response of selected wildlife to mesquite removal...
TRANSCRIPT

Response of selected wildlife tomesquite removal in desert grassland
Item Type text; Thesis-Reproduction (electronic)
Authors Germano, David Joseph
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 26/07/2018 18:46:53
Link to Item http://hdl.handle.net/10150/566644

RESPONSE OF SELECTED WILDLIFE TO MESQUITE
REMOVAL IN DESERT GRASSLAND
byDavid Joseph Germane
A Thesis Submitted to the Faculty of the
SCHOOL OF RENEWABLE NATURAL RESOURCES
In Partial Fulfillment of the Requirements • For the Degree of
MASTER OF SCIENCE WITH A MAJOR IN WILDLIFE ECOLOGY
In the Graduate College
THE UNIVERSITY OF ARIZONA
1 9 7 8

STATEMENT BY AUTHOR
This thesis has been submitted in partial fulfillment of requirements for an advanced degree at The University of Arizona and is deposited in the University Library to be made available to borrowers under rules of the Library.
Brief quotations from this thesis Eire allowable without special permission, provided that accurate acknowledgment of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in his judgment the proposed use of the material is in the interests of scholarship. In all other instances, however, permission must be obtained from the author.
SIGNED:
APPROVAL BY THESIS DIRECTOR
This thesis has been approved on the date shown below:
Professor of Wildlife EcologyV Date

ACKNOWLEDGMENTS
This study was funded by the U.S. Forest Service, Rocky Moun
tain Forest and Range Experiment Station. It was under the supervision
of Dr. S. Clark Martin of the Forest Science Laboratory, Tempe,
Arizona.
I thank my friend and advisor, Dr. C. Roger Hungerford, for
his guidance throughout this study and for his faith in my abilities.
Also, his help with all phases of writing of this thesis is deeply
appreciated.
I thank Dr. Norman S. Smith and Dr. S. Clark Martin for
serving on my committee and for critically analyzing my thesis.
I also thank Dr. Robert 0. Kuehl, Statistician, Remote Sensing
Laboratory, University of Arizona, and Alan R. Fisher, a graduate stu
dent with the Arizona Cooperative Wildlife Research Unit, The Univer
sity of Arizona, for their assistance in analyzing my data and
suggesting correct statistical tests. I also wish to acknowledge
Patrick Schlecht who began this project.
Lastly, I thank my wife, Rhonda, for her love and help. She
typed many drafts of this thesis and assisted in the field. Her
moral support throughout this study is gratefully acknowledged.
iii

TABLE OF CONTENTS
LIST OF TABLES................................................ vi
LIST OF ILLUSTRATIONS.............. yii
ABSTRACT................................. viii
INTRODUCTION .................................................. 1
DESCRIPTION OF STUDY A R E A .................................... 5
Location.............. 5Vegetation . . . . . . . ........................... • • • 5Climate . ........ . . . . . . . . . . . . ........ • » 11S o i l s .................................................... 14G r a z i n g ............ 14
METHODS ....................................................... 15
Visual C e n s u s ............ 15Pellet Counts . . . . . . . . . . . . . . . . . . . . . . 18Call C o u n t s ...................... 19Quantifying E d g e ......................................... 19Data A n a l y s i s ............................................ 19
R E S U L T S ...................................................... 21
Visual Census . . . . . ............. . . . . . . . . . . 21Pellet and Scat Counts . . . . . . . . . . . . . . . . . . 23Call C o u n t s ....................................... 23Use of Openings . . . . . . . . . . . . . . . . . . . . . 26Cattle Influence on Wildlife Sightings ................. . 26Quantifying E d g e .......................................... 29
DISCUSSION.................................................... 30
Desert Mule Deer . . . . . . . . . . . . . . . . . . . . . 30Javelina ............. . ............. . . . . . . . . . . 31Coyote.................................................... 32Black-tailed Jackrabbit ................................. 35Antelope Jackrabbit ...................................... 3^Desert Cottontail . . . . . . . . . . . . . . . . . . . . 35
Page
iv

V
TABLE OF CONTENTS— Continued
Page
Mourning D o v e s ................................... .. 36White-winged Doves . . . . .......... . . . . . . . . . . 37Scaled Quail ........................... .............. .. 38Gambel* s Quail . . . .Reptiles . ...........
Zebra-tailed LizardDesert Spiny Lizard ........ . ......... . . . . . . 4lLesser Earless L i z g r d ........... . 4lTree Lizard ................. 42Western Whiptail ............. . . . . . . . . . . . . 42Sonora Spotted Whiptail ................. . . . . . . 43All Reptiles.................... 44Unidentified Lizards . . . . . . . . . . . . . . . . . 44
Other Species ................. . . . . . . . . . . . . . 45
SUMMARY AND CONCLUSIONS ..................................... 46APPENDIX A: STATISTICAL ANALYSIS OF VISUAL CENSUS.......... 48
APPENDIX B: STATISTICAL ANALYSIS OF PELLET C O U N T .......... 50
APPENDIX C: STATISTICAL ANALYSIS OF CALL COUNT D A T A ........ 51
APPENDIX D: STATISTICAL ANALYSIS OF VISUAL CENSUS WITH ORWITHOUT THE PRESENCE OF C A T T L E ................. 52
LIST OF REFERENCES........................................... 53
SSiS

LIST OF TABLES
1. The number of mammalst birds, and reptiles sightedduring the visual census of the three pastures . . . . . 22
2. Total number of pellets and scats counted at three monthintervals from December 1976 to June 1978 ........ .. 24
3» Call counts of doves and quail at two week intervalsfrom April to June, 1977 and 1978 . . . . . . . . . . . 25
4. The number of mammals, birds, and reptiles sighted in the two habitat types of pasture 22 from September 1976to June 1978 . . ........................................ 27
5. The total number of mammals and birds sighted for thethree pastures when cattle were absent or present . . . 28
Table Page
vi

LIST OF ILLUSTRATIONS
1. The desert grasslands of the Southwest ............... 2
2. Location of the Santa Rita Experimental R a n g e ........ 63. Relative size and position of the three pastures used
in this s t u d y ..................... ............. 74. Typical vegetation of pasture 2S (undisturbed mesquite) 85. Typical vegetation of pasture 1 (mesquite-free) . . . . 10
6. Clearing B (top) and mesquite (bottom) directly opposite clearing B showing typical vegetation ofpasture 2 2 ................................. 12
7. Clearing E (top) and mesquite (bottom) directly opposite clearing E showing typical vegetation ofpasture 2 2 .............................................. 13
8. Relative size and placement of the seven clearingscreated in pasture 2 2 .................................. 16
9. Placement of transect lines in each p a s t u r e ............ 17
Figure Page
vii

ABSTRACT
I studied selected wildlife on an undisturbed mesquite range,
a mesquite-free range, and a mesquite range with clearings on the
Santa Rita Experimental Range. I visually censused six mammals and
four birds monthly from September 1976 to June 1978. I visually cen
sused six lizards monthly from April to September 1977 and from April
to June 1978. I counted pellets and scats every three months starting
December 1976. I counted calls of doves and quail every two weeks
from April to June, 1977 and 1978.
The mesquite with clearings contained significantly more
black-tailed jackrabbits, antelope, jackrabbits, gambel’s quail,
western whiptails, and all reptiles sighted than the mesquite-free
range. The undisturbed mesquite contained significantly more black
tailed and antelope jackrabbits, zebra-tailed lizards, western whip-
tails, and all reptiles sighted than the mesquite-free range. I
counted significantly more lagomorph pellets on the mesquite areas
than the mesquite-free range. I heard significantly more calls in
the mesquite ranges.
Creating clearings in mesquite is not detrimental to the
selected wildlife I studied. Totally clearing mesquite from ranges
is harmful to wildlife in some instance.
viii

INTRODUCTION
Desert grasslands are primarily found in southeastern Arizona,
southcentral and southwestern New Mexico and southwest Texas (Humphrey
1958) (Fig# 1). Sauer (1950) indicates these grasslands are found in
areas with dry seasons or occasional short periods of dry weather
during which the ground cover dries out and in areas of smooth to roll
ing land surfaces. According to Humphrey (1958) the desert grasslands
occur as local grasslands, not being interspersed widely with other
types. He further typifies them as lying in broad belts around the
bases of southwestern mountain ranges principally at 3,000 to 3,500 feet in elevation.
Within the last century, mesquite (Prosopis juliflora) has in
vaded the desert grassland (Brown 1950; Parker and Martin 1952; Martin
and Reynolds 1975; Martin 1975)• Originally, mesquite was confined
mostly to bottomlands and drainages (Reynolds and Martin 1968). The
mesquite invasion has coincided with the influx of White men into the
Southwest and may partially be due to fire suppression and cattle
overgrazing (Brown 1950; Sauer 1950; Parker and Martin 1952; Humphrey
1958; Ellison i960. By 1952 Parker and Martin thought mesquite was a
problem in Arizona, New Mexico and Texas and that about seventy million
acres had been invaded. As the mesquite increased on the range, the
amount of grass cover decreased (Parker and Martin 1952; Caraher 1970;
Martin 1975)• Parker and Martin (1952) and Dahl, Sosebee and Goen
1

Figure 1. The desert grasslands of the Southwest. — Adapted from Brown and Lowe 1977•
rv

3(197*0 also found that during dry years grass did better in areas where
mesquite was controlled. Because much of the Southwest is grazed by
cattle, today many areas are being cleared of mesquite to help promote
grass growth.
Most of Arizona's ranges are publicly owned lands and are
managed for multiple use. One of the implications of multiple use is
that the needs of wildlife should be considered when planning the use
of rangeland (Arizona Interagency Range Committee 1973» 1975)• Grazing
land is seen then as land producing natural vegetation which can be
grazed by domestic cattle, as habitat for wildlife, as a watershed, or
for aesthetic purposes (Arizona Interagency Range Committee 1972).
Total eradication of mesquite which would benefit cattle could
be harmful to wildlife. Total tree or brush eradication has been shown
to be harmful to deer (Davis and Winkler 1968; McCulloch 1972; Whitson,
Beasom and Scifres 1977)• On the Santa Rita Experimental Range in
Arizona, a study by McCormick (1975) showed lower numbers of gambol's
quail, scaled quail, white-winged doves, mourning doves, desert cotton
tails and black-tailed jackrabbits in mesquite cleared range than in
undisturbed mesquite range.
Partial eradication of mesquite on invaded grasslands which
are publicly owned may benefit both cattle and wildlife. On privately
owned ranges, this compromise may also be the best solution. Clearing
strips of mesquite may benefit Texas cattle ranchers because of the
money they can receive from hunters using their rangeland (Whitson
et al. 1977)• Private land ownership and wildlife may be compatible
according to Martin (1975, p. 26), "If beef is the only salable ranch

k
product, complete clearing may be a reasonable objective. On the other
hand, if opportunities to develop income from hunting privileges or
other recreational enterprises are good, it may be desirable and
profitable to leave patches of mesquite for food and cover.” Estab
lishing irregular shaped clearings in mesquite should enhance grass
production while still retaining wildlife habitat. Edge will be in
creased by this method. Leopold (1953) notes the desirability of
wildlife having simultaneous access to more than one environment.
Wildlife numbers may in fact increase because of the clearings.
The purpose of this study is to monitor the response of se
lected wildlife.species to the creation of irregular shaped clearings
in mesquite on desert grassland range.

