samba, a xenopus hnrnp expressed in neural and neural … · samba, a xenopus hnrnp expressed in...
TRANSCRIPT

PATTERNS & PHENOTYPES
Samba, a Xenopus hnRNP Expressed in Neuraland Neural Crest TissuesChao Yun Irene Yan,1* Paris Skourides,2 Chenbei Chang,3 and Ali Brivanlou4
RNA binding proteins regulate gene expression at the posttranscriptional level and play important roles inembryonic development. Here, we report the cloning and expression of Samba, a Xenopus hnRNP that ismaternally expressed and persists at least until tail bud stages. During gastrula stages, Samba is enrichedin the dorsal regions. Subsequently, its expression is elevated only in neural and neural crest tissues. In thelatter, Samba expression overlaps with that of Slug in migratory neural crest cells. Thereafter, Samba ismaintained in the neural crest derivatives, as well as other neural tissues, including the anterior andposterior neural tube and the eyes. Overexpression of Samba in the animal pole leads to defects in neuralcrest migration and cranial cartilage development. Thus, Samba encodes a Xenopus hnRNP that isexpressed early in neural and neural crest derivatives and may regulate crest cells migratory behavior.Developmental Dynamics 238:204–209, 2009. © 2008 Wiley-Liss, Inc.
Key words: Xenopus; hnRNP; Samba; Slug; neural crest migration; RNA binding protein (RBP)
Accepted 26 October 2008
INTRODUCTION
RNA binding proteins (RBPs) regulateRNA splicing, stability, nuclear ex-port, subcellular localization andtranslation. They play important rolesduring early metazoan embryogene-sis. One illustrative example is the es-tablishment and maintenance of themRNAs gradient that determines an-terior–posterior axis in Drosophila byRNA binding (Minakhina and Stew-ard, 2005). In Xenopus, several RBPscontaining the heterogeneous ribonu-cleoprotein (hnRNP) domains modu-late early development by means ofspatial restriction of RNA localization.Vg1-RBP/Vera mediates vegetal local-ization of Vg1 RNA, which encodes acritical mesendodermal inducing fac-
tor (Havin et al., 1998, Choo et al.,2005). Similarly, 40LoVe binds to veg-etal localization elements in vegetal-specific RNAs for polarized distribu-tion of these transcripts in Xenopusoocytes (Czaplinski et al., 2005).These RBPs thus participate in ma-ternal determination of animal–veg-etal axis. In addition to setting up theearly embryonic axis, RBPs also regu-late late developmental processes. Forexample, hnRNP Hermes is expressedin the egg and throughout develop-ment, but is also present in the devel-oping heart where its misexpressioninduces cardiac malformations (Ger-ber et al., 2002). Vg1-RBP is presentmaternally and is later restricted toneural folds, eyes, and brachial arches.
In addition to its early role in partition-ing Vg-1 mRNA, Vg1-RBP also modu-lates neural crest migration at late neu-rula stages (Yaniv et al., 2003).
RESULTS
In the course of our expression libraryscreening for genes that regulate earlyXenopus development, we identified aXenopus hnRNP, which was namedSamba after its effect on dissociated an-imal cap cells plated on fibronectin sub-strate. Samba overexpression causesdissociated animal cap cells to oscillateback and forth on the same spot as op-posed to the migratory behavior dis-played by the control cells (not shown).The cDNA for Samba encodes a proteinwith 313 amino acids and a calculated
1Department of Cell and Developmental Biology, Universidade de Sao Paulo, Sao Paulo, SP, Brazil2Department of Biological Sciences, University of Cyprus, Nicosia, Cyprus3Department of Cell Biology, University of Alabama at Birmingham, Birmingham, Alabama4Laboratory of Molecular Vertebrate Embryology, The Rockefeller University, New York, New York*Correspondence to: Chao Yun Irene Yan, Department of Cell and Developmental Biology, Universidade de Sao Paulo, AvProf. Lineu Prestes, 1524, Sao Paulo, SP, 05508-900 Brazil. E-mail: [email protected]
DOI 10.1002/dvdy.21826Published online 17 December 2008 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 238:204–209, 2009
© 2008 Wiley-Liss, Inc.

