role of enterobactin and intracellular iron in cell lethality during near-uv irradiation in...
TRANSCRIPT

Photochemistry and Photobiology, 1996, 64(3): 537-541
Research Note
Role of Enterobactin and lntracellular Iron in Cell Lethality During Near-UV Irradiation in Escherichia coli
J. Hoerter”, A. Pierce2, C. Troupe’, J. Eppersonl and A. Eisenstark3 ’Department of Biological Sciences, Ferris State University, Big Rapids, MI, USA; 2Department of Natural Sciences, Stephens College, Columbia, MO, USA and 3Cancer Research Center, Columbia, MO, USA
Received 17 January 1996; accepted 3 June 1996
ABSTRACT In Escherichia coli, fur mutants that constitutively ex- press their native iron chelating agent, enterobactin, are significantly more sensitive to near-UV radiation (NUV) than wild type. An entA mutant, which is incapable of synthesizing enterobactin, is equal to wild type in resis- tance to NUV irradiation. However, the addition of Fe+3 enterobactin but not Al’j enterobactin to entA cell sus- pensions just prior to irradiation results in an increased sensitivity to NUV irradiation. A fes mutant, which is unable to reduce and release iron from enterobactin, is significantly more sensitive to NUV irradiation than wild type. The addition of nontoxic levels of H202 (5 pM) just prior to irradiation significantly increases sensitivity of both fur and fes mutants. These results suggest that one mechanism by which NUV irradiation leads to cell le- thality is by creating a transient iron overload, producing very favorable conditions for the production of highly deleterious free radicals through a variety of mechanisms that lead to oxidative stress and DNA damage including lethal and mutagenic lesions. These results are consistent with the hypothesis that enterobactin is an endogenous chromophore for NUV and contributes to cell lethality via the destruction of its ligand, releasing Fe+2 into the cytoplasm to catalyze the production of highly reactive hydroxyl radicals and other toxic oxygen species via the Haber-Weiss reaction.
INTRODUCTION There is increasing concern that additional solar near-UV light (NUV,? 29WOO nm) may reach the earth’s surface as a result of the depletion of the global stratospheric ozone
*To whom correspondence should be addressed at: Department of Biological Sciences, College of Arts and Sciences, 808 Campus Drive, Ferris State University, Big Rapids, MI 49307-2225, USA. Fax: 616-592-2627; e-mail: [email protected].
tAbbreviutions: LB, Luria-Bertani broth; NUV, near UV light 290- 400 nm; SOD, superoxide dismutase; UVA, 3 2 0 4 0 0 nm radia- tion; UVB, 290-320 nm radiation.
0 1996 American Society for Photobiology 003 1-R655/96 $5.00+0.00
shield. During NUV irradiation of both prokaryotic and eu- karyotic cells, it appears that a large and varied number of endogenous photosensitizers are involved, some of which may only act as intermediates in conveying the incident en- ergy to important cellular molecules such as DNA (1). It is well established that NUV irradiation can cause cell death and mutations, but the mechanisms involved are complex and are not yet clearly identified (2). Previous studies sug- gest that NUV light exerts toxic effects on Escherichia coli via oxidative mechanisms, and toxic oxygen radical photo- products are strongly implicated in NUV-induced lethality (3). Multiple lines of evidence implicate NUV as a generator of excited states of oxygen via endogenous chromophores acting as photosensitizers (4). The NUV-induced lethality is oxygen dependent ( 5 ) , and several active oxygen interme- diates are generated by NUV irradiation of various biologi- cal molecules (6). Further evidence that NUV irradiation generates active oxygen species comes from studies utilizing Salmonella ryphimurium lacking the oxidative defense regu- Ion (AoxyR); these strains are hypersensitive to NUV irra- diation (7). The nature of the intermediate reactive species involved is not identified, but singlet oxygen and hydroxyl radicals have been implicated (4). The generation of super- oxide radicals during NUV irradiation is suggested by stud- ies showing increased NUV sensitivity and mutation rate in E. coli strains deficient in superoxide dismutase (SOD) (8).
