged ice forming on the seabed) are very rare.Hangar Cove is a soft sediment habitat supporting anabundant infaunal assemblage, whilst South Cove con-sists of a semi-stable cobbled substratum and ischaracterised by a rich epifauna. Detailed descriptionsof the study sites are given by Brown et al. (2004) and
Malacostraca Cheirimedon femoratus ▵ F BothIndet. lysianassid amphipod ▵ F HC
Anopla Parborlasia corrugatus F BothOphiuroidea Ophionotus victoriae F BothAsteroidea Odontaster validus F Both
Cryptasterias turqueti F SCPerknaster fuscus antarcticus F SC
Actinopterygii Notothenia coriiceps F BothEchinoidea Sterechinus neumayeri Q BothGastropoda Marsenioposis mollis Q SCGastropoda Austrodoris kerguelenensis Q HCPolychaeta Flabelligera mundata Q HC
▵ Denotes taxa seen feeding only in winter.

Fig. 4. Graphical representations of bait dominance throughout all experimental deployments at Adelaide Island, Antarctica. Bars represent‘dominance’ periods when the scavenging group contributed to greater than 50% of the total abundance of fauna feeding on bait.
409D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
Bowden (2005). In total, 14 experimental deploymentswere carried out, 4 during both summer and winter atHangar Cove and 3 in both seasons at South Cove.Replicate deployments at the same site were conducted atleast 20 m away from each other to maintain indepen-dence. In addition, control deployments for bait and asingle control deployment for light were conducted. Baitcontrols were normal camera deployments without thebait and were run at each site in both seasons. The lightcontrol was a normal deployment without the use of thelamp, except for 1 min every 4 h to observe the bait. Thiswas conducted at Hangar Cove during winter and, byminimising the use of artificial light, tested for the in-fluence of photo-attraction on the behaviour of scavengersduring the experimental deployments.
2.3. Data generation and analysis
The wet mass (tissue dried) of the remaining bait aftereach 72-hour deployment was measured (±0.1 g) in order
Table 4Maximum feeding abundances of scavengers during both seasons andboth study sites at Adelaide Island, Antarctica
Hangar Cove South Cove
Winter Summer Winter Summer
Amphipods 7000 0 46 0Nemerteans 16 98 0 1Ophiuroids 6 0 26 10Asteroids 1 1 50 16
Amphipod abundances were estimated from the collected bait, whilstall other taxa were quantified from video footage.
to compare consumption rates between seasons and sites.Each deployment generated∼8 h of video footage, whichwas later used to record the time each individual enteredthe quadrat, the residence time of taxa at the bait and thetotal number of animals feeding directly on the bait.However, amphipod abundance could not be estimated inthis way due to the very high numbers of small-sizedindividuals present. The amphipods feeding on the bait atthe end of the experiment were collected and theirabundance estimated. The total number of individuals thatmoved into the quadrat during each deployment was alsorecorded. This facilitated comparisons between experi-mental and bait control deployments and permitted theanalysis of taxa feeding around (rather than on) the baititself (e.g. echinoids, ophiuroids and nudibranch gastro-pods). General linear model (GLM) analysis was used inMinitab 14.0 to test the significance of differences inconsumption rates between sites and seasons. Intra-sitevariation in the maximum feeding abundances ofscavengers between seasons was analysed with one-wayANOVAs. All means are presented±standard error.
Table 5One-way ANOVAs of maximum feeding abundances for eachscavenging group observed during winter and summer (each siteanalysed separately)
Hangar Cove (D.F=7) South Cove (D.F=5)
F P F P
Amphipods 18.58 0.005⁎ 3.89 0.120Nemerteans 110.66 0.000⁎ 1.00 0.374Ophiuroids 5.42 0.059 2.21 0.211Asteroids 0.00 1.000 16.53 0.015⁎
Significant seasonal differences are marked with an asterisk.
