researcharticle ouabain modulates zymosan-induced ... · researcharticle ouabain modulates...
TRANSCRIPT

Research ArticleOuabain Modulates Zymosan-Induced Peritonitis in Mice
Jacqueline Alves Leite1 Anne Kaliery De Abreu Alves2 Joseacute Guilherme Marques Galvatildeo2
Mariana Pires Teixeira3 Luiz Henrique Agra Cavalcante-Silva2 Cristoforo Scavone1
Alexandre Morrot4 Vivian Mary Rumjanek3 and Sandra Rodrigues-Mascarenhas25
1 Instituto de Ciencias Biomedicas Departamento de Farmacologia Laboratorio de Neurofarmacologia MolecularUniversidade de Sao Paulo 05508-900 Sao Paulo SP Brazil2Centro de Biotecnologia Laboratorio de Imunofarmacologia Programa de Pos-Graduacao em Fisiologia MulticentricoUniversidade Federal da Paraıba 58059-900 Joao Pessoa PB Brazil3Instituto de Bioquımica Medica Leopoldo de Meis Laboratorio de Imunologia Tumoral Universidade Federal do Rio de Janeiro21941-902 Rio de Janeiro RJ Brazil4Laboratorio de Biologia do Sistema Imune Instituto de Microbiologia Universidade Federal do Rio de Janeiro21941-902 Rio de Janeiro RJ Brazil5Centro de Ciencias da Saude Programa de Pos-Graduacao em Produtos Naturais e Sinteticos BioativosUniversidade Federal da Paraıba 58059-900 Joao Pessoa PB Brazil
Correspondence should be addressed to Sandra Rodrigues-Mascarenhas sandramascarenhasltfufpbbr
Received 20 March 2015 Accepted 22 April 2015
Academic Editor Elzbieta Kolaczkowska
Copyright copy 2015 Jacqueline Alves Leite et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Ouabain a potent inhibitor of the Na+ K+-ATPase was identified as an endogenous substance Recently ouabain was shown toaffect various immunological processes We have previously demonstrated the ability of ouabain to modulate inflammation butlittle is known about the mechanisms involved Thus the aim of the present work is to evaluate the immune modulatory role ofouabain on zymosan-induced peritonitis inmice Our results show that ouabain decreased plasma exudation (33) After inductionof inflammation OUA treatment led to a 46 reduction in the total number of cells as a reflex of a decrease of polymorphonuclearleukocytes which does not appear to be due to cell death Furthermore OUA decreased TNF-120572 (57) and IL-1120573 (58) levelswithout interfering with IL-6 and IL-10 Also in vitro experiments show that ouabain did not affect endocytic capacity Moreoverelectrophoretic mobility shift assay (EMSA) shows that zymosan treatment increased (85) NF-120581B binding activity and thatouabain reduced (30) NF-120581B binding activity induced by zymosan Therefore our data suggest that ouabain modulated acuteinflammatory response reducing the number of cells and cytokines levels in the peritoneal cavity as well as NF120581B activationsuggesting a new mode of action of this substance
1 Introduction
Ouabain a steroid capable of inhibiting Na+K+-ATPaseis an adrenal and hypothalamus-derived hormone presentin mammalian plasma and tissue which appears to beproduced in response to volume expansion angiotensin IIandor adrenocorticotropic hormone stimulation [1ndash4] Ithas also been suggested that endogenous ouabain levels aremodulated by stress conditions [5 6] Moreover elevatedlevels of ouabain were found in hypertensive patients andnormotensive individuals after physical exercise [5 7]
Ouabain is able tomodulate several immunological func-tions inhibiting in vitro proliferation of lymphocytes inducedby different mitogens [8ndash11] increasing the expression of themolecule CD69 on the thymocyte surface [12] and decreas-ing the levels of phosphorylated MAPK p38 and nuclearfactor of activated T cells (NFATc1) [13] The combination ofouabain plus corticoids produced a synergic effect inducingthymic atrophy in vivo [14] Additionally ouabain was shownto modulate function and cytokine production of monocytesin vitro [15] and dendritic cells [16] Ouabain negativelymodulated acute peritoneal inflammatory response induced
Hindawi Publishing CorporationMediators of InflammationVolume 2015 Article ID 265798 12 pageshttpdxdoiorg1011552015265798
2 Mediators of Inflammation
by Leishmania amazonensis infection in Swiss mice [17] Itwas also reported that ouabain presented anti-inflammatoryand antinociceptive effects [18] but themechanisms involvedwere still unclear Nonetheless some ouabain effects are con-troversial It has also been described using different modelsthat ouabain is capable of increasing the activation of NF-120581B VCAM-1 and iNOS expression NO and inflammatorycytokines levels [19ndash22]
During inflammation a complex process of intracellularsignal transduction and transcription events driven by mul-tiple proinflammatory mediators and cytokines is activatedAcute inflammation is characterized by exudation of protein-rich fluid edema vasodilatation and cell migration into thesite of injury [23] Intraperitoneal injection of zymosan apolysaccharide component of cell wall from Saccharomycescerevisiae represents a self-resolving model of acute inflam-mation and has been widely used for the quantification ofparticular cell types and inflammation-related soluble factors[24ndash26] When administered in mouse peritoneal cavityzymosan induces an increased vascular extravasation whichis one of the primary signs of inflammation This is a keystep in inflammatory exudate formation and is followed bya time-dependent recruitment of migratory cells speciallyneutrophils andmonocytesmacrophages [26ndash32]Thereforeto better understand the anti-inflammatory effect of ouabainwe aimed to assess its action in vascular leakage leukocytemigration as well as apoptosis of leukocytes cytokine pro-duction and endocytosis
2 Material and Methods
21 Animals Female Swiss albino mice (2 months old) wereobtained from Thomas George animal house of Centro deBiotecnologia (CBiotec UFPB Joao Pessoa Brazil) Animalswere kept under standard laboratory conditions on a constant12 h lightdark cycle with temperature (21 plusmn 1∘C) Food andwater were given ad libitum All procedures adopted in thisstudy were approved by Institutional Ethics Committee ofCentro de Biotecnologia (Protocol 050411)
22 In Vivo Treatment with Ouabain In all in vivo exper-iments 056mgkg ouabain (Sigma Aldrich) [14 17 18] orphosphate buffered saline (PBS) was given intraperitoneally(ip) for three consecutive days
23 Assessment of Vascular Permeability Mice were ran-domly divided into four groups (119899 = 6) and each group ofmice was administrated intraperitoneally (ip) once a dayfor 3 consecutive days with either vehicle (PBS) or ouabain(056mgkg) One hour after the last injection on day 3 micewere injected iv into the caudal vein with Evans blue dyesuspended in saline at 10mgmL (03mLmouse) The dyeinjection was immediately followed by an ip injection ofzymosan The animals were euthanized after 30min and theperitoneal cavity was lavaged with 15mL of saline Lavagefluid was then centrifuged for 10min at 350 gThe absorbanceof cell-free lavage fluid was measured at 650 nm [32]
24 Induction of Peritonitis Peritoneal inflammation wasinduced as previously described [26 32]Micewere randomlydivided into four groups (119899 = 8) and each group receivedip the same volume (200120583L) of vehicle (PBS) or ouabain(056mgkg) once a day for 3 consecutive days Zymosan Awas freshly prepared (2mgmL) in sterile 09wv saline and05mLwas injected ip one hour after the last injection onday3 At either four or twenty-four hours after stimulus animalswere euthanized by cervical dislocationThe peritoneal cavitywas then lavagedwith 1mLof cold PBS Exudateswere pooledand the number of leukocytes present was determined byoptical microscopy in Hemocytometer using Turkrsquos solution(001 crystal violet in 3 acetic acid) Finally the exudateswere centrifuged and an aliquot of the supernatant wascollected and stored at minus20∘C for cytokine analysis
25 Leukocytes Subsets Analysis of the cell populations in theperitoneal cavity was performed by flow cytometry based onsize and granularity parameters as well as on Gr-1 (Ly-6GandLy-6C) expression andMac-3 expression to identify neu-trophils andmacrophages respectivelyMonocytes expressedboth Gr-1 and Mac-3 (Gr1+Mac3+) Leukocytes collectedfrom peritoneal lavage were stained with the antibodieskits (eBioScience) according to the manufacturerrsquos protocolBriefly 1 times 106 cellsmL were incubated with a saturatingamount of anti-Gr-1 PE-conjugated and anti-Mac-3 FITC-conjugated (eBioScience) for 30min at 4∘C Cells were thenwashed with cold PBS and resuspended in the same solutionAnalyses of Gr-1 andMac-3 expression were performed usingFACSCalibur flow cytometer In these experiments data wereacquired in a mode of 10000 events FITC was measuredin FL-1 and PE was measured in FL-2 channels Data wereanalyzed by WinMID software
