resveratrol prevents fibrosis, nf-κb activation and tgf-β increases induced by chronic ccl4...
TRANSCRIPT

RESVERATROL PREVENTS LIVER FIBROSIS 35
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
JOURNAL OF APPLIED TOXICOLOGYJ. Appl. Toxicol. 2008; 28: 35–43Published online 11 April 2007 in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/jat.1249
Resveratrol prevents fibrosis, NF-κκκκκB activation and TGF-βββββincreases induced by chronic CCl4 treatment in rats
Enrique Chávez,1 Karina Reyes-Gordillo,1 José Segovia,2 Mineko Shibayama,3 Victor Tsutsumi,3
Paula Vergara,2 Mario G. Moreno1 and Pablo Muriel1,*
1 Sección Externa de Farmacología, Cinvestav-IPN, Apdo Postal 14-740, México 07000, D.F. México2 Departamento de Fisiología Biofísica y Neurociencias, Cinvestav-IPN, Apdo Postal 14-740, México 07000, D.F. México3 Departamento de Patología Experimental, Cinvestav-IPN, Apdo Postal 14-740, México 07000, D.F. México
Received 8 January 2007; Revised 19 February 2007; Accepted 21 February 2007
ABSTRACT: Resveratrol is a nonflavonoid polyphenol with antioxidant, anticancer and antiinflammatory properties.
Moreover, it has been reported that this compound inhibits NF-κκκκκB, which regulates the transcription of several genes
including cytokines such as the profibrogenic TGF-βββββ . The aim of this work was to evaluate the pharmacological effects
of resveratrol on CCl4-induced cirrhosis in the rat. Four groups were formed: the control group that received the vehicles
only; the CCl4 group that received the toxin (0.4 g kg−−−−−1, i.p., three times a week, for 8 weeks); the CCl4 plus resveratrol
(10 mg kg−−−−−1, daily) group; and the resveratrol alone group. Alanine aminotransferase, alkaline phosphatase and bilirubins
were increased by CCl4, but resveratrol afforded some degree of protection. Glycogen was decreased markedly by CCl4
and resveratrol prevented almost completely this effect. No antioxidant effect of resveratrol was observed. One of the most
prominent effects was on fibrosis which increased near 5-fold (hydroxyproline) in the CCl4 group; resveratrol preserved
the content of collagen. These results were corroborated by histopathology. To elucidate the antifibrogenic mechanism of
resveratrol, the activation of NF-κκκκκ B and the production of TGF-βββββ were measured; in both cases CCl4 increased them and
resveratrol abolished them; however, changes in NF-κκκκκ B were modest and did not reach statistical significance, while the
increase in TGF-βββββ was about three fold and resveratrol decreased it under control values. Together, the present results
indicate that resveratrol possesses a strong antifibrogenic effect at least in the CCl4 model of cirrhosis. Moreover, the
action mechanism is probably associated with its ability to reduce NF-κκκκκB activation and TGF-βββββ content. Copyright © 2007
John Wiley & Sons, Ltd.
KEY WORDS: resveratrol; cytokines; liver injury; carbon tetrachloride; necrosis; inflammation; fibrosis; cirrhosis; NF-κB;
TGF-β
* Correspondence to: Dr Pablo Muriel, Sección Externa de Farmacología,
Cinvestav-IPN, Apdo Postal 14-740, México 07000, D.F., México.
E-mail: [email protected]
Contract/grant sponsor: Conacyt; contract/grant number: 42721.
particular transforming growth factor-β (TGF-β) has been
associated with the fibrotic process and thus it has been
the aim of some studies to interfere with the disease
(Muriel, 2007b). Nuclear factor κB (NF-κB) is an induc-
ible factor that functions to enhance the transcription
of a variety of genes including those for cytokines and it
is well known for its involvement in inflammatory
responses (Ghosh et al., 1998). Therefore, NF-κB is an-
other interesting target in the treatment of liver diseases.
On the other hand, resveratrol (3,5,4′-trans-trihydro-
xystilbene), a natural phytoalexin present in grapes,
peanuts and red wine, has various pharmacological
effects including antiinflammatory and antioxidant prop-
erties, modulation of lipid metabolism and prevention
of cancer (Fremont, 2000; Dong, 2003; Aggarwal et al.,
2004). In addition, recently it was demonstrated that
resveratrol protects against acetaminophen-induced toxi-
city in mice (Sener et al., 2006a). Importantly, it has been
reported that resveratrol inhibits the activation of NF-κB;
some authors described a reduced nuclear appearance
of NF-κB factor (Tsai et al., 1999) and others showed
an interference with their transcription activity (Pendurthi
Introduction
Experimental CCl4-induced cirrhosis seems to reproduce
the main characteristics of human cirrhosis: the liver is
grossly nodular; there is portal hypertension in most of
the animals, and the normal architecture is replaced by
nodules of regenerating liver surrounded by fibrotic septa
with proliferating bile ducts. Therefore, the CCl4-induced
cirrhosis is the most widely used model to reproduce
human cirrhosis in experimental animals to study patho-
physiological mechanisms of the disease, and many
possible antifibrotic drugs are tested in this model before
using them in humans (Muriel, 2007a). Hepatic stellate
cells (HSC) are activated as a result of liver injury. Once
activated, HSC increase their production of extracellular
matrix (ECM) proteins finally leading to fibrosis and
cirrhosis. Activation of HSC is dependent on cytokines, in

36 E. CHAVEZ ET AL.
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
et al., 1999). Furthermore, it has been shown that
resveratrol decreases proinflammatory cytokine expres-
sion in vitro (Wadsworth and Koop, 1999). Therefore this
work investigated whether resveratrol was capable of
preventing CCl4-induced liver cirrhosis. It was found
that CCl4 produced fibrosis, activation of NF-κB and
production of TGF-β; resveratrol prevented fibrosis
completely and this effect was accompanied by preven-
tion of the translocation of NF-κB to the nucleus and
attenuation of the profibrotic cytokine TGF-β.