The finding that H,O, is a photoproduct of NUV (9) has important implications in vivo because naturally occurring iron complexes may react with H,02 and generate highly reactive hydroxyl radicals via a superoxide-driven Fenton reaction capable of damaging DNA and other targets by a variety of mechanisms (10,l l) . Because ferrous iron is re- leased from ferritin by UVA (320-400 nm) (12,13) and UVB (290-320 nm) (14). this suggests the possibility that enterobactin, the native iron-chelating agent in E. coli, may be a chromophore for NUV, leading to the photoreduction and release of intracellular iron to catalyze free radical gen- eration. Furthermore, the superoxide anion (a possible pho- toproduct of NUV (8)) has also been shown to reduce ferritin (1516). Recent studies utilizingfir and SOD mutants (sodA so&) suggest superoxide-mediated iron reduction is respon- sible for creating those conditions that lead to oxidative
537
538 J. Hoerter et al.
stress and DNA damage including lethal and mutagenic le- sions (17). Because the Fenton reaction is driven by the con- tinual reduction of ferric iron to the ferrous state by super- oxide anion, these findings suggest that intracellular iron pools may have a major influence on the generation of active oxygen species during NUV irradiation, contributing to cell lethality.
Under normal physiological conditions, the level of intra- cellular iron in E. coli is controlled in part by the manufac- ture, secretion and transport of its native iron-chelating agent, enterobactin. Perhaps about 30 genes of the E. coli chromosome are involved in iron uptake, and all are in some way regulated by the ferric uptake regulator Vur) (18). Transport of ferric enterobactin through the cell envelope requires an outer membrane receptor, product of fepA gene ( I 9). The biosynthesis of enterobactin requires the products of at least seven biosynthetic genes, one of which is entA (20). Intracellular release and reduction of iron from entero- bactin requires the product of thefes gene (21). The final destination of iron after it is released from enterobactin has not been determined. The pathways leading to the reduction of ferric iron and its incorporation into proteins is not clear. Recently, it has been shown that iron metabolism deregula- tion in fur mutants produces an iron overload, leading to oxidative stress and DNA damage including lethal and mu- tagenic lesions (17). Fur also appears to regulate iron and manganese SOD, key enzymes in the defense against oxygen toxicity (22).
Utilizing E. coli strains with mutations in these genes to alter both the level of enterobactin and intracellular iron pools, this study determined in E. coli ( I ) the extent that the iron chelator, enterobactin, acts as a photosensitizer for NUV and (2) the effect of increased intracellular iron on NUV- induced lethality. Our working hypothesis was that if the Fenton reaction plays a significant role during NUV irradi- ation in generating hydroxyl radicals capable of damaging DNA and other targets by a variety of mechanisms, an in- crease in the level of intracellular Fe+2 will increase cell lethality.
MATERIALS AND METHODS
Strains ofE. coli. MC4100: F araD139 A(lacI0PZYA) U169 rpsL thiA; MK3: same as MC4100 but entA::mini-kan; MK67: same as MC4100 butfes::mini-kan; MFT5: same as MC4100 butfur::TnlO. All of these strains were provided by Dr. Mark Mclntosh’s labora- tory at the University of Missouri-Columbia.