molecular weight of approximately 34kDa (Fig. 1). Sequence analysis re-vealed several structural features thatare characteristic of hnRNPs—namely,two RNA recognition motifs (RRMs)and a glycine rich-carboxyl terminus(Akindahunsi et al., 2005). RRM con-sists of 80–100 amino acids that arehighly conserved among eukaryotes; itnormally contains consensus RNA-binding sequences designated as RNP1and RNP2, respectively, with other con-served hydrophobic residues inter-
spersed in the motif (Burd and Drey-fuss, 1994; Merrill et al., 1988). InSamba, the first conserved RRM isfound between amino acids 54–108.The second RRM is located between res-idues 138–208 (Merrill et al., 1988).The carboxyl terminal of Samba has aglycine-rich region that is common topreviously characterized hnRNP-A/Bproteins. Although Samba does not con-tain a traditional nuclear localizationsignal, nor is there one present in Xeno-pus laevis hnRNP A1/A2 carboxyl-
terminus (Siomi and Dreyfuss, 1995), itdoes contain a conserved CBFNT do-main at its amino terminus betweenresidues 1–53. The CBFNT domain hasbeen identified in the CARG-bindingfactor A protein as the domain that in-teracts with the promoter of immuno-globulin K (Bemark et al., 1998).
Sequence alignment between Sambaand RNPs from different speciesshowed that Samba is highly ho-mologous to zebrafish hnRNP A/B(AAH48898), chick single strandedD-box binding factor (NP_990659,89% identical), and human hnRNPA/B (AAH09359, 89% identical). Inaddition, we identified two XenopushnRNPs that show significant iden-tity to Samba: BC074212.1, a geneidentified through the Xenopus se-quencing initiative (96% identical;Klein et al., 2002) and BC043814.1(84% identical). The singular differ-ence between Samba and these twohnRNPs is the absence of an amino-terminal insert in Samba, which ispresent in the other two proteins.The high sequence homology be-tween Samba and BC074212.1 sug-gests that they are likely pseudoal-leles in the Xenopus genomegenerated through partial genomicduplication during evolution (Hell-sten et al., 2007).
To examine temporal and spatialexpression of Samba, we performedreverse transcriptase-polymerase chainreaction (RT-PCR) and in situ hybrid-ization. Samba is expressed maternally,its zygotic expression peaks during neu-rula stages and is maintained until atleast tail bud stages (Fig. 2A). Detailedanalysis of Samba distribution at gas-trula stages by tissue dissection showedthat, although Samba is present in allembryonic regions, it is enriched in thedorsal and vegetal domains during gas-trulation (Fig. 2B). Whole-mount in situhybridization revealed that Sambatranscripts are concentrated in the neu-ral plate at early neurula stages (Fig.3B). As development proceeds, Sambais localized to the neural as well as theneural crest tissues. By the time of neu-ral tube closure, Samba is expressedboth in the migratory neural crest andthe neural tube (Fig. 3C). Double in situhybridization for the neural crestmarker Slug indicated that Samba in-deed overlaps with Slug expression inthe neural crest; however, Samba also
Fig. 1. Samba is homologous to hnRNPs. The amino-terminus of the predicted sequence showshigh homology with CBFNT domains (black bar), immediately followed by two RNA recognitionmotifs: RRM1-like (hatched bar) and RRM2-like (white bar).
SAMBA, A XENOPUS NEURAL hnRNP 205

has additional expression in the neuraltissues that is not seen in Slug expres-sion pattern. At tail bud stages, Sambaexpression is maintained in neural andneural crest derivatives, including thebrain, the spinal cord, eyes, and bra-chial arches (Fig. 3F–I).