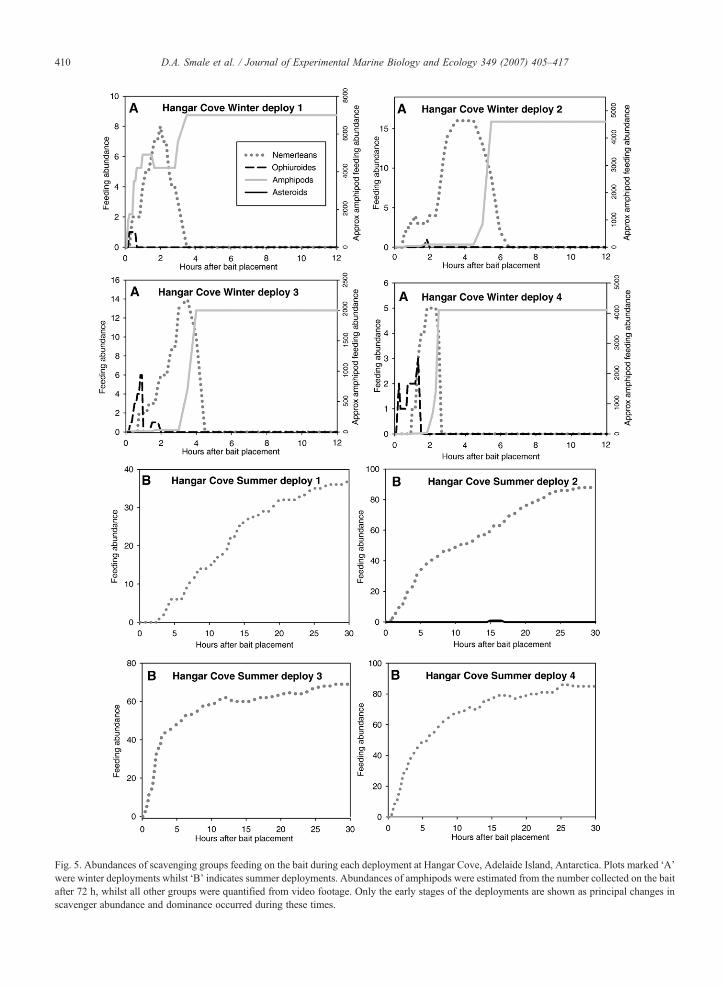
Fig. 5. Abundances of scavenging groups feeding on the bait during each deployment at Hangar Cove, Adelaide Island, Antarctica. Plots marked ‘A’were winter deployments whilst ‘B’ indicates summer deployments. Abundances of amphipods were estimated from the number collected on the baitafter 72 h, whilst all other groups were quantified from video footage. Only the early stages of the deployments are shown as principal changes inscavenger abundance and dominance occurred during these times.
410 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
411D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
3. Results
3.1. Consumption rates
In every experimental deployment scavengers wereobserved feeding and a reduction in the mass of bait wasrecorded (Table 1). The highest rate of bait consump-tion occurred at Hangar Cove during winter (80% or2.2 g h−1) and the lowest at South Cove during summer(10% or 0.3 g h−1). The mean rate of consumption atHangar Cove (winter=2.0±0.1 g h−1, summer 1.5±0.2 g h−1) was about double that of South Cove(winter=0.8±0.1 g h−1, summer 0.8±0.3 g h−1) in bothseasons. The rate of consumption was significantlyhigher at Hangar Cove, but seasonal differences, bothoverall and within site, were non-significant (Table 2).The rate of bait consumption during the ‘light control’was 1.9 g h−1 (compared with an experimental mean of2.0 g h−1) (Fig. 3) and similar patterns of taxa arrival wererecorded. This would suggest the constant light sourcerequired for the experimental deployments had anegligible effect on consumption rates and the behaviourof the scavenging guild at this site. However, due tologistical constraints, this control was only conductedonce and therefore we cannot reject the possibility ofphoto-attraction influencing scavenger behaviour duringthe baited deployments. A total of 8 taxa were recordedactively feeding on the bait and a further 4 taxa were
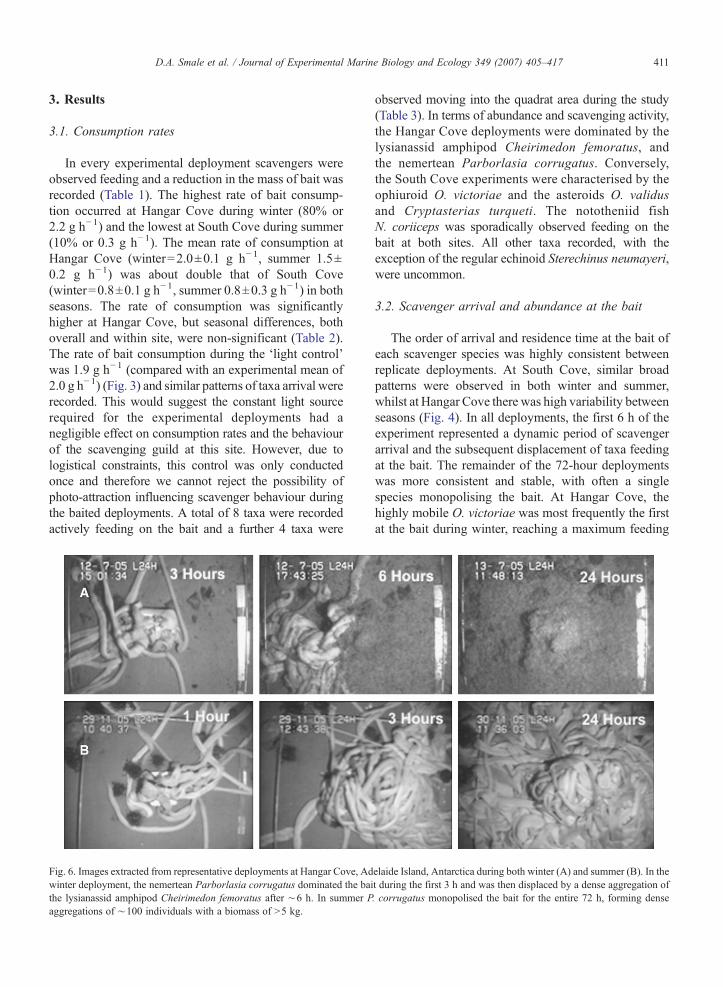
Fig. 6. Images extracted from representative deployments at Hangar Cove, Adwinter deployment, the nemertean Parborlasia corrugatus dominated the baithe lysianassid amphipod Cheirimedon femoratus after ∼6 h. In summer Paggregations of ∼100 individuals with a biomass of N5 kg.