26 Measurement of Cytokines Levels by an Enzyme-LinkedImmunosorbent Assay (ELISA) TNF-120572 IL-1120573 IL-6 and IL-10 levels in the peritoneal fluid were evaluated by mousespecific sandwich ELISA according to the manufacturerrsquosinstructions (eBioScience) Optical density was read using amicroplate spectrophotometer (microplate reader VersaMaxtunable BN 2529 Molecular Devices)
27 Apoptosis Assessment by Flow Cytometry Apoptoticleukocytes were identified quantitatively by Annexin V-PEApoptosis Detection Kit (BD Bioscience) that enables cellstainingwithAnnexinV and propidium iodide (PI) AnnexinV binds to phosphatidylserine exposed on the outer leaflet ofthe plasma membrane of apoptotic dying cells while PI is avital dye that enters any necrotic cell Leukocytes collectedfrom peritoneum were stained with this kit according to themanufacturerrsquos protocol Briefly cells were washed twice withcold PBS and resuspended in binding buffer (10mM Hepes140mM NaCl and 25mM CaCl
2) Then 25120583L of Annexin
V-PE and 10 120583L of PI (50 gmL) were added to cell sus-pensions (5 times 105 cells100 120583L binding buffer) and incubatedfor 15min at room temperature in the dark Fluorescencemeasurement was performed using a flow cytometer (BD
Mediators of Inflammation 3
PBS Ouabain
Zymosan00
05
10
15
Opt
ical
den
sity
(OD
)
lowastlowast
Ouabainzymosan
Figure 1 Ouabain decreases plasma extravasation induced byzymosan Swiss mice (119899 = 6) were pretreated with 056mgkg ofouabain (ip) or PBS for 3 consecutive days One hour after thelast dose of treatment animals were stimulated with zymosan (ip)The results were expressed as absorbance for the Evans blue dyepresent in the peritoneal exudate Results were expressed as mean plusmnSEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquosposttest where all groups were compared 119875 lt 0001 versus PBSgroup and lowastlowast119875 lt 001 versus zymosan group
FACSCalibur) Annexin V-PE was measured in FL-2 and PIin FL-1 channels Results were analyzed byWinMID software
28 Analysis of Macrophage Viability Peritoneal exudate waselicited in mice with an ip injection of 3mgmL of thiogly-colate (Sigma Aldrich) Four days after the ip thioglycolateinjection animals were euthanized by cervical displacementand the peritoneal cavity was washed with 8mL of completeRPMI-1640medium (Gibco) (streptomycin 10mgmL peni-cillin 6mgmL and kanamycin 2mgmL) supplementedwith 10 fetal bovine serum (FBS) (Gibco) Cell suspensionobtained from peritoneal lavage was centrifuged at 350 g for15 minutes (4∘C) Supernatant was discarded and the pelletwas resuspended in 1mL of complete RPMI medium Viablecells were counted with a Neubauer chamber (Hemocytome-ter L OptikATC-111020) using Trypan blue solution (Merck)Macrophages were then enriched by adherence to plasticFor that viable peritoneal cells were seeded in 96-well platesat a concentration of 4 times 105 cellswell in a final volume of200120583L and incubated for 2 h with FBS-free culture mediumin an atmosphere of 5 CO
2at 37∘C Then nonadherent
cells were removed by wash with PBS Remaining cells werefurther incubated for 24 h with complete RPMI mediumin the presence and absence of different concentrations ofouabain (1 10 and 100 nM) Cell viability was estimated byMTT (3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazoliumbromide) assay For that 200120583L of RPMImedium containing20120583L of MTT solution (5mgmL of MTT in PBS) wasadded to each well After 4 h of incubation MTT-containingmedium was removed and the precipitate was solubilized inDMSO solution (200 120583L) The optical density was read at570 nm using a microplate spectrophotometer (SpectraMaxMolecular Devices Sunnyvale CA)
0
10
20
30
PBS
Oua
bain
Zym
osan
lowastlowastlowast
4h 24h
Cel
l num
bertimes
106
(mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Figure 2 Total cell number in the peritoneal cavity 4 and 24 hafter injection with zymosan Swiss mice (119899 = 8) were treated with056mgkg of ouabain (ip) or PBS for 3 consecutive days Onehour after the last dose of treatment animals were stimulated withzymosan (ip) Four and 24 hours after challenge with zymosanperitoneal exudate was collected and total leukocyte counts wereperformed Results were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where allgroups were compared 119875 lt 0001 versus PBS group and lowastlowastlowast119875 lt001 versus zymosan group
29 Quantification of Fluorescein Isothiocyanate- (FITC-)Dextran Endocytosis by Flow Cytometry Analysis Residentperitoneal cells were placed in sterile 24-well plates at theconcentration of 2 times 106 cellsmL (final volume = 05mL)and incubated for 2 h with RPMI-1640 medium containing10 FCS in an atmosphere of 5 CO
2and 37∘C Following
the incubation period cells were washed twice with PBSfor the removal of nonadherent cells Remaining cells werefurther incubated in a final volume (500 120583L) of RPMI-1640medium containing 10 FCS with 05mgmL of 40 kDaFITC-dextran (Sigma Aldrich) in the presence or absenceof different concentrations of ouabain (1 10 and 100 nM)for 1 h at 37∘C or 4∘C (control endocytic activity) or inRPMI-1640 medium containing 10 FCS in the presenceor absence of different concentrations of ouabain for 24 hat 37∘C followed by the addition of RPMI-1640 mediumcontaining 10 FCS with FITC-dextran (05mgmL) for 1 hat 37∘C or 4∘C Cells were then washed with ice-cold PBSand fluorescence measurement was performed using a BDFACSCalibur flow cytometer Fluorescence signs correspond-ing to dextran-FITC uptake were analyzed by Summit v43software (Dako USA) using a macrophageneutrophil gatewhich was determined according to cell size and complexityparameters In a manner somewhat similar to the previousarticle fromTeixeira and Rumjanek [15] results are presentedboth as percentage of cells that endocytosed dextran andas MFI (median fluorescence intensity) a measurement thatin the present work is representative of the amount offluorescence per cell
4 Mediators of Inflammation
84 11
3 2
5 3
73 18
60 14
21 5
21 5
73 1
28 11
33 27
26 14
33 27
4 hours
3 2
72 23
4 3
69 24
24 hours
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
GR-
1G
R-1
GR-
1G
R-1
MAC-3 MAC-3
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
20
40
60
lowastlowast
o
f cel
l Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(b)
Figure 3 Continued
Mediators of Inflammation 5
0
20
40
60
o
f cel
l Mac3+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(c)
0
20
40
60
o
f cel
l Mac3+
Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 3 Effect of ouabain on peritoneal cavity leukocyte subsets Cells were obtained from peritoneal lavage 4 and 24 h after the injection ofthe stimuli ip Neutrophils monocytes and macrophages were distinguished by flow cytometry using anti-Gr-1 and anti-Mac-3 antibodies(a) Representative experiment showing the patterns of Gr-1+ and Mac-3 expression as well as the leukocyte populations defined by thesemolecules Values of the percentage of neutrophils (Gr-1+) macrophages (Mac-3+) and monocytes (Gr-1+ and Mac-3+) were also added tothe dot plots ((b) (c) and (d)) Data were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttestwhere all groups were compared The results were obtained from 6 animals per group
119875 lt 0001 and 119875 lt 005 versus PBS group
lowastlowast
119875 lt 001 versus zymosan group
210 TransmigrationAssay Twodifferent groups of cells weretested for their migratory capacity in vitro The control groupconsisted of resident peritoneal cells obtained from animalsinjected once a day in the three previous days with PBSand the experimental pool obtained from animals injectedwith ouabain (056mgkg) once a day for 3 consecutive daysChemotaxis was measured by migration through a polycar-bonate filter of 5 120583m pore size in 24-well transwell cham-bers (Corning Costar Cambridge MA) DMEM containing05 FCS (500120583L) plus lipopolysaccharide (100 ngmL) ormedium alone as control for spontaneous migration wasadded to the lower chambers 106 cells (100 120583L) were added tothe upper chambers and were incubated at 37∘C in a 5 CO
2
humidified atmosphere After 4 h migration was defined bycounting the cells thatmigrated to the lower chambers by flowcytometry (BD FACSCalibur flow cytometer)
211 Nuclear Extracts Nuclear extracts of peritoneal cellswere prepared based on an adapted version of Rong andBaudry [33] Briefly cells were lysed in cold phosphatebuffered saline (PBS) [supplemented with 2120583gmL leupeptin2 120583gmL antipain and 05mM PMSF (all obtained fromSigma Aldrich)] and centrifuged at 4∘C for 2min at 12000 gPellets were resuspended in lysis buffer [10mM HEPESpH 79 15mM MgCl
2 10mM KCl 01mM EDTA 05mM
PMSF 2 120583gmL leupeptin 2120583gmL antipain 3mM sodiumorthovanadate 30mM sodium fluoride and 20mM sodiumpyrophosphate (all obtained from Sigma Aldrich)] and incu-bated on ice for 15min After that NP-40 (Sigma Aldrich)(final concentration of 05) was added and samples werevigorously mixed and centrifuged for 30 s at 12000 g Pelletswere then resuspended in extraction buffer [20mM HEPESpH 79 25 glycerol 15mMMgCl
2 300mMNaCl 025mM