DESCRIPTION OF STUDY AREA
Location
The study area is located in the southeast corner of the Santa
Rita Experimental Range. The range is located in the semidesert grass
land biotic community (Brown and Lowe 1977)• It is approximately 48
km. south of Tucson, Arizona (Fig. 2)• Three pastures were used for
this study: pasture 22, pasture 1 and pasture 2S (Fig. 3)• Elevations
ranged from 1170 meters to 1300 meters (Martin 1966).
Vegetation
Plant names follow Nickerson, Brink and Feddema (1976) • The
major shrub components of the Santa Rita Experimental Range are mes-
quite, cacti (Opuntia spp.) and burroweed (Haplopappus tenuisectus)
(Martin and Reynolds 1973)• Pasture 2S is an undisturbed mesquite range (Fig. 4). The major shrubs are mesquite with an understory of
burroweed, Englemann pricklypear (Opuntia englemannii) and staghorn •
cholla versicolor and 0. spinosior). Other shrubs found in lesser
numbers are false mesquite (Calliandra eriophylla), catclaw (Acacia
greggii) (personal observation), desert zinnia (Zinnia pumila), long-
leaf ephedra (Ephedra trifurea), blue paloverde (Cercidium floridum)
and Arizona carlowrightia (Carlowrightia arizonica) (U.S. Forest Ser
vice n.d.). Forbs are seasonal, growing mostly in the July to August
rainy season. Forbs include ragweed (Ambrosia spp.), indianroot
5

6
ARIZONA
SANTA CATALINA MOUNTAINS
RINCONMOUNTAINS
TUCSON
SANTA RITAEXPERIMENTALRANGE
SANTA RITA MOUNTAINS
Figure 2. Location of the Santa Rita Experimental Range,

V
-x3
Figure 3. Relative size and position of the three pastures used in this study.

8
Figure 4. Typical vegetation of pasture 2S (undisturbed mesquite)•

9(Aristolochia watsonii), spiderling (Boerhaevia spp.), Arizona
evolvulus (Bvolvulus arizonicus), sida (Sida spp.) and silverleaf
nightshade (Solanum elaeagnifolium) (U.S. Forest Service, n.d.).
Grasses are of lowest densities in this pasture. Sixteen species were
recorded on pasture 25 with Santa Rita threeawn (Aristida glabrata),
Rothrock gramma (Bouteloua rothrockii) and Lehmann's lovegrass
(Eragrostis lehmanniana) being dominate grasses (U.S. Forest Service
n.d.).
Pasture 1 is an almost mesquite-free range (Fig. 5)• Between
1954 and 1957, this range was cleared of mesquite (Cable and Martin
1975)• Very light stands of mesquite still remain. Shrubs also found
in pasture 1 in low numbers are burroweed, Whitehall acacia (Acacia
angustissima), agave (Agave spp.), false mesquite, Wright's buckwheat
(Briogonum wrightii), ocotillo (Fouquieria splendens), littleleaf
krameria (Krameria parvifolia), mammillaria (Mammillaria spp.), cat-
claw mimosa (Mimosa biuncifera) and velvetpod mimosa (M. dysocarpa)
(U.S. Forest Service n.d.). Forbs present include ragweed, spiderling,
Arizona evolvulus, sida, silverleaf nightshade, trailing four o'clock
(Alliopia spp.), milkvetch (Astragalus spp.), thistle (Cirsium spp.),
sacred datura (Datura metaloides), spurge (Euphorbia spp.), wire let
tuce (Stephanomeria spp.) and threadleaf groundsel (Senicio longilobus)
(U.S. Forest Service n.d.). Grasses dominate this pasture and are
quite dense. Nineteen species have been recorded, of which Lehmann's
lovegrass, slender gramma (Boutelous filiformis), sideoats gramma (B.
curtipendula) and hairy gramma (B. hirsuta) predominate (U.S. Forest
Service n.d.).

10
IPf W - * . V «*
c : : '
•:4 ..4V■ 2 ^ . *, A #
" # ••
w#' BeBE . \
Figure 5* Typical vegetation of pasture 1 (mesquite-free)

11Pasture 22 is a mosaic area. Mesquite dominates areas not
cleared with large numbers of burroweed underneath. Also present in
lower numbers are Englemann pricklypear, staghorn cholla, false raes-
quite, velvetpod mimosa, catclaw (personal observation), littleleaf
krameria, Arizona carlowrightia and desert zinnia (U.S. Forest Service
n.d.). Forbs include ragweed, milkvetch, spiderling, Arizona evolvulus,'
sida and silverleaf nightshade (U.S. Forest Service n.d.). Grasses are
sparse under the mesquite but become more dense in the clearings (Figs.
6 and 7)• Nineteen species were recorded, of which Santa Rita threeawn,
hairy gramma, Rothrock gramma and curlymesquite (Hilaria belangeri)
are the dominate species (U.S. Forest Service n.d.).
Climate
The climate of the Santa Rita Experimental Range is typical of
the arid Southwest with low relative humidity and daily temperatures
sometimes exceeding 38°C during the summer (Reynolds and Martin 1968).
About one half of the precipitation comes between July and September,
two fifths between October and March and the rest during the dryest
part of the year from April to June (Reynolds and Martin 1968). Aver
age rainfall for the lower elevations of the study site is 36 cm. up to 43 cm. for the higher elevations (Martin 1966). Recorded rainfall
from five rainfall gauges near the study site averaged 32.99 cm. from July, 1975 to June, 1976, 33.12 cm. from July, 1976 to June, 1977 and 43.69 cm. from July, 1977 to June, 1978 (U.S. Forest Service n.d.).

Figure 6. Clearing B (top) and mesquite (bottom) directly opposite clearing B showing typical vegetation of pasture 22.

13

14Soils
Similar soils made up the three ranges. The 310 hectares of
pasture 1 are made up of Comoro (l-59» slope), White House (0-1Q& slope),
Caralampi (10-4096 slope) and Lampshire-Chiricahua (l5-509» slope) soils;
the 311 hectares of pasture 22 are made up of Comoro, White House and
Sonoita soils (1-8% slope); the 1426 hectares of pasture 2S are mostly
Comoro, White House, Sonoita and Continental (1-10% slope) soils
(Clemmons and Wheeler 1970)•
Grazing
A rotational grazing schedule with moderate grazing was fol
lowed on all three pastures. About fifty head were grazed in pastures
22 and 1. Pasture 2S, which was larger, was grazed by 170 animals.
Cattle were in pasture 22 from the end of October, 1976 until
the end of February, 1977 and from March, 1978 until the end of the
study. Cattle were in pasture 2S from March 1977 until the end of
October, 1977 and in pasture 1 from March, 1977 until the end of
November, 1977*

METHODS
To create clearings in the mesquite on pasture 22, the area was
chained from 7-10 July, 1976. Two bulldozers dragged a ship’s anchor
chain between them and knocked over mesquite. Seven clearings of dif
ferent size and shape were created (Fig. 8). Clearing A is 18.6 hec
tares, B is 12.2 hectares, C is 2.8 hectares, D is 2.8 hectares, E is 30.4 hectares and F is 7.7 hectares. Sometimes mesquite which was
knocked over congregated to create "brush piles." Individual trees
were usually left where they fell. Pasture 25 was used as an experi
mental range of undisturbed mesquite. Pasture 1 was used as a control
and is an almost mesquite-free range. In each pasture four transect
lines, each 1200 meters long, were established (Fig. 9) perpendicular
to the major drainage channels.
Visual Census
The mammals and birds censused were selected because of their
economic importance and relative abundance. The reptiles censused
were selected because of their abundance. Also, much work has been
done on the species chosen and a large amount "of literature is avail
able to review. Although many other species exist on this range, the
mammals, birds and reptiles censused may provide insight into the re
sponse of other wildlife species to the treatments.
I visually censused mourning doves (Zenaida macroura), white
winged doves (Z. asiatica), gambel's quail (Lophortyx gambelii),
15

16
PASTURE 22
Figure 8. Relative size and placement of the seven clearings created in pasture 22.
\
17
PASTURE 1PASTURE 26 PASTURE 22
TRANSECT LINES
PASTURE BOUNDARIES
Figure-9. Placement of transect lines in each pasture.

18scaled quail (Callipepla squamata), desert mule deer (Odocoileus
hemionus), javelina (Dicotyles tajacu), coyote (Canis latrans)t black
tailed jackrabbit (Lepus californicus), antelope jackrabbit (L. alleni)
and desert cottontail (Sylvilagus auduboni). I" did this each month
from September, 1976 to June, 1978. .1 began walking transect lines in
the morning when it became light enough to see. I censused only two
lines each morning.
I also visually censused reptiles monthly from April, 1977 to
September, 1977 and from April, 1978 until June, 1978. The time that
I censused reptiles (mainly lizards) varied each month. Pianka (1970)
notes that whiptails (Cnemidophorus spp.) exhibit unimodal activity in
the spring and bimodal activity in the summer. This is directly re
lated to soil temperatures. Milstead (1957) found their activity began
in the morning when soil temperatures were 28-30*0 and ended when soil temperatures reached 50-52*0. The same is true of many other lizards.
I used 7 x 35 binoculars to identify all wildlife seen.
Pellet Counts
Every three months I counted pellets and pellet groups. Ten
stakes were placed at 6 chain intervals in each transect line. Per
pendicular to the line at each stake on each side, an additional stake
was placed approximately 2 chains out. Thus, there were thirty stakes
for each transect line and one hundred and twenty stakes for each
pasture. D. J. Neff (1968) noted that for general coverage of an area
it is best to use a series of circular plots. At each stake a circular
plot of .001 acre was used for lagomorph pellets and a .01 acre plot

19was used for deer and other scats. I followed the guidelines of Smith,
Neff and McCulloch (1969) for searching pellet plots. Pellets were
removed from each plot after they were counted. Deer pellets were
counted as groups and all others were counted individually. No accept
able method was found to distinguish between the three lagomorphs.
Call Counts
Between April and June, 1977 and 1978, I counted calls from
gambel1s quail, scaled quail, mourning doves and white-winged doves.
These months are similar to times other Arizona investigators have
used to census calls (Gallizioli 1961; Kufeld and Fitzhugh 1962; Brown,
Cochran and Waddell 1978) • I counted calls at two week intervals. An
often used technique described by Gallizioli and Smith (1962) involves
driving roads and stopping every mile and listening for three minutes.
Instead of driving, I walked each transect line with a map of the area
and wrote down calls continuously for each pasture. I walked the
transect lines and recorded number of calls six times a year in each
pasture.
Quantifying Edge
The amount of edge in each pasture compared to a circle of the
same area was determined by the method developed by Patton (1975).
Data Analysis
Because the visual census data were not normally distributed,
I used a method (Bhapkar 1968) which categorizes the data into counts
per transect to test for significant differences (P<.05). Each

