role of the monkey substantia nigra pars reticulata in orienting behaviour and visually triggered...
TRANSCRIPT

Neurosciem'e Let ters , 64 (1986) 109 115 109 Elsevier Scientific Publishers Ireland Lid.
NSL 03755
ROLE OF THE MONKEY SUBSTANTIA NIGRA PARS RETICULATA IN ORIENTING BEHAVIOUR AND VISUALLY TRIGGERED ARM M O V E M E N T S
F. LESTIENNE and P. CAILLIER
I,ahoraloirc d~' Ph.vsiolo~ie Neurosensorielle du ('NRS, 15, rue de l 'Ecole de M&h, cim', F-75270 Pari.~ ("&h'v
06 : b)'am'~, :
tReceivcd August 12th. 1985: Revised version received November 6lh, 1985: Accepted November Sth. 19851
Key u'or,(w substantia nigra pars reticulata monkey orienting behavior arm movement
The role of the substantia mgra pars rcticulata (SNpr) has been studied in the head-free monkey during orienting behaviour in response to visual instruction signals triggering head positioning and conditioncd arm movement. During the behavioural responses we recorded the electromyographic activities of neck muscles and triceps brachii, head movement, horizontal eleclrooculogram and single unit activity of SNpr neurons. Activity of 38 neurons located in the medial part of SNpr were analysed during the visuo-motor task. Forty percent of these units showed a moderate decrease in tonic liring rate during postural prepara- tion preceding the orientation toward eccentric visual signal. This decrease, unrelated with saccadic eye movements per se, was followed by a marked pause observed when the rewarded stimulus was swhched tm and the conditioned arm movement was executed to get the reward. These data suggest that the pause in discharge of thesc SNpr neurons are time locked with hehaviourally relevant visual stimuli and or ap- propriate motor responses.
Earlier studies on the role of the substant ia nigra pars reticulata (SNpr) have lent
s trong support to the theory that the activity o f the SNpr is p redominan t ly related
to oral-face movemen t s [14]. Recently, the visual and o c u l o m o t o r funct ions o f the
monkey SNpr have been studied extensively by Hikosaka and Wurtz [8]. They found
that the activity o f the lateral part of the S N p r is related to visual stimuli and saccadic
eye movements . In the cat similar results were obta ined [9].
E lec t roana tomica l studies o f the rat basal ganglia have suggested that the SNpr
exerts a control on a large spectrum of subcort ical structures which play an irnpor-
tant role in the e labora t ion o f eye and head m o t o r activity involved in or ient ing be-
haviour [3]. Fur thermore , it has been shown that act ivat ion o f non-dopaminerg ic
nigral efferents which originate from the S N p r influences postural and m o t o r mecha-
nisms [6]. Recent neurophysiological studies revealed that a few cells o f the cat SNpr
were related to the active head movemen t and in a few other cells the sensory re-
sponses were modula ted by the subsequent or ient ing behaviour . Similar neuronal ac-
tivity has been observed in the monkey S N p r [10]. These neurons were predominant ly
located in the medial part o f the SNpr. On the basis o f these previous results, the
present study was under taken to establish in the monkey the par t ic ipat ion o f the me-
0304-3940/86/$ 03.50 © 1986 Elsevier Scientilic Publishers Ireland Ltd.