Material and Methods
Chemicals
Resveratrol, chloramine-T, methyl cellosolve sodium
thiosulphate, p-dimethylaminobenzaldehyde, antrone,
carboxymethylcellulose, thiobarbarbituric acid, γ-
glutamyl-p-nitroanilide, L-γ-glutamyl-p-nitroaniline,
p-nitrophenyl phosphate and bovine serum albumin
were purchased from the Sigma Chemical Company
(St Louis, MO, USA). Carbon tetrachloride, sodium
acetate, sodium hydroxide, glacial acetic acid, hydro-
chloric acid, sulfuric acid, ethanol, methanol, potassium
hydroxide and formaldehyde were obtained from J.T.
Backer (Xalostoc, Mexico).
Study Design
Wistar male rats weighing initially 90–100 g and fed
a Purina chow rat diet ad libitum were used. Cirrhosis
was produced by i.p. administration of CCl4 (0.4 g kg−1
of body weight) dissolved in mineral oil three times
per week for 8 weeks. Four groups were performed.
Group 1 (n = 8), consisted of control animals receiving
the vehicle only (oil). In group 2 (n = 15), CCl4 was
administered for 8 weeks. Animals in group 3 (n = 15)
received resveratrol (10 mg kg−1 suspended in 0.7%
carboxymethyl-cellulose (CMC) p.o., for 8 weeks) plus
CCl4. The rats in group 4 (n = 8) received resveratrol
only. All the animals were killed under light ether
anesthesia 72 h after the last dose of CCl4 or mineral oil.
All determinations with the exception of NF-κB (see
below) were measured at this time. In order to determine
the activation of NF-κB another set of animals weighing
around 200 g received the same treatments, but CCl4 was
given only once and resveratrol administered 15 min
before CCl4; in this particular case the animals were
killed 30 min after CCl4 or mineral oil because this is
approximately the time in which NF-κB is activated
and translocated to the nucleus. Blood was collected
by cardiac puncture and the liver was rapidly removed.
All samples were kept on ice until analysis. All animals
received humane care according to the institution’s
guidelines and the Mexican Official Norm (NOM-
062-ZOO-1999) regarding technical specifications for
production, care and use of laboratory animals.
Serum Enzyme Activities and BilirubinDetermination
Serum was obtained for determination of liver damage
by measuring the activities of alanine aminotransferase
(ALT) (Reitman and Frankel, 1957), alkaline phosphatase
(ALP) (Bergmeyer et al., 1983) and bilirubin content (kit
Bioxon-Mexico).
Assessment of Lipid Peroxidation
The extent of lipid peroxidation was estimated in liver
homogenates by measurement of malondialdehyde
(MDA) formation using the thiobarbituric acid method
(Okawa et al., 1979). Protein was determined according
to Bradford (1976) using bovine serum albumin as
standard.
Reduced and Oxidized GlutathioneDeterminations in Liver
GSH and GSSG were prepared on the day of the experi-
ment in 0.1 M sodium phosphate 0.005 M EDTA buffer
(pH 8.0) and kept on ice until used. The OPT solution
was prepared in reagent-grade absolute methanol just
prior to use.
Liver samples were homogenized on ice using a
polytron homogenizer. The solution used for homogeni-
zation consisted of 3.75 ml of the phosphate-EDTA
buffer and 1 ml of 25% H3PO4, which was used as a pro-
tein precipitant. The total homogenate was centrifuged
at 4 °C at 100 000 g for 30 min to obtain the supernatant
for the assay of GSH and GSSG.
Determination of GSH was performed according to
Hissin and Hilf (1976). To 0.01 ml of the 100 000 g
supernatant, 4.9 ml of the phosphate-EDTA buffer
pH 8.0 was added. The final assay mixture (2.0 ml) con-
tained 100 μl of the diluted tissue supernatant, 1.8 ml of
phosphate-EDTA buffer, and 100 μl of the OPT solution,
containing 100 μg of OPT. After through mixing and
incubation at room temperature for 15 min, the solu-
tion was transferred to a quartz cuvette. Fluorescence at
420 nm was determined with the activation at 350 nm.
For the GSSG assay, a 0.5 ml portion of the original
100 000 g supernatant was incubated at room temperature
with 200 μl of 0.04 M N-ethylmaleimide (NEM) for
30 min to interact with the GSH present in the tissue. To
this mixture, 4.3 ml of 0.1 N NaOH was added. A 100 μl
portion of this mixture was taken for the measurement of

RESVERATROL PREVENTS LIVER FIBROSIS 37
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
GSSG, as outlined above for the GSH assay, except
that 0.1 N NaOH was employed as a diluent rather than
phosphate-EDTA buffer.
Glycogen Determination
Small pieces of liver (0.5 g) were separated for glycogen
determination using the anthrone reagent according to
Seifter et al. (1950).
Histology
Samples of liver were taken from all the animals and
fixed with 10% formaldehyde in phosphate buffered
saline for 24 h. Those tissue pieces were washed with tap
water, dehydrated in alcohol and embedded in paraffin.
Sections of 6–7 μm were mounted on glass slides and
covered with silane. Masson’s trichromic stains were
performed on each slide.
Western Blot Assays
The TriPure reagent (Roche Diagnostics) was used to
isolate total protein from samples of liver tissue. Fresh
tissue was homogenized in 1 ml of TriPure reagent, then,
0.2 ml of chloroform was added to homogenates, and the
lower phase was treated with isopropanol to precipitate
total protein. Samples were centrifuged at 12 000 rpm
for 10 min at 4 °C, then, 3 washes were performed with
0.3 M guanidine hydrochloride in 95% ethanol. A final
wash was performed with 100% ethanol, samples were
centrifuged as previously described (Pérez-Severiano
et al., 2002), and the pellet resuspended in 1% SDS.