observed moving into the quadrat area during the study(Table 3). In terms of abundance and scavenging activity,the Hangar Cove deployments were dominated by thelysianassid amphipod Cheirimedon femoratus, andthe nemertean Parborlasia corrugatus. Conversely,the South Cove experiments were characterised by theophiuroid O. victoriae and the asteroids O. validusand Cryptasterias turqueti. The nototheniid fishN. coriiceps was sporadically observed feeding on thebait at both sites. All other taxa recorded, with theexception of the regular echinoid Sterechinus neumayeri,were uncommon.
3.2. Scavenger arrival and abundance at the bait
The order of arrival and residence time at the bait ofeach scavenger species was highly consistent betweenreplicate deployments. At South Cove, similar broadpatterns were observed in both winter and summer,whilst at Hangar Cove there was high variability betweenseasons (Fig. 4). In all deployments, the first 6 h of theexperiment represented a dynamic period of scavengerarrival and the subsequent displacement of taxa feedingat the bait. The remainder of the 72-hour deploymentswas more consistent and stable, with often a singlespecies monopolising the bait. At Hangar Cove, thehighly mobile O. victoriae was most frequently the firstat the bait during winter, reaching a maximum feeding
elaide Island, Antarctica during both winter (A) and summer (B). In thet during the first 3 h and was then displaced by a dense aggregation of. corrugatus monopolised the bait for the entire 72 h, forming dense

412 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
abundance of 6 individuals, but it was not observedfeeding at this site during summer. In winter thenemertean P. corrugatus consistently dominated (interms of biomass and abundance) the bait for the first 2–3 h of experiments, forming feeding aggregationsconsisting of up to 16 individuals (Table 4). Theamphipod population increased dramatically after 2–5 h, which coincided with the nemerteans retreating fromthe bait. The amphipod population, dominated byC. femoratus (over 99%), monopolised the bait for the
Fig. 7. Abundances of scavenging groups feeding on the bait during each depwere winter deployments whilst ‘B’ indicates summer deployments. Scavendeployments are shown as principal changes in scavenger abundance and domfeeding by Antarctic Cod, Notothenia coriiceps, are indicated by ‘F.D’ (fish
remainder of the experiment in all four experimentaldeployments (Fig. 4). In contrast, ophiuroids and amphi-pods were completely absent in the summer deploymentsat Hangar Cove, whilst P. corrugatus numbers increasedto form feeding aggregations of nearly 100 individuals(Table 4). A one-way ANOVA of the maximum numberof each scavenging species observed feeding duringsummer and winter shows that C. femoratus was sig-nificantly more abundant in winter, whilst P. corrugatuswas present in significantly higher numbers during
loyment at South Cove, Adelaide Island, Antarctica. Plots marked ‘A’gers were quantified from video footage. Only the early stages of theinance occurred during these times. Periods of intensive and disruptivedisturbance).
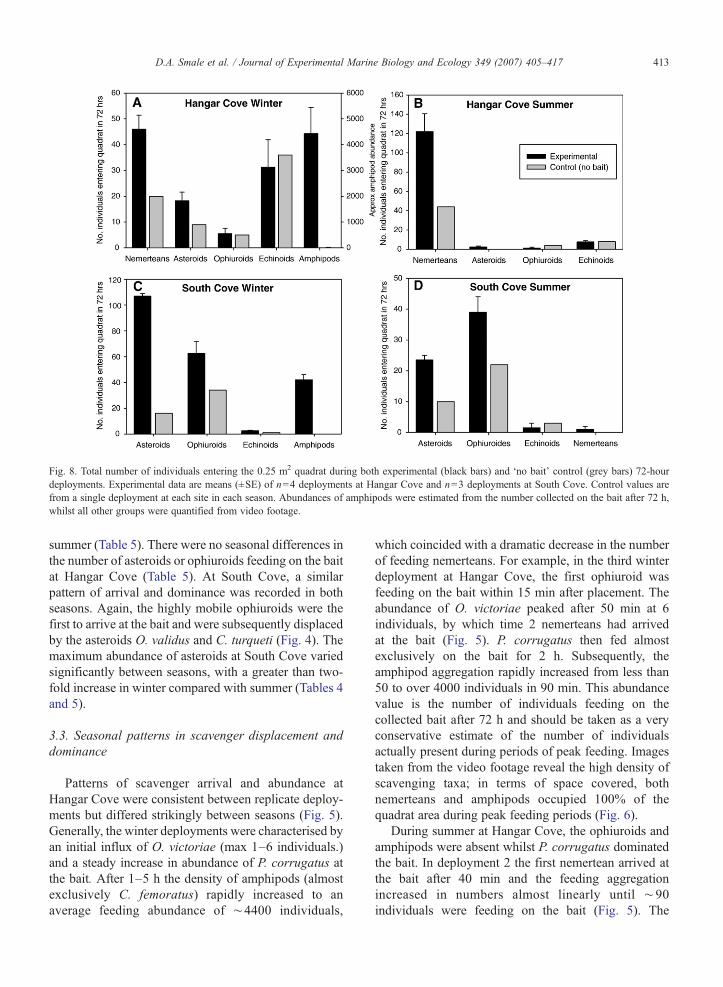
Fig. 8. Total number of individuals entering the 0.25 m2 quadrat during both experimental (black bars) and ‘no bait’ control (grey bars) 72-hourdeployments. Experimental data are means (±SE) of n=4 deployments at Hangar Cove and n=3 deployments at South Cove. Control values arefrom a single deployment at each site in each season. Abundances of amphipods were estimated from the number collected on the bait after 72 h,whilst all other groups were quantified from video footage.