Media and growth conditions. All experiments used late log- phase suspensions (ODO,K, of 1 .O) started from a I : 100 dilution of an overnight culture grown in Luria-Bertani broth (LB; 10 g/L tryp- tone [Difco Laboratories, Detroit, MI], 5 g/L yeast extract [Difco] and 10 g/L NaCl [Fisher Scientific Co., Springfield, NJ], 40 pg/mL of kanamycin added when appropriate [Sigma Chemical Co.. St. Louis, MO]). Before NUV irradiation, late log-phase cells growing in LB were diluted 1:100 in M9 salts (23) in order to eliminate indirect radiation effects due to exposure of medium constituents. The NUV irradation was delivered at a rate of 22 Wlm2 measured at 365 nm with a UV meter (Spectroline, Westbury, NY). It was performed in an aluminum foil-lined box that contained six 15 W GE FISTSB black-light bulbs mounted on the inside cover. The sides of the box were open and a fan was mounted on one side to maintain a temperature of approximately 34°C. Cell suspensions (50 mL in M9 salts) were irradiated in 9 cm glass dishes covered with Pyrex petri plates. In some experiments. Mylar plastic film was placed over the Pyrex cover to block the passage of the UVB com-
10:
1 7
1:
.01 :
.w1 I
1
wild
wlld
fes
OW1 I 100 200 300 4 0 0
Fluence (KJlm2)
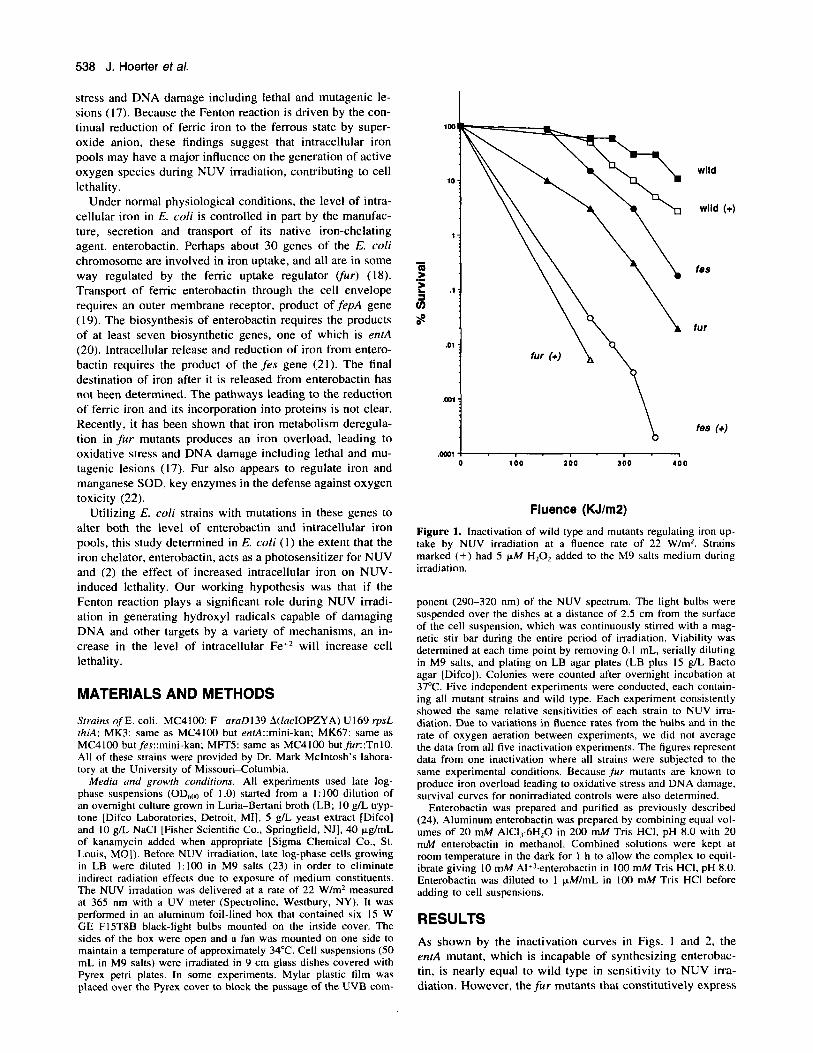
Figure 1. Inactivation of wild type and mutants regulating iron up- take by NUV irradiation at a fluence rate of 22 W/m’. Strains marked (+) had 5 IJ.M HzOz added to the M9 salts medium during irradiation.