Volumes equivalent to 50 μg of proteins (determined by
the bis-cinconinic acid method) were transferred onto
12% polyacrylamide gel; separated proteins were trans-
ferred onto nitrocellulose paper (Amersham). Next, blots
were blocked with 5% skim milk and 0.05% Tween-20
for 30 min at room temperature and independently
incubated overnight at 4 °C with antibodies selective
against TGF-β (MAB 1032 from Chemicon Int. Inc.).
The following day, the membranes were washed and then
exposed to a secondary peroxidase-labeled antibody
(Zymed) diluted 1:4000 in the blocking solution for 1 h
at room temperature. Blots were washed and protein
developed using the ECL detection system (NEN Life
Sciences Products). Blots were stripped and incubated
with a monoclonal antibody directed against β-actin
(García-Tovar et al., 2001), which was used as a control
to normalize cytokine protein expression levels. The pro-
cedure to strip membranes was as follows. First, blots
were washed four times with phosphate-saline buffer
pH 7.4 (0.015 M, 0.9% NaCl), then immersed in stripping
buffer (2-mercaptoethanol 100 mM, sodium dodecyl
sulfate 2% and Tris-HCl 62.5 mM, pH 6.7) for 30 min at
60 °C with gentle shaking, membranes were then washed
five times with 0.05% Tween-20 in phosphate-saline
buffer.
Images were digitalized using the BioDoc-It System
(UVP) and then analysed densitometrically using the Lab
Works 4.0 Image Acquisition and Analysis software
(UVP), as previously described (Pérez-Severiano et al.,
2002).
Nuclear Protein Isolation and Shift Assays forNF-κκκκκB
The isolation of intact and stable nuclei consisted of the
iso-osmotic lysis procedure as reported by Dyer and
Hersog (1995). With this lysis procedure, the nuclear
envelope remains intact even during the further manipu-
lations of washing, freezing and ultracentrifugation and
provides nuclear extract for use in NF-κB assay.
NF-κB was estimated using a commercial kit (Non-
Radioactive NF-κB p50/p65 Transcription Factor Assay,
SGT510 Chemicon, Mexico City). This kit combines
the principle of the electrophoretic mobility shift assay
(EMSA) with the enzyme-linked immunosorbent assay
(ELISA).
Statistical Analysis
Data are expressed as mean value ± SEM. Comparisons
were carried out by analysis of variance followed by
Dunnett’s or Tukey’s test, as appropriate, using Sigma
Stat for Windows (2.0 Version). Differences were consid-
ered statistically significant when P < 0.05.
Results
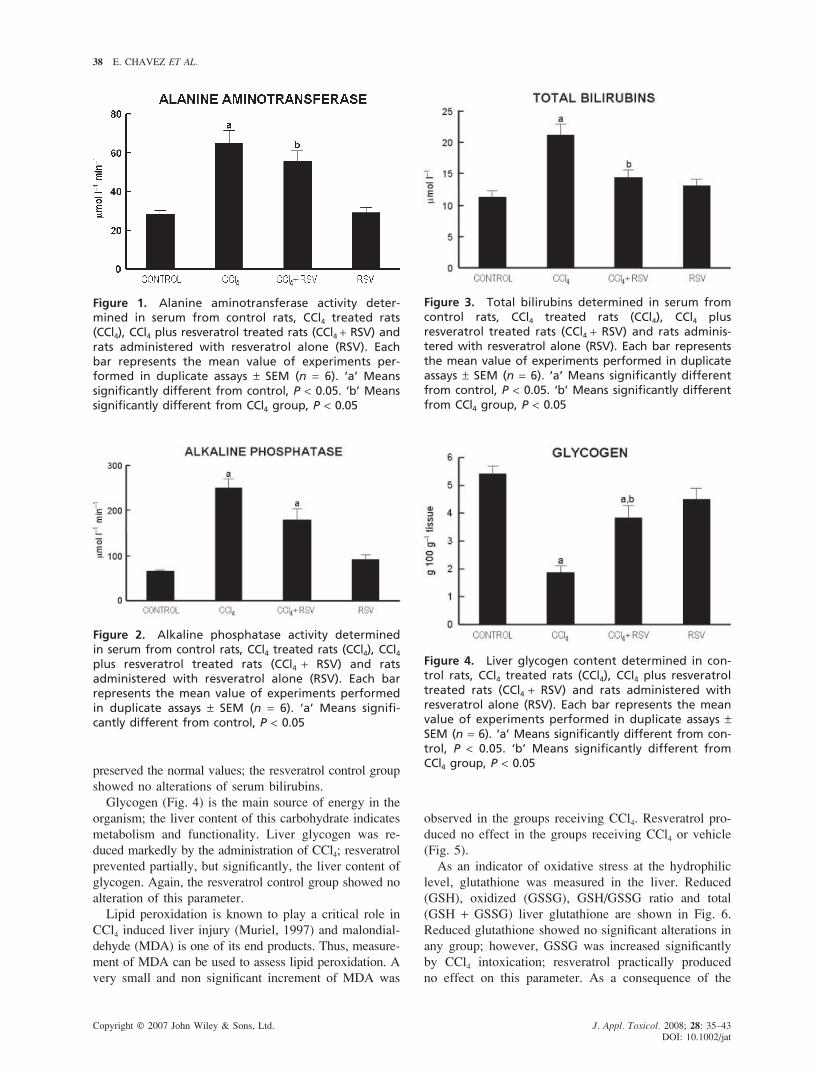
The activity of ALT in serum is shown in Fig. 1. ALT is
a cytosolic enzyme of the hepatocyte and an increase of
enzyme activity reflects an increase in plasma membrane
permeability, which, in turn, is associated with cell death
(Rosen and Keeffe, 2000). Chronic CCl4 treatment in-
creased significantly ALT activity; resveratrol administra-
tion prevented significantly this effect. Resveratrol by
itself did not modify ALT activity.
Serum enzyme activity of ALP is an indicator
of cholestasis. Carbon tetrachloride induced ALP activity
by about 3-fold (Fig. 2). Resveratrol partially prevented
the increase of this enzyme; however, the statistical
analysis shows that this effect was not significant.