EDTA 05mM PMSF 2 120583gmL leupeptin and 2 120583gmLantipain (all obtained from Sigma Aldrich)] incubated for20min on ice and centrifuged for 20min at 13000 g at 4∘CThe resulting supernatants containing nuclear proteins werestored at minus80∘C Protein concentration was determined usingthe Bio-Rad (Richmond CA USA) colorimetric assay [34]
212 Electrophoretic Mobility Shift Assay Electrophoreticmobility shift assay (EMSA) for NF-120581B was performedusing gel shift assay kit from Promega (Madison WI USA)[33] NF120581B double-stranded consensus oligonucleotide(51015840-AGTTGAGGGGACTTTCCCAGGC-31015840) was end-labeled with 120574-32P-ATP using T4 polynucleotide kinaseUnincorporated nucleotides were removed by passing thereaction mixture through a Sephadex G-25 spin column(Amersham-Pharmacia Uppsala Sweden) Purified 32P-labeled probe (25000 cpm) was incubated in 20 120583L with5 120583g of nuclear extracts in a binding reaction mixturecontaining 50mM NaCl 02mM EDTA 05mM DTT 4glycerol 10mM TrisndashHCl (pH 75) and 005 120583g of poly(dIndashdC) for 30min at room temperature DNA-proteincomplexes were separated by electrophoresis using a 6nondenaturing acrylamide bisacrylamide (375 1) gel in 05X Tris-borateEDTA (TBE) for 2 h at 150V Gels were vacuumdried and analyzed by autoradiography Autoradiographswere visualized with a photo documentation system DP-001-FDC (Vilber Lourmat Marne la Vallee France) andquantified in NIH ImageJ software (Bethesda MD USA)Several exposure times were analyzed to ensure the linearityof the band intensities
213 Statistical Analysis All data were expressed as mean plusmnSEM and analyzed by Graphpad Prism 50 software using
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Mediators of Inflammation
by Leishmania amazonensis infection in Swiss mice [17] Itwas also reported that ouabain presented anti-inflammatoryand antinociceptive effects [18] but themechanisms involvedwere still unclear Nonetheless some ouabain effects are con-troversial It has also been described using different modelsthat ouabain is capable of increasing the activation of NF-120581B VCAM-1 and iNOS expression NO and inflammatorycytokines levels [19ndash22]
During inflammation a complex process of intracellularsignal transduction and transcription events driven by mul-tiple proinflammatory mediators and cytokines is activatedAcute inflammation is characterized by exudation of protein-rich fluid edema vasodilatation and cell migration into thesite of injury [23] Intraperitoneal injection of zymosan apolysaccharide component of cell wall from Saccharomycescerevisiae represents a self-resolving model of acute inflam-mation and has been widely used for the quantification ofparticular cell types and inflammation-related soluble factors[24ndash26] When administered in mouse peritoneal cavityzymosan induces an increased vascular extravasation whichis one of the primary signs of inflammation This is a keystep in inflammatory exudate formation and is followed bya time-dependent recruitment of migratory cells speciallyneutrophils andmonocytesmacrophages [26ndash32]Thereforeto better understand the anti-inflammatory effect of ouabainwe aimed to assess its action in vascular leakage leukocytemigration as well as apoptosis of leukocytes cytokine pro-duction and endocytosis
2 Material and Methods
21 Animals Female Swiss albino mice (2 months old) wereobtained from Thomas George animal house of Centro deBiotecnologia (CBiotec UFPB Joao Pessoa Brazil) Animalswere kept under standard laboratory conditions on a constant12 h lightdark cycle with temperature (21 plusmn 1∘C) Food andwater were given ad libitum All procedures adopted in thisstudy were approved by Institutional Ethics Committee ofCentro de Biotecnologia (Protocol 050411)
22 In Vivo Treatment with Ouabain In all in vivo exper-iments 056mgkg ouabain (Sigma Aldrich) [14 17 18] orphosphate buffered saline (PBS) was given intraperitoneally(ip) for three consecutive days
23 Assessment of Vascular Permeability Mice were ran-domly divided into four groups (119899 = 6) and each group ofmice was administrated intraperitoneally (ip) once a dayfor 3 consecutive days with either vehicle (PBS) or ouabain(056mgkg) One hour after the last injection on day 3 micewere injected iv into the caudal vein with Evans blue dyesuspended in saline at 10mgmL (03mLmouse) The dyeinjection was immediately followed by an ip injection ofzymosan The animals were euthanized after 30min and theperitoneal cavity was lavaged with 15mL of saline Lavagefluid was then centrifuged for 10min at 350 gThe absorbanceof cell-free lavage fluid was measured at 650 nm [32]
24 Induction of Peritonitis Peritoneal inflammation wasinduced as previously described [26 32]Micewere randomlydivided into four groups (119899 = 8) and each group receivedip the same volume (200120583L) of vehicle (PBS) or ouabain(056mgkg) once a day for 3 consecutive days Zymosan Awas freshly prepared (2mgmL) in sterile 09wv saline and05mLwas injected ip one hour after the last injection onday3 At either four or twenty-four hours after stimulus animalswere euthanized by cervical dislocationThe peritoneal cavitywas then lavagedwith 1mLof cold PBS Exudateswere pooledand the number of leukocytes present was determined byoptical microscopy in Hemocytometer using Turkrsquos solution(001 crystal violet in 3 acetic acid) Finally the exudateswere centrifuged and an aliquot of the supernatant wascollected and stored at minus20∘C for cytokine analysis
25 Leukocytes Subsets Analysis of the cell populations in theperitoneal cavity was performed by flow cytometry based onsize and granularity parameters as well as on Gr-1 (Ly-6GandLy-6C) expression andMac-3 expression to identify neu-trophils andmacrophages respectivelyMonocytes expressedboth Gr-1 and Mac-3 (Gr1+Mac3+) Leukocytes collectedfrom peritoneal lavage were stained with the antibodieskits (eBioScience) according to the manufacturerrsquos protocolBriefly 1 times 106 cellsmL were incubated with a saturatingamount of anti-Gr-1 PE-conjugated and anti-Mac-3 FITC-conjugated (eBioScience) for 30min at 4∘C Cells were thenwashed with cold PBS and resuspended in the same solutionAnalyses of Gr-1 andMac-3 expression were performed usingFACSCalibur flow cytometer In these experiments data wereacquired in a mode of 10000 events FITC was measuredin FL-1 and PE was measured in FL-2 channels Data wereanalyzed by WinMID software
26 Measurement of Cytokines Levels by an Enzyme-LinkedImmunosorbent Assay (ELISA) TNF-120572 IL-1120573 IL-6 and IL-10 levels in the peritoneal fluid were evaluated by mousespecific sandwich ELISA according to the manufacturerrsquosinstructions (eBioScience) Optical density was read using amicroplate spectrophotometer (microplate reader VersaMaxtunable BN 2529 Molecular Devices)
27 Apoptosis Assessment by Flow Cytometry Apoptoticleukocytes were identified quantitatively by Annexin V-PEApoptosis Detection Kit (BD Bioscience) that enables cellstainingwithAnnexinV and propidium iodide (PI) AnnexinV binds to phosphatidylserine exposed on the outer leaflet ofthe plasma membrane of apoptotic dying cells while PI is avital dye that enters any necrotic cell Leukocytes collectedfrom peritoneum were stained with this kit according to themanufacturerrsquos protocol Briefly cells were washed twice withcold PBS and resuspended in binding buffer (10mM Hepes140mM NaCl and 25mM CaCl
2) Then 25120583L of Annexin
V-PE and 10 120583L of PI (50 gmL) were added to cell sus-pensions (5 times 105 cells100 120583L binding buffer) and incubatedfor 15min at room temperature in the dark Fluorescencemeasurement was performed using a flow cytometer (BD
Mediators of Inflammation 3
PBS Ouabain
Zymosan00
05
10
15
Opt
ical
den
sity
(OD
)
lowastlowast
Ouabainzymosan
Figure 1 Ouabain decreases plasma extravasation induced byzymosan Swiss mice (119899 = 6) were pretreated with 056mgkg ofouabain (ip) or PBS for 3 consecutive days One hour after thelast dose of treatment animals were stimulated with zymosan (ip)The results were expressed as absorbance for the Evans blue dyepresent in the peritoneal exudate Results were expressed as mean plusmnSEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquosposttest where all groups were compared 119875 lt 0001 versus PBSgroup and lowastlowast119875 lt 001 versus zymosan group
FACSCalibur) Annexin V-PE was measured in FL-2 and PIin FL-1 channels Results were analyzed byWinMID software