ponent (290-320 nm) of the NUV spectrum. The light bulbs were suspended over the dishes at a distance of 2.5 cm from the surface of the cell suspension, which was continuously stirred with a mag- netic stir bar during the entire period of irradiation. Viability was determined at each time point by removing 0. I mL, serially diluting in M9 salts, and plating on LB agar plates (LB plus 15 g/L Bacto agar [Difco]). Colonies were counted after overnight incubation at 37°C. Five independent experiments were conducted, each contain- ing all mutant strains and wild type. Each experiment consistently showed the same relative sensitivities of each strain to NUV irra- diation. Due to variations in fluence rates from the bulbs and in the rate of oxygen aeration between experiments, we did not average the data from all five inactivation experiments. The figures represent data from one inactivation where all strains were subjected to the same experimental conditions. Because fur mutants are known to produce iron overload leading to oxidative stress and DNA damage. survival curves for nonirradiated controls were also determined.
Enterobactin was prepared and purified as previously described (24). Aluminum enterobactin was prepared by combining equal vol- umes of 20 mM AICI3.6H2O in 200 mM Tris HCI, pH 8.0 with 20 mM enterobactin in methanol. Combined solutions were kept at room temperature in the dark for 1 h to allow the complex to equil- ibrate giving 10 mM AIt3.enterobactin in 100 mM Tris HCI, pH 8.0. Enterobactin was diluted to I pMImL in 100 mM Tris HCI before adding to cell suspensions.
RESULTS As shown by the inactivation curves in Figs. 1 and 2, the enrA mutant, which is incapable of synthesizing enterobac- tin, is nearly equal to wild type in sensitivity to NUV irra- diation. However, the fur mutants that constitutively express
I
Photochemistry and Photobiology, 1996, 64(3) 539
DISCUSSION
.01 1
entA
entA (Ak3)
\ .001:
0 1 0 0 2 0 0 3 0 0 4 0 0
Fluence (KJ/m2)
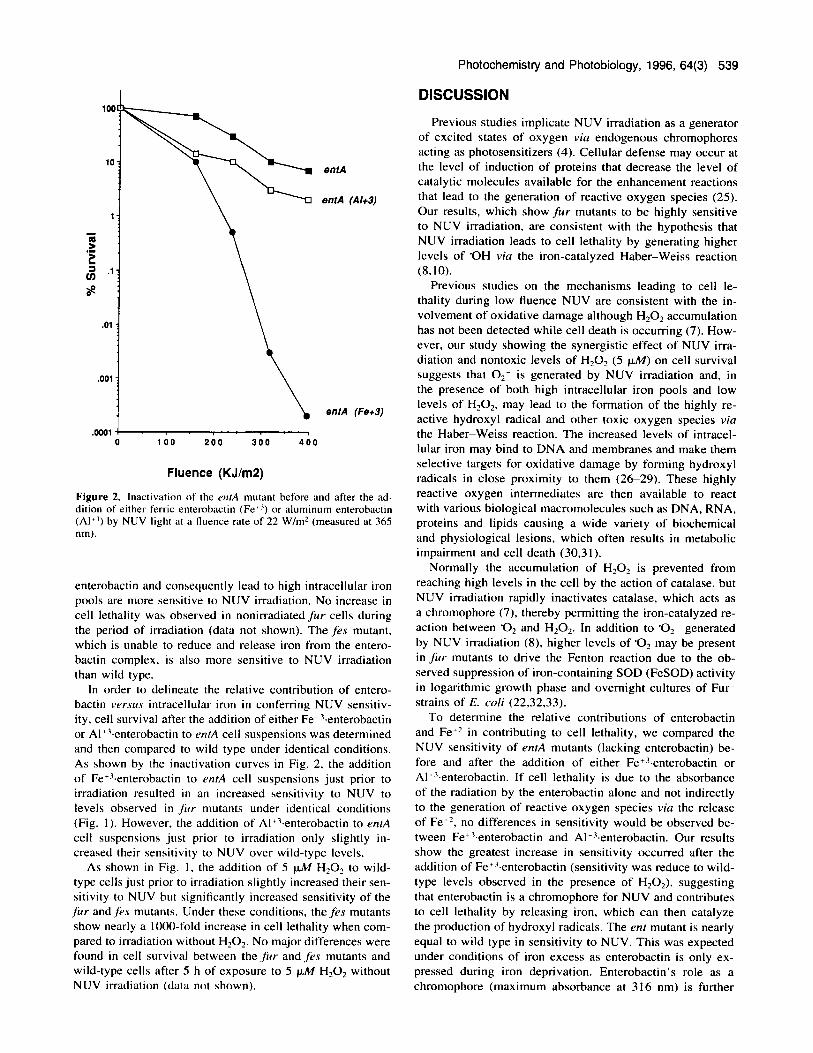
Figure 2. Inactivation of the emA mutant before and after the ad- dition of either ferric enterobactin (Fet3) or aluminum enterobactin (Al+3) by NUV light at a Ruence rate of 22 W/m2 (measured at 365 nm).