413D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
summer (Table 5). There were no seasonal differences inthe number of asteroids or ophiuroids feeding on the baitat Hangar Cove (Table 5). At South Cove, a similarpattern of arrival and dominance was recorded in bothseasons. Again, the highly mobile ophiuroids were thefirst to arrive at the bait and were subsequently displacedby the asteroids O. validus and C. turqueti (Fig. 4). Themaximum abundance of asteroids at South Cove variedsignificantly between seasons, with a greater than two-fold increase in winter compared with summer (Tables 4and 5).
3.3. Seasonal patterns in scavenger displacement anddominance
Patterns of scavenger arrival and abundance atHangar Cove were consistent between replicate deploy-ments but differed strikingly between seasons (Fig. 5).Generally, the winter deployments were characterised byan initial influx of O. victoriae (max 1–6 individuals.)and a steady increase in abundance of P. corrugatus atthe bait. After 1–5 h the density of amphipods (almostexclusively C. femoratus) rapidly increased to anaverage feeding abundance of ∼4400 individuals,
which coincided with a dramatic decrease in the numberof feeding nemerteans. For example, in the third winterdeployment at Hangar Cove, the first ophiuroid wasfeeding on the bait within 15 min after placement. Theabundance of O. victoriae peaked after 50 min at 6individuals, by which time 2 nemerteans had arrivedat the bait (Fig. 5). P. corrugatus then fed almostexclusively on the bait for 2 h. Subsequently, theamphipod aggregation rapidly increased from less than50 to over 4000 individuals in 90 min. This abundancevalue is the number of individuals feeding on thecollected bait after 72 h and should be taken as a veryconservative estimate of the number of individualsactually present during periods of peak feeding. Imagestaken from the video footage reveal the high density ofscavenging taxa; in terms of space covered, bothnemerteans and amphipods occupied 100% of thequadrat area during peak feeding periods (Fig. 6).
During summer at Hangar Cove, the ophiuroids andamphipods were absent whilst P. corrugatus dominatedthe bait. In deployment 2 the first nemertean arrived atthe bait after 40 min and the feeding aggregationincreased in numbers almost linearly until ∼90individuals were feeding on the bait (Fig. 5). The

414 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
biomass of P. corrugatus feeding on the bait at the endof the deployment was over 5 kg and the aggregationcompletely covered the 0.25 m−2 study area (Fig. 6). AtSouth Cove, all deployments were dominated by theasteroids O. validus and (in lesser numbers) C. turqueti(Fig. 7). The ophiuroid O. victoriae was observedfeeding at the initial stages of most deployments,peaking at 26 individuals feeding in winter and 10 insummer. During the deployments the feeding aggrega-tions of the asteroids steadily increased to ∼50individuals in winter and ∼15 in summer (Fig. 7). Inboth seasons, periods of intensive scavenging byN. coriiceps were observed. Up to 6 fish were recordedfeeding simultaneously, for periods of ∼3 h. Sustainedfeeding by N. coriiceps resulted in a decrease inabundance of invertebrate scavengers on the bait andan above average consumption rate for the deployment.
The ‘bait control’ deployments provided informationon the natural level of invertebrate activity during winterand summer. At Hangar Cove, there was little variationin the total number of asteroids, ophiuroids andechinoids moving into the quadrat whether bait waspresent or absent (Fig. 8A and B). Conversely, theabundance of nemerteans and amphipods increaseddramatically when bait was present. At South Cove, thenumber of echinoids and nemerteans entering thequadrat was similar in baited and non-baited treatmentswhilst the abundance of asteroids, ophiuroids andamphipods seemed more dependent on the presence ofthe bait (Fig. 8C and D).
4. Discussion
The deployment of the baited camera facilitated twoimportant behavioural observations. 1) The shallowwater scavengers at a high Antarctic site, AdelaideIsland, are opportunistic and will feed during winter,resulting in similar consumption rates compared withsummer. 2) The composition of the scavenging guild at asite can vary greatly with site and season andsuperabundant winter fauna may be absent in summer,thus influencing competitive displacement processes atthe food source.