20treatment involved eighty-eight transect counts. To test significance
between treatmentst I used the "least significant difference" method
(P <.05) (Steel and Torrie I960).
Deer pellet data were not normally distributed. I used Bhapkar
(1968) to test for significant differences (P <.05)• The lagoraorph
pellet counts were normally distributed and I used a four way analysis
of variance (P <.05) (Nie et al. 1975) to test for significant differ
ences. I tested for differences among factors with the least signifi
cant difference method (P<.05) (Steel and Torrie I960).
Call count data were poisson distributed. I used a one way
analysis of variance ( P < .05) (Steel and Torrie i960), using the square root of the data to determine significant differences. I determined
significant differences between treatments by the least significant
difference method (P <.05) (Steel and Torrie i960).To test for differences between the number of animals visually
censused in the mesquite and the number in the clearings of pasture
22, I used the nonparametrie sign test (P <.05) (Steel and Torrie I960). I used the sign test for deer pellet groups and the number of plots
with cattle droppings. To test for differences in lagomorph pellets
between these two habitat types, I used a paired t-test (P <.05)
(Steel and Torrie i960).To test for significant differences in the numbers of animals
visually censused when cattle were present on the pastures and when
cattle were absent, I used the method by Bhapkar (1968) (P <.05)•

RESULTS
Visual Census
The numbers of desert mule deer, javelina, coyotes, mourning
doves, scaled quail, desert spiny lizards (Sceloporus magister), lesser
earless lizards (Holbrookia maculata) and unidentified lizards seen
showed no significant differences (P <.05) between treatments. Not
enough data were collected for desert cottontail and white-winged
doves to test for significant differences (Table 1).
The mesquite with clearings range contained significantly more
black-tailed jackrabbits, antelope jackrabbits, gambel’s quail, western
whiptails (Cnemidophorus tigris) and all reptiles sighted (Table 1)
than the mesquite-free range.
The mesquite with clearings range contained significantly more
tree lizards (Urosaurus ornatus) and sonora spotted whiptails
(Cnemidophorus sonorae) sighted (Table 1) than the undisturbed mesquite
range.
The undisturbed mesquite range contained significantly more
black-tailed jackrabbit, antelope jackrabbit, zebra-tailed lizards
(Callisaurus draconoides), western whiptails and all reptiles sighted
(Table 1) than the mesquite-free range.
The undisturbed mesquite range contained significantly more
zebra-tailed lizards sighted (Table 1) than the mesquite with clear
ings range.
21

22• Table 1. The number of mammalst birds, and reptiles sighted during
the visual census of the three pastures.*
Species
Vegetative CharacteristicsMesquite with
ClearingsUndisturbed
MesquiteMesquite-
free
Desert mule deer 27 33 18Javelina 17 26 0Coyote 7 5 iBlack-tailed Jackrabbit 38a 39° 6Antelope Jackrabbit 59a 39° 10Desert cottontail 2 4 3Mourning dove 399 59 108White-winged dove 0 1 1Scaled quail 36 32 24Gambel's quail 77a 22 1Zebra-tailed lizard 20 6lc,d 0Desert spiny lizard 5 12 0Lesser earless lizard 31 18 22Tree lizard 42b 16 22Western whiptail 210a 303° 3Sonora spotted whiptail 53b 8 70*Unidentified lizards 4l 30 55All reptiles 422a 468c 187
♦Significant differences (P <.05) among treatments determined by the least significant difference method.
^Significant difference between the mesquite with clearings and the mesquite-free pasture.
^Significant difference between the mesquite with clearings and the undisturbed mesquite pasture.^Significant difference between the undisturbed mesquite and the mesquite-free pasture.^Significant difference between the undisturbed mesquite and the mesquite with clearings pasture.^Significant difference between the mesquite-free pasture and the undisturbed mesquite pasture.

23The mesquite-free range contained significantly more sonora
spotted whiptails sighted (Table l) than the undisturbed mesquite
range. The statistical analysis of the visual census data are shown
in Appendix A.
Pellet and Scat Counts
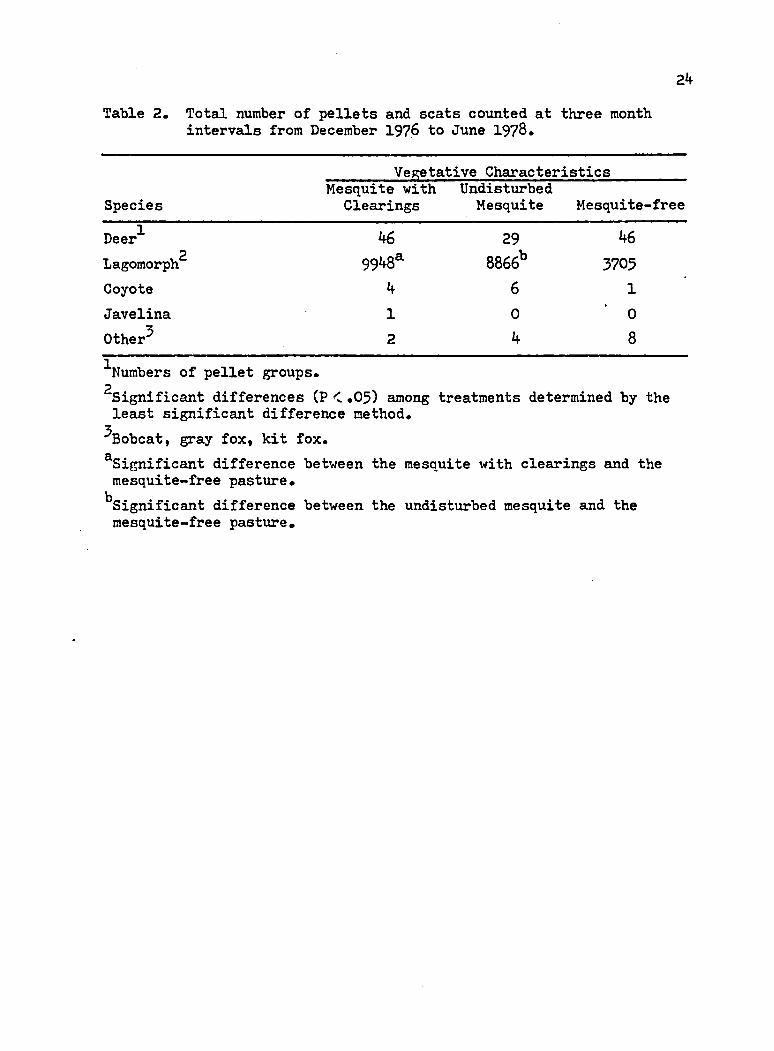
The number of deer pellet groups counted were not significantly
different (P <.05) between treatments. Not enough scats were collected
for coyotes or javelina to test for significant differences (Table 2).
Both the mesquite with clearings and the undisturbed mesquite
ranges contained significantly more lagomorph pellets than the
mesquite-free range (Table 2). The statistical analysis of the pellet
and scat counts are shown in Appendix B1 and B2.
Call Counts
No significant differences (P <.05) in the number of scaled
quail heard calling were found between pastures (Table 3)• Signifi
cantly more calls were heard in the mesquite with clearings range than
the mesquite-free range for mourning doves and gambel* s quail (Table J>).
Significantly more calls were heard in the mesquite with clearings
range than the undisturbed mesquite range for gambel*s quail (Table 3). Significantly more calls were heard in the undisturbed mesquite range
than the mesquite-free range for mourning doves, white-winged doves
and gambel* s quail (Table 3)• The statistical analysis of call count
data are shown in Appendix C.
24
Table 2. Total number of pellets and scats counted at three month intervals from December 1976 to June 1978.
________ Vegetative CharacteristicsMesquite with Undisturbed
Species Clearings Mesquite Mesquite-free
Deer'*' 46 29 462Lagomorph 9948* 8866b 3705Coyote 4 6 1Javelina 1 0 0Other** 2 4 8^Numbers of pellet groups.^Significant differences (P <.05) among treatments determined by the least significant difference method.
^Bobcat, gray fox, kit fox.^Significant difference between the mesquite with clearings and the mesquite-free pasture.“Significant difference between the undisturbed mesquite and the mesquite-free pasture.

25Table 3* Call counts of doves and quail at two week intervals from
April to June, 1977 and 1978.*
________ Vegetative CharacteristicsMesquite with Undisturbed
Species Clearings Mesquite Mesquite-free
Mourning dove 194* 268c 105White-winged dove 22 55c 11Scaled quail 43 61 16Gambel's quail llla,b 52c 1
•Significant differences (P <.05) among treatments determined by the least significant difference method.^Significant difference between the mesquite with clearings and the mesquite-free pasture.
^Significant difference between the mesquite with clearings and the undisturbed mesquite pasture.^Significant difference between the undisturbed mesquite and the mesquite-free pasture.
#