enterobactin and consequently lead to high intracellular iron pools are more sensitive to NUV irradiation. No increase in cell lethality was observed in nonirradiated fu r cells during the period of irradiation (data not shown). The fes mutant, which is unable to reduce and release iron from the entero- bactin complex, is also more sensitive to NUV irradiation than wild type.
In order to delineate the relative contribution of entero- bactin versus intracellular iron in conferring NUV sensitiv- ity, cell survival after the addition of either Fe+3.enterobactin or Al+3.enterobactin to entA cell suspensions was determined and then compared to wild type under identical conditions. As shown by the inactivation curves in Fig. 2, the addition of Fe+3.enterobactin to enrA cell suspensions just prior to irradiation resulted in an increased sensitivity to NUV to levels observed in fur mutants under identical conditions (Fig. I ) . However, the addition of Al+'.enterobactin to entA cell suspensions just prior to irradiation only slightly in- creased their sensitivity to NUV over wild-type levels.
As shown in Fig. 1, the addition of 5 FM H202 to wild- type cells just prior to irradiation slightly increased their sen- sitivity to NUV but significantly increased sensitivity of the ,fur and fes mutants. Under these conditions, the fes mutants show nearly a 1000-fold increase in cell lethality when com- pared to irradiation without H202. N o major differences were found in cell survival between the fur andjes mutants and wild-type cells after 5 h of exposure to 5 pM H,O, without NUV irradiation (data not shown).
Previous studies implicate NUV irradiation as a generator of excited states of oxygen viu endogenous chromophores acting as photosensitizers (4). Cellular defense may occur at the level of induction of proteins that decrease the level of catalytic molecules available for the enhancement reactions that lead to the generation of reactive oxygen species (25). Our results, which show fur mutants to be highly sensitive to NUV irradiation, are consistent with the hypothesis that NUV irradiation leads to cell lethality by generating higher levels of 'OH via the iron-catalyzed Haber-Weiss reaction (&lo).
Previous studies on the mechanisms leading to cell le- thality during low fluence NUV are consistent with the in- volvement of oxidative damage although H,O2 accumulation has not been detected while cell death is occurring (7). How- ever, our study showing the synergistic effect of NUV irra- diation and nontoxic levels of HzO, ( 5 ~ k f ) on cell survival suggests that 0,- is generated by NUV irradiation and, in the presence of both high intracellular iron pools and low levels of H,O,, may lead to the formation of the highly re- active hydroxyl radical and other toxic oxygen species via the Haber-Weiss reaction. The increased levels of intracel- Mar iron may bind to DNA and membranes and make them selective targets for oxidative damage by forming hydroxyl radicals in close proximity to them (26-29). These highly reactive oxygen intermediates are then available to react with various biological macromolecules such as DNA, RNA, proteins and lipids causing a wide variety of biochemical and physiological lesions, which often results in metabolic impairment and cell death (30.3 1 ).