The concentration of total bilirubins in serum is
depicted in Fig. 3. Chronic CCl4 treatment increased
bilirubins significantly, while co-treatment with resveratrol
38 E. CHAVEZ ET AL.
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
Figure 1. Alanine aminotransferase activity deter-mined in serum from control rats, CCl4 treated rats(CCl4), CCl4 plus resveratrol treated rats (CCl4 + RSV) andrats administered with resveratrol alone (RSV). Eachbar represents the mean value of experiments per-formed in duplicate assays ± SEM (n = 6). ‘a’ Meanssignificantly different from control, P < 0.05. ‘b’ Meanssignificantly different from CCl4 group, P < 0.05
preserved the normal values; the resveratrol control group
showed no alterations of serum bilirubins.
Glycogen (Fig. 4) is the main source of energy in the
organism; the liver content of this carbohydrate indicates
metabolism and functionality. Liver glycogen was re-
duced markedly by the administration of CCl4; resveratrol
prevented partially, but significantly, the liver content of
glycogen. Again, the resveratrol control group showed no
alteration of this parameter.
Lipid peroxidation is known to play a critical role in
CCl4 induced liver injury (Muriel, 1997) and malondial-
dehyde (MDA) is one of its end products. Thus, measure-
ment of MDA can be used to assess lipid peroxidation. A
very small and non significant increment of MDA was
Figure 2. Alkaline phosphatase activity determinedin serum from control rats, CCl4 treated rats (CCl4), CCl4plus resveratrol treated rats (CCl4 + RSV) and ratsadministered with resveratrol alone (RSV). Each barrepresents the mean value of experiments performedin duplicate assays ± SEM (n = 6). ‘a’ Means signifi-cantly different from control, P < 0.05
Figure 3. Total bilirubins determined in serum fromcontrol rats, CCl4 treated rats (CCl4), CCl4 plusresveratrol treated rats (CCl4 + RSV) and rats adminis-tered with resveratrol alone (RSV). Each bar representsthe mean value of experiments performed in duplicateassays ± SEM (n = 6). ‘a’ Means significantly differentfrom control, P < 0.05. ‘b’ Means significantly differentfrom CCl4 group, P < 0.05
Figure 4. Liver glycogen content determined in con-trol rats, CCl4 treated rats (CCl4), CCl4 plus resveratroltreated rats (CCl4 + RSV) and rats administered withresveratrol alone (RSV). Each bar represents the meanvalue of experiments performed in duplicate assays ±SEM (n = 6). ‘a’ Means significantly different from con-trol, P < 0.05. ‘b’ Means significantly different fromCCl4 group, P < 0.05
observed in the groups receiving CCl4. Resveratrol pro-
duced no effect in the groups receiving CCl4 or vehicle
(Fig. 5).
As an indicator of oxidative stress at the hydrophilic
level, glutathione was measured in the liver. Reduced
(GSH), oxidized (GSSG), GSH/GSSG ratio and total
(GSH + GSSG) liver glutathione are shown in Fig. 6.
Reduced glutathione showed no significant alterations in
any group; however, GSSG was increased significantly
by CCl4 intoxication; resveratrol practically produced
no effect on this parameter. As a consequence of the

RESVERATROL PREVENTS LIVER FIBROSIS 39
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
Figure 5. Liver lipid peroxidation determined asmalondialdehyde (MDA) content in control rats, CCl4treated rats (CCl4), CCl4 plus resveratrol treated rats(CCl4 + RSV) and rats administered with resveratrolalone (RSV). Each bar represents the mean value ofexperiments performed in duplicate assays ± SE (n = 6)
increase in GSSG, the GSH/GSSG ratio decreased
significantly in the groups that received CCl4; the total
glutathione showed no significant alterations in any group.
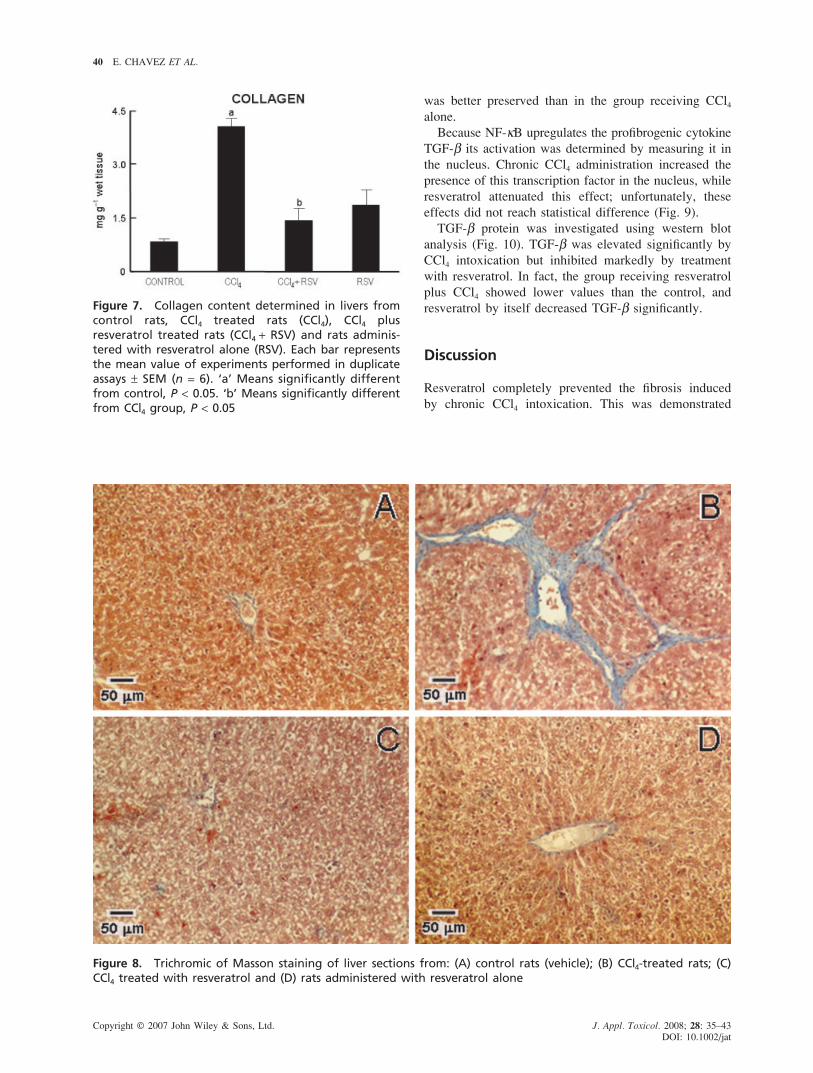
Fibrosis is the main characteristic of any chronic liver
damage and collagen is the main protein accumulating in
fibrotic tissue. Therefore, fibrosis was analysed from two
points of view: biochemically by measuring hydroxypro-
line (an amino acid exclusive of collagen) (Fig. 7),
and histologically by visualizing the blue color of the
collagen fibers in the trichromic stained liver sections
(Fig. 8). Hydroxyproline was increased by nearly 5-fold
by chronic CCl4 intoxication and, importantly, resveratrol
prevented completely the increase in collagen (Fig. 7).