4.1. Multi-seasonal feeding frenzies
Unlike primary consumers, many Antarctic scaven-gers are independent of seasonal fluctuations in primaryproductivity (Arntz et al., 1994) and, perhaps unsur-prisingly, most taxa fed during both winter and summer.At this latitude scavenging opportunities are likely to bemuch reduced in winter because of a reduction in ice
induced mortalities and the absence of many highervertebrates, both of which are potential food sources forinvertebrate scavengers (Jones et al., 1998; Slattery andOliver, 1986). Although many Antarctic organisms havethe physiological capabilities to fast for sustainedperiods, and indeed many do (Brockington et al.,2001; Fraser et al., 2002; Harper and Peck, 2003;Janecki and Rakusa-Suszczewski, 2006), it is clear thatif a food source is present in winter it will be exploitedby scavengers. The rate of consumption recorded duringthis study (a maximum mean of 2.0 g h−1) appears lowif directly compared with similar studies. For example, adolphin carcass in the abyssal Atlantic Ocean can beconsumed at a rate of 400 g h−1 (Jones et al., 1998).Crucially, however, the consumption rate of bait isintimately related to its surface area to volume ratio,which complicates comparisons of the rate of scavengeractivity between studies. The surface area of baitavailable to scavengers in such studies is rarely measured,so a standardisation of consumption rate (g h−1) with themass of bait used (kg) before inter-study comparisons aremade is the best practical solution (although not ideal).Weterm this value ‘bait mass standardised consumption rate’,or Cm. The Cm value for the current Antarctic study was10.0 g h−1 kg−1 whilst the Cm for the dolphin carcass intheAtlanticOcean (Jones et al., 1998)was 6.7 g h−1 kg−1,which suggests a similar rate of scavenging activity. Inshallow Mediterranean waters Cm can reach 81.9 g h−1
kg−1; over 8 times greater than the results of the presentstudy (Bozzano and Sarda, 2002). The only comparableAntarctic experiments were conducted on the SouthGeorgia slope at water depths greater than 600 m;similarly, the Cm value was considerably higher than thepresent study (∼125.0 g h−1 kg−1) (Yau et al., 2002).However, comparable Cm values of 0.6–10.3 g h−1 kg−1
have been recorded in the shallow waters of the Irish Sea(Ramsay et al., 1997).
Comparing consumption rates with other studies isrequired to place the current findings in a wider context,but such comparisons are complicated and should betreated cautiously. For example, the surface area of thebait that is available to scavengers in each study willhave a marked effect on the rate of scavenging activity,and this information is generally not available. Due tosuch limitations there are currently not enough data toinvestigate the large-scale effects of water depth andlatitude on consumption rates. Ultimately, the type ofscavenger inhabiting the study location will be the keyfactor driving consumption rates of carrion. Forexample, in the current study no large decapods (beingabsent from these waters) and very few fish fed on thebait and consumption rates were comparatively low. On

415D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
the South Georgia slope, where consumption rates werehigh, stone crabs and Patagonian toothfish wereabundant and were frequently observed taking largepieces of bait (Yau et al., 2002). Intuitively, the size,number and feeding mechanisms of dominant scaven-gers strongly influence the rate of carrion consumption.
At Adelaide Island, we observed highly contrastingpatterns of scavenger arrival and behaviour betweenstudy sites, which are just 1 km apart. This may beattributable, to some degree, to the backgroundpopulation density of each scavenging species at eachsite (Smale, unpublished data). The two coves differmarkedly in substratum type and exposure to distur-bance (Smale et al., 2006), hence the densities ofsubstrate-specific scavengers (e.g. Cherimedon femor-atus prefers soft bottoms) also varies between the sites.The results suggest that the physical characteristics atthe site of a food-fall play a major role in determiningthe way it is processed, even at small spatial scales.However, at South Cove the asteroid O. validus,dominated the bait in both seasons but, despite alsobeing present at Hangar Cove, individuals wereobserved on the bait only twice. It is unknown whetherthe O. validus population at Hangar Cove is utilising analternative food source or if the high backgrounddensities of nemerteans and amphipods at this siteinfluence its behaviour. In a similar study in the shallowwaters of the Irish Sea, Ramsay et al. (1997) showed theresponses of scavenging species can vary betweenhabitats, despite having similar background densities.
Although consumption rates were comparativelylow, scavenger densities at the bait were high, with thenumber of amphipods, nemerteans and asteroids peak-ing at ∼7000, ∼100 and 50 respectively. Reports ofsimilarly high densities of scavenging crustacea arecommon (Bozzano and Sarda, 2002; Jones et al., 1998;Presler, 1986; Sainte-Marie, 1986; Slattery and Oliver,1986) and Zamorano et al. (1986) recorded similarabundances of asteroids in comparable field manipula-tions. However, the observed feeding aggregation of100 P. corrugatus is very dense, and considerablyhigher than previous field reports of about 20 indivi-duals (Heine et al., 1991; Zamorano et al., 1986).Aggregations of P. corrugatus on (icescour associated)damaged benthos at Signy Island, South Orkneys alsorarely exceed 20 individuals (Barnes personal observa-tion). P. corrugatus is a large (by Antarctic or globalcomparison) nemertean and a voracious predator andscavenger with highly developed sensory organs(Gibson, 1983). The dense aggregations observed atHangar Cove represent a considerable biomass (N5 kg)and energy sink (Heine et al., 1991). The high densities
observed may be partly attributable to the fact thatP. corrugatus has no known predators (Gibson, 1983)and possesses strong chemical defences (Heine et al.,1991).