As Samba is expressed in neuraland neural crest cells, we examinedwhether increased expression ofSamba affected the development ofthese cells. We co-injected Samba withbeta-galactosidase in one blastomerein two-cell stage embryos and exam-ined the expression of neural plate(Sox2, Sox3) and neural crest marker(Slug, Msx1, Twist) expression. We
found that there was no change inSox2, Sox3, Slug, Msx1, and Twist ex-pression when Samba was overex-pressed, suggesting that Samba didnot alter neural plate or neural crestcell fate (Fig. 4). Thereafter, we ana-lyzed the effect of Samba expressionon neural crest behavior. BecauseSamba was expressed in migratingcrest streams, we analyzed neuralcrest migration. At neurula stages,the neural fold from green fluorescentprotein (GFP) -injected embryos wastransplanted isotopically into a redfluorescent protein (RFP) -injectedhost embryo (Borchers et al., 2000). Inthis experimental paradigm, trans-planted cells from control donors in-jected with GFP only showed the char-acteristic streams of migratory neuralcrest (Fig. 5A). In contrast, neuralcrest cells derived from donors thatwere co-injected with Samba and GFPdid not migrate efficiently (83% withabnormal streams; n � 12). Most ofthe cells formed tight clusters aroundthe dorsal neural tube; some of thecells that did migrate out did notreach beyond the ventral region of theeyes (compare Fig. 5B with 5A).Transplantation of GFP-expressingcrest cells into Samba-overexpressinghost embryos resulted in normal mi-gratory patterns (0% with abnormalstreams, n � 12; Fig. 5E), suggestingthat Samba acts cell-autonomously inneural crest cells, but its level in tis-sues through which neural crest mi-grates is not critical for migratory be-havior. Consistent with an inhibitoryeffect of Samba on neural crest migra-tion, we observed that, when Sambawas injected into the animal pole of asingle blastomere of two-cell stage em-bryos, the resulting tadpoles dis-played defects in neural crest-derivedhead cartilage without any obvious al-terations in other embryonic tissues(Fig. 5I). Because overexpression ofSamba did not affect neural crestspecification as seen by marker ex-pression, but clearly affected crest cellmigration, these results suggest thatSamba impairs neural crest migra-tion, which ultimately leads to defectsin the formation of cranial crest deriv-atives.
Other RBPs, such as Vg1-RBP,Hermes, Quaker (Xqua), and YY1,have been shown to affect anteriorneural structure development as well
(Zorn et al., 1997; Gerber et al., 2002;Yaniv et al., 2003; Morgan et al.,2004). Among these, only Vg1-RBPhas been reported previously to affectneural crest migration (Yaniv et al.,2003). However, Vg1-RBP and Sambaact differently on crest development.Whereas overexpression of Samba in-hibits crest migration, it is Vg1-RBPknockdown that has this effect (Yanivet al., 2003). This apparent contradic-tion should be considered in light ofour current knowledge of the role ofRBPs in cell migration. Vg1-RBP, andits homologues ZBP-1 and ZBP-2 areselectively enriched in migratory lead-ing edges where they associate withactin mRNA and are required for itsaccumulation and increased transla-tion (Gu et al., 2002; Oleynikov andSinger, 2003; Leung et al., 2006).These data indicate that Vg1-RBP lev-els and distribution control local cy-toskeletal dynamics during cell migra-tion. In contrast, another family ofRBPs, hnRNP K and E1, play a func-tional role in protein complexes re-sponsible for regulating the initialsteps of cell spreading. In this setting,loss of RBP function increased cellspreading (de Hoog et al., 2004). Thus,it is clear that different RBPs playdistinct roles in cell spreading and mi-gration and alterations in their levelscould affect both phenomena. In thisscenario, we believe that endogenousSamba might act similarly to hnRNPK in regulating neural crest cellspreading required for proper migra-tion.
EXPERIMENTALPROCEDURES
Embryo Manipulations andRNA Preparation
Xenopus embryos were obtained andcultured as previously described (Wil-son and Melton, 1994) and staged ac-cording to Nieuwkoop and Faber.Samba coding sequence was cloned inthe pCS2�� vector, which was linear-ized with AscI and transcribed usingmMessage mMachine kit (Ambion) forRNA synthesis. Samba RNA was in-jected at 200-pg dose into the animalpoles of two-cell stage embryos, withor without co-injection of 500 pg ofGFP RNA.
Fig. 2. Samba is expressed throughout earlydevelopmental stages. A: Reverse transcrip-tase-polymerase chain reaction for Samba withembryos at different stages shows that Sambais a maternal transcript that is expressed untillate tail bud stages. Histone was used as anormalizing control. Numbers indicate the cor-responding Nieuwkoop and Faber stages. B:Samba is enriched in the dorsal and the vegetalregions in stage 11 gastrula embryos. As con-trols for different regions, we used the markersChordin for the dorsal mesoderm, Wnt8 for ven-trolateral mesoderm, Xbra for mesoderm, Vg1for the vegetal region. WE, whole embryo; �RT,samples without reverse transcriptase; An, an-imal pole; Vg, vegetal pole; Do, dorsal meso-derm; Vn, ventral mesoderm.