110
A
a
b
C
d
e
I I
k _ . / - -
I
I J
r / - - T -
t
I I f I ~ 1 "1 r l ' l r i , I
i t i I
J F - - - - - ]
h __I 0 0 (D
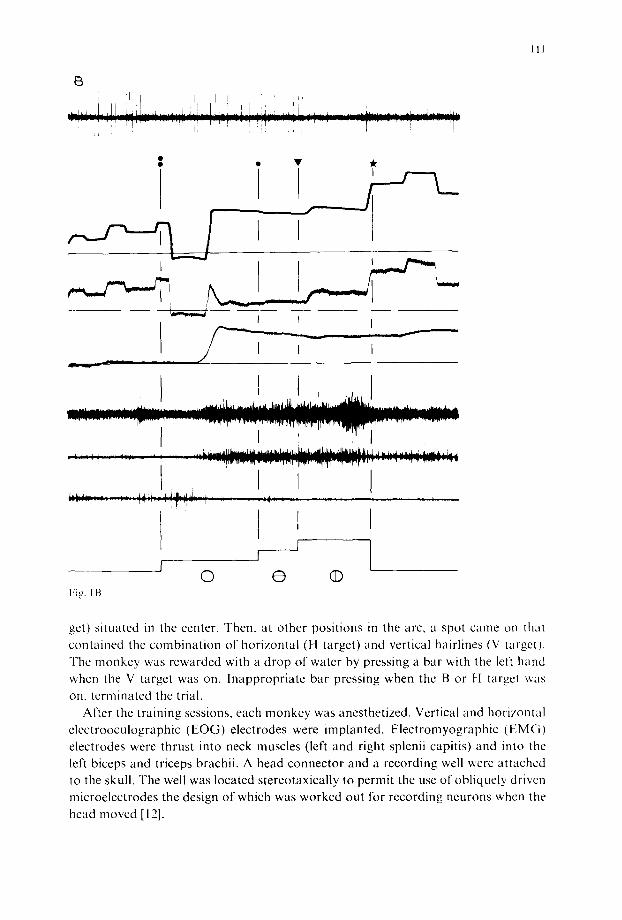
Fig. 1. Behavioral events associated with a 20' right visual target during trigger (A) and predictive (B) eye-head coordination, a, SNpr cell discharge; b, gaze shift; c, horizontal EOG; d, head movement; e g, EMG activity of left triceps brachii, right and left sp!enius capitis; h, sequence of visual targets. Double point, single point and triangle are respectively the onset of B, H and V targets. Star is the onset of bar- pressing. The horizontal lines of the tracings b, c and d represent the central position of gaze, eye and head, respectively. Time scale: 500 ms. Calibration vertical line: 20'L
dial part of the SNpr in orienting behaviour in response to visual instruction signals triggering head positioning and conditioned arm movement.
The experiments were performed on 3 adult rhesus monkeys. The animals were trained to make a visual discrimination between horizontal and vertical hairlines superimposed on a luminous target spot. Target spots were placed on a horizontal perimeter arc. By sequential presentation of target spots we induced the monkeys to make natural, coordinated eye-head movements to direct their gaze at the targets. The target sequence consisted first of the presentation of a blank target spot (B tar-
1 1 1
13
i T "
'
I i i
;'~!;! {
i~iL,
I t , F l I
f I © @ (D
F i g . I B .
I I I
'. ; : : • : I!dl . . . . . , ~ r ~ , , ~ . , r ~ r . , , . ~ r l - r ~ r . , ~ , ~ . , - r r ~ r ~ ~ ~r~, " " , ~ ~.T -w- . ,m, , - -T • r , I.
I
get) situated in the center. Then, at other positions in the arc, a spot came on that contained the combination of horizontal (H target) and vertical hairlines (V target). The monkey was rewarded with a drop of water by pressing a bar with the left hand when the V target was on. Inappropriate bar pressing when the B or H target was
on, terminated the trial. After the training sessions, each monkey was anesthetized. Vertical and horizontal
electrooculographic (EOG) electrodes were implanted. Electromyographic (EMG) electrodes were thrust into neck muscles (left and right splenii capitis) and into tile left biceps and triceps brachii. A head connector and a recording well were attached to the skull. The well was located stereotaxically to permit the use of obliquely driven microelectrodes the design of which was worked out for recording neurons whcn the head moved [12].
112
A • • •
I I . . . . . ~ ° . . . . . ~ • . ° o . . . . . . • . . . . , . . . .
" " ' ' " ° ° " " ' ' ° " " t " . . . . . . . . . . . . . + . . . .
° , . . , . ° . ° ° ~ . . , , . . . . . . I o . . ~ . . . . . ° . . . . . . . , . o . . ° + . . . . ~ . . . . . . . . . • . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . ~ . . . . . . • . . . . . ~ . . . . . . . . . . . • . . . . . . . o . + . . . . . o . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . ~
• . . . o . . ° + . . . . . ° o ° o . . . . ° . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . o . ° . . . . • . . . . . . ° . . . . . . . . . ° .
• . . ° . o ° ~ . . . . . o , , , ° ° . . . . . . . . . . . . . . . . . . . . . . .
• . . . . . . . . . . . ° . . . . . . . . . . . ° ~ . . . . ° . . . . . . . . . . ~ . . . .