2 6
Use of Openings
The numbers of all the selected mammals, birds and reptiles
(except for the zebra-tailed and tree lizards) sighted in the mesquite
were not significantly different (P <.05) than the clearings of pasture
22 (Table 4).
The tree lizard was seen significantly more often in the
clearings than the mesquite of pasture 22 (Table 4). The zebra-tailed
lizard was seen significantly more often in the mesquite than the
clearings of pasture 22 (Table 4).
I counted seven deer pellet groups from the clearings and 39
groups from the mesquite of pasture 22. Deer pellet groups were sig
nificantly higher (P <.05) in the mesquite than the clearings (using
the sign test). I counted 5138 lagomorph pellets in the clearings and
4810 lagomorph pellets in the mesquite of pasture 22. Lagomorph
pellets were not significantly different (PC.05) between the mesquite
and the clearings of pasture 22 (using the paired t-test). Eighty-four
plots in the clearings contained cattle droppings and sixty plots in
the mesquite contained cattle droppings. No significant difference
(P•< 05) was seen in the number of plots with cattle droppings between the two habitat types of pasture 22 (using the sign test).
Cattle Influence on Wildlife Sightings
I did not see any significant differences (P <.05) in the num
ber of selected birds and mammals sighted when cattle were present or
absent from the pastures (Table 5). Cattle grazed rotationally on each
pasture. Also, cattle grazing was only moderate. Other systems of
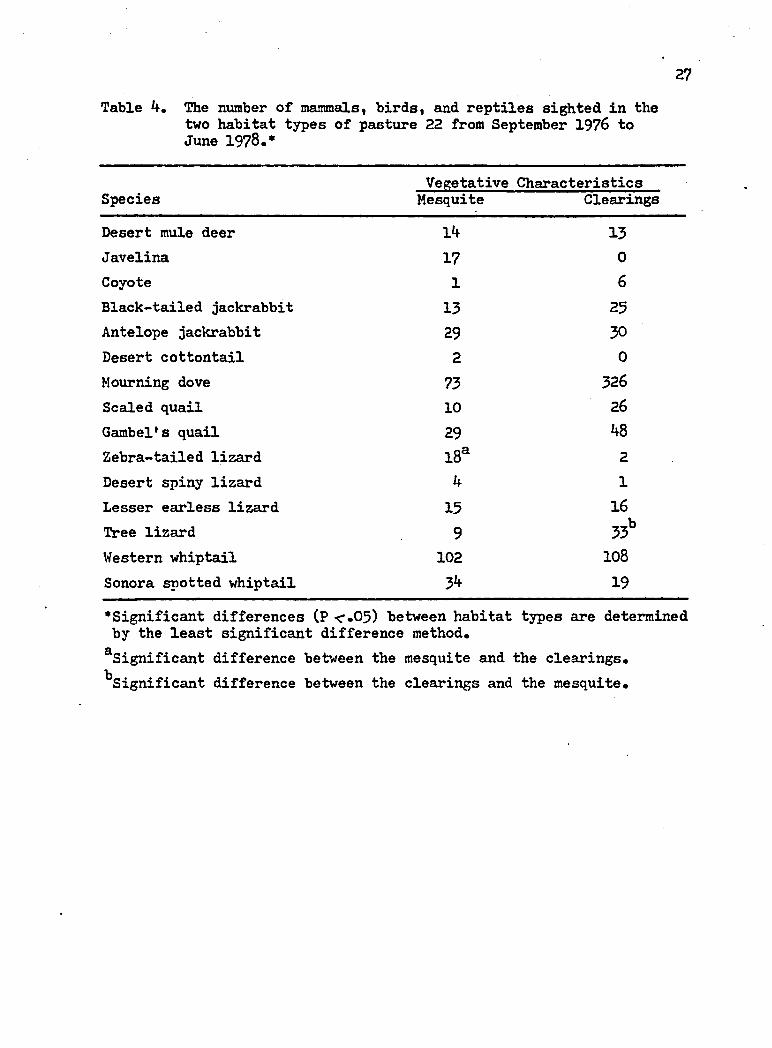
27Table 4. The number of mammalst birds, and reptiles sighted in the
two habitat types of pasture 22 from September 1976 to June 1978.*
Vegetative CharacteristicsSpecies Mesquite Clearings
Desert mule deer 14 13Javelina 17 0Coyote 1 6Black-tailed jackrabbit 13 25Antelope jackrabbit 29 30Desert cottontail 2 0Mourning dove 73 326Scaled quail 10 26Gambel's quail 29 48Zebra-tailed lizard l8a 2Desert spiny lizard 4 1Lesser earless lizard 15 16Tree lizard 9 33bWestern whiptail 102 108Sonora spotted whiptail 34 19
•Significant differences (P <.05) between habitat types are determined by the least significant difference method.^Significant difference between the mesquite and the clearings."Significant difference between the clearings and the mesquite.

28Table 5- The total number of mammals and birds sighted for the three
pastures when cattle were absent or present.
Species Cattle Absent Cattle Present
Desert mule deer 65 13Javelina 20 23Coyote 6 7Black-tailed jackrabbit 49 34Antelope jackrabbit . 77 31Mourning dove 468 98Scaled quail 57 35Gambel’s quail 66 34

29cattle grazing should be studied and for a longer period of time to
understand more fully cattle's effect on wildlife in mesquite con
trolled habitats. The statistical analysis of cattle's influence on
wildlife is shown in Appendix D.
Quantifying Edge
Pasture 1 (mesquite free) has a perimeter of 8,986 meters and
an area of 310 hectares. Its diversity index (Patton 1975) is 1.44.
Pasture 25 (undisturbed mesquite) has a perimeter of 15t789 meters and
an area of 1,426 hectares. The diversity index for pasture 2S is l.l8.
Pasture 22 (mesquite with clearings) has a total perimeter of 16,388
meters (including clearings) and an area of 311 hectares. Its diver
sity index is 2.62. Two species of low radius and mobility were found
in greater numbers in the pasture with the highest diversity index.
Gambel's quail numbers in the mesquite with clearings were 2.85 times
those in the undisturbed mesquite and 77 times the numbers in the
mesquite-free range. Tree lizard numbers in the mesquite with clear
ings were 2.62 times those in the undisturbed mesquite and 1.91 times those in the mesquite-free range.

DISCUSSION
The overall effect of spot clearing in mesquite habitat was
not detrimental to selected wildlife and was beneficial in some in
stances. The number of indicator mammalst birds and reptiles visually
consused was generally higher than the number counted in the mesquite-
free range and at about the same level as the undisturbed mesquite
range. Call counts indicate similar results. Lagomorph pellet counts
also indicate a higher population in the mesquite with clearings than
the mesquite-free range.
Desert Mule Deer
Visual censusing showed no significant difference between pas
tures , nor did counts of pellet groups indicate any difference. This
may in part be due to the size of their home range. Rodgers (1977)«
working on the Santa Rita Experimental Range, found mule deer had an
average home range size of 2.9 square miles. This may partially be
the reason an equivalent number of mule deer were seen in the mesquite-
free range as in the other two ranges. Mule deer could easily travel
into the mesquite-free area and not be very- far from mesquite. In
effect, because of their size and mobility, the whole mesquite-free
range could be considered a clearing in a mesquite habitat. If the
mesquite-free range was much larger, it would probably not receive
much deer use.
30

31Mule deer diet was found to include only a trace to 2.6#
grasses during the year but 31«7 to 48.0# shrubs and half shrubs, 14.1
to 55.9# cacti fruit and, in July and August, 24.1# mesquite (Short
1977). Mesquite, catclaw acacia and mountain hackberry are utilized
heavily by mule deer during droughts (Anthony 1976). In southeastern
Arizona, Anthony and Smith (1977) found mule deer to be associated with
a habitat type of mesquite and ocotillo. I did not find a significant
difference between sightings of mule deer in the clearings or the mes
quite of pasture 22, but I did find significantly more pellet groups
in the mesquite than the clearings. This may be related to the time
deer spent feeding in different areas. Although my study did not indi
cate any preference by mule deer, mesquite and associated shrubs and
cacti are prime habitat. Short (1977) thinks extensive destruction of
mesquite would be detrimental to deer habitat. I think opening only
clearings in mesquite as a compromise would not harm desert mule deer
and may attract them due to an increase in forb production.
Javelina
I found no significant difference in the number of javelina
visually censused, and scat counts did not provide enough data to test
for significance. The results from the test on visual census data are
somewhat misleading and may be a result of the nature of the test. I
saw 17 javelina in the mesquite with clearings from one herd and 26 javelina from several sightings in the undisturbed mesquite range. I
did not see any javelina nor any signs of their presence in the
mesquite-free range. Javelina have a smaller home range size than

32
mule deer. Schweinsburg (1971) found Arizona home ranges of 0.2, 0.3,
0.5 and 0.6 square miles. Ellisor and Harwell (1969) found home ranges
of 311 and 548 acres in Texas mesquite. Jennings and Harris (1953)
think Texas javelina home ranges do not exceed 3/4 of a mile, but Sowls
(1974) indicates home ranges vary from 0.2 to 1.5 square miles in
variable Arizona habitats.
I found no sign of javelina using the mesquite-free pasture as
part of their home range. Knipe (1957) cites mesquite thickets as a
favorite habitat, with washes being preferred. Dawson (1950) stated
preferred habitat is pricklypear and mesquite covered pastures. On the
Santa Rita Experimental Range, Eddy (1961) found that pricklypear and
century plants (Agave spp.) are the most important foods with seasonal
supplements of mesquite beams, amnual forbs and barrel cactus fruits.
Grasses are not important in the diet (Knipe 1957i Eddy 1961). Clado-
phylls of pricklypear are especially important in late spring and early
summer because of water content (Sowls 1966). Even though my data do
not deny or support javelina favoring mesquite areas, the evidence in
the literature does. I believe javelina did not use the mesquite-free
range and totally clearing mesquite from a range would be harmful to
javelina. With clearings in mesquite, javelina would have suitable
habitat remaining to make use of this type of range.
Coyote
Visual census data of coyotes showed no significant differences
and I lacked enough scats to test. Kreftling (1969) states coyotes are
opportunistic and will eat anything. Coyotes feed on rodents,

33lagomorphs, birds, reptiles, carrion, invertebrates and vegetable mat
ter (Grater 19^3; Korschgen 1957; Hawthorne 1972)• They also feed on
javelina (Jennings and Harris 1953; Knipe 1957) and deer (Murie 1940;
Hawthorne 1972). Feeding habits from Nebraska show a rabbit-mouse-
pocket gopher complex to be very important to coyotes (Fichter, Shild-
man and Gather 1955)• At times, mice and pocket gophers are very im
portant (Murie 1940; Hawthorne 1972) and at other times lagomorphs are
of greatest importance (Grater 1943; Korschgen 1957; Clark 1972).
This may be due to relative abundance of prey items in an area. Wagner
and Stoddart (1972) believe though that coyotes caused a drop in black
tailed jackrabbit population in Utah from 1963 to 1967 and consequently
showed a major drop in their own population. Coyotes also eat mes-
quite beans in season (Grater 1943)• Home range size of coyotes in
southeastern Arizona has been reported from $4.4 square kilometers
for adults, 6.7 square kilometers for pups and yearlings and 32.4
square kilometers for the overall coyote home range size (Danner 1976).
I found no significant difference in sightings of coyote between the
mesquite and the clearings of pasture 22. Because of their wide range
in food selection, mobility and low habitat preference, coyotes would
be expected to maintain similar population sizes in any of the treat
ment areas.
Black-tailed Jackrabbit
I found no significant difference between the undisturbed mes
quite range and the mesquite with clearings. Both were significantly
higher in black-tailed jackrabbits seen than the mesquite-free range.

34
Lagomorph pellets were also significantly higher in both ranges than
the mesquite-free range. This is consistent with data from Taylor,
Vorhies and Lister (1935)• They found more jackrabbits on grazed
lands than ungrazed, indicating a preference for less dense grass
growth. Lechleitner (1958) found black-tailed jackrabbits preferred
moderately open areas. Similarly, Vorhies and Taylor (1933) found
black-tailed jackrabbits more abundant on areas of less grass cover.
The difference between black-tailed jackrabbits in clearings and in
the mesquite was not significant, and pellet count data indicated the
same results. Vorhies and Taylor (1933) also found that overall,
black-tailed jackrabbits consumed 24# grasses, $6# mesquite and 3«3#
cactus. Griffing and Davis (1976) reported that food of black-tailed
jackrabbits tended to be green succulent matter such as leaves and
seeds of mesquite. The denser grass growth and lack of abundant suc
culent growth for food keep black-tailed jackrabbit numbers low on the
mesquite-free range.
#Antelope Jackrabbit
As with the black-tailed jackrabbit, I saw significantly more
antelope jackrabbits in the undisturbed mesquite and the mesquite with
clearings pactures than the mesquite-free pasture. Vorhies and Taylor
(1933) found antelope jackrabbits more abundant in moderate grass
growth than heavy grass growth. Although all three pastures had been
grazed periodically, the mesquite-free range always had a thicker
growth of grass. Arnold (1942) notes mesquite is favored as a browse
species. Vorhies and Taylor (1933) found that antelope jackrabbits

35seemed to prefer areas where grass, mesquite and catclaws abound# They
also found that on the Santa Rita Experimental Range antelope jack-
rabbits were found where grasses were less abundant and burroweed was
high. This is the case on the mesquite with clearings and undisturbed
mesquite ranges. Pellet count data also indicated lagomorphs spent
more time in the mesquite of pasture 22 than the clearings.
Desert Cottontail
I did not see enough cottontails to test for differences.
Lagomorph pellet counts were significantly higher on the mesquite with
clearings and undisturbed mesquite ranges than the mesquite-free range,
but most of the pellets were from jackrabbits. Sowls (1957) indicates
the desert cottontail in Arizona is found in many habitats of which
the desert grassland characterized by mesquite, yucca (Yucca elata) and
many grasses is one. McCormick (1975)» working on the Santa Rita ex
perimental Range, found cottontails to be more numerous on the un
disturbed mesquite range than the mesquite-free range. Kundaeli and
Reynolds (1972) found more cottontails in areas where shrubs were of
a density of about 85 per acre. They also noted that fallen trees
which were left on the ground were good cover. Desert cottontails eat
approximately 3 % grasses, forbs and ZOfo shrubs (Turkowski 1975) •
Opuntia spp. and lovegrass are also important to desert cottontails
during drought (Turkowski 1975)• The clearings in mesquite ranges
with fallen mesquite which can be used as cover may provide excellent
habitat for desert cottontails.