28 Analysis of Macrophage Viability Peritoneal exudate waselicited in mice with an ip injection of 3mgmL of thiogly-colate (Sigma Aldrich) Four days after the ip thioglycolateinjection animals were euthanized by cervical displacementand the peritoneal cavity was washed with 8mL of completeRPMI-1640medium (Gibco) (streptomycin 10mgmL peni-cillin 6mgmL and kanamycin 2mgmL) supplementedwith 10 fetal bovine serum (FBS) (Gibco) Cell suspensionobtained from peritoneal lavage was centrifuged at 350 g for15 minutes (4∘C) Supernatant was discarded and the pelletwas resuspended in 1mL of complete RPMI medium Viablecells were counted with a Neubauer chamber (Hemocytome-ter L OptikATC-111020) using Trypan blue solution (Merck)Macrophages were then enriched by adherence to plasticFor that viable peritoneal cells were seeded in 96-well platesat a concentration of 4 times 105 cellswell in a final volume of200120583L and incubated for 2 h with FBS-free culture mediumin an atmosphere of 5 CO
2at 37∘C Then nonadherent
cells were removed by wash with PBS Remaining cells werefurther incubated for 24 h with complete RPMI mediumin the presence and absence of different concentrations ofouabain (1 10 and 100 nM) Cell viability was estimated byMTT (3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazoliumbromide) assay For that 200120583L of RPMImedium containing20120583L of MTT solution (5mgmL of MTT in PBS) wasadded to each well After 4 h of incubation MTT-containingmedium was removed and the precipitate was solubilized inDMSO solution (200 120583L) The optical density was read at570 nm using a microplate spectrophotometer (SpectraMaxMolecular Devices Sunnyvale CA)
0
10
20
30
PBS
Oua
bain
Zym
osan
lowastlowastlowast
4h 24h
Cel
l num
bertimes
106
(mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Figure 2 Total cell number in the peritoneal cavity 4 and 24 hafter injection with zymosan Swiss mice (119899 = 8) were treated with056mgkg of ouabain (ip) or PBS for 3 consecutive days Onehour after the last dose of treatment animals were stimulated withzymosan (ip) Four and 24 hours after challenge with zymosanperitoneal exudate was collected and total leukocyte counts wereperformed Results were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where allgroups were compared 119875 lt 0001 versus PBS group and lowastlowastlowast119875 lt001 versus zymosan group
29 Quantification of Fluorescein Isothiocyanate- (FITC-)Dextran Endocytosis by Flow Cytometry Analysis Residentperitoneal cells were placed in sterile 24-well plates at theconcentration of 2 times 106 cellsmL (final volume = 05mL)and incubated for 2 h with RPMI-1640 medium containing10 FCS in an atmosphere of 5 CO
2and 37∘C Following
the incubation period cells were washed twice with PBSfor the removal of nonadherent cells Remaining cells werefurther incubated in a final volume (500 120583L) of RPMI-1640medium containing 10 FCS with 05mgmL of 40 kDaFITC-dextran (Sigma Aldrich) in the presence or absenceof different concentrations of ouabain (1 10 and 100 nM)for 1 h at 37∘C or 4∘C (control endocytic activity) or inRPMI-1640 medium containing 10 FCS in the presenceor absence of different concentrations of ouabain for 24 hat 37∘C followed by the addition of RPMI-1640 mediumcontaining 10 FCS with FITC-dextran (05mgmL) for 1 hat 37∘C or 4∘C Cells were then washed with ice-cold PBSand fluorescence measurement was performed using a BDFACSCalibur flow cytometer Fluorescence signs correspond-ing to dextran-FITC uptake were analyzed by Summit v43software (Dako USA) using a macrophageneutrophil gatewhich was determined according to cell size and complexityparameters In a manner somewhat similar to the previousarticle fromTeixeira and Rumjanek [15] results are presentedboth as percentage of cells that endocytosed dextran andas MFI (median fluorescence intensity) a measurement thatin the present work is representative of the amount offluorescence per cell
4 Mediators of Inflammation
84 11
3 2
5 3
73 18
60 14
21 5
21 5
73 1
28 11
33 27
26 14
33 27
4 hours
3 2
72 23
4 3
69 24
24 hours
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
GR-
1G
R-1
GR-
1G
R-1
MAC-3 MAC-3
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
20
40
60
lowastlowast
o
f cel
l Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(b)
Figure 3 Continued
Mediators of Inflammation 5
0
20
40
60
o
f cel
l Mac3+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(c)
0
20
40
60
o
f cel
l Mac3+
Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 3 Effect of ouabain on peritoneal cavity leukocyte subsets Cells were obtained from peritoneal lavage 4 and 24 h after the injection ofthe stimuli ip Neutrophils monocytes and macrophages were distinguished by flow cytometry using anti-Gr-1 and anti-Mac-3 antibodies(a) Representative experiment showing the patterns of Gr-1+ and Mac-3 expression as well as the leukocyte populations defined by thesemolecules Values of the percentage of neutrophils (Gr-1+) macrophages (Mac-3+) and monocytes (Gr-1+ and Mac-3+) were also added tothe dot plots ((b) (c) and (d)) Data were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttestwhere all groups were compared The results were obtained from 6 animals per group
119875 lt 0001 and 119875 lt 005 versus PBS group
lowastlowast
119875 lt 001 versus zymosan group
210 TransmigrationAssay Twodifferent groups of cells weretested for their migratory capacity in vitro The control groupconsisted of resident peritoneal cells obtained from animalsinjected once a day in the three previous days with PBSand the experimental pool obtained from animals injectedwith ouabain (056mgkg) once a day for 3 consecutive daysChemotaxis was measured by migration through a polycar-bonate filter of 5 120583m pore size in 24-well transwell cham-bers (Corning Costar Cambridge MA) DMEM containing05 FCS (500120583L) plus lipopolysaccharide (100 ngmL) ormedium alone as control for spontaneous migration wasadded to the lower chambers 106 cells (100 120583L) were added tothe upper chambers and were incubated at 37∘C in a 5 CO
2
humidified atmosphere After 4 h migration was defined bycounting the cells thatmigrated to the lower chambers by flowcytometry (BD FACSCalibur flow cytometer)
211 Nuclear Extracts Nuclear extracts of peritoneal cellswere prepared based on an adapted version of Rong andBaudry [33] Briefly cells were lysed in cold phosphatebuffered saline (PBS) [supplemented with 2120583gmL leupeptin2 120583gmL antipain and 05mM PMSF (all obtained fromSigma Aldrich)] and centrifuged at 4∘C for 2min at 12000 gPellets were resuspended in lysis buffer [10mM HEPESpH 79 15mM MgCl
2 10mM KCl 01mM EDTA 05mM
PMSF 2 120583gmL leupeptin 2120583gmL antipain 3mM sodiumorthovanadate 30mM sodium fluoride and 20mM sodiumpyrophosphate (all obtained from Sigma Aldrich)] and incu-bated on ice for 15min After that NP-40 (Sigma Aldrich)(final concentration of 05) was added and samples werevigorously mixed and centrifuged for 30 s at 12000 g Pelletswere then resuspended in extraction buffer [20mM HEPESpH 79 25 glycerol 15mMMgCl
2 300mMNaCl 025mM
EDTA 05mM PMSF 2 120583gmL leupeptin and 2 120583gmLantipain (all obtained from Sigma Aldrich)] incubated for20min on ice and centrifuged for 20min at 13000 g at 4∘CThe resulting supernatants containing nuclear proteins werestored at minus80∘C Protein concentration was determined usingthe Bio-Rad (Richmond CA USA) colorimetric assay [34]
212 Electrophoretic Mobility Shift Assay Electrophoreticmobility shift assay (EMSA) for NF-120581B was performedusing gel shift assay kit from Promega (Madison WI USA)[33] NF120581B double-stranded consensus oligonucleotide(51015840-AGTTGAGGGGACTTTCCCAGGC-31015840) was end-labeled with 120574-32P-ATP using T4 polynucleotide kinaseUnincorporated nucleotides were removed by passing thereaction mixture through a Sephadex G-25 spin column(Amersham-Pharmacia Uppsala Sweden) Purified 32P-labeled probe (25000 cpm) was incubated in 20 120583L with5 120583g of nuclear extracts in a binding reaction mixturecontaining 50mM NaCl 02mM EDTA 05mM DTT 4glycerol 10mM TrisndashHCl (pH 75) and 005 120583g of poly(dIndashdC) for 30min at room temperature DNA-proteincomplexes were separated by electrophoresis using a 6nondenaturing acrylamide bisacrylamide (375 1) gel in 05X Tris-borateEDTA (TBE) for 2 h at 150V Gels were vacuumdried and analyzed by autoradiography Autoradiographswere visualized with a photo documentation system DP-001-FDC (Vilber Lourmat Marne la Vallee France) andquantified in NIH ImageJ software (Bethesda MD USA)Several exposure times were analyzed to ensure the linearityof the band intensities
213 Statistical Analysis All data were expressed as mean plusmnSEM and analyzed by Graphpad Prism 50 software using
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
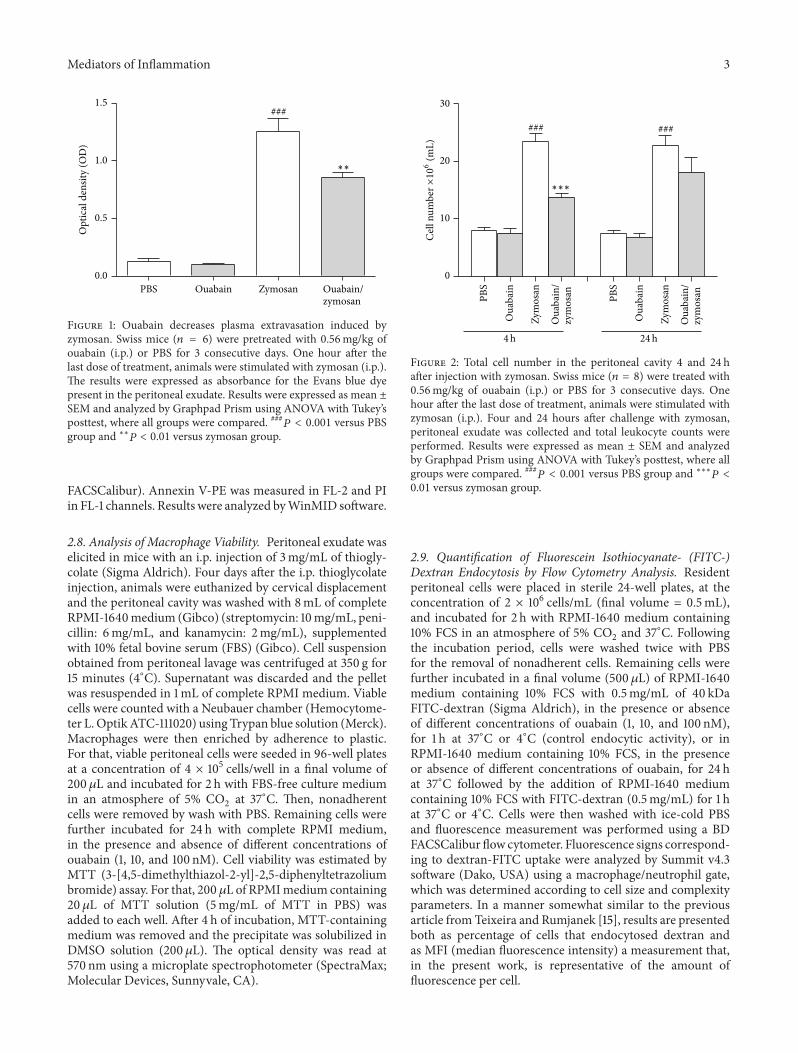
Mediators of Inflammation 3
PBS Ouabain
Zymosan00
05
10
15
Opt
ical
den
sity
(OD
)
lowastlowast
Ouabainzymosan
Figure 1 Ouabain decreases plasma extravasation induced byzymosan Swiss mice (119899 = 6) were pretreated with 056mgkg ofouabain (ip) or PBS for 3 consecutive days One hour after thelast dose of treatment animals were stimulated with zymosan (ip)The results were expressed as absorbance for the Evans blue dyepresent in the peritoneal exudate Results were expressed as mean plusmnSEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquosposttest where all groups were compared 119875 lt 0001 versus PBSgroup and lowastlowast119875 lt 001 versus zymosan group