The histopathological analysis revealed that chronic
CCl4 intoxication produced a marked increase in collagen
deposition around the portal triad, also nodules of
hepatocytes surrounded by variable size collagen bands
(blue stained) were frequently observed, the normal
architecture was lost and extended necrotic areas were
present. In agreement with the biochemical analysis, the
group treated simultaneously with resveratrol and CCl4
showed no accumulation of collagen and the parenchyma
Figure 6. Reduced (GSH) and oxidized (GSSG) glutathione, GSH/GSSG ratio and total glutathione (GSH + GSSG)determined in livers from control rats, CCl4 treated rats (CCl4), CCl4 plus resveratrol treated rats (CCl4 + RSV) andrats administered with resveratrol alone (RSV). Each bar represents the mean value of experiments performed induplicate assays ± SEM (n = 6). ‘a’ Means significantly different from control, P < 0.05. ‘b’ Means significantly dif-ferent from CCl4 group, P < 0.05
40 E. CHAVEZ ET AL.
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
Figure 7. Collagen content determined in livers fromcontrol rats, CCl4 treated rats (CCl4), CCl4 plusresveratrol treated rats (CCl4 + RSV) and rats adminis-tered with resveratrol alone (RSV). Each bar representsthe mean value of experiments performed in duplicateassays ± SEM (n = 6). ‘a’ Means significantly differentfrom control, P < 0.05. ‘b’ Means significantly differentfrom CCl4 group, P < 0.05
was better preserved than in the group receiving CCl4
alone.
Because NF-κB upregulates the profibrogenic cytokine
TGF-β its activation was determined by measuring it in
the nucleus. Chronic CCl4 administration increased the
presence of this transcription factor in the nucleus, while
resveratrol attenuated this effect; unfortunately, these
effects did not reach statistical difference (Fig. 9).
TGF-β protein was investigated using western blot
analysis (Fig. 10). TGF-β was elevated significantly by
CCl4 intoxication but inhibited markedly by treatment
with resveratrol. In fact, the group receiving resveratrol
plus CCl4 showed lower values than the control, and
resveratrol by itself decreased TGF-β significantly.
Discussion
Resveratrol completely prevented the fibrosis induced
by chronic CCl4 intoxication. This was demonstrated
Figure 8. Trichromic of Masson staining of liver sections from: (A) control rats (vehicle); (B) CCl4-treated rats; (C)CCl4 treated with resveratrol and (D) rats administered with resveratrol alone

RESVERATROL PREVENTS LIVER FIBROSIS 41
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
Figure 9. Analysis of NF-κB DNA-binding activity insamples of liver tissue from control rats, CCl4-treatedrats (CCl4), CCl4 treated with resveratrol (CCl4 + RSV)and rats administered with resveratrol alone (RSV).See the conditions of animal treatment to determinethis factor. NF-κB was estimated using a commercial kit(Non-Radioactive NF-κB p50/p65 Transcription FactorAssay) that combines the principle of the electrophore-tic mobility shift assay (EMSA) with the enzyme-linkedimmunosorbent assay (ELISA). HeLa whole cell (TNF-αtreated) was used as a positive control. Results are ex-pressed as relative optical density (OD) values obtainedusing spectrophometric analysis. Each bar representsthe mean value of six rats ± SEM. ‘a’ Means signifi-cantly different from control, P < 0.05. ‘b’ Means sig-nificantly different from CCl4 group, P < 0.05
both biochemically and histologically. To investigate the
action mechanism of the compound its effect on NF-κB
activation and the levels of one of its products, the
profibrotic cytokine TGF-β, which is involved in HSC
Figure 10. Resveratrol blockade of TGF-β proteinin samples of liver tissue determined by westernblot analysis from CCl4 treated rats (CCl4), CCl4 plusresveratrol treated rats (CCl4 + RSV), and rats adminis-tered with resveratrol alone (RSV). β-actin was usedas an internal control. Signal intensities were deter-mined by densitometric analysis of treated blots andvalues calculated as the ratio of TNF-α /β-actin. Eachbar represents the mean value of six rats ± SEM
activation and production of extracellular matrix proteins
were measured. It was found that CCl4 effectively in-
creased both NF-κB activation and TGF-β production;
resveratrol prevented TGF-β elevation providing a satis-
factory antifibrotic mechanism. However, other actions
of resveratrol cannot be discarded. The antifibrotic effects
of resveratrol have been observed in other organs and
in vitro (Godichaud et al., 2000; Olson et al., 2005;
Sener et al., 2006b; Abergel et al., 2006; Godichaud
et al., 2006). However, this is the first report on the
antifibrogenic effect of resveratrol on liver cirrhosis.