Normally the accumulation of H,O, is prevented from reaching high levels in the cell by the action of catalase, but NUV irradiation rapidly inactivates catalase, which acts as a chromophore (7), thereby permitting the iron-catalyzed re- action between '02 and H,O,. In addition to *02- generated by NUV irradiation (8), higher levels of'O, may be present in fur mutants to drive the Fenton reaction due to the ob- served suppression of iron-containing SOD (FeSOD) activity in logarithmic growth phase and overnight cultures of Fur- strains of E. coli (22.32.33).
To determine the relative contributions of enterobactin and Fe'? in contributing to cell lethality, we compared the NUV sensitivity of entA mutants (lacking enterobactin) be- fore and after the addition of either Fe+"enterobactin or Al+3.enterobactin. If cell lethality is due to the absorbance of the radiation by the enterobactin alone and not indirectly to the generation of reactive oxygen species via the release of Fe'?, no differences in sensitivity would be observed be- tween Fe+3.enterobactin and Al+3.enterobactin. Our results show the greatest increase in sensitivity occurred after the addition of Fec3.enterobactin (sensitivity was reduce to wild- type levels observed in the presence of H,O,), suggesting that enterobactin is a chromophore for NUV and contributes to cell lethality by releasing iron, which can then catalyze the production of hydroxyl radicals. The ent mutant is nearly equal to wild type in sensitivity to NUV. This was expected under conditions of iron excess as enterobactin is only ex- pressed during iron deprivation. Enterobactin's role as a chromophore (maximum absorbance at 316 nm) is further

540 J. Hoerter et a/.
REFERENCES Enterobactin Fe"
Fe" NUV+ 4
H202 + 02'- + 0 2 + OH- + 'OH f f 4
catnlsse t NUV inactivation
DNA Damage
Figure 3. Proposed model to explain how the release of iron from enterobactin during NUV irradiation in concert with other known effects of NUV drives the Fenton reaction leading to the production of highly deleterious hydroxyl radicals.
supported by our finding that Mylar film (blocks wave- lengths below 320 nm) reduced cell lethality in fes mutants to wild-type levels (data not shown).
There are at least three mechanisms that might explain the observed increase in sensitivity of fes mutants despite the fact that under normal conditions they are unable to increase the intracellular iron pools via the release and reduction of Fe+' from enterobactin: (1) Enterobactin is a chromophore for NUV leading to the release of iron via the destruction of the ligand, resulting in a transient iron overload. The ele- vated levels of iron produce 'OH at an accelerated rate upon reaction with low levels of H202 generated by NUV irradi- ation (27). This possibility is supported by the recent finding that ferritin is photoreduced by UVA (12) and UVB (14) irradiation. (2) The superoxide anion possibly generated by NUV irradiation triggers the release and reduction of iron from enterobactin in a way similar to that in which super- oxide anion is able to reduce ferritin (33.34). (3) Low levels of hydrogen peroxide, generated by NUV irradiation react with elevated levels of Fe+' bound to enterobactin to gen- erate hydroxyl radicals in a manner similar to that observed in vitro by ferric iron chelates ( 1 1,34-36). The increased sensitivity of the feu mutant over wild type may be due to the sudden burst of iron released from high intracellular con- centrations of Fe''.enterobactin. This strain may also contain higher concentrations of enterobactin for absorbing more ra- diant energy, leading to oxidative stress and increased cell lethality. Figure 3 illustrates our hypothesis for explaining how the release of iron from enterobactin during NUV ir- radiation drives the Fenton reaction leading to the production of highly deleterious hydroxyl radicals.
We recognize that the sensitivity of fur mutants may also be related to the absence of other gene products regulated by Fur. An increasing number of genes have been found to be Fur regulated, although they have no direct involvement with iron transport systems. However, our studies utilizing other known genes affecting the iron transport system strongly suggest that one mechanism by which NUV irra- diation leads to cell lethality is by creating a transient iron overload, producing very favorable conditions for the pro- duction of highly deleterious free radicals through a variety of mechanisms that lead to oxidative stress and DNA dam- age including lethal and mutagenic lesions.