206 YAN ET AL.

RT-PCR
RT-PCR was performed as previouslydescribed (Wilson and Melton, 1994).The following primers were used inthe PCR reaction: Samba: (U: GATGCT GCG AGT GTG GAT AAG G; D:GTC ATA GCC TGG TCC ATA TCC);ODC (U: CCA AGG CTA AAG TTGCAG; D: AAT GGA TTT CAG AGACCA); Chordin (U: CAG TCA GATGGA GCA GGA TC; D: AGT CCC ATTGCC CGA GTT GC); Wnt-8 (U: GTTCAA GCA TTA CCC CGG AT; D: CTCCTC AAT TCC ATT CTG CG); Xbra(U: GGA TCG TTA TCA CCT CTG; D:GTG TAG TCT GTA GCA GCA); andVg1 (U: CCA TTG CTT AAT CCAAGC; D: GAC CAT ATG TGC CAGTAC).
Whole-Mount In SituHybridization
Whole-mount in situ hybridizationwas performed according to themethod described previously (Har-land, 1991). Double in situ hybridiza-tion was performed with the probeslabeled with digoxigenin-UTP and flu-orescein-UTP, respectively, and thechromogenic reaction was developedsequentially with BM purple andNBT-BCIP. For the in situ hybridiza-tions analysis of the effect of Sambaon neural plate and crest markers,200 pg of Samba was co-injected with100 pg of beta-galactosidase RNA.Beta-galactosidase activity was de-tected with Red-gal (Simonson et al.,
Fig. 3. Samba is expressed in the neural and neural crest derivatives as seen by in situ hybrid-ization. A: Vegetal view of stage 10 embryos with dorsal side to the right. Samba expression isconcentrated in the dorsal region. B: Neurula stage embryo. Samba is present in the neural plateand crest domain. C: Anterior–dorsal view of late neurula stage embryo. Arrows point to the outeredge of the forming migratory crest domain, which expresses Samba. D: Double in situ for Slug andSamba show that Samba overlaps with Slug-expressing migrating streams but is not restricted toneural crest domain. Slug probe was developed with a violet stain and Samba with blue stain. E:Anterior-lateral view of tail bud stage embryo indicates that Samba is expressed in migrating lateralstreams of crest cells (arrow) and optic vesicles. F: Lateral view of tail bud stage embryo showingthat Samba remains in the crest cells, optic vesicle, and now appears in the otic vesicle as well. G:Dorsal view of a late tail bud stage embryo. H,I: Higher magnification of cranial expression inFigures F and G.
Fig. 4. Overexpression of Samba does not affect neural and crest markers. Embryos co-injected with 200 pg of Samba and 100 pg of beta-galactosidase in one blastomere at two-cell stage were processed for in situ hybridization for the neural markers Sox2 and Sox3 and the crest markersSlug, Msx1, and Twist. None of the markers altered their expression pattern in the presence of excess Samba.
SAMBA, A XENOPUS NEURAL hnRNP 207
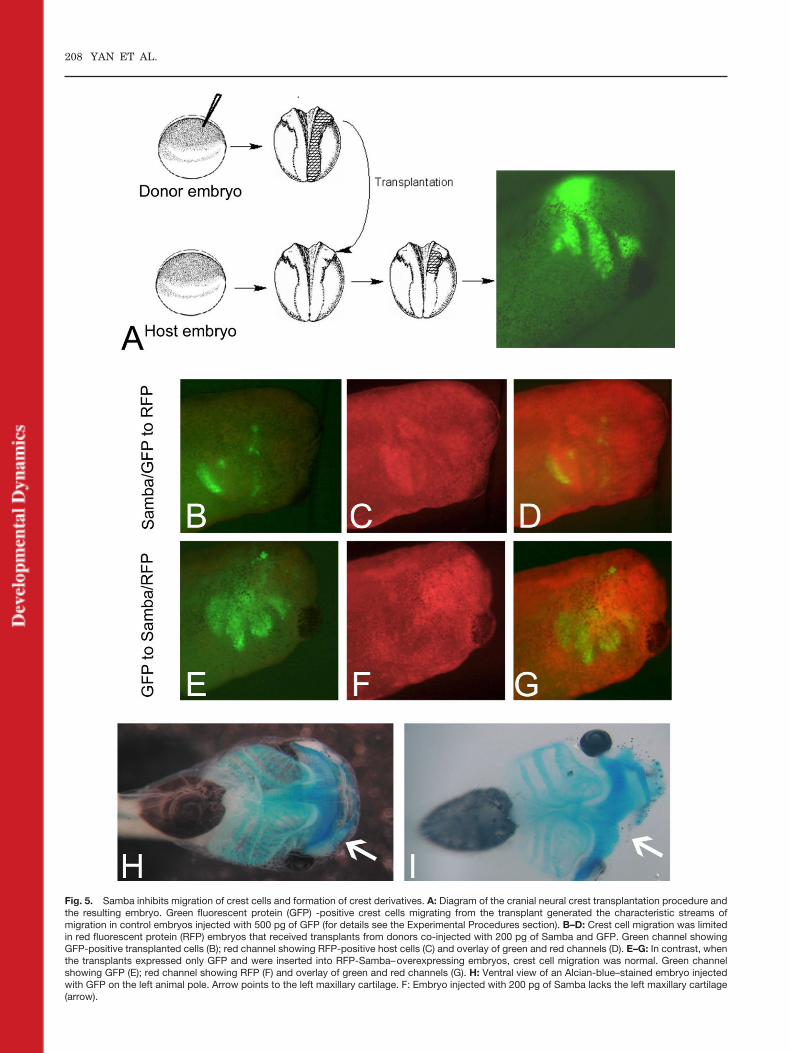