At South Cove, the number of O. validus feeding insummer was significantly less than during the winterdeployments. Also, at both study sites O. victoriae wasmore common in the early stages of winter experimentscompared with summer deployments. These ubiquitous,omnivorous echinoderms use chemoreception to detectfood items and often form dense aggregations at food falls(Kidawa, 2005). Seasonal necrophagy and high dietplasticity in O. validus was reported by Arnaud (1977),whilst Pearse (1965) found large masses of diatoms inthe stomachs of O. validus collected during summer atRoss Island. At Adelaide Island, small aggregationsof 2–4 individuals feeding on clumps of algae werefrequently observed during summer (personal observa-tion). We suggest that O. validus at Adelaide Island ispreferentially phytophagous during summer, although anumber of individuals will still feed on carrion (in ourstudy, the bait). It is possible that O. victoriae and otherspecies also exhibit a diet shift towards necrophagyduring the winter at this location.
4.2. Where did the super abundant amphipods go insummer?
During winter at Hangar Cove, the nemerteanP. corrugatus quickly arrived at the bait but was subse-quently displaced by a rapidly increasing C. femoratusaggregation. The mechanism of displacement is un-known, but perhaps the movements of thousands ofamphipods prompted the nemerteans to retreat from thebait. However, in the absence of C. femoratus duringsummer the nemerteans were not displaced at the foodsource and they monopolised the bait. The lysianassidamphipod C. femoratus has seasonal patterns of growthand reproduction, linked with the annual cycle ofprimary production (Bregazzi, 1972). Although fre-quently regarded as necrophagous, ovigerous femalesand newly hatched young obligatorily feed on photo-synthetic material (algae) during summer (Bregazzi,1972). We suggest that C. femoratus does not feed oncarrion (as it did not consume any of our bait) duringsummer but instead consumed algal material anddetritus either beneath the deteriorating fast ice or inthe newly formed tide cracks and openings.
Seasonal patterns of the abundance of C. femoratus inthe shallow sublittoral have been reported previously. In acomprehensive study, Presler (1986) deployed a series ofbaited traps along two depth transects in order to describe

416 D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
the necrophagic assemblages of Admiralty Bay, KingGeorge Island. C. femoratus was present in much highernumbers during winter compared with summer in trapsdeployed at 5–30 m depth. Interestingly, Presler (1986)found 5 species of amphipod in the 15mdepth traps duringwinter, whilst only 2 species were collected in the currentstudy. Furthermore, during summer the scavengingassemblage was dominated by the amphipods Hippome-don kergueleni and Orchomene rotundifrons; a starkcontrast to the Adelaide Island deployments whereamphipods were totally absent in summer and asteroidsor nemerteans dominated the bait. The reasons for apparentabsence of H. kergueleni in the soft sediments of HangarCove remain unknown, as this species is a commonburrower with a circum-Antarctic distribution and hasbeen reported from ∼15 m depth (Thurston, 1972).
Jazdzewski et al. (2001) sampled the very shallowsublittoral (i.e. 0.1 m depth) throughout the year andalthough C. femoratus was not an abundant member ofthe amphipod taxocene, small peaks in abundance ofC. femoratus, which lasted for about a month, wereobserved in both winter and summer. Also, Bregazzi(1972) describes specimens collected from 1–5 m depthduring summer that had consumed a variety of plantmaterial. It seems the C. femoratus population at HangarCove, Adelaide Island, exhibits opportunistic seasonalscavenging during winter and perhaps switches itsfeeding preference to plant material and detritus in theshallow sublittoral during early summer. However, with-out conducting feeding preference experiments undercontrolled conditions, it is not possible to conclude thatC. femoratus prefers plant material during summer.Other explanations for its absence at the bait duringsummer include a migration of the population to anothersite or depth and predator avoidance behaviour duringsummer, all of which would require further investigationbefore conclusions can be drawn.
Over ecological time scales the intense scouring ofshallow seabeds is probably the most fundamental factorshaping polar benthic communities. We have shown thatlike many things in Antarctica, polar scavenging is slow.The current study has demonstrated that carrion, which isfrequently created by ice disturbance, is scavenged in apredictable manner but one which is very differentbetween sites and seasons. This variability in carrionprocessing by scavengers not only emphasises thecomplexity of interactions within a seemingly simpleguild, but may also be vital to the survival of some ofAntarctica's most ecologically important benthos. Thenemertean P. corrugatus was consistently displaced (inwinter) by populations of an amphipod, C. femoratus.However, the nemertean can dominate carrion in summer
whenC. femoratus is absent. Like ice scour, scavenging isclearly important to polar benthos, in some wayspredictable and yet strikingly variable in time and space.Decades ago ecological processes were generally thoughtto be simple in Antarctica. Yet a preliminary investigationcovering just two sites and seasons has shown thatinteractions within the scavenger guild are anything butsimple. Uncovering the processes of displacement, therelationships between changes in feeding strategies andenvironmental conditions and trophic links are the nextkey step to understanding even the ‘simple’.