B • • • * J t i
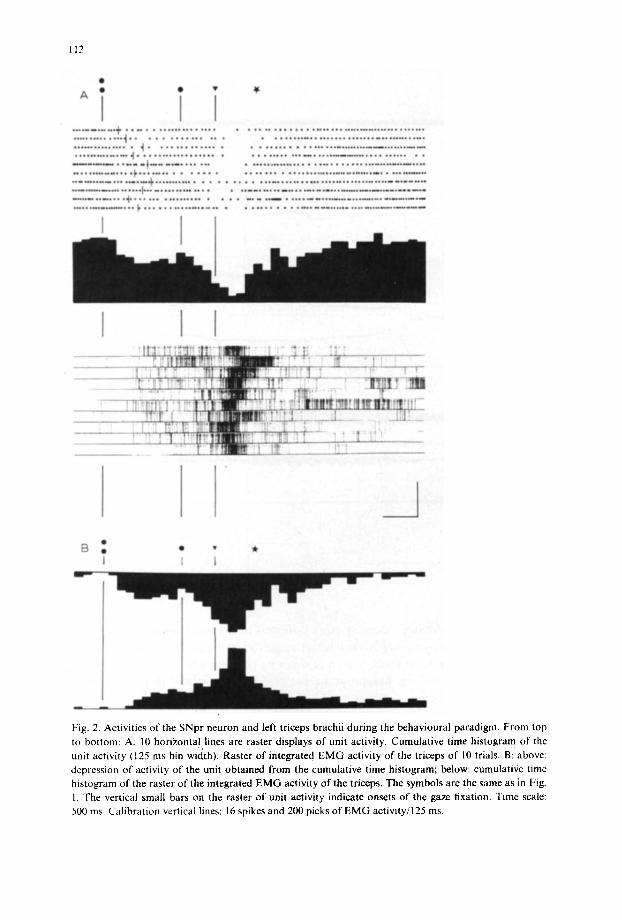
Fig. 2. Activities of the SNpr neuron and left triceps brachii during the behavioural paradigm, From top to bottom: A: 10 horizontal lines are raster displays of unit activity. Cumulative time histogram of the unit activity (125 ms bin width). Raster of integrated E M G activity of the triceps of 10 trials. B: above: depression of activity of the unit obtained from the cumulative time histogram; below: cumulative time histogram of the raster o f the integrated EMG activity of the triceps. The symbols are the same as in Fig. 1. The vertical small bars on the raster of unit activity indicate onsets o f the gaze fixation. Time scale: 500 ms. Calibration vertical lines: 16 spikes and 200 picks of EMG activity/125 ms.

113
The location of the recording tracks and of the small lesions placed at the lowest points of certain tracks were determined histologically.
In this study, electrode penetrations were principally targeted towards the medial part of the SN. In this area the activities of a total of 73 single neurons in 3 monkeys were recorded the positions of which were verified histologically as being in the pars compacta (n=26) and pars reticulata (n=38) . The pars reticulata neurons dis- charged spontaneously with rates frequently as high as 20-40 spikes/s, and the im- pulses were usually shorter than 1 ms. The rate of discharges was not modulated dur- ing spontaneous eye and head movements as long as the monkey was sitting in darkness. In contrasL during the performance of the learnt orienting task, these neu- rons showed a pronounced decrease in activity related to the visual stimuli and motor e v e n t s .
In this report we will describe responses of histologically verified neurons (n = 19) related to specific sensory signals of the paradigm and subsequent orienting behav- iour to those sensory stimuli. Each of these neurons with signal-to-noise ratios of 5:1 was studied extensively for different target locations ( 10 , 20 , 30 left and right) and l\~r each target a minimum of 20 trials was performed. Furthermore, each of these neurons was tested for somatosensory inputs by touching the face, trunk and hand and for vestibular and neck proprioceptive inputs.