Fig. 5. Samba inhibits migration of crest cells and formation of crest derivatives. A: Diagram of the cranial neural crest transplantation procedure andthe resulting embryo. Green fluorescent protein (GFP) -positive crest cells migrating from the transplant generated the characteristic streams ofmigration in control embryos injected with 500 pg of GFP (for details see the Experimental Procedures section). B–D: Crest cell migration was limitedin red fluorescent protein (RFP) embryos that received transplants from donors co-injected with 200 pg of Samba and GFP. Green channel showingGFP-positive transplanted cells (B); red channel showing RFP-positive host cells (C) and overlay of green and red channels (D). E–G: In contrast, whenthe transplants expressed only GFP and were inserted into RFP-Samba–overexpressing embryos, crest cell migration was normal. Green channelshowing GFP (E); red channel showing RFP (F) and overlay of green and red channels (G). H: Ventral view of an Alcian-blue–stained embryo injectedwith GFP on the left animal pole. Arrow points to the left maxillary cartilage. F: Embryo injected with 200 pg of Samba lacks the left maxillary cartilage(arrow).
208 YAN ET AL.

1995) before whole-mount in situ pro-cedures.
Neural CrestTransplantation
Neural crest transplantation was per-formed using a modified version ofBorchers’ protocol (Borchers et al.,2000). Briefly, the donor embryoswere injected in the animal pole of oneblastomere at two-cell stage with 500pg of GFP RNA and the host embryoswith 200 pg of RFP. To verify the ef-fect of Samba on migrating cells, thedonor embryos were co-injected with200 pg of Samba and GFP, whereas inthe converse experiment, the donorembryos were injected with GFP onlyand the host embryos with Samba andRFP RNA. When the embryos reachedstage 14, the donor embryos were po-sitioned alongside with host siblingsin embryo-sized troughs made of non-toxic modeling clay. The neural foldepidermis of the host embryo waspeeled back and the neural crest re-gion was carefully removed from thehost embryo. The same region was re-moved from the donor embryo andtransplanted into the host embryo.The pliability of the clay allowed us toadjust the borders so as to cover theembryo after transplantation to fixthe transplant in position. The em-bryos were released from the troughsafter 30 min and cultured normallyuntil tail bud or tadpole stages for as-sessment of crest migration and dif-ferentiation.
REFERENCES
Akindahunsi AA, Bandiera A, Manzini G.2005. Vertebrate 2xRBD hnRNP pro-teins: a comparative analysis of genome,mRNA and protein sequences. ComputBiol Chem 29:13–23.
Bemark M, Olsson H, Heinegard D, Lean-derson T. 1998. Purification and charac-terization of a protein binding to the SP6kappa promoter. A potential role forCArG-box binding factor-A in kappatranscription. J Biol Chem 273:18881–18890.
Borchers A, Epperlein HH, Wedlich D.2000. An assay system to study migra-tory behavior of cranial neural crest cellsin Xenopus. Dev Genes Evol 210:217–222.