Acknowledgements
Thanks to J. Coldron, and K. Snell for assisting theWinter 2005 dive team above and beyond the call of duty.A. Porter is greatly appreciated for his numerous repairsand modifications to technical equipment. Thanks to L. S.Peck for constructive comments on an early draught of thepaper. [RH]
References
Arnaud, P.M., 1977. Adaptations within the Antarctic marine benthicsystem. In: Llano, G.A. (Ed.), Adaptations Within AntarcticEcosystems. Gulf, Houston, pp. 135–157.
Arntz, W.E., Brey, T., Gallardo, V.A., 1994. Antarctic zoobenthos.Oceanogr. Mar. Biol. Annu. Rev. 32, 251–303.
Barnes, D.K.A., Clarke, A., 1995. Seasonality of feeding activity inAntarctic suspension feeders. Polar Biol. 15, 335–340.
Bowden, D.A., 2005. Quantitative characterization of shallow marinebenthic assemblages at Ryder Bay, Adelaide Island, Antarctica.Mar. Biol. 146, 1235–1249.
Bozzano, A., Sarda, F., 2002. Fishery discard consumption rate andscavenging activity in the northwest Mediterranean Sea. ICES J.Mar. Sci. 59, 15–28.
Bregazzi, P.K., 1972. Life cycles and seasonal movements of Cheiri-medon femoratus (Pfeffer) and Tryphosella kergueleni (Miers)(Crustacea: Amphipoda). Br. Antarct. Surv. Bull. 30, 1–34.
Brockington, S., 2001. The seasonal energetics of the Antarctic bivalveLaternula elliptica (King and Broderip) at Rothera Point, AdelaideIsland. Polar Biol. 24, 523–530.
Brockington, S., Clarke, A., Chapman, A.L.G., 2001. Seasonality offeeding and nutritional status during the australwinter in theAntarcticsea urchin Sterechinus neumayeri. Mar. Biol. 139, 127–138.
Brown, K.M., Fraser, K.P.P., Barnes, D.K.A., Peck, L.S., 2004. Linksbetween the structure of an Antarctic shallow-water communityand ice-scour frequency. Oecologia 141, 121–129.
Collins, M.A., Yau, C., Nolan, C.P., Bagley, P.M., Priede, I.G., 1999.Behavioural observations on the scavenging fauna of thePatagonian slope. J. Mar. Biol. Assoc. U.K. 79, 963–970.
Dayton, P.K., Robbiliard, G.A., Paine, R.T., 1969. Anchor ice formationat McMurdo Sound, Antarctica, and its biological effects. Science163, 273–274.
Dayton, P.K., Robbiliard, G.A., Paine, R.T., Dayton, L.B., 1974.Biological accommodation in the benthic community at McMurdoSound, Antarctica. Ecol. Monogr. 44, 105–128.

417D.A. Smale et al. / Journal of Experimental Marine Biology and Ecology 349 (2007) 405–417
De Broyer, C., Scailteur, Y., Chapelle, G., Rauschert, M., 2001.Diversity of epibenthic habitats of gammaridean amphipods in theeastern Weddell Sea. Polar Biol. 24, 744–753.
Dearborn, J.H., 1977. Food and feeding characteristics of Antarcticasteroids and ophiuroids. In: Llano, G.A. (Ed.), AdaptationsWithinAntarctic Ecosystems. Gulf Publication, Houston, pp. 293–326.
El-Sayed, S.Z., 1988. Productivity of the Southern Ocean: a closerlook. Comp. Biochem. Physiol. 90B, 489–498.
Fraser, K.P.P., Clarke, A., Peck, L.S., 2002. Feast and famine inAntarctica: seasonal physiology in the limpet Nacella concinna.Mar. Ecol. Prog. Ser. 242, 169–177.
Gibson, R., 1983. Antarctic nemerteans: the anatomy, distribution, andbiology of Parborlasia corrugatus (McIntosh, 1876) (Hetero-nemertea, Lineidae). Antarct. Res. Ser. 39, 289–316.
Groenewold, S., Fonds, M., 2000. Effect on scavengers of discards anddamaged benthos produced by the beam-trawl fishery in thesouthern North Sea. ICES J. Mar. Sci. 57, 1395–1406.
Gutt, J., Piepenburg, D., 2003. Scale-dependent impact on diversity ofAntarctic benthos caused by grounding of icebergs. Mar. Ecol.Prog. Ser. 253, 77–83.
Gutt, J., Starmans, A., Dieckmann, G., 1996. Impact of iceberg scouringon polar benthic habitats. Mar. Ecol. Prog. Ser. 137, 311–316.