36
Mourning Doves
Even though I saw four times as many mourning doves in the
mesquite with clearings than the mesquite-free range, the differences
were not significant. I saw the least amount of mourning doves in the
undisturbed mesquite range. Call counts showed a significantly higher
amount of mourning doves calling from the undisturbed mesquite and the
mesquite with clearings ranges than the mesquite-free range. This
indicates a much higher breeding population in the ranges that had
higher call counts. Perhaps these doves are not as easily censused
visually as are doves feeding on the ground.
Mourning doves eat mainly seeds of grasses, forbs and weeds
(Ward 1964). When available, cereal crops such as corn and wheat com
prise much of their diet (Korschgen 1955)• In the semidesert part of
New Mexico, the forb caltrop (Kallstroemia spp.) was a very important
part of the diet (C. A. Davis 1974). In an uncultivated area of New
Mexico, croton (Croton spp.), spurges (Euphorbia spp.) and panic
grasses (Panicum spp.) were important (Griffing and Davis 1974). For
supplying food, dense mesquite cover would not be beneficial. Fields
of grasses and forbs would be best. Nesting though often takes place
in trees (Cowan 1952; Schroeder 1970). Soutiere and Bolen (1973) found
that a loss of large mesquite trees as nesting sites was accompanied
by the occurrence of ground nesting. They also found predation to be
no greater on the ground than in the trees. So large trees may not be
absolutely necessary for nesting. However, most of the calling in my
study area came from the two ranges with mesquite. When trees are
available, they seem to be favored at least as perches for calling

37males. Doves are highly mobile and can fly great distances in one day
to take advantage of a particular resource an area can offer. I
counted the most mourning doves in the pasture with the clearings and
heard about the same calling from this area as the undisturbed mes-
quite pasture. I think creating clearings in mesquite will be a bene
ficial practice for mourning doves, leading to increased food supply
and leaving mesquite trees as preferred nesting sites. I did not see
a significant difference in the number of mourning doves seen in the
mesquite or the clearings of pasture 22. Mourning doves would make
use of both the clearings and the mesquite if both habitats are avail
able.
White-winged Doves
I did not see enough white-winged doves to test. I heard sig
nificantly more white-winged doves calling in the mesquite with clear
ings pasture than the mesquite-free pasture. White-winged doves are
closely associated with mesquite thickets usually in riparian areas
(Neff 1940b; Goodwin 1977). Neff (1940a) found white-winged doves fed
on fruits of saguaro (Carnegiea gigantea), hackberry (Celtis spp.),
Condalia lycioides, C. spathulata and also on seeds of catclaw, flower
parts of ocotillo and on oats, barley and wheat. Desert grasslands
provided little of either food or cover to white-winged doves and
mesquite invasion has helped extend their range into these areas
(Cottam and Trefethen 1968). Populations here in past years may have
been an overflow from active colonial nesting in mesquite thickets
along the nearby Santa Cruz drainage. Many of these mesquite thickets

are disappearing along the major drainages and activity may never again
be high in upper elevations as compared to mourning doves. Very little
white-winged dove activity was seen on the study area. Creating clear
ings in mesquite would still leave ample habitat for the presently
small white-winged dove population.
Scaled Quail
The numbers of scaled quail were not significantly different
between pastures. I did hear significantly more scaled quail calling
in the undisturbed mesquite area than the mesquite-free range. Ligon
(1961) stated the distribution of scaled quail is largely coextensive
with that of mesquite, Condalia spp. and cholla cactus. Goodwin and
Hungerford (1977) indicate good scaled quail habitat is characterized
by low growing grass with forbs and shrubs. Also, trees and shrubs
should be less than two meters tall. They found that scaled quail
avoided tall dense growth. Lehmann and Ward (1941) found that scaled
quail consumed large amounts of seeds of woody plants. They also think
cacti were needed for cover. Davis, Barkley and Haussamen (1975) found
that scaled quail ate large seeds and that mesquite and snakeweed
(Gutierrezia sarothrae) were important in the diet. In winter, they
found that these two plants comprised 3/4 of the diet. They suggest
there is a need to leave some mesquite in clearing operations. I found
no significant difference in the distribution of scaled quail, but this
may be a function of the size of the mesquite-free range. Scaled quail
found in the mesquite-free range may be able to get to the mesquite
bordering this range. I also did not find a significant difference in
38

39the amount of scaled quail seen in the mesquite of the clearings of
pasture 22, indicating they will probably make use of the clearings in
mesquite ranges.
Gambel* s Quail
In almost two years I saw only one gambel's quail in the
mesquite-free range. Significantly more gambel*s quail were seen in
the mesquite with clearings than the mesquite-free range. Call counts
were significantly higher in the mesquite with clearings than either
of the other two pastures. Mesquite is known to be a favorite habitat
of gambel's quail (Goodwin and Hungerford 1977)• Besides providing
cover, it provides food from buds, flowers and seeds (Gorsuch 193*0 •
Gambel's quail also eat moist berries of shrubs, fruits of mistletoe
and cacti (Hungerford 1960b). Gambel's quail may go without drinking
water but need green annual plants for reproduction to be successful
(Hungerford 1960a). I found a much greater number of gambel's quail
in the mesquite with clearings pasture. They were often associated
with the edge of the clearings and many males called from the dead
mesquite in the clearings. This study shows gambel's quail greatly
benefited from the clearings when the dead mesquite was left which
agrees with the statement by Gorsuch (1934:47), "Pure stands of well
developed mesquite or other trees are not good quail cover, for dense
shade prohibits the growth of the small herbs and grasses used as
food, and the shrubs utilized as roosts." Gambel's quail seemed to
use both mesquite and clearings and no significant difference was seen
between these two habitat types of pasture 22. Clearings can be very

40
beneficial if raesquite is close by. The large amount of edge as indi
cated by pasture 22's diversity index is significant in regard to
gambel's quail populations.
Reptiles
Zebra-tailed Lizard
The undisturbed mesquite range contained significantly more
zebra-tailed lizards than the mesquite-free range. In fact, I did not
see any zebra-tailed lizards in the mesquite-free range. Zebra-tailed
lizards utilize a wide variety of habitats in the arid Southwest in
cluding washes, flood plains of rivers, sand dunes and flat land
deserts (Vitt and Ohmart 1977a). They are particularly associated with
open space where they can run between shrubs (Pianka and Parker 1972;
Tanner and Krogh 1975)• These conditions are met by the mesquite areas
where large sandy washes are found and the vegetation around the mes
quite is sparse with many open areas. On the mesquite-free range, the
drainages are not sandy but are covered with grasses and forbs. Also,
there are no open spaces between shrubs because of, the dense grass
growth. In pasture 22 all the zebra-tailed lizards were seen in the
mesquite area, usually in sandy washes. The zebra-tailed lizard is
insectivorous (Kay, Miller and Miller 1970). It often feeds on insects
found on either overhanging annual vegetation or overhanging peren
nials such as catclaw and mesquite (Vitt and Ohmart 1977a). A mesquite
free range in which grasses dominate would eliminate zebra-tailed
lizard populations. Mesquite needs to remain, especially around
washes, if zebra-tailed lizards are to be successful.

41Desert Spiny Lizard
I found no significant difference between pastures. As with
javelina, this may partially be due to the nature of the test used.
I did not see any desert spiny lizards in the mesquite-free range.
Desert spiny lizards are primarily arboreal or sometimes saxicolous
(Parker and Pianka 1973i Vitt and Ohmart 1974). Tinkle (1976) found
them primarily arboreal on cottonwoods, mesquite and willows. Parker
and Pianka (1973) found the desert spiny lizard often in trees with
pack rat nests at the base. I also observed this on the mesquite
pastures. These lizards were quite wary and often would jump out of
a mesquite tree and run down a hole in a pack rat nest. As with the
zebra-tailed lizard, a mesquite-free environment would not provide the
habitat needed by desert spiny lizards. Mesquite remaining from spot
clearing will provide the necessary habitat for desert spiny lizards.
Lesser Earless Lizard
I found no significant differences in the amount of lesser ear
less lizards seen between pastures. I also saw no significant differ
ence in sightings between the mesquite and the clearings of pasture 22.
> Gennaro (1972) found the lesser earless lizard fed in open areas with
sparse vegetation. Degenhardt (1966), working in the Big Bend National
Park in Texas, found that the lesser earless lizard seemed to prefer
flat areas with sparse vegetation. I did not find this occurring on
my study area. The lesser earless lizard was seen approximately as
often in the dense grass areas as in areas with sparse vegetation.
Stebbins (1966) shows the lesser earless lizard to occupy a wide range

of habitats including washes, sandy stream banks, sand dunes, short
grass prairies, mesquite woodlands and farmlands. The lesser earless
lizard apparently would not decrease significantly in numbers if mes
quite were totally removed.
Tree Lizard
Tree lizards were significantly more abundant in the mesquite
with clearings range than the undisturbed mesquite range. They are
found in a variety of habitats but are mostly arboreal (Aspland 1964).
They are rarely found in areas where trees are absent and then usually
in rocky terrain (Aspland 1964; Milstead 1970). I found about the same
number in the mesquite-free range as the undisturbed mesquite range.
This may be partially due to my inability to see the tree lizards on
mesquite trees as often as they occurred. On the mesquite-free range,
I saw them often on stumps of dead mesquite. I saw significantly more
tree lizards in the clearings of the mesquite range and almost always
on dead mesquite. In winter tree lizards often aggregate in large
numbers under the bark of mesquite (Vitt 1974). Also the termite
Kalotermes minor is often eaten by the tree lizard and this termite
is found in standing or fallen dry wood (Aspland 1964). The dry mes
quite in pasture 22 is a source of both food and hibernating sites for
tree lizards and probably is the reason I found significantly more
tree lizards in this pasture.
Western Whiptail
I saw significantly more western whiptails in the undisturbed
mesquite and the mesquite with clearings ranges than the mesquite-free
42