FACSCalibur) Annexin V-PE was measured in FL-2 and PIin FL-1 channels Results were analyzed byWinMID software
28 Analysis of Macrophage Viability Peritoneal exudate waselicited in mice with an ip injection of 3mgmL of thiogly-colate (Sigma Aldrich) Four days after the ip thioglycolateinjection animals were euthanized by cervical displacementand the peritoneal cavity was washed with 8mL of completeRPMI-1640medium (Gibco) (streptomycin 10mgmL peni-cillin 6mgmL and kanamycin 2mgmL) supplementedwith 10 fetal bovine serum (FBS) (Gibco) Cell suspensionobtained from peritoneal lavage was centrifuged at 350 g for15 minutes (4∘C) Supernatant was discarded and the pelletwas resuspended in 1mL of complete RPMI medium Viablecells were counted with a Neubauer chamber (Hemocytome-ter L OptikATC-111020) using Trypan blue solution (Merck)Macrophages were then enriched by adherence to plasticFor that viable peritoneal cells were seeded in 96-well platesat a concentration of 4 times 105 cellswell in a final volume of200120583L and incubated for 2 h with FBS-free culture mediumin an atmosphere of 5 CO
2at 37∘C Then nonadherent
cells were removed by wash with PBS Remaining cells werefurther incubated for 24 h with complete RPMI mediumin the presence and absence of different concentrations ofouabain (1 10 and 100 nM) Cell viability was estimated byMTT (3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazoliumbromide) assay For that 200120583L of RPMImedium containing20120583L of MTT solution (5mgmL of MTT in PBS) wasadded to each well After 4 h of incubation MTT-containingmedium was removed and the precipitate was solubilized inDMSO solution (200 120583L) The optical density was read at570 nm using a microplate spectrophotometer (SpectraMaxMolecular Devices Sunnyvale CA)
0
10
20
30
PBS
Oua
bain
Zym
osan
lowastlowastlowast
4h 24h
Cel
l num
bertimes
106
(mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Figure 2 Total cell number in the peritoneal cavity 4 and 24 hafter injection with zymosan Swiss mice (119899 = 8) were treated with056mgkg of ouabain (ip) or PBS for 3 consecutive days Onehour after the last dose of treatment animals were stimulated withzymosan (ip) Four and 24 hours after challenge with zymosanperitoneal exudate was collected and total leukocyte counts wereperformed Results were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where allgroups were compared 119875 lt 0001 versus PBS group and lowastlowastlowast119875 lt001 versus zymosan group
29 Quantification of Fluorescein Isothiocyanate- (FITC-)Dextran Endocytosis by Flow Cytometry Analysis Residentperitoneal cells were placed in sterile 24-well plates at theconcentration of 2 times 106 cellsmL (final volume = 05mL)and incubated for 2 h with RPMI-1640 medium containing10 FCS in an atmosphere of 5 CO
2and 37∘C Following
the incubation period cells were washed twice with PBSfor the removal of nonadherent cells Remaining cells werefurther incubated in a final volume (500 120583L) of RPMI-1640medium containing 10 FCS with 05mgmL of 40 kDaFITC-dextran (Sigma Aldrich) in the presence or absenceof different concentrations of ouabain (1 10 and 100 nM)for 1 h at 37∘C or 4∘C (control endocytic activity) or inRPMI-1640 medium containing 10 FCS in the presenceor absence of different concentrations of ouabain for 24 hat 37∘C followed by the addition of RPMI-1640 mediumcontaining 10 FCS with FITC-dextran (05mgmL) for 1 hat 37∘C or 4∘C Cells were then washed with ice-cold PBSand fluorescence measurement was performed using a BDFACSCalibur flow cytometer Fluorescence signs correspond-ing to dextran-FITC uptake were analyzed by Summit v43software (Dako USA) using a macrophageneutrophil gatewhich was determined according to cell size and complexityparameters In a manner somewhat similar to the previousarticle fromTeixeira and Rumjanek [15] results are presentedboth as percentage of cells that endocytosed dextran andas MFI (median fluorescence intensity) a measurement thatin the present work is representative of the amount offluorescence per cell
4 Mediators of Inflammation
84 11
3 2
5 3
73 18
60 14
21 5
21 5
73 1
28 11
33 27
26 14
33 27
4 hours
3 2
72 23
4 3
69 24
24 hours
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
GR-
1G
R-1
GR-
1G
R-1
MAC-3 MAC-3
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
20
40
60
lowastlowast
o
f cel
l Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(b)
Figure 3 Continued
Mediators of Inflammation 5
0
20
40
60
o
f cel
l Mac3+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(c)
0
20
40
60
o
f cel
l Mac3+
Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 3 Effect of ouabain on peritoneal cavity leukocyte subsets Cells were obtained from peritoneal lavage 4 and 24 h after the injection ofthe stimuli ip Neutrophils monocytes and macrophages were distinguished by flow cytometry using anti-Gr-1 and anti-Mac-3 antibodies(a) Representative experiment showing the patterns of Gr-1+ and Mac-3 expression as well as the leukocyte populations defined by thesemolecules Values of the percentage of neutrophils (Gr-1+) macrophages (Mac-3+) and monocytes (Gr-1+ and Mac-3+) were also added tothe dot plots ((b) (c) and (d)) Data were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttestwhere all groups were compared The results were obtained from 6 animals per group
119875 lt 0001 and 119875 lt 005 versus PBS group
lowastlowast
119875 lt 001 versus zymosan group
210 TransmigrationAssay Twodifferent groups of cells weretested for their migratory capacity in vitro The control groupconsisted of resident peritoneal cells obtained from animalsinjected once a day in the three previous days with PBSand the experimental pool obtained from animals injectedwith ouabain (056mgkg) once a day for 3 consecutive daysChemotaxis was measured by migration through a polycar-bonate filter of 5 120583m pore size in 24-well transwell cham-bers (Corning Costar Cambridge MA) DMEM containing05 FCS (500120583L) plus lipopolysaccharide (100 ngmL) ormedium alone as control for spontaneous migration wasadded to the lower chambers 106 cells (100 120583L) were added tothe upper chambers and were incubated at 37∘C in a 5 CO
2
humidified atmosphere After 4 h migration was defined bycounting the cells thatmigrated to the lower chambers by flowcytometry (BD FACSCalibur flow cytometer)
211 Nuclear Extracts Nuclear extracts of peritoneal cellswere prepared based on an adapted version of Rong andBaudry [33] Briefly cells were lysed in cold phosphatebuffered saline (PBS) [supplemented with 2120583gmL leupeptin2 120583gmL antipain and 05mM PMSF (all obtained fromSigma Aldrich)] and centrifuged at 4∘C for 2min at 12000 gPellets were resuspended in lysis buffer [10mM HEPESpH 79 15mM MgCl
2 10mM KCl 01mM EDTA 05mM
PMSF 2 120583gmL leupeptin 2120583gmL antipain 3mM sodiumorthovanadate 30mM sodium fluoride and 20mM sodiumpyrophosphate (all obtained from Sigma Aldrich)] and incu-bated on ice for 15min After that NP-40 (Sigma Aldrich)(final concentration of 05) was added and samples werevigorously mixed and centrifuged for 30 s at 12000 g Pelletswere then resuspended in extraction buffer [20mM HEPESpH 79 25 glycerol 15mMMgCl
2 300mMNaCl 025mM
EDTA 05mM PMSF 2 120583gmL leupeptin and 2 120583gmLantipain (all obtained from Sigma Aldrich)] incubated for20min on ice and centrifuged for 20min at 13000 g at 4∘CThe resulting supernatants containing nuclear proteins werestored at minus80∘C Protein concentration was determined usingthe Bio-Rad (Richmond CA USA) colorimetric assay [34]
212 Electrophoretic Mobility Shift Assay Electrophoreticmobility shift assay (EMSA) for NF-120581B was performedusing gel shift assay kit from Promega (Madison WI USA)[33] NF120581B double-stranded consensus oligonucleotide(51015840-AGTTGAGGGGACTTTCCCAGGC-31015840) was end-labeled with 120574-32P-ATP using T4 polynucleotide kinaseUnincorporated nucleotides were removed by passing thereaction mixture through a Sephadex G-25 spin column(Amersham-Pharmacia Uppsala Sweden) Purified 32P-labeled probe (25000 cpm) was incubated in 20 120583L with5 120583g of nuclear extracts in a binding reaction mixturecontaining 50mM NaCl 02mM EDTA 05mM DTT 4glycerol 10mM TrisndashHCl (pH 75) and 005 120583g of poly(dIndashdC) for 30min at room temperature DNA-proteincomplexes were separated by electrophoresis using a 6nondenaturing acrylamide bisacrylamide (375 1) gel in 05X Tris-borateEDTA (TBE) for 2 h at 150V Gels were vacuumdried and analyzed by autoradiography Autoradiographswere visualized with a photo documentation system DP-001-FDC (Vilber Lourmat Marne la Vallee France) andquantified in NIH ImageJ software (Bethesda MD USA)Several exposure times were analyzed to ensure the linearityof the band intensities
213 Statistical Analysis All data were expressed as mean plusmnSEM and analyzed by Graphpad Prism 50 software using