Harper, E.M., Peck, L.S., 2003. Predatory behaviour and metaboliccosts in the Antarctic muricid gastropod Trophon longstaffi. PolarBiol. 26, 208–217.
Heine, J.N., McClintock, J.B., Slattery, M., Weston, J., 1991. Energeticcomposition, biomass, and chemical defense in the commonantarctic nemertean Parborlasia corrugatus McIntosh. J. Exp.Mar. Biol. Ecol. 153, 15–25.
Janecki, T., Rakusa-Suszczewski, S., 2006. Biology and metabolism ofGlyptonotus antarcticus (Eights) (Crustacea: Isopoda) fromAdmiralty Bay, King George Island, Antarctica. Polar Biosci. 19,29–42.
Jazdzewski, K., De Broyer, C., Pudlarz, M., Zielinski, D., 2001.Seasonal fluctuations of vagile benthos in the uppermost sublittoralof a maritime Antarctic fjord. Polar Biol. 24, 910–917.
Jones, E.G., Collins, M.A., Bagley, P.M., Addison, S., Priede, I.G.,1998. The fate of cetacean carcasses in the deep sea: observationson consumption rates and succession of scavenging species in theabyssal north-east Atlantic Ocean. Proc. Roy. Soc. Lond. B Biol.Sci. 265, 1119–1127.
Kellogg, D.E., Kellogg, T.B., Dearborn, J.H., Edwards, K.C., Fratt, D.B.,1983. Diatoms from brittle star stomach contents: implications forsediment reworking. Antarc. J. U.S. 17, 167–169.
Kidawa, A., 2005. Behavioural and metabolic responses of theAntarctic sea star Odontaster validus to food stimuli of differentconcentration. Polar Biol. 28, 449–455.
McClintock, J.B., Pearse, J.S., Bosch, I., 1988. Population structureand energetics of the shallow-water Antarctic sea star Odontastervalidus in contrasting habitats. Mar. Biol. 99, 235–246.
Pearse, J.S., 1965. Reproductive periodicities in several contrastingpopulations of Odontaster validus Koehler, a common Antarcticasteroid. In: Llano, G.A. (Ed.), Biology of the Antarctic seas II.American Geophysical Union, Washington D.C, pp. 39–85.
Peck, L.S., Brockington, S., Vanhove, S., Beghyn, M., 1999.Community recovery following catastrophic iceberg impacts in asoft-sediment shallow-water site at Signy Island, Antarctica. Mar.Ecol. Prog. Ser. 186, 1–8.
Peck, L.S., Barnes, D.K.A., Willmott, J., 2005. Responses to extremeseasonality in food supply: diet plasticity in Antarctic brachiopods.Mar. Biol. 147, 453–463.
Premke, K., Klages, M., Arntz, W.E., 2006. Aggregations of Arcticdeep-sea scavengers at large food falls: temporal distribution,consumption rates and population structure. Mar. Ecol. Prog. Ser.325, 121–135.
Presler, P., 1986. Necrophagous invertebrates of the Admiralty Bay ofKing George Island (South Shetland Islands, Antarctica). Pol. PolarRes. 7, 25–61.
Ramsay, K., Kaiser, M.J., Moore, P.G., Hughes, R.N., 1997.Consumption of fisheries discards by benthic scavengers: utilisa-tion of energy subsidies in different marine habitats. J. Anim. Ecol.66, 884–896.
Sainte-Marie, B., 1986. Feeding and swimming of lysianassidamphipods in a shallow cold-water bay. Mar. Biol. 91, 219–229.
Slattery, P.N., Oliver, J.S., 1986. Scavenging and other feeding habitsof lysianassid amphipods (Orchomene spp.) from McMurdoSound, Antarctica. Polar Biol. 6, 171–177.
Smale, D.A., Barnes, D.K.A., Fraser, K.P.P., 2006. The influence ofdepth, site exposure and season on the intensity of iceberg scouring innearshore Antarctic waters. Polar Biol. doi:10.1007/s00300-006-0236-0.
Teixido, N., Garrabou, J., Gutt, J., Arntz, W.E., 2004. Recovery inAntarctic benthos after iceberg disturbance: trends in benthiccomposition, abundance and growth forms. Mar. Ecol. Prog. Ser.278, 1–16.
Thurston,M.H., 1972. The crustacea amphipoda of Signy Island, SouthOrkney Islands. British Antarctic Survey Scientific Reports 71.
Whitaker, T.M., 1982. Primary production of phytoplankton off SignyIsland, South Orkney Islands, the Antarctic. Proc. Roy. Soc. Lond.B Biol. Sci. 214, 169–189.
Yau, C., Collins, M.A., Bagley, P.M., Everson, I., Priede, I.G., 2002.Scavenging by megabenthos and demersal fish on the SouthGeorgia slope. Antarct. Sci. 14, 16–24.
Zamorano, J.H., Duarte, W.E., Moreno, C.A., 1986. Predation uponLaternula elliptica (Bivalva, Anatinidae): a field manipulation inSouth Bay, Antarctica. Polar Biol. 6, 139–143.