43
range. Out of 516 western whiptails seen, only 3 were seen in the
mesquite-free range. Western whiptails inhabit more xeric habitats
(Medica 196?), often with sparse vegetation and open areas (Burkholder
and Walker 1973; Schall 1977)• They forage in shrubs and run swiftly
between these shrubs (Vitt and Ohmart 1977b). Their food consists
largely of termites (Echternacht 1967; Pianka 1970). The dense grasses
of the mesquite-free range hinders both movements and foraging be
havior of the western whiptail. I did not find a significant differ
ence between the clearings and the mesquite of pasture 22 in the
amount of western whiptails seen. Less western whiptails are expected
in the clearings as the grass cover becomes more dense. But overall,
western whiptails should be found in good numbers in the mesquite left
from spot clearing.
Sonora Spotted Whiptail
The other whiptail species found is somewhat a problem to
identify. It is probably the sonora spotted whiptail as defined by
Lowe and Wright (1964), but some non-western whiptails seen may"be
hybrids of the bisexual western whiptail and the parthenogenic sonora
spotted whiptail (Lowe, Wright, Cole and Bezy 1970). I have used the name
sonora spotted whiptail to designate the non-western whiptails I found.
Both the mesquite-free and the mesquite with clearings ranges
contained significantly more sonora spotted whiptails than the undis
turbed mesquite range. The sonora spotted whiptail is found in the
southeast corner of Arizona with the chihuahua whiptail (Cnemidophorus
exsanguis) occurring in Arizona only near the Arizona-New Mexico border

44
(Wright and Lowe 1968). Echternacht (1967), working on the Santa Rita
Experimental Range found what he called the chihuahua whip tail (more
likely the sonora spotted whiptail) at higher elevations but not in
grasslands. He thought their numbers diminished as the elevation
lowered. Wright and Lowe (1968) indicate the sonora spotted whiptail's
habitat as essentially woodlands with local populations occurring in
desert-grassland and desert scrub. The sonora spotted whiptail is
found between 1065 to 2130 meters (Lowe and Wright 1964). I found more
sonora spotted whiptails as the elevation increased. I also found no
significant difference between the mesquite and the clearings of pas
ture 22 which indicates no habitat preference. At elevations where
sonora spotted whiptails do occur, clearings in mesquite would not be
harmful to their population.
All Reptiles
Significantly more reptiles (totalling numbers for all species,
including species not listed because of low number of sightings) were
seen in the mesquite ranges than the mesquite-free range. The diver
sity of habitat types contained in a mesquite range allow for greater
numbers of reptiles.
Unidentified Lizards
The number of unidentified lizards was not significantly higher
in any one pasture. At times lizards would seek refuge in a dense
shrub or down a hole very quickly. When this happened, positive iden
tification was not possible. But numbers of unidentified lizards in
each pasture were similar and should not bias results.

4$Other Species
Another specie which seems to avoid the grasslands in Arizona
is the bobcat (Lynx rufus). Lawhead (1978), using radio telemetry,
found that an area of grassland in the Three Bar Wildlife Refuge of
Arizona was contained in three bobcat home ranges. Only once did he
locate a bobcat in this grassland. He thinks bobcats are less effec
tive hunters in large areas of grassland as compared to areas with
dense shrubbery. Although the grasslands on the Three Bar Wildlife
Refuge are on steeper slopes, the behavior of bobcats may be the same
on desert grasslands.
Nocturnal rodent populations are apparently little affected
by vegetation removal (Vaughn 1976). Vaughn (1976), working on the
Santa Rita Experimental Range, found little difference for the number
of Dipodomys merriami and Perognathus penicillatus between his cleared
and uncleared ranges. He found higher numbers of Perognathus baileyi
and P . flavus and slightly higher numbers of Peromyscus eremicus, P .
maniculatus, Reithrodontomys montanus and R. fulvescens on the cleared
range. He found higher numbers for Neotoma albigula on the uncleared
range. Overall biomass was slightly higher on the uncleared range.
Spot clearing should not be harmful to populations of bobcats
and nocturnal rodents. Totally clearing mesquite from ranges may
substantially lower bobcat numbers.

SUMMARY AND CONCLUSIONS
Creating clearings in mesquite is far less detrimental to wild
life than completely clearing whole ranges. In some cases, it benefits
wildlife. For the rancher, grass production will increase. For those
interested in wildlife, clearings in mesquite will still leave valuable
food and cover. "Edge" is greatly increased by irregular shaped clear
ings. Gambel’s quail seemed to have benefited greatly from the in
creased edge. Populations of cottontail were extremely low, however,
if they are studied when the population is at a high, cottontail may
show increased use of the clearings. Scaled quail, which seemed to
prefer a more open habitat, may increase more rapidly with clearings
and mesquite available. Numbers of most selected mammals and birds
were lower in the mesquite-free range. Total destruction of mesquite
is detrimental to the indicator mammals and birds I studied and may be
for many other species.
Zebra-tailed lizards, desert spiny lizards and western whip-
tails would suffer from the elimination of mesquite, but spot clearing
would not severely lower numbers. Tree lizard numbers would increase
with clearings as long as dead mesquite was left. The sonora spotted
whiptail does well in mesquite with clearings.
Aesthetically, spot clearing of mesquite may be much more de
sirable to the public than total clearing of mesquite. This is one
aspect of shrub control which should be studied.
46

4?Wildlife response to spot clearing should be studied after five
to ten years following treatment. The effects I found may be ephemeral
and not applicable to wildlife many years after treatment. Mesquite
require persistent periodic control to maintain a brush-free range
(Cable and Martin 1973)• Cable and Martin (1973) also found reinvasion
is not dependent on the size of range. Range managers will have to
weigh the cost versus benefit of keeping ranges open in spots for in
creased grass production.
The response of wildlife to mesquite removal in desert grass
land is favorable for spot clearing and unfavorable for totally re
moving mesquite. Spot clearing in mesquite gives a greater diversity
of habitat, has less visual impact than total clearing of mesquite, is
less detrimental to all classes of vertebrates and still accomplishes
range management objectives for more grass.
APPENDIX A
STATISTICAL ANALYSIS OF VISUAL CENSUS
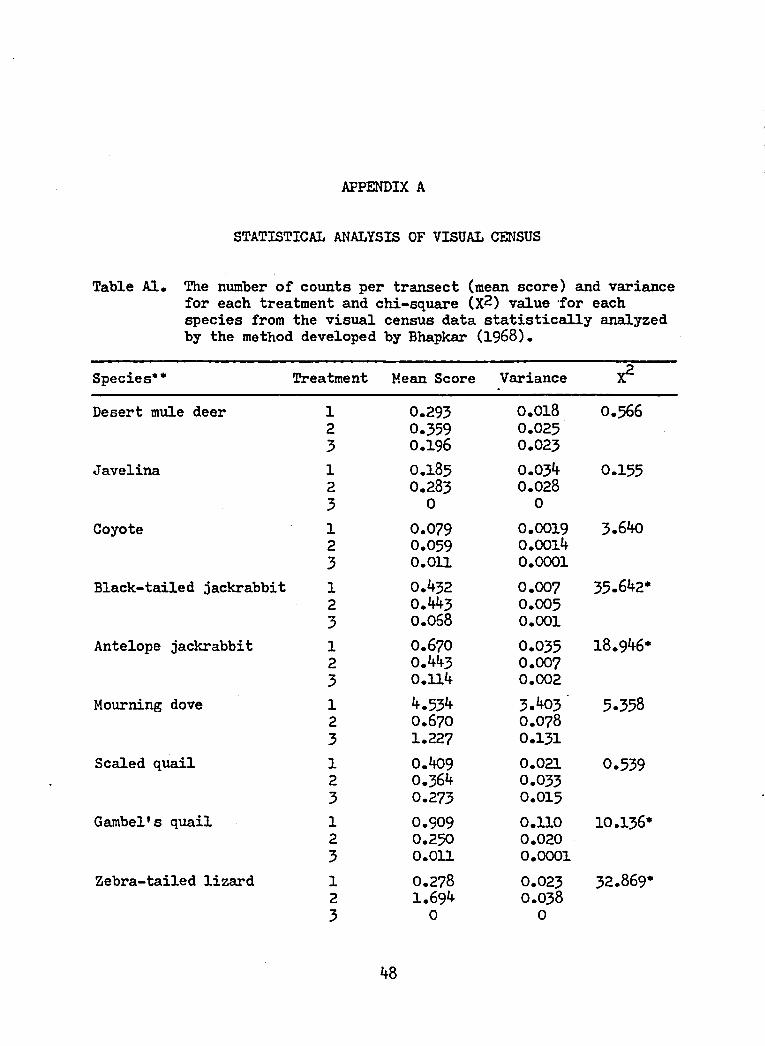
Table Al. The number of counts per transect (mean score) and variancefor each treatment and chi-square (X2) value for eachspecies from the visual census data statistically analyzedby the method developed by Bhapkar (1968).
Species** Treatment Mean Score Variance X2
Desert mule deer 1 0.293 0.018 0.5662 0.359 0.0253 0.196 0.023
Javelina 1 0.185 0.034 0.1552 O.283 0.0283 0 0
Coyote 1 0.079 0.0019 3.6402 0.059 0.00143 0.011 0.0001
Black-tailed jackrabbit 1 0.432 0.007 35.642*2 0.443 0.0053 0.068 0.001
Antelope jackrabbit 1 0.670 0.035 18.946*2 0.443 0.0073 0.114 0.002
Mourning dove 1 4.534 3.403 ' 5.3582 0.670 0.0783 1.227 0.131
Scaled quail 1 0.409 0.021 0.5392 0.364 0.0333 0.273 0.015
Gambel's quail 1 O.909 0.110 10.136*2 0.250 0.0203 0.011 0.0001
Zebra-tailed lizard 1 0.278 0.023 32.869*2 1.694 0.0383 0 0
48