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
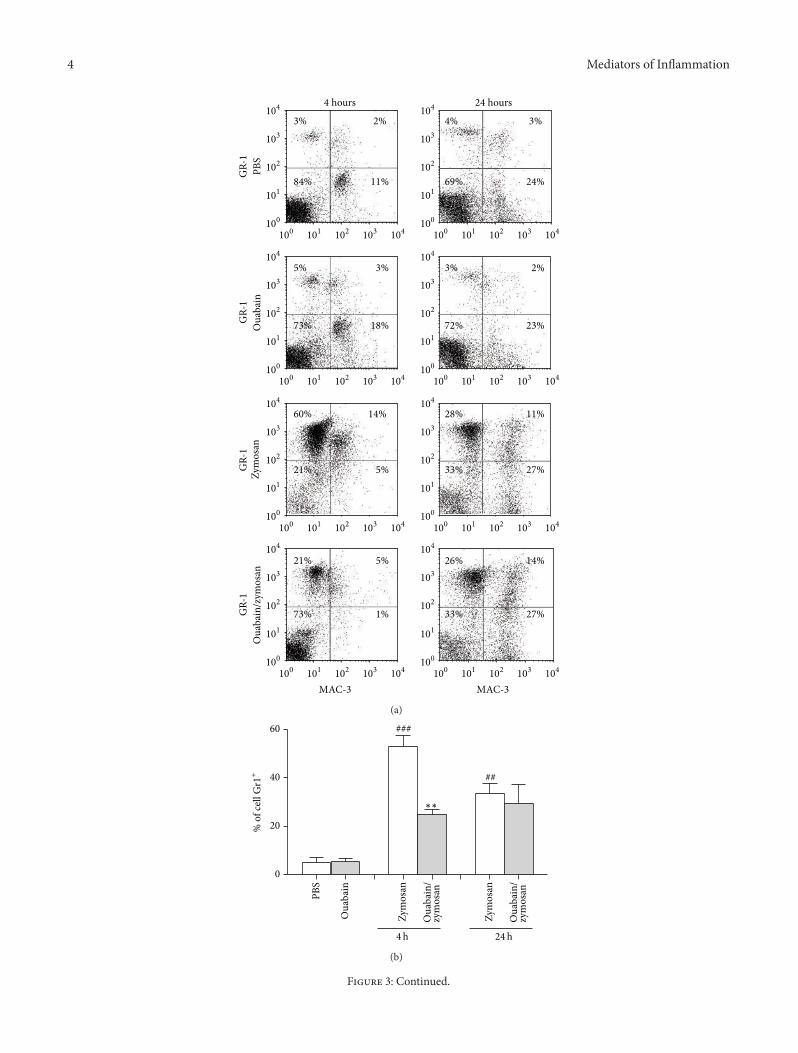
4 Mediators of Inflammation
84 11
3 2
5 3
73 18
60 14
21 5
21 5
73 1
28 11
33 27
26 14
33 27
4 hours
3 2
72 23
4 3
69 24
24 hours
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
GR-
1G
R-1
GR-
1G
R-1
MAC-3 MAC-3
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
20
40
60
lowastlowast
o
f cel
l Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(b)
Figure 3 Continued
Mediators of Inflammation 5
0
20
40
60
o
f cel
l Mac3+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(c)
0
20
40
60
o
f cel
l Mac3+
Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 3 Effect of ouabain on peritoneal cavity leukocyte subsets Cells were obtained from peritoneal lavage 4 and 24 h after the injection ofthe stimuli ip Neutrophils monocytes and macrophages were distinguished by flow cytometry using anti-Gr-1 and anti-Mac-3 antibodies(a) Representative experiment showing the patterns of Gr-1+ and Mac-3 expression as well as the leukocyte populations defined by thesemolecules Values of the percentage of neutrophils (Gr-1+) macrophages (Mac-3+) and monocytes (Gr-1+ and Mac-3+) were also added tothe dot plots ((b) (c) and (d)) Data were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttestwhere all groups were compared The results were obtained from 6 animals per group
119875 lt 0001 and 119875 lt 005 versus PBS group
lowastlowast
119875 lt 001 versus zymosan group
210 TransmigrationAssay Twodifferent groups of cells weretested for their migratory capacity in vitro The control groupconsisted of resident peritoneal cells obtained from animalsinjected once a day in the three previous days with PBSand the experimental pool obtained from animals injectedwith ouabain (056mgkg) once a day for 3 consecutive daysChemotaxis was measured by migration through a polycar-bonate filter of 5 120583m pore size in 24-well transwell cham-bers (Corning Costar Cambridge MA) DMEM containing05 FCS (500120583L) plus lipopolysaccharide (100 ngmL) ormedium alone as control for spontaneous migration wasadded to the lower chambers 106 cells (100 120583L) were added tothe upper chambers and were incubated at 37∘C in a 5 CO
2
humidified atmosphere After 4 h migration was defined bycounting the cells thatmigrated to the lower chambers by flowcytometry (BD FACSCalibur flow cytometer)
211 Nuclear Extracts Nuclear extracts of peritoneal cellswere prepared based on an adapted version of Rong andBaudry [33] Briefly cells were lysed in cold phosphatebuffered saline (PBS) [supplemented with 2120583gmL leupeptin2 120583gmL antipain and 05mM PMSF (all obtained fromSigma Aldrich)] and centrifuged at 4∘C for 2min at 12000 gPellets were resuspended in lysis buffer [10mM HEPESpH 79 15mM MgCl
2 10mM KCl 01mM EDTA 05mM
PMSF 2 120583gmL leupeptin 2120583gmL antipain 3mM sodiumorthovanadate 30mM sodium fluoride and 20mM sodiumpyrophosphate (all obtained from Sigma Aldrich)] and incu-bated on ice for 15min After that NP-40 (Sigma Aldrich)(final concentration of 05) was added and samples werevigorously mixed and centrifuged for 30 s at 12000 g Pelletswere then resuspended in extraction buffer [20mM HEPESpH 79 25 glycerol 15mMMgCl
2 300mMNaCl 025mM
EDTA 05mM PMSF 2 120583gmL leupeptin and 2 120583gmLantipain (all obtained from Sigma Aldrich)] incubated for20min on ice and centrifuged for 20min at 13000 g at 4∘CThe resulting supernatants containing nuclear proteins werestored at minus80∘C Protein concentration was determined usingthe Bio-Rad (Richmond CA USA) colorimetric assay [34]
212 Electrophoretic Mobility Shift Assay Electrophoreticmobility shift assay (EMSA) for NF-120581B was performedusing gel shift assay kit from Promega (Madison WI USA)[33] NF120581B double-stranded consensus oligonucleotide(51015840-AGTTGAGGGGACTTTCCCAGGC-31015840) was end-labeled with 120574-32P-ATP using T4 polynucleotide kinaseUnincorporated nucleotides were removed by passing thereaction mixture through a Sephadex G-25 spin column(Amersham-Pharmacia Uppsala Sweden) Purified 32P-labeled probe (25000 cpm) was incubated in 20 120583L with5 120583g of nuclear extracts in a binding reaction mixturecontaining 50mM NaCl 02mM EDTA 05mM DTT 4glycerol 10mM TrisndashHCl (pH 75) and 005 120583g of poly(dIndashdC) for 30min at room temperature DNA-proteincomplexes were separated by electrophoresis using a 6nondenaturing acrylamide bisacrylamide (375 1) gel in 05X Tris-borateEDTA (TBE) for 2 h at 150V Gels were vacuumdried and analyzed by autoradiography Autoradiographswere visualized with a photo documentation system DP-001-FDC (Vilber Lourmat Marne la Vallee France) andquantified in NIH ImageJ software (Bethesda MD USA)Several exposure times were analyzed to ensure the linearityof the band intensities
213 Statistical Analysis All data were expressed as mean plusmnSEM and analyzed by Graphpad Prism 50 software using
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
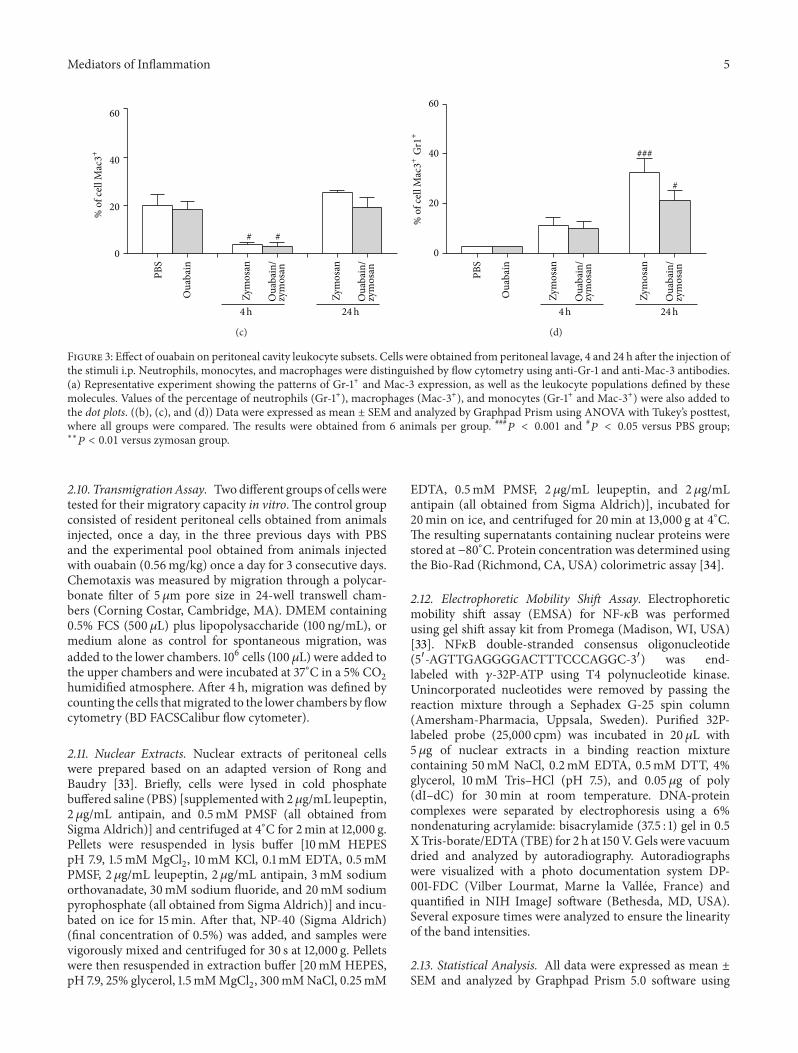
Mediators of Inflammation 5
0
20
40
60
o
f cel
l Mac3+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(c)
0
20
40
60
o
f cel
l Mac3+
Gr1
+
4h 24h
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 3 Effect of ouabain on peritoneal cavity leukocyte subsets Cells were obtained from peritoneal lavage 4 and 24 h after the injection ofthe stimuli ip Neutrophils monocytes and macrophages were distinguished by flow cytometry using anti-Gr-1 and anti-Mac-3 antibodies(a) Representative experiment showing the patterns of Gr-1+ and Mac-3 expression as well as the leukocyte populations defined by thesemolecules Values of the percentage of neutrophils (Gr-1+) macrophages (Mac-3+) and monocytes (Gr-1+ and Mac-3+) were also added tothe dot plots ((b) (c) and (d)) Data were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttestwhere all groups were compared The results were obtained from 6 animals per group
119875 lt 0001 and 119875 lt 005 versus PBS group
lowastlowast
119875 lt 001 versus zymosan group
210 TransmigrationAssay Twodifferent groups of cells weretested for their migratory capacity in vitro The control groupconsisted of resident peritoneal cells obtained from animalsinjected once a day in the three previous days with PBSand the experimental pool obtained from animals injectedwith ouabain (056mgkg) once a day for 3 consecutive daysChemotaxis was measured by migration through a polycar-bonate filter of 5 120583m pore size in 24-well transwell cham-bers (Corning Costar Cambridge MA) DMEM containing05 FCS (500120583L) plus lipopolysaccharide (100 ngmL) ormedium alone as control for spontaneous migration wasadded to the lower chambers 106 cells (100 120583L) were added tothe upper chambers and were incubated at 37∘C in a 5 CO
2
humidified atmosphere After 4 h migration was defined bycounting the cells thatmigrated to the lower chambers by flowcytometry (BD FACSCalibur flow cytometer)
211 Nuclear Extracts Nuclear extracts of peritoneal cellswere prepared based on an adapted version of Rong andBaudry [33] Briefly cells were lysed in cold phosphatebuffered saline (PBS) [supplemented with 2120583gmL leupeptin2 120583gmL antipain and 05mM PMSF (all obtained fromSigma Aldrich)] and centrifuged at 4∘C for 2min at 12000 gPellets were resuspended in lysis buffer [10mM HEPESpH 79 15mM MgCl
2 10mM KCl 01mM EDTA 05mM
PMSF 2 120583gmL leupeptin 2120583gmL antipain 3mM sodiumorthovanadate 30mM sodium fluoride and 20mM sodiumpyrophosphate (all obtained from Sigma Aldrich)] and incu-bated on ice for 15min After that NP-40 (Sigma Aldrich)(final concentration of 05) was added and samples werevigorously mixed and centrifuged for 30 s at 12000 g Pelletswere then resuspended in extraction buffer [20mM HEPESpH 79 25 glycerol 15mMMgCl
2 300mMNaCl 025mM
EDTA 05mM PMSF 2 120583gmL leupeptin and 2 120583gmLantipain (all obtained from Sigma Aldrich)] incubated for20min on ice and centrifuged for 20min at 13000 g at 4∘CThe resulting supernatants containing nuclear proteins werestored at minus80∘C Protein concentration was determined usingthe Bio-Rad (Richmond CA USA) colorimetric assay [34]
212 Electrophoretic Mobility Shift Assay Electrophoreticmobility shift assay (EMSA) for NF-120581B was performedusing gel shift assay kit from Promega (Madison WI USA)[33] NF120581B double-stranded consensus oligonucleotide(51015840-AGTTGAGGGGACTTTCCCAGGC-31015840) was end-labeled with 120574-32P-ATP using T4 polynucleotide kinaseUnincorporated nucleotides were removed by passing thereaction mixture through a Sephadex G-25 spin column(Amersham-Pharmacia Uppsala Sweden) Purified 32P-labeled probe (25000 cpm) was incubated in 20 120583L with5 120583g of nuclear extracts in a binding reaction mixturecontaining 50mM NaCl 02mM EDTA 05mM DTT 4glycerol 10mM TrisndashHCl (pH 75) and 005 120583g of poly(dIndashdC) for 30min at room temperature DNA-proteincomplexes were separated by electrophoresis using a 6nondenaturing acrylamide bisacrylamide (375 1) gel in 05X Tris-borateEDTA (TBE) for 2 h at 150V Gels were vacuumdried and analyzed by autoradiography Autoradiographswere visualized with a photo documentation system DP-001-FDC (Vilber Lourmat Marne la Vallee France) andquantified in NIH ImageJ software (Bethesda MD USA)Several exposure times were analyzed to ensure the linearityof the band intensities