Liver cirrhosis is irreversible and a worldwide leading
cause of death. Today, there is no effective pharmaco-
logical treatment for this disease and fibrosis is the main
feature of the pathology. Liver fibrosis is characterized
by an excessive deposition of extracellular matrix com-
ponents. It is well known that most extracellular matrix
proteins are deposited by liver myofibroblasts. Myofibro-
blasts are very rare in the normal liver. Most studies
suggest that myofibroblast derive from HSC (Friedman,
1993). These precursor cells undergo phenotypic modifi-
cations that are collectively referred to as activation. The
principal phenotypic modifications are morphological
changes, de novo expression of α smooth muscle actin,
increased cellular proliferation and extracellular matrix
components synthesis (Bataller and Brenner, 2005).
The main factor involved in stellate cell activation is
the cytokine TGF-β (Gressner et al., 2002), making
this cytokine a suitable target to interfere with the fibro-
tic process (Gressner and Weiskirchen, 2006). In this
sense, the diminution of TGF-β protein afforded by
resveratrol observed herein is an outstanding effect of this
phytoalexin.
The TGF-β superfamily consists of multiple family
members, including the highly homologous TGF-β (TGF-
β1 TGF-β2 and TGF-β3) isoforms. TGF-β1 has shown
the most prominent fibrogenic properties (Border and
Ruoslahti, 1992; Massague et al., 1992; Muriel 2007b)
and is the isoform measured in this work. TGF-β1 seems
to lead to fibrogenesis through autocrine and paracrine
effects on HSC (Hellerbrand et al., 1999; Gressner and
Weiskirchen, 2006), the cells responsible for fibrogenesis.
Given the prominent role of TGF-β in hepatic fibrogene-
sis, a number of approaches to abrogate the effect of this
cytokine have been advanced. For example, neutralizing
antibodies (Jester et al., 1997; Yamamoto et al., 1999),
dominant negative TGF-β receptors (Qi et al., 1999) and
chimeric soluble TGF-β receptor (Yata et al., 2002) have
been shown to reduce fibrosis in a variety of models. In
this work a different approach was utilized to interfere
with TGF-β signaling; resveratrol, a non toxic and natu-
rally occurring compound, decreased TGF-β in animals
treated with CCl4 or vehicle accompanied by a complete
abrogation of CCl4-fibrosis. Resveratrol is a safer, easier
and cheaper way to interfere with the fibrogenic process,
at least in this model of liver cirrhosis.

42 E. CHAVEZ ET AL.
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
The NF-κB proteins are latent gene regulatory proteins
that lie at the heart of most inflammatory responses.
There are five NF-κB proteins in mammals (RelA, RelB,
c-Rel, NF-κB1 and NF-κB2), and they form a variety of
homodimers and heterodimers, each of which activates its
own characteristic set of genes (Karin et al., 2004). RelA
is a heterodimer of P50 and P65 and is responsible for
the production of many cytokines including TGF-β(Gressner and Weiskirchen, 2006). It is widely accepted
that resveratrol inhibits NF-κB activity (Tsai et al., 1999;
Pendurthi et al., 1999; Wadsworth and Koop, 1999).
Since NF-κB upregulates TGF-β it is possible that the
effects observed on TGF-β were mediated by this factor
and therefore account for the antifibrogenic properties of
resveratrol. Glycogen is the main source of energy in
liver and is also utilized to maintain blood glucose levels.
Chronic administration of CCl4 decreased importantly and
significantly the liver glycogen content. It is worth noting
that resveratrol preserved almost completely this para-
meter. Probably, resveratrol preserved glycogen by its
antifibrotic mechanism because fibrosis disrupts the nor-
mal architecture and blood flow into the liver therefore
not allowing nutrients to be absorbed by the hepatocytes.
In addition, necrosis was elevated by CCl4 and prevented
partially by resveratrol (as glycogen content), obviously
necrotic cells do not store glycogen.
Resveratrol has been reported to prevent oxidative
stress and lipid peroxidation processes (Frankel and
Waterhouse, 1993; Chanvitayapongs et al., 1997;
Ignatowicz and Baer-Dubowska, 2001). Therefore, lipid
peroxidation and glutathione were studied in our groups.
Unexpectedly, lipid peroxidation was not increased sig-
nificantly by CCl4, despite it being well known that this
agent is converted to trichloromethyl radical (CCl3•) that
reacts with the fatty acids of membranes producing lipid
peroxidation (Muriel, 1997; Muriel, 2007b). The most
likely explanation is that the dose used in chronic liver
damage is relatively small (0.4 g kg−1) compared with
the dose administered to produce acute liver damage
(4 g kg−1) (Muriel, 2007a); more importantly, after acute
liver damage, lipid peroxidation is increased after a few
hours of intoxication, while in this work it was measured
72 h after the last dose. Therefore, it is possible that
MDA was produced in small amounts and that it was
depurated within 72 h. These circumstances did not allow
the effect of resveratrol on lipid peroxidation to be tested.
Glutathione is a useful parameter for studying oxida-
tive stress in the cytosol of hepatocytes. The GSH/GSSG
ratio decreased by CCl4 but resveratrol was not able to
prevent this effect. These results are in apparent contrast
to those obtained by Sener et al. (2006c) who found that
resveratrol preserved GSH in ischemia/reperfusion-
induced oxidative injury in rats. The main difference is
that Sener et al. (2006c) utilized 30 mg of resveratrol
and this work used only 10 mg kg−1. In addition, no sig-
nificant changes in GSH were observed, and the diminu-
tion in the ratio GSH/GSSG was due to an increased
concentration of GSSG, which was slightly prevented by
resveratrol.