Table Al— Continued
Species** Treatment Mean Score Variance X2
Desert spiny lizard 1 0.139 0.003 1.1072 0.333 0.0313 0 0
Lesser earless lizard 1 0.861 0.023 3.1012 0.500 0.0213 0.611 0.016
Tree lizard 1 1.16? 0.073 6.232*2 0.444 0.0123 0.611 0.031
Western whiptail 1 5.833 0.970 73.454*2 8.417 1.6703 0.083 0.002
Sonora spotted whiptail 1 1.472 0.081 35.984*2 0.222 0.0053 1.944 1 0.148
Unidentified lizards 1 1.139 0.057 4.6572 0.833 0.0333 1.528 0.073
All reptiles 1 11.722 2.293 10.597"2 12.056 2.4443 5.194 0.674
•Significant difference (P <.05)•••For each species the degrees of freedom are 2.1 = mesquite with clearings.2 = undisturbed mesquite.3 = mesquite-free.
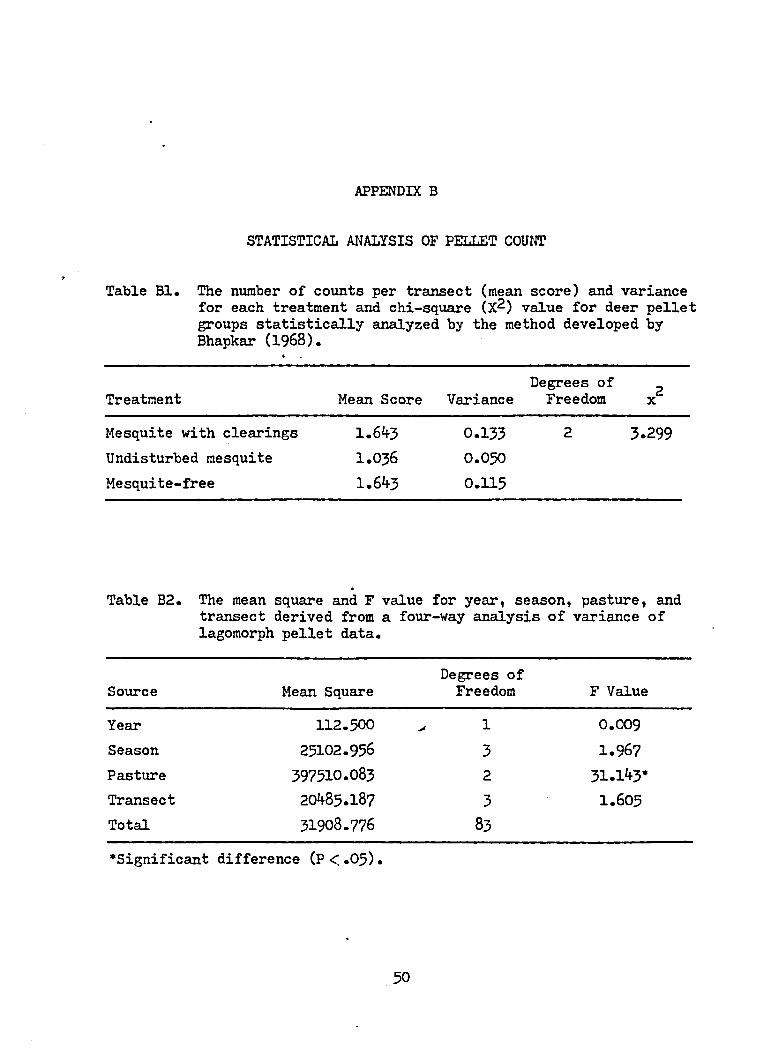
APPENDIX B
STATISTICAL ANALYSIS OF PELLET COUNT
Table Bl. The number of counts per transect (mean score) and variancefor each treatment and chi-square (X^) value for deer pelletgroups statistically analyzed by the method developed byBhapkar (1968).
Treatment Mean Score VarianceDegrees of Freedom x2
Mesquite with clearings 1.643 0.133 2 3.299Undisturbed mesquite 1.036 0.050Mesquite-free 1.643 0.115
Table B2. The mean square and F transect derived from lagomorph pellet data.
value for year, season, pasture, and a four-way analysis of variance of
Source Mean SquareDegrees of Freedom F Value
Year 112.500 > 1 0.009Season 25102.956 3 1.967Pasture 397510.083 2 31.143*Transect 20485.18? 3 1.605Total 31908.776 83♦Significant difference (P <.05)•
50

APPENDIX C
STATISTICAL ANALYSIS OF CALL COUNT DATA
Table Cl. The mean square and F values for each dove and quail species derived by the one-way analysis of variance of the call count data.
SourceDegrees of Freedom
Mean SquareMourning
DoveWhite-winged
DoveScaledQuail
Gambol's Quail
Among 2 9.834 4.032 5.275 21.452
Within 33 0.968 1.075 0.927 1.091
F Value 10.159* 3.751* 5.690* 19.663*
^Significant difference (P <.05)•
51

APPENDIX D
STATISTICAL ANALYSIS OF VISUAL CENSUS WITH OR WITHOUT THE PRESENCE OF CATTLE
Table Dl. The number of counts per transect (mean score) and variancefor the presence or absence of cattle from the pastures andchi-square (X2) value for each species from the visualcensus data statistically analyzed by the method developedby Bhapkar (1968).
Species Cattle Mean Score Variance X2
Desert mule deer present 0.13 0.00? 1.900absent 0.32 0.012
Javelina present 0.23 0.032 0.471absent 0.10 0.005
Coyote present 0.07 0.002 0.408absent 0.10 0.0002
Black-tailed jackrabbit present 0.34 0.004 2.040absent 0.24 0.004
Antelope jackrabbit present 0.31 0.010 0.300absent 0.38 0.006
Mourning dove present 0.98 0.162 1.374absent 2.02 0.662
Scaled quail present ' 0.35 0.020 0.191absent 0.28 0.006
Galbel's quail present 0.34 0.02? 0.008absent 0.32 0.019
T o r each species the degrees of freedom are 2.
52

LIST OF REFERENCES
Anthony, R. G. 1976. Influence of drought on diets and numbers of desert deer. J. Wildl. Manage. 40(l):l40-l44.
Anthony, R. G., and N. S. Smith. 1977. Ecological relationships of mule deer and white-tailed deer in southeastern Arizona.Ecol. Monogr. 47:255-277.
Arizona Interagency Range Committee. 1972. Proper use and management of grazing land. Univ. of Ariz. Publ. No. 1. 48 pp.
__________ • 1973* Grazing systems for Arizona ranges. Univ. ofAriz. Publ. No. 2. 36 pp.
__________ • 1975» Interrelationship of livestock and wildlife onArizona rangeland. Univ. of Ariz. Publ. No. 3*
Arnold, J. F. 1942. Forage consumption and preferences of experimentally fed arizona and antelope jackrabbits. Univ. of Arizona Tech. Bull. No. 98. 86 pp.
Aspland, K. K. 1964. Seasonal variation in the diet of Urosaurus ornatus in a riparian community. Herpetelogica 20(2):91-94.
Bhapkar, V. P. 1968. On the analysis of contingency tables with a quantitative response. Biometrica 24:329-338.
Brown, A. L. 1950. Shrub invasion of southern Arizona desert grassland. J. Range Manage. 3(3)i172-177•
Brown, D. E., C. L. Cochran, and T. E. Waddell. 1978. Using call-counts to predict hunter success for scaled quail. J. Wildl. Manage. 42(2):281-287.
• Brown, D. E., and C. H. Lowe. 1977. . Map of the biotic communities of the Southwest. USDA Forest Service General Tech, report RM-41.
Burkholder, G. L., and J. K. Walker. 1973. Habitat and reproduction of the desert whiptail, Cnemidophorus tigris Baird and Girard in southwestern Idaho at the northern part of its range. Herpetelogica 29(l):76-83.
53

54
Cable, D. R., and S. C. Martin. 1973• Invasion of semidesert grassland by velvet mesquite and associated vegetation changes.J. Ariz. Acad. Sci. 8(3):127-134.
__________ . 1975• Vegetation responses to grazing, rainfall, sitecondition, and mesquite control on semidesert range. USDA Forest Service research paper HM-149. 24 pp.
Caraher, D. L. 1970. Effects of longtime livestock exclusion versus grazing on the desert grassland of Arizona. Master’s thesis. University of Arizona. 4$ pp.
Clark, F. W. 1972. Influence of jackrabbit density on coyote population change. J. Wildl. Manage. 36(2):343-356.
Clemmons, S., and L. D. Wheeler. 1970. Soils report for the Santa Rita Experimental Range and Coronado National Forest, S. W. region. U.S.D.A. Forest Service. 4l pp.
Cottarn, C., and J. B. Trefethen. 1968. Whitewings. D. Van Nostrand Co., Inc., Princeton. 348 pp.
Cowan, J. B. 1952. Life history and productivity of a population of western mourning doves in California. Calif. Fish and Game 38(4):503-521.
Dahl, B. E., R. E. Sosebee, and J. P. Goen. 1974. Influence of mesquite on grass yeilds. Noxious brush and weed control research highlights 1974. Texas Tech. Univ. Vol. 5-10-12.
Danner, D. A. 1976. Coyote home range, social organization, and scent post visitation. Master's thesis, Univ. of Arizona.86 pp.
Davis, C. A. 1974. Mourning dove foods in semi-desert south-central New Mexico. J. Wildl. Manage. 38(4):941-944.
Davis, C. A., R. C. Barkley, and W. C. Haussamen. 1975* Scaled quail foods in southeastern New Mexico. J. Wildl. Manage. 39(3): 496-502.
Davis, R. B., and C. K. Winkler. 1968. Brush vs. cleared range as deer habitat in southern Texas. J. Wildl. Manage. 32(2): 321-329.
Dawson, E. T. 1950. The peccary (Pecari angulatus Cope). Texas Game Fish Comm. Wildlife profile leaflet, Dawson series. No. 68m . .2 pp.
Degenhardt, W. G. 1966. A method of counting some diurnal groundlizards of the genus Holbrookia and Cneraidophorus with results from the Big Bend National Park. Amer. Mid. Nat. 75(l):6l-100.

55Echternacht, A. C. 196?. Ecological relationships of two of the
lizard genus Cnemidophorus in the Santa Rita mountains of Arizona. Amer. Mid. Nat. 78(2):448-559•
Eddy, T. A. 1961. Foods and feeding patterns of the collared peccary in southern Arizona. J. Wildl. Manage. 25(3)i248-257•
Ellison, L. I960. Influence of grazing on plant succession of range- lands. Botanical Review 26:1-78.
Ellisor, J. E., and V. F. Harwell. 1969. Mobility and home ranges of collared peccary in southern Texas. J. Wildl. Manage.33(2):425-427.
Fichter, E., G. Shildman, and J. H. Gather. 1955. Some feeding patterns of coyotes in Nebraska. Ecol. Monogr. 25:1-37.
Gallizioli, S. 1961. Gambel quail population trend techniques.Ariz. Game and Fish Dept. Project W-78-R-5, WP1, J3« 11 pp.
Gallizioli, S., and R. Smith. 1962. Gambel quail population trend techniques. Ariz. Game and Fish Dept. Project W-78-R-6,WP1, J3. 5 pp.
Gennaro, A. L. 1972. Home range and movements of Holbrookia maculata in eastern New Mexico. Herpetelogica 28(2):165-168.
Goodwin, D. 1977. Pigeons and doves of the world. Cornell Univ. Press, New York. 446 pp.
Goodwin, J. G., Jr., and C. R. Hungerford. 1977. Habitat use bynative gambel's and scaled quail and released masked bobwhite quail in southern Arizona. USDA Forest Service Research Paper RM-197. 8 pp.
Gorsuch, D. M. 1934. Life history of gambel's quail in Arizona.Univ. of Ariz. Bio. Sci’. Bull. No. 2. 8l pp.
Grater, R. K. 1943. Coyote foods near Boulder Dam. J. Wildl. Manage. 7(4):422-423.
Griffing, J. P., and C. A. Davis. 1974. Mourning dove foods in an uncultivated area of New Mexico. J. Wildl. Manage. 38(2): 375-376.
. 1976. Black-tailed jackrabbits in southeastern NewMexico: population structure, reproduction, feeding, and useof forms. New Mexico State Univ. Agric. Exp. Stn. Research report 318. 5 pp»
Hawthorne, V. M. 1972. Coyote food habits in Sagehen Creek Basin, northeastern California. Calif. Fish and Game 58(1):4-12.