213 Statistical Analysis All data were expressed as mean plusmnSEM and analyzed by Graphpad Prism 50 software using
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
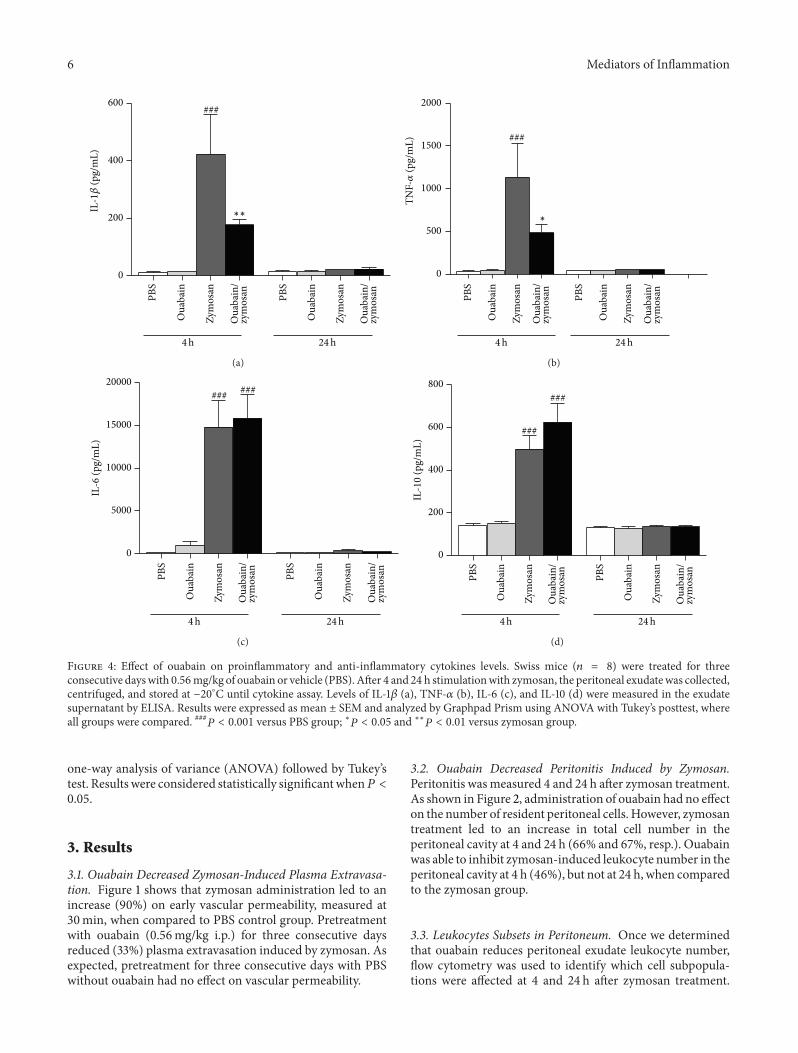
6 Mediators of Inflammation
0
200
400
600
lowastlowast
PBS
Oua
bain
Zym
osan
4h 24h
IL-1120573
(pg
mL)
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an(a)
lowast
4h 24h
0
500
1000
1500
2000
TNF-120572
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(b)
4h 24h
0
5000
10000
15000
20000
IL-6
(pg
mL)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(c)
4h 24h
0
200
400
600
800
IL-1
0 (p
gm
L)
PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an PBS
Oua
bain
Zym
osan
Oua
bain
zy
mos
an
(d)
Figure 4 Effect of ouabain on proinflammatory and anti-inflammatory cytokines levels Swiss mice (119899 = 8) were treated for threeconsecutive dayswith 056mgkg of ouabain or vehicle (PBS) After 4 and 24 h stimulationwith zymosan the peritoneal exudatewas collectedcentrifuged and stored at minus20∘C until cytokine assay Levels of IL-1120573 (a) TNF-120572 (b) IL-6 (c) and IL-10 (d) were measured in the exudatesupernatant by ELISA Results were expressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest whereall groups were compared 119875 lt 0001 versus PBS group lowast119875 lt 005 and lowastlowast119875 lt 001 versus zymosan group
one-way analysis of variance (ANOVA) followed by Tukeyrsquostest Results were considered statistically significantwhen119875 lt005
3 Results
31 Ouabain Decreased Zymosan-Induced Plasma Extravasa-tion Figure 1 shows that zymosan administration led to anincrease (90) on early vascular permeability measured at30min when compared to PBS control group Pretreatmentwith ouabain (056mgkg ip) for three consecutive daysreduced (33) plasma extravasation induced by zymosan Asexpected pretreatment for three consecutive days with PBSwithout ouabain had no effect on vascular permeability
32 Ouabain Decreased Peritonitis Induced by ZymosanPeritonitis wasmeasured 4 and 24 h after zymosan treatmentAs shown in Figure 2 administration of ouabain had no effecton the number of resident peritoneal cells However zymosantreatment led to an increase in total cell number in theperitoneal cavity at 4 and 24 h (66 and 67 resp) Ouabainwas able to inhibit zymosan-induced leukocyte number in theperitoneal cavity at 4 h (46) but not at 24 h when comparedto the zymosan group
33 Leukocytes Subsets in Peritoneum Once we determinedthat ouabain reduces peritoneal exudate leukocyte numberflow cytometry was used to identify which cell subpopula-tions were affected at 4 and 24 h after zymosan treatment
Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 7
PBS
Oua
bain
Zym
osan
Oua
bain
zym
osan
PIPI
PIPI
Annexin Annexin
05 4
95 08
24 hours
04 2
97 06
4 4
94 2
05 3
95 2
07 5
92 3
4 hours
2 5
90 3
2 13
777
2 15
480
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
104
103
102
101
100
104103102101100
(a)
0
5
10
15
Early-apoptosis Late-apoptosis Necrosis
Cel
l (
)
lowastlowastlowast
lowast
PBS
PBS
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in PB
SO
uaba
in
Oua
bain
Oua
bain
Zym
osan
4h
zym
osan
4h
Oua
bain
zym
osan
24
hO
uaba
in
Zym
osan
24h
Zym
osan
24h
Zym
osan
24h
(b)
Figure 5 Effect of ouabain on peritoneal leukocytes viability Viability of the leukocytes present in peritoneal lavage was assessed by flowcytometry For that cells obtained by peritoneal lavage were stained with Annexin V and propidium iodide in order to evaluate the percentageof cells undergoing apoptosis and necrosis respectively (a) Representative dot plots showing the percentage of cells undergoing initialapoptosis (Annexin V+) late-apoptosis (Annexin V+ and PI+) and necrosis (PI+) (b) Data were expressed as mean plusmn SEM and analyzedby Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Results were obtained from 6 animals per grouplowastlowastlowast
119875 lt 0001 and lowast119875 lt 005 versus PBS group
As expected no significant difference in cell subpopula-tions was observed between PBS and ouabain groups (Fig-ures 3(a) 3(b) 3(c) and 3(d)) However four hours afterzymosan ip injection there was a significant increase (90)in neutrophils (Gr1+) at the site Also ouabain pretreat-ment was able to reduce neutrophil cell population (53)when compared to zymosan group (Figure 3(b)) Monocyte(Gr1+Mac3+) and macrophage (Mac3+) cell numbers wereonly enhanced significantly 24 h after zymosan injection atthis time-point ouabain pretreatment was not able to inhibitmonocyte or macrophages cell number (Figures 3(c) and3(d))
34 Ouabain Modulates Cytokines Induced by ZymosanReduced neutrophil migration promoted by ouabain couldbe a consequence of the modulation of cytokines levels(TNF-120572 IL-1120573 IL-6 and IL-10) at 4 and 24 h after zymosanstimulation Figure 4 indicates that animals stimulated withzymosan showed increased levels of TNF-120572 IL-1120573 IL-6 andIL-10 (96 97 99 and 72 resp) in the peritoneal cavityat 4 h but these levels returned to baseline values at 24 hOuabain pretreatment for three consecutive days reducedsignificantly IL-1120573 (58) and TNF-120572 (57) levels (Figures4(a) and 4(b)) at 4 h but not at 24 h when compared to thezymosan group On the other hand ouabain did not affect
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
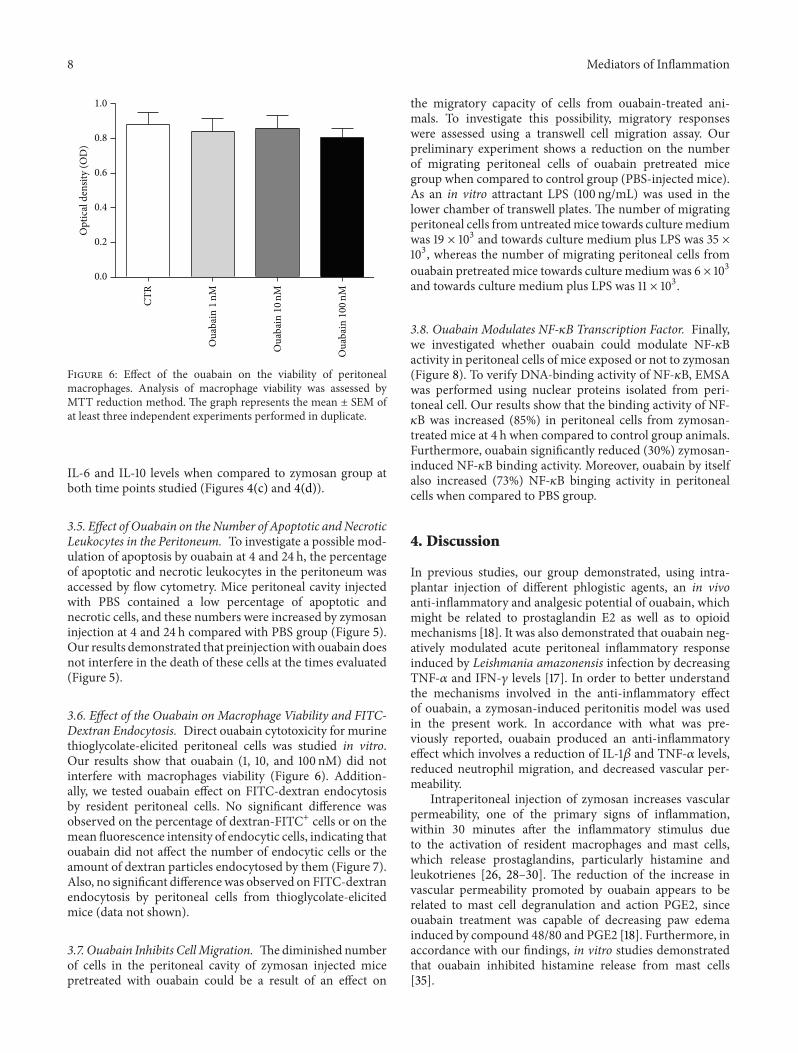
8 Mediators of Inflammation
00
02
04
06
08
10
Opt
ical
den
sity
(OD
)
CTR
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Figure 6 Effect of the ouabain on the viability of peritonealmacrophages Analysis of macrophage viability was assessed byMTT reduction method The graph represents the mean plusmn SEM ofat least three independent experiments performed in duplicate
IL-6 and IL-10 levels when compared to zymosan group atboth time points studied (Figures 4(c) and 4(d))
35 Effect of Ouabain on the Number of Apoptotic andNecroticLeukocytes in the Peritoneum To investigate a possible mod-ulation of apoptosis by ouabain at 4 and 24 h the percentageof apoptotic and necrotic leukocytes in the peritoneum wasaccessed by flow cytometry Mice peritoneal cavity injectedwith PBS contained a low percentage of apoptotic andnecrotic cells and these numbers were increased by zymosaninjection at 4 and 24 h compared with PBS group (Figure 5)Our results demonstrated that preinjectionwith ouabain doesnot interfere in the death of these cells at the times evaluated(Figure 5)
36 Effect of the Ouabain on Macrophage Viability and FITC-Dextran Endocytosis Direct ouabain cytotoxicity for murinethioglycolate-elicited peritoneal cells was studied in vitroOur results show that ouabain (1 10 and 100 nM) did notinterfere with macrophages viability (Figure 6) Addition-ally we tested ouabain effect on FITC-dextran endocytosisby resident peritoneal cells No significant difference wasobserved on the percentage of dextran-FITC+ cells or on themean fluorescence intensity of endocytic cells indicating thatouabain did not affect the number of endocytic cells or theamount of dextran particles endocytosed by them (Figure 7)Also no significant difference was observed on FITC-dextranendocytosis by peritoneal cells from thioglycolate-elicitedmice (data not shown)
37 Ouabain Inhibits CellMigration Thediminished numberof cells in the peritoneal cavity of zymosan injected micepretreated with ouabain could be a result of an effect on
the migratory capacity of cells from ouabain-treated ani-mals To investigate this possibility migratory responseswere assessed using a transwell cell migration assay Ourpreliminary experiment shows a reduction on the numberof migrating peritoneal cells of ouabain pretreated micegroup when compared to control group (PBS-injected mice)As an in vitro attractant LPS (100 ngmL) was used in thelower chamber of transwell plates The number of migratingperitoneal cells fromuntreatedmice towards culturemediumwas 19 times 103 and towards culture medium plus LPS was 35 times103 whereas the number of migrating peritoneal cells fromouabain pretreatedmice towards culture mediumwas 6 times 103and towards culture medium plus LPS was 11 times 103
38 Ouabain Modulates NF-120581B Transcription Factor Finallywe investigated whether ouabain could modulate NF-120581Bactivity in peritoneal cells of mice exposed or not to zymosan(Figure 8) To verify DNA-binding activity of NF-120581B EMSAwas performed using nuclear proteins isolated from peri-toneal cell Our results show that the binding activity of NF-120581B was increased (85) in peritoneal cells from zymosan-treated mice at 4 h when compared to control group animalsFurthermore ouabain significantly reduced (30) zymosan-induced NF-120581B binding activity Moreover ouabain by itselfalso increased (73) NF-120581B binging activity in peritonealcells when compared to PBS group