Behavioural events associated with the visual targets. Fig. 1 shows the activity of one and the same SNpr neuron during two typical recordings of the syncrgy between eye, head and the conditioned left arm movement triggered by a 20 right target (Fig. IA) and in anticipation of the appearance of the same target (Fig. I B). This ligure is a representative example of the distinct neuromuscular characteristics depending upon the mode (triggered or predictive) of eye head coordination previously de- scribed by Bizzi [1]. Most of the eye head coordinations were found to pertain to the category of the predictive mode (85%). Indeed for each target, a series of l(l move- mcnts vvas induced by the presentation of the H-V target at a fixed location and at regular intervals. However, independent of the mode of coordination a striking simi- larity was observed between the pattern of the EMG activities. The onset o f thc activ- ity of the triceps was time locked with that of the neck agonist activity. We always observed a sustained tonic EMG lasting for the entire duration of the fixation of gaxe on the H-V target (between 0.5 and 1.5 s). This activity is accompanied by the tonic contraction of the biceps. On the basis of previous data, this cocontraction may be related to an obligatory synergy between the flexor and extensor of the arm and also thc neck muscles to stabilize the shoulder and neck joints during the postural prepa- ration of phasic movement and fixation of gaze [11]. Following the tonic activity, the triceps showed a distinct burst of activity, specific to the bar-pressing.
Behavioural correlates ~/neuronal activity. We found that one-half of the 3~ neu- rons located in the medial part of the SNpr were characterized by systematic and significant changes in their activity during the visuo-motor task but not during spon- taneous saccades executed in the dark. A common property of most of these neurons (n = 14) was that their changes in activity were always a decrease in discharge rate and followed the different sequences of the visual signals. The typical responses of

114
this class of neurons are illustrated by the raster display of 10 successive predictive eye-head coordinations (Fig. 2). Rasters of unit activity are aligned with the onset of the B target. The thin vertical lines indicate the time of occurrence of the gaze shift towards the predictive location of a 30 ° right target. The star represents the mean value of the onset of the bar-pressing. The cells are responsive to the onset of the B target. Latencies range from 80 to 300 ms (mean latency: 190 ms). We failed to find a clear correlation between this decrease in response and the onset of the gaze shift which lagged between 120 and 700 ms behind that of the B target. Following this first response the firing rate increased but never resumed the level of the sponta- neous activity. A second and more pronounced decrease was observed in responsc to the H-V target. Latencies were comparable to those of the first response. A final
observation was that the burst of triceps activity was associated with the clear sup- pression of the cell activity.
At this point it is important to stress that changes in the neuronal discharge rate showed a reciprocal relationship with the average E M G activity of the triceps mus- cle displayed as a raster of the integrated EMG. This observation is illustrated in Fig. 2B, which displays: (1) the depression of activity of the neuron shown by the cumulative time histogram of Fig. 2A and (2) the cumulative time histogram pro- duced by the summation of the rasters of integrated EMG. Every unit in this class of SNpr neurons was found to have this profile of depression of activity clearly related to the EMG activity of triceps. In order to assess the extent to which the neu- ronal responses reflected the pattern of EMG activity of triceps, we recorded the activ- ity of the same neurons during free arm extension directed to the bar and performed in no connection with the behavioural paradigm. Only a small percentage of cells responded during these tests. For these few cells the depression of the neuronal activi- ty was considerably reduced (70%) and was seen only during large bursts of EMG activity. We also failed to observe any modulation of neuron activity during active head movement not related to the performance of the learned task. Furthen'nore, when the H-V target was randomly suppressed during the conventional paradigm, correct fixation on the "remembered' location of the target was commonly observed and rewarded. In this paradigm, we observed a reduction of 40-50% in the second depression of the neuron in response to the fixation of the ' remembered' target and the arm movement. When the H-V target was not rewarded, we observed a rapid and total extinction of the response after a few repetitions.
The findings of the present study strongly suggest that a substantial number of neurons in the medial part of the SNpr are concerned with the orienting behaviour underlying conditioned arm movement. We have shown that activity of the SNpr is modulated by visual signals and active movements of the upper limbs. The response of the neurons took the form of a decrease or a pause of a tonic discharge. One of the most striking results was the enhancement of the depression of the activity observed when the visual stimulus was associated with the occurrence of the muscle activity of the arm and not with the onset of the saccade as has been shown in the lateral part of the SNpr by Hikosaka and Wurtz [8]. From anatomical studies reviewed by Chevalier [2] it is known that the targets of the lateral part of the SNpr

115
are mainly the thalamic nucleus ventralis anterior and the intermediate layers of the superior colliculus, while the target of the medial part of the SNpr is the thalamic nucleus ventralis lateralis. In light of these differential anatomical connections and according to the schematic organization of the basal ganglia [5], the present study suggests a different role in visual orienting behaviour for the medial SNpr as com- pared to the lateral. Namely, the medial SNpr should be more involved in postural adjustments during orienting and visually triggered arm movements, whereas the lateral SNpr appears to be mainly concerned with the extraoculomotor system.