56
Humphrey, R. R. 1958. The desert grassland — a history of vegeta- tional chamges and an analysis of causes. Univ. of Arizona Press, Tucson. 73 PP«
Hungerford, C. R. 1960a. The factors affecting the breeding ofgambel’s quail in Arizona. Doctoral dissertation, Univ. of Arizona. 94 pp.
_________ 1960b. Water requirements of gambol's quail. Trans. N.Am. Wildl. Nat. Resour. Conf. 25*231-238.
Jennings, W. S., and J. T. Harris. 1953• The collared peccary inTexas. Texas Game Fish Comm. FA Report Series No. 12. 31 pp.
Kay, F. R., B. W. Miller, and C. L. Miller. 1970. Food habits andreproduction of Callisaurus draconoides in Death Valley, California. Herpetelogica 26(4):431-436.
Knipe, T. 1957* Javelina in Arizona. Ariz. Game and Fish Dept.Wildl. Bull. No. 2. 96 pp.
Korschgen, L. J. 1955* Food habits of the mourning dove in Missouri. Missouri Fish and Game Div. P-R Series No. 12. 31 PP*
__________ . 1957* Food habits of the coyote in Missouri. J. Wildl.Manage. 21(4):424-434.
Kreftling, L. W. 1969* The rise and fall of the coyote on Isle Royale. Naturalist 20(4):24-31.
Kufeld, R. C., and L. Fitzhugh. 1962. Dove survey and hunt information. Ariz. Game and Fish Dept. Project W-53-R-12, WP3, J2.7 pp.
Kundaeli, J. N., and H. J. Reynolds. 1972. Desert cottontail use ofnatural and modified pinyon-juniper woodland. J. Range Manage.25(2):116-118.
Lawhead, D. N. 1978. Home range, density, and habitat preference of the bobcat on the Three Bar Wildlife Area. Master's thesis, Univ. of Arizona. 53 PP*
Lechleitner, R. R. 1958. Certain aspects of behavior of the blacktailed jackrabbit. Amer. Mid. Nat. 60(1):145-155*
Lehmann, V. W., and H. Ward. 1941. Some plants valuable to quail in southwestern Texas. J. Wildl. Manage. 5(2):131-135*
Leopold, A. 1933* Game management. Charles Scribner's Sons. New York. 48l pp.

57Ligon, J. S. 1961. New Mexico birds and where to find them. Univ.
of New Mexico Press. Albuquerque. 360 pp.
Lowe, C. H., and J. W. Wright. 1964. Species of the Cnemidophorus exsanguis subgroup of whiptail lizards. J. Ariz. Acad. Sci. 3(2):78-bO.
Lowe, C. H., J. W. Wright, C. J. Cole, and R. L. Bezy. 1970. Natural hybridization between the Teiid lizards Cnemidophorus sonorae (Parthenogenic) and Cnemidophorus tigris (Bisexual). Systematic Zoology 19(2):114-127.
Martin, S. C. 1966. The Santa Rita Experimental Range: a center forresearch on improvement and management of semidesert range- lands. USDA Forest Service, Research paper RM-22. 24 pp.
__________ . 1975• Ecology and management of Southwestern semidesertgrass-shrub ranges: the status of our knowledge. USDA ForestService, Research paper RM-I56. 39 pp.
Martin, S. C., and H. G. Reynolds. 1973• The Santa Rita Experimental Range: your facility for research on semi-desert ecosystems.J. Ariz. Acad. Sci. 8(2):56-67.
McCormick, D. P. 1975- Effect of mesquite control on small game populations. Master's thesis, Univ. of Arizona. 66 pp.
McCulloch, C. Y. 1972. Deer foods and brush control in southern Arizona. J. Ariz. Acad. Sci. 7(3):113-119•
Medica, P. A. 1967. Food habits, habitat preference, reproduction, and diurnal activity in four sympatric species of whiptail lizards (Cnemidophorus) in southcentral New Mexico. Bull. S. Calif. Acad. Sci. 66:251-276.
Milstead, W. W. 1957. Observations on the natural history of four species of whiptail lizards, Cnemidophorus (Sauria, Teiidae) in Trans-Pecos Texas. Southwestern Nat. 2(2-3):105-121.
__________ . 1970. Late summer behavior of the lizards Sceloporusmerriami and Urosaurus ornatus in the field. Herpetelogica 26(3):343-354.
Murie, A. 1940. Ecology of the coyote in the Yellowstone. USDIFauna of the National Parks of the U.S. Bull. No. 4. 206 pp.
Neff, D. J. 1968. The pellet-group count technique for big gametrend, census, and distribution: a review. J. Wildl. Manage.32(3):597-614.

Neff, J. A. 1940a. Range, population, and game status of the western white-winged dove in Arizona. J. Wildl. Manage. 4(2):117-127•
__________ • 1940b. Notes on nesting and other habits of the westernwhite-winged dove in Arizona. J. Wildl. Manage. 4(3):279-290.
Nickerson, M. F., G. E. Brink, and C. Feddema. 1976. Principal range plants of the central and southern Rocky Mountains: names andsymbols. USDA Forest Service, General Tech. Report RM-20.121 pp.
Nie, N. H., C. H. Hull, J. G. Jenkins, K. Steinbrenner, and D. H. Bent. 1975• Statistical package for the social sciences; second edition. McGraw-Hill Book Co., Inc., New York. 675 pp.
Parker, K. W., and S. C. Martin. 1952. The mesquite problem on southern Arizona ranges. USDA Cir. 908. 70 pp.
Parker, W. S., and E. R. Pianka. 1973• Notes on the ecology of the iguanid lizard, Sceloporus magister. Herpetelogica 29(2): 143-152.
Patton, D. R. 1975. A diversity index for quantifying habitat "edge." Wildl. Soc. Bull. 3(4):171-173.
Pianka, E. R. 1970. Comparative autecology of the lizard Cnemido- phorus tigris in different parts of its geographic range. Ecology 51:703-720.
Pianka, E. R., and W. S. Parker. 1972. Ecology of the iguanid lizard Callisaurus draconoides. Copeia 1972(3):493-508.
Reynolds, H. G., and S. C. Martin. 1968. Managing grass-shrub cattle ranges in the Southwest. USDA Forest Service. Agric. handbook No. 162. 44 pp.
Rodgers, K. J. 1977. Seasonal movement of mule deer on the SantaRita Experimental Range. Master's thesis. Univ. of Arizona.63 pp.
Sauer, C. 0. 1950. Grassland climax, fire, and man. J. Range Manage.3(1):16-21.
Schall, J. J. 1977. Thermal ecology of five sympatric species ofCnemidophorus (Sauria: Teiidae). Herpetelogica 33(3):261-272.
Schroeder, M. H. 1970. Mourning dove production in a Kansas osage orange planting. J. Wildl. Manage. 34(2):344-348.
Schweinsburg, R. E. 1971. Home range, movements, and herd integrity of the collared peccary. J. Wildl. Manage. 35(3):455-460.
58

59Short, H. L. 1977• Food habits of mule deer in a semidesert grass-
shrub habitat. J. Range Manage. 30(3):206-209.
Smith, R. H., D. J. Neff, and C. Y. McCulloch. 1969* A model for the installation and use of a deer pellet group survey. Ariz.Game and Fish Dept. Special Report No. 1. 30 pp.
Soutiere, B. C., and E. G. Bolen. 1973• Role of fire in mourning dove nesting ecology. Proc. Ann. Tall Timbers Fire Ecology Conf. 12:277-288.
Sowls, L. K. 1957• Reproduction in the audubon cottontail in Arizona. J. Mammal. 38(2):234-243.
__________ . 1966. Reproduction in the collared peccary (Tayassutajacu). In Comparative biology of reproduction in mammals, ed. I. W. Rowlands, pp. 155-172. Symp. No. 15, Zool. Soc. London. New York: Academic Press.
__________ • 1974. Social behavior of the collared peccary (Dicotylestajacu L.). IUCN Inter. Symp. on the Behavior of Ungulates and its Relation to Management. Merges, Switzerland, pp. 144- 165.
Stebbins, R. C. 1966. A field guide to western reptiles and amphibians. Houghton Mifflin Co., Boston. 279 PP»
Steel, R. G. D., and J. H. Torrie. i960. Principles and procedures of statistics. McGraw-Hill Book Co., Inc., New York. 48l pp.
Tanner, W. W., and J. E. Krogh. 1975* Ecology of the zebra-tailed lizard Callisaurus draconoides at the Nevada test site. Herpetelogica 31(5):502-3l6.
Taylor, W. P., C. T. Vorhies, and P. B. Lister. 1935• The relation of jackrabbits to grazing in southern Arizona. J. Forestry 33(5):490-498.
Tinkle, D. W. 1976. Comparative data on the population ecology of the desert spiny lizard, Sceloporus magister. Herpetelogica 32(1):l-6.
Turkowski, F. J. 1975. Dietary adaptability of the desert cottontail. J. Wildl. Manage. 39(4):748-756.
U.S. Forest Service, n.d. Unpublished data, Rocky Mountain Forest and Range Experimental Station, Tempe.
Vaughn, T. C. 1976. Effects of woody vegetation removal on rodent population at Santa Rita Experimental Range, Arizona.Doctoral dissertation, Univ. of Arizona. 95 pp.

6oVitt, L. J. 1974. Winter aggregations, size classes, and relative
tail breaks in the tree lizard Urosaurus ornatus (Sauria: Iguanidae). Herpetelogica 30(2):182-1
Vitt, L« J., and R. D. Ohmart. 1974. Reproduction and ecology of a Colorado river population of Sceloporus raagister (Sauria: Iguanidae). Herpetelogica 30(4):410-417.
__________ . 1977a. Ecology and reproduction of lower Colorado riverlizards: I. Callisaurus draconoides (Iguanidae). Herpetelogica 33(2):214-222.
. 1977b. Ecology and reproduction of lower Colorado riverlizards: II. Cnemidophorus tigris (Iguanidae). Herpetelogica33(2):223-234.
Vorhies, C. T., and W. P. Taylor. 1933• The life histories of jack- rabbits, Lepus alleni and Lepus californicus spp. in relation to grazing in Arizona. Univ. of Ariz. Tech. Bull. No. 49.116 pp.
Wagner, F. H., and L. C. Stoddart. 1972. Influence of coyote predation on black-tailed jackrabbit populations in Utah. J. V/ildl. Manage. 36(2):329-342.
Ward, A. L. 1964. Foods of the mourning dove in eastern Colorado.J. Wildl. Manage. 28(1):152-157.
Whitson, R. E., S. L. Beasom, and C. J. Scifres. 1977. Economicevaluation of cattle and white-tailed deer responses to aerial spraying of mixed brush. J. Range Manage. 30(3)!214-217.
Wright, J. W., and C. H. Lowe. 1968. Weeds, polyploids, parthenogenesis, and the geographical and ecological distribution of all-female species of Cnemidophorus. Copeia 1968(1):128-138.

!: ^289a