Leiro et al. (2004) investigated the effects of the cis
isomer of resveratrol on the responses of inflamma-
tory murine peritoneal macrophages. The results that
they obtained were similar to those generated by trans
resveratrol (the isoform used here) in different sys-
tems. Cis resveratrol at 100 μM decreased inducible nitric
oxide synthase and COX-2 in lipopolysaccharide and
interferon-γ treated macrophages. The Leiro et al. (2004)
results show that cis resveratrol significantly attenuates
several components of the inflammatory response to
proinflammatory stimuli. Furthermore, Leiro et al. (2005)
found that cis resveratrol has a significant modulatory
effect on the NF-κB signaling pathway. These results
suggest that the isomeric form of resveratrol may be not
relevant for their biological activity, although compara-
tive studies are necessary between both isomers.
Quercetin, a flavonoid has shown similar results to
those observed by resveratrol in the same model of liver
damage (Pavanato et al., 2003); however, the mechanism
of action seems to be different. Quercetin acts by increas-
ing total peroxyl radical-trapping antioxidant capacity of
liver, while resveratrol failed to prevent oxidative stress
but decreased the profibrogenic cytokine TGF-β. Anyway
both substances afforded beneficial effects on fibrosis and
other markers of liver damage.
In general we were unable to observe the antioxidant
properties of resveratrol in this model and at the dose
employed. However, resveratrol completely prevented
fibrosis and totally abolished the increase in TGF-β and
NF-κB (although this latter effect was not significant).
Therefore, in this model of fibrosis and at the dose
employed, resveratrol is an excellent antifibrogenic drug.
Furthermore, it seems that the protective effects of the
compound is more associated with its ability to reduce
the profibrogenic cytokine TGF-β, maybe by inactivating
NF-κB, than to its antioxidant properties.
Acknowledgements—The authors express their gratitude to Mr BenjamínSalinas Hernández, Mr Ramón Hernández and MVZ Ricardo Gaxiolafor their excellent technical assistance. IF Enrique Chavez was a fellowof Conacyt. This work was supported in part by Conacyt grant 42721(JS).
References
Abergel A, Sapin V, Dif N, Chassard C, Darcha C, Marcand-SauvantJ, Gaillard-Martinie B, Rock E, Dechelotte P, Sauvant P. 2006.Growth arrest and decrease of alpha-SMA and type I collagenexpression by palmitic acid in the rat hepatic stellate cell line PAV-1. Dig. Dis. Sci. 51: 989–995.
Aggarwal BB, Bhardwaj A, Aggarwal RS, Seeram NP, Shishoda S,Takada Y. 2004. Role of resveratrol in prevention and therapy ofcancer: preclinical and clinical studies. Anticancer Res. 24: 2783–2840.
Bataller R, Brenner DA. 2005. Liver fibrosis. J. Clin. Invest. 115: 209–218.

RESVERATROL PREVENTS LIVER FIBROSIS 43
Copyright © 2007 John Wiley & Sons, Ltd. J. Appl. Toxicol. 2008; 28: 35–43
DOI: 10.1002/jat
Bergmeyer HU, Grabl M, Walter HE. 1983. Enzymes. In Methods of
Enzymatic Analysis, Bergmeyer J, Grabl M (eds). Verlag-Chemie:Weinheim; 269–270.
Border WA, Ruoslahti E. 1992. Transforming growth factor-beta indisease: the dark side of tissue repair. J. Clin. Invest. 90: 1–7.
Bradford MM. 1976. A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of proteindye binding. Anal. Biochem. 72: 248–254.
Chanvitayapongs S, Draczynska-Lusiak B, Sun AY. 1997. Ameliorationof oxidative stress by antioxidants and resveratrol in PC12 cells.NeuroReport 8: 1499–1502.
Dong Z. 2003. Molecular mechanisms of the chemopreventive effect ofresveratrol. Mutat. Res. 523: 145–150.
Dyer RB, Hersog NK. 1995. Isolation of intact nuclei for nuclearextract preparation from a fragile B lymphocyte cell line.Biotechniques 19: 192–195.
Frankel EN, Waterhouse AL. 1993. Inhibition of human LDL oxidationby resveratrol. Lancet 341: 1103–1104.
Freemont L. 2000. Biological effects of resveratrol. Life Sci. 66: 663–673.
Friedman SL. 1993. The cellular basis of hepatic fibrosis. Mechanismsand treatment strategies. N. Engl. J. Med. 328: 1828–1835.
Garcia-Tovar C, Perez A, Luna J, Mena R, Osorio B, Aleman V,Mondragon R, Mornet D, Rendon A, Hernandez JM. 2001. Bio-chemical and histochemical analysis of 71 kDa dystrophin isoform(Dp 71f) in rat brain. Acta Histochem. 103: 209–224.
Ghosh S, May EB, Kopp EB. 1998. NF-kappaB and Rel proteins:evolutionarily conserved mediators of immune responses. Annu. Rev.
Immunol. 16: 225–260.Godichaud S, Krisa S, Couronne B, Dubuisson L, Merillon JM,
Desmouliere A, Rosenbaum J. 2000. Deactivation of cultured humanliver myofibroblast by trans-resveratrol, a grapevine-derivedpolyphenol. Hepatology 31: 922–931.
Godichaud S, Si-Tayeb K, Auge N, Desmouliere A, Balabaud C,Payrastre B, Negre-Salvayre A, Rosenbaum J. 2006. The grape-derived polyphenol resveratrol differentially affects epidermal andplatelet-derived growth factor signaling in human liver myofibroblast.Int. J. Biochem. Cell. Biol. 38: 629–637.
Gressner AM, Weiskirchen R. 2006. Modern pathogenetic concepts ofliver fibrosis suggest stellate cells and TGF-beta as major players andtherapeutic targets. J. Cell. Mol. Med. 10: 76–99.
Gressner AM, Weiskirchen R, Breitkopf K, Dooley S. 2002. Roles ofTGF-beta in hepatic fibrosis. Front Biosci. 7: 793–807.
Hellerbrand C, Stefanovic B, Giordano F, Burchardt ER, Brener DA.1999. The role of TGFbeta 1 in initiating hepatic stellate cell activa-tion in vivo. J. Hepatol. 30: 77–87.