Burd CG, Dreyfuss G. 1994. Conservedstructures and diversity of functions ofRNA-binding proteins. Science 265:615–621.
Choo S, Heinrich B, Betley JN, Chen Z,Deshler JO. 2005. Evidence for commonmachinery utilized by the early and lateRNA localization pathways in Xenopusoocytes. Dev Biol 278:103–117.
Czaplinski K, Kocher T, Schelder M, SegrefA, Wilm M, Mattaj IW. 2005. Identifica-tion of 40LoVe, a Xenopus hnRNP Dfamily protein involved in localizing aTGF-b-related mRNA during oogenesis.Dev Cell 8:505–515.
de Hoog CL, Foster LJ, Mann M. 2004.RNA and RNA binding proteins partici-pate in early stages of cell spreadingthrough spreading initiation centers.Cell 117:649–662.
Gerber WV, Vokes SA, Zearfoss NR, KriegPA. 2002. A role for the RNA-bindingprotein, hermes, in the regulation ofheart development. Dev Biol 247:116–126.
Gu W, Pan F, Zhang H, Bassell GJ, SingerRH. 2002. A predominantly nuclear pro-tein affecting cytoplasmic localization ofbeta-actin mRNA in fibroblasts and neu-rons. J Cell Biol 156:41–51.
Harland RM. 1991. In situ hybridization:an improved whole-mount method forXenopus embryos. Methods Cell Biol 36:685–695.
Havin L, Git A, Elisha Z, Oberman F, Ya-niv K, Schwartz SP, Standart N, YisraeliJK. 1998. RNA-binding protein con-served in both microtubule- and micro-filament-based RNA localization. GenesDev 12:1593–1598.
Hellsten U, Khokha MK, Grammer TC,Harland RM, Richardson P, Rokhsar DS.2007. Accelerated gene evolution andsubfunctionalization in the pseudotet-
raploid frog Xenopus laevis. BMC Biol5:31.
Klein SL, Strausberg RL, Wagner L, Pon-tius J, Clifton SW, Richardson P. 2002.Genetic and genomic tools for Xenopusresearch: the NIH Xenopus initiative.Dev Dyn 225:384–391.
Leung KM, van Horck FP, Lin AC, Allison R,Standart N, Holt CE. 2006. Asymmetricalbeta-actin mRNA translation in growthcones mediates attractive turning to ne-trin-1. Nat Neurosci 9:1247–1256.
Merrill BM, Stone KL, Cobianchi F, WilsonSH, Williams KR. 1988. Phenylalaninesthat are conserved among several RNA-binding proteins form part of a nucleicacid-binding pocket in the A1 heteroge-neous nuclear ribonucleoprotein. J BiolChem 263:3307–3313.
Minakhina S, Steward R. 2005. Axes for-mation and RNA localization. Curr OpinGenet Dev 15:416–421.
Morgan MJ, Woltering JM, In der RiedenPM, Durston AJ, Thiery JP. 2004. YY1regulates the neural crest-associatedslug gene in Xenopus laevis. J Biol Chem279:46826–46834.
Oleynikov Y, Singer RH. 2003. Real-timevisualization of ZBP1 association withbeta-actin mRNA during transcriptionand localization. Curr Biol 13:199–207.
Simonson MS, LePage DF, Walsh K. 1995.Rapid characterization of growth-arrestgenes in transient transfection assays.Biotechniques 18:434–436.
Siomi H, Dreyfuss G. 1995. A nuclear lo-calization domain in the hnRNP A1 pro-tein. J Cell Biol 129:551–560.
Wilson PA, Melton DA. 1994. Mesodermalpatterning by an inducer gradient de-pends on secondary cell-cell communica-tion. Curr Biol 4:676–686.
Yaniv K, Fainsod A, Kalcheim C YisraeliJK. 2003. The RNA-binding protein Vg1RBP is required for cell migration duringearly neural development. Development130:5649–5661.
Zorn AM, Grow M, Patterson KD, EberoleTA, Chen Q, Artzt K, Krieg PA. 1997.Remarkable sequence conservation oftranscripts encoding amphibian andmammalian homologous of quaking, aKH domain RNA-binding protein. Gene188:199–206.
SAMBA, A XENOPUS NEURAL hnRNP 209