In addition to these considerations, an important question is raised by the impor- tance of the behavioural contingencies for occurrence of responses in neurons. In- deed, our results demonstrate clearly that the responses were not necessarily linked to the sensory inputs or motor outputs themselves. On the basis of our tindings and recent results on the lateral part of the SNpr, the putamen and the globus pallidus [5, 7, 8, 13], it has been argued that the responses of many neurons belonging to the striato-pallidal system are mainly 'cognitively' driven.
I Bizzi, E., Kalil, R.E. and Morasso, P., Two modes of active eye head coordination in monke_~s, Brain
Res., 40 (1972) 45 48. 2 ('hevalier, (i., Analyse des relations entre ganglions de la base cl centres moteurs: dtude anatomo-phy-
siologique des projections nigroteetale et nigro-thalamique, Thdse de Doctoral d'Etat, 1985, 62 pp. 3 Chewdier, (i., Vacher, S., Deniau, J.M. and Albe-Fessard, D., Tonic nigral control of tecto spinal
recto diencephalic branched neurons: a possible implication of basal ganglia in orienting bchaviour.
In .I.S. McKenzie, R.E. Kemm and L.N. Wilcock (Eds.). The Basal Ganglia, Plenum, 1984, pp. 247 25~L
4 Dckong, M.R., Crutcher, M.D. and Georgopoulos, A.P., Primate globus pallidus and subthakmnc
nucleus: functional organization, J. Neurophysiol., 53 (I 985) 53{1 543. 5 l)eLong, M.R., Georgopolous, A.P. and Crulcher, M.D., Corlicobasal ganglia relations and coding
of motor performance, Exp. Brain Res., Suppl. 7 (1983) 30 40. 6 l)ichiara, G., Olianas, M., Delfiacco, M., Spano, P.F. and Tagliamonte, A., Intranigral kainic acid
is exidencc that nigral non-dopaminergic neurons control posture, Nature {London), 268 (i977) 743 745.
7 Ii~arts, E.V., Kimura. M., Wurtz, R.tt . and Iilkosaka, O., Behavioural correlates of activitx in basal
ganglia neurons, TINS, II (1984) 447 453, 8 1 likosaka, O. and Wurtz, R.11., Visual and oculomotor functions of monkey substantia nigra pars reti-
culala. 1: Relation of visual and autilory responses to saccades, J. Neurophysiol., 49 (1983) 1230 1253.
9 Joseph, ,I.P. and Boussaoud, D., Role of the cat substantia nigra pars reliculata in eye and head move- ments, 1. Neural activity, Exp. Brain Res., 57 {1985) 45 53.
10 Lestiennc, F. and Caillier, P., Role of the monkey substantia nigra pars reticulata in behavorial rc sponses to visual stimuli, Ncurosci. l.ett., Suppl. 22 (1985) $484.
I 1 Lesticnne, [7.. Vidal, P.P. and Berthoz, A., Gaze changing behaviour in head retrained monkey. Exp.
Brain Res., 53 (1984) 349 356. 12 kes~icnne, F.. Whittington. D.A. and Bizzi, E., Single cell recording from the pontinc reticular lk~rma-
tion in monkey: behaviour of preoculomotor neurons during eye head coordination, In A.F. | :uchs and W. Bcckcr (Eds.), Progress in Oculomotor Research, Proceedings of the International Conference, Schloss Rcisensburg, near Uhn, West Gcrmany, 28-31 July 1980 (Developments in Neuroscience, Vol. 12), EIsevier, 1981,pp, 325 333.
13 Liles, S.g., Activity of neurons in putamen during active and passive movements of wrist, J. Ncuroph_~
sio1.. 53 (1985) 217 236. 14 Mora. F.. Mogenson, G.J. and Rolls, l'i.F.. Activity of neurons in the regiorl of the substanlia nigra
durmg feeding in the monkey. BrainRes. . 133(1977) 267 276.