Hissin PJ, Hilf R. 1976. A fluorometric method for determinationof oxidized and reduced glutathione in tissues. Anal. Biochem. 74:214–226.
Ignatowicz E, Baer-Dubowska W. 2001. Resveratrol, a naturalchemopreventive agent against degenerative diseases. Polish J.
Pharmacol. 53: 557–569.Jester JV, Barry-Lane PA, Petrol WM, Olsen DR, Cavanagh HD. 1997.
Inhibition of corneal fibrosis by topical application of blocking anti-bodies to TGF beta in the rabbit. Cornea 16: 177–187.
Karin M, Yamamoto Y, Wang QM. 2004. The IKK NF-κB system: atreasure trove for drug development. Nature Rev 3: 17–26.
Leiro J, Alvarez E, Arranz JA, Laguna R, Uriarte E, Orallo F. 2004.Effects of cis-resveratrol on inflammatory murine macrophages:antioxidant activity and down-regulation of inflammatory genes.J. Leukoc. Biol. 75: 1156–1165.
Leiro J, Arranz JA, Fraiz N, Sanmartin ML, Quezada E, Orallo F. 2005.Effect of cis-resveratrol on genes involved in nuclear factor kappa Bsignaling. Int. Immunopharmacol. 5: 393–406.
Massague J, Cheifetz S, Laiho M, Ralph DA, Weis FM, Zentella A.1992. Transforming growth factor-beta. Cancer Surv. 12: 81–103.
Muriel P. 1997. Peroxidation of lipids and liver damage. In Antioxi-
dants, Oxidants, and Free Radicals, Baskin SI, Salem H (eds). Taylor& Francis: Washington DC; 237–257.
Muriel P. 2007a. Some experimental models of liver damage. InHepatotoxicity: From Genomics to In Vitro and In Vivo Models,Sahu S (ed.). John Wiley & Sons: Chichester (in press).
Muriel P. 2007b. Cytokines in liver diseases. In Hepatotoxicity: From
Genomics to In Vitro and In Vivo Models, Sahu S (ed.). John Wiley& Sons: Chichester (in press).
Okawa H, Ohmishi N, Yagi K. 1979. Assay for lipid peroxides inanimal tissues by the thiobarbituric acid reaction. Anal. Biochem.
95: 351–358.Olson ER, Naugle JE, Zhang X, Bomser JA, Meszaros JG. 2005.
Inhibition of cardiac fibroblast proliferation and myofibroblast dif-ferentiation by resveratrol. Am. J. Physiol. Heart Circ. Physiol. 288:H1131–H1138.
Pavanato A, Tunon MJ, Sanchez-Campos S, Marroni CA, Llesuy S,Gonzalez-Gallego J, Marroni N. 2003. Effects of quercetin on liverdamage in rats with carbon tetrachloride-induced cirrhosis. Dig. Dis.
Sci. 48: 824–829.Pendurthi UR, Williams JT, Rao LV. 1999. Resveratrol, a polyphenolic
compound found in wine, inhibits tissue factor expression in vascu-lar cells: a possible mechanism for the cardiovascular benefits asso-ciated with moderate consumption of wine. Arterioscler. Thromb.
Vasc. Biol. 19: 419–426.Pérez-Severiano F, Escalante B, Vergara P, Ríos C, Segovia J. 2002.
Age-dependent changes in nitric oxide synthase activity and proteinexpression in striata of mice transgenic for the Huntington’s diseasemutation. Brain Res. 951: 36– 42.
Qi Z, Atsuchi N, Ooshima A, Takeshita A, Ueno H. 1999. Blockade oftype beta transforming growth factor signaling prevents liver fibrosisand dysfunction in the rat. Proc. Natl Acad. Sci. USA 96: 2345–2349.
Reitman S, Frankel S. 1957. A colorimetric method for determinationof serum oxaloacetic and glutamic pyruvic transaminases. Am. J.
Clin. Pathol. 28: 56–63.Rosen HR, Keeffe EB. 2000. Evaluation of abnormal liver enzymes,
use of liver test, and the serology of viral hepatitis. In Liver Disease
Diagnosis and Management, Bacon BR, Di Bisceglie AM (eds).Churchill: New York; 24–35.
Seifter S, Dayton S, Novic B, Muntwyler E. 1950. The estimation ofglycogen with the anthrone reagent. Arch. Biochem. 25: 191–200.
Sener G, Toklu HZ, Sehirli O, Velioglu-Ogunc A, Cetinel S, Gedic N.2006a. Protective effects of resveratrol against acetaminophen-induced toxicity in mice. Hepatol. Res. 35: 62– 68.
Sener G, Topaloglu N, Ozer Sehirli A, Ercan F, Gedik N. 2006b.Resveratrol alleviates bleomicin-induced lung injury in rats. Pulm.
Pharmacol. Ther. (in press). PMID: 17035056.Sener G, Tugtepe H, Yuksel M, Centinel S, Gedik N, Yegen BC.
2006c. Resveratrol improves ischemia/reperfusion-induced oxidativerenal injury in rats. Arch. Med. Res. 37: 822–829.
Tsai SH, Lin-Shiau SY, Lin JK. 1999. Suppression of nitric oxidesynthase and the down regulation of the activation of NF kappa B inmacrophages by resveratrol. Br. J. Pharmacol. 126: 673–680.
Wadsworth TL, Koop DR. 1999. Effects of the wine polyphenolicsquercetin and resveratrol on pro-inflammatory cytokine expression inRAW 264.7 macrophages. Biochem. Pharmacol. 57: 941–949.
Yamamoto T, Takagawa S, Katayama I, Nishioka K. 1999. Anti-sclerotic effect of transforming growth factor-beta antibody in a mousemodel of bleomycin-induced scleroderma. Clin. Immunol. 92: 6–13.
Yata Y, Gotwals P, Koteliansky V, Rockey DC. 2002. Dose-dependentinhibition of hepatic fibrosis in mice by a TGF-β soluble receptor:implications for antifibrotic therapy. Hepatology 35: 1022–1030.