Acknowledgemenrs-The authors thank Professor Mark McIntosh for bacterial strains and for his guidance during this study. This research was made possible by an NlH Academic Research En- hancement Award (ES06399-02SI) to J.H. and by a National Insti- tutes of Environmental Health Sciences grant (ES04889) to A.E.
1. Tyrrell, R. M. and S. M. Keyse (1990) The interaction of UVA radiation with cultured cells. J. Phofochem. Phorobiol. 4, 349- 363.
2. Favre, A,, E. Hajinsdorf, K. Thiam and A. Calderia de Araujo (1985) Mutagenesis and growth delay induced in Escherichiu coli by near-UV radiation. Biochimie 67, 335-342.
3. Jagger, J. ( I 985) Solar-UV Acrions on Living CeIls. Praeger, New York.
4. Peak, J. G., M. J. Peak and C. S. Foote (1982) Effects of glyc- erol upon the biological actions of near-ultraviolet light; spectra and concentration dependence for transforming DNA and for Escherichia coli Blr. Phorochem. Phorobiol. 36, 41 3 4 16.
5. Webb, R. B. and J. R. Lorenz (1970) Oxygen dependence and repair of lethal effects of near-UV and visible light. Phororhern. Photobid. 12, 383-389.
6. Ahmad, S. I. (1981) Synergistic killing of coliphage T7 by near- ultraviolet radiation plus hydrogen peroxide: possible role of superoxide radicals. Phorobiochem. Phorobiophys. 2, 173- I 80.
7. Kramer, G. F. and B. N. Ames (1987) Oxidative mechanisms of toxicity of low-intensity near-UV light. J. Bacreriol. 169,
8. Hoerter, J., A. Eisenstark and D. Touati (1989) Mutations by near-UV radiation in Escherichia coli strains lacking superoxide dismutase. Murar. Res. 215, 161-165.
9. Ananthaswamy, H. N. and A. Eisenstark (1976) Near UV-in- duced breaks in phage DNA: sensitization by hydrogen peroxide (a tryptophan photoproduct). Photochem. Phorobiol. 24, 439- 442.
10. Imlay, J. A,, S. M. Chin and S. Linn (1988) Toxic DNA damage by hydrogen peroxide through the Fenton reaction in vivo and in virro. Science 240, 640-642.
11. Tachon, P. ( 1989) Ferric and cupric ions requirement for DNA single-stranded breakage by hydrogen peroxide. Free Rudical Res. Commun. 7, 1-10,
12. Aubailly, M., R. Santus and S. Salmon ( 1991) Ferrous ion re- lease from ferritin by ultraviolet-A radiation. J. Phorochem. Phorobiol. 54, 769-173.
13. Laulhere. J. P., A. M. Laboure' and J. F. Briat (1990) Photo- reduction and incorporation of iron into ferritins. Biochem. J . 269, 79-84.
14. Aubailly, M., S. Salmon and R. Santus (1990) UV (A,B)-in- duced ferrous ion release from ferritin. Phorochem. Phorobiol. 51, 34.
15. Thomas, C. E., L. A. Morehouse and S. D. Aust (1985) Ferritin and superoxide dependent lipid peroxidation. J. Bid. Chem.
16. Boyer, R. F. and C. J. McCleary (1987) Superoxide ion as a primary reductant in ascorbate-mediated ferritin ion release. Free Radical B i d . Med. 3. 389-395.
17. Touati, D., M. Jacques, B. Tardat, L. Bouchard and S. Despied (1995) Lethal oxidative damage and mutagenesis are generated by iron in A& mutants of Escherichia coli: protective role of superoxide dismutase. J. Bacreriol. 177, 2305-23 14.
18. Bagg, A. and J. B. Neilands (1987) Ferris uptake regulation protein acts as a repressor, employing iron (11) as a cofactor to bind the operator of an iron transport operon in Eschrrirhiu coli. Biochemisrry 26, 547 1-5477.
19. Pugsley, A. P. and P. Reeves (1976) Transport of ferric entero- bactin through the cell envelope requires outer membrane re- ceptor, product of fepA gene. J . Bacreriol. 127, 2 18-228.
20. Nahlik. M. S., T. P. Fleming and M. A. McIntosh (1987) Cluster of genes controlling synthesis and activation of 2.3-dihydroxy- benzoic acid in production of enterobactin in Escherichiu coli. J. Bucreriol. 169, 4 163-4 170.
21. Langman. L., I. G. Young, G. E. Frost, H. Rosenberg and F. Gibson (1972) Enterochelin system of iron transport in Esche- richia coli mutation affecting ferric-enterochelin esterase. J . Bacreriol. 112, 1142-1 149.
22. Niederhoffer, E. C., C. M. Naranjo, K. L. Bradley and J. A. Fee (1990) Control of E. coli superoxide dismutase (sodA and sr/dB) genes by the ferric uptake regulation (fur) locus J. Bacreriol. 172, 1930-1938.
2259-2266.
260, 3275-3280.

Photochemistry and Photobiology, 1996, 64(3) 541
23. Miller, J . H. (1972) Experinienrs in Molecular Generics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
24. Hollifield, W. C. and J. B. Neilands (1978) Ferric enterobactin transport system in Escherichia coli K- 12. Extraction, assay, and specificity of the outer membrane receptor. Biochemistry 17,
25. Hoerter, J. and A. Eisenstark (1990) Patterns of protein synthe- sis in a growth delay mutant (nuv) of E. coli after treatment of near-UV radiation or hydrogen peroxide. J . Phorochem. Pho- tohiol. B Biol. 6 , 283-289.
26. Halliwell, B. (1987) Oxidants and human disease; some new concepts. FEBS J. 1, 358-364.
27. Halliwell, B. (1994) Free radicals, antioxidants, and human dis- ease. Lancer 344, 721-724.
28. Czapski, G. (1984) On the use of hydroxyl radical scavengers in biological systems. Isr. J. Chem. 24. 19.
29. Goldstein, S. and G. Czapski (1986) The role and mechanism of metal ions and their complexes in enhancing damage in bi- ological systems or in protecting these systems from the toxicity of superoxide radical. J. Free Radicals Biol. Med. 2, 3-1 I .
1922-1 928.
30. Jagger, J. (1981) Near-UV radiation effects on microorganisms. Phorochem. Phorobiol. 34, 761-768.
3 1. Farr, S. B., D. Touati and T. Kogoma (1988) Effects of oxygen stress on membrane functions in Escherichia coli: role of HPI catalase. J. Bacteriol. 170, 1837-1 842.
32. Boyer, R. F. and C. J. McCleary (1987) Superoxide ion as a primary reductant in ascorbate-mediated ferritin ion release. Free Radical Biol. Med. 3, 389-395.
33. Bando, Y. and K. Aki (1990) Superoxide-mediated release of iron from ferritin by some flavoenzymes. Biochem. Biophys. Res. Commun. 168, 389-395.
34. Larson, R. A.. R. E. Lloyd, K. A. Marley and R. W. Tuveson (1992) Ferric-ion-photosensitized damage to DNA by hydroxyl and non-hydroxyl radical mechanisms. J. Phorochem. Phoro- b id . B Biol. 14, 345-357.
35. Singer, B. and H. Fraenkel-Conrat (1965) Effects of light in the presence of iron salts on ribonucleic and model compounds. Biochemistry 4, 226-233.
36. Cernohorsky, I. J. and G. M. Blackurn (1971) Photodynamic effect of Fe+3 upon bases of nucleic acids. Proc. First European Biophysics Congr. Radial. Biophys. Free Radicals 2, 29-3 I .