4 Discussion
In previous studies our group demonstrated using intra-plantar injection of different phlogistic agents an in vivoanti-inflammatory and analgesic potential of ouabain whichmight be related to prostaglandin E2 as well as to opioidmechanisms [18] It was also demonstrated that ouabain neg-atively modulated acute peritoneal inflammatory responseinduced by Leishmania amazonensis infection by decreasingTNF-120572 and IFN-120574 levels [17] In order to better understandthe mechanisms involved in the anti-inflammatory effectof ouabain a zymosan-induced peritonitis model was usedin the present work In accordance with what was pre-viously reported ouabain produced an anti-inflammatoryeffect which involves a reduction of IL-1120573 and TNF-120572 levelsreduced neutrophil migration and decreased vascular per-meability
Intraperitoneal injection of zymosan increases vascularpermeability one of the primary signs of inflammationwithin 30 minutes after the inflammatory stimulus dueto the activation of resident macrophages and mast cellswhich release prostaglandins particularly histamine andleukotrienes [26 28ndash30] The reduction of the increase invascular permeability promoted by ouabain appears to berelated to mast cell degranulation and action PGE2 sinceouabain treatment was capable of decreasing paw edemainduced by compound 4880 and PGE2 [18] Furthermore inaccordance with our findings in vitro studies demonstratedthat ouabain inhibited histamine release from mast cells[35]
Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 9
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(a)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
(b)
0
20
40
60
80
100
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM
Dex
tran
-FIT
C+ce
lls (
)
(c)
0
10
20
30
40
50
MFI
of d
extr
an-F
ITC
(au
)
CTR4∘ C
CTR37∘ C
Oua
bain
1nM
Oua
bain
10nM
Oua
bain
100
nM(d)
Figure 7 Effect of ouabain on in vitro dextran endocytosis by resident peritoneal leukocytes ((a) (b)) Resident peritoneal cells wereincubated with 1 10 and 100 nM ouabain and 05mgmL of FITC-conjugated dextran for 1 hour at 37∘C or 4∘C (control endocytic activity)and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (a) and the medianfluorescence intensity of dextran-FITC (b) ((c) (d)) Prior to the endocytic assay resident peritoneal cells were cultured in the absence orpresence of ouabain for 24 hours After this culture period these cells were then incubated for 1 hour with FITC-conjugated dextran at 37∘Cor 4∘C and fluorescence signals were analyzed by flow cytometry Scatter plots indicate the percentage of dextran-FITC+ cells (c) and themedian fluorescence intensity of dextran-FITC (d) Lines refer to the median of the results obtained from at least 7 animals per group
Zymosan injection in Swiss mice induces massive poly-morphonuclear leukocytes (PMN) influx with maximal cellaccumulation at the 4th hour of peritonitis [27 28 32]In the present study ouabain pretreatment for three con-secutive days produced anti-inflammatory effects decreas-ing the number of intraperitoneal PMNs at the 4th hourThese data are consistent with earlier studies by our groupwhere ouabain reduced the number of PMNs in modelsof peritonitis induced by Concanavalin A and Leishmaniaamazonensis [17 18] Our results show a reduction of thenumber of peritoneal cells of ouabain pretreated mice groupsuggesting that inhibition of cell migration may be a key
event for the ouabain-induced decrease in the number ofperitoneal leukocytes observed by us Furthermore ourpreliminary in vitro study using a transwell model indicatedthat peritoneal cavity cells from ouabain treatedmicemigrateless than those of control animals In addition other studieshave demonstrated that ouabain inhibits in vitro neutrophilmigration by interfering with IL-8 receptor recycling [36]Besides that oleandrin another cardiac glycoside potentiallyinhibited IL-8-mediated biological responses in diverse celltypes [37] Furthermore in studies with lung tumor cellscardiotonic glycosides ouabain and odoroside A inhibitedICAM-1 adhesion molecule expression [38]
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
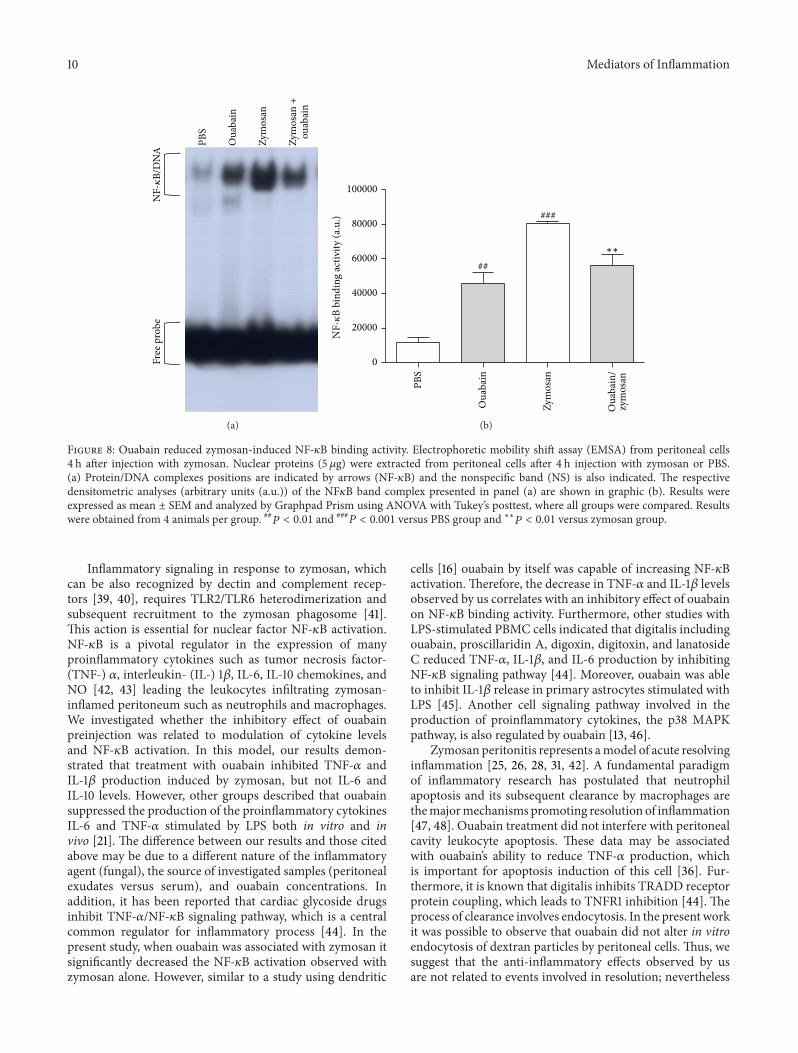
10 Mediators of Inflammation
Free
pro
beN
F-120581
BD
NA
PBS
Oua
bain
Zym
osan
Zym
osan
+ou
abai
n
(a)
0
20000
40000
60000
80000
100000
PBS
Oua
bain
Zym
osan
NF-120581
B bi
ndin
g ac
tivity
(au
)
lowastlowast
Oua
bain
zy
mos
an
(b)
Figure 8 Ouabain reduced zymosan-induced NF-120581B binding activity Electrophoretic mobility shift assay (EMSA) from peritoneal cells4 h after injection with zymosan Nuclear proteins (5 120583g) were extracted from peritoneal cells after 4 h injection with zymosan or PBS(a) ProteinDNA complexes positions are indicated by arrows (NF-120581B) and the nonspecific band (NS) is also indicated The respectivedensitometric analyses (arbitrary units (au)) of the NF120581B band complex presented in panel (a) are shown in graphic (b) Results wereexpressed as mean plusmn SEM and analyzed by Graphpad Prism using ANOVA with Tukeyrsquos posttest where all groups were compared Resultswere obtained from 4 animals per group 119875 lt 001 and
119875 lt 0001 versus PBS group and lowastlowast119875 lt 001 versus zymosan group
Inflammatory signaling in response to zymosan whichcan be also recognized by dectin and complement recep-tors [39 40] requires TLR2TLR6 heterodimerization andsubsequent recruitment to the zymosan phagosome [41]This action is essential for nuclear factor NF-120581B activationNF-120581B is a pivotal regulator in the expression of manyproinflammatory cytokines such as tumor necrosis factor-(TNF-) 120572 interleukin- (IL-) 1120573 IL-6 IL-10 chemokines andNO [42 43] leading the leukocytes infiltrating zymosan-inflamed peritoneum such as neutrophils and macrophagesWe investigated whether the inhibitory effect of ouabainpreinjection was related to modulation of cytokine levelsand NF-120581B activation In this model our results demon-strated that treatment with ouabain inhibited TNF-120572 andIL-1120573 production induced by zymosan but not IL-6 andIL-10 levels However other groups described that ouabainsuppressed the production of the proinflammatory cytokinesIL-6 and TNF-120572 stimulated by LPS both in vitro and invivo [21] The difference between our results and those citedabove may be due to a different nature of the inflammatoryagent (fungal) the source of investigated samples (peritonealexudates versus serum) and ouabain concentrations Inaddition it has been reported that cardiac glycoside drugsinhibit TNF-120572NF-120581B signaling pathway which is a centralcommon regulator for inflammatory process [44] In thepresent study when ouabain was associated with zymosan itsignificantly decreased the NF-120581B activation observed withzymosan alone However similar to a study using dendritic
cells [16] ouabain by itself was capable of increasing NF-120581Bactivation Therefore the decrease in TNF-120572 and IL-1120573 levelsobserved by us correlates with an inhibitory effect of ouabainon NF-120581B binding activity Furthermore other studies withLPS-stimulated PBMC cells indicated that digitalis includingouabain proscillaridin A digoxin digitoxin and lanatosideC reduced TNF-120572 IL-1120573 and IL-6 production by inhibitingNF-120581B signaling pathway [44] Moreover ouabain was ableto inhibit IL-1120573 release in primary astrocytes stimulated withLPS [45] Another cell signaling pathway involved in theproduction of proinflammatory cytokines the p38 MAPKpathway is also regulated by ouabain [13 46]
Zymosan peritonitis represents amodel of acute resolvinginflammation [25 26 28 31 42] A fundamental paradigmof inflammatory research has postulated that neutrophilapoptosis and its subsequent clearance by macrophages arethemajormechanisms promoting resolution of inflammation[47 48] Ouabain treatment did not interfere with peritonealcavity leukocyte apoptosis These data may be associatedwith ouabainrsquos ability to reduce TNF-120572 production whichis important for apoptosis induction of this cell [36] Fur-thermore it is known that digitalis inhibits TRADD receptorprotein coupling which leads to TNFR1 inhibition [44] Theprocess of clearance involves endocytosis In the present workit was possible to observe that ouabain did not alter in vitroendocytosis of dextran particles by peritoneal cells Thus wesuggest that the anti-inflammatory effects observed by usare not related to events involved in resolution nevertheless
Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 11
more detailed studies should be performed to confirm thishypothesis
5 Conclusion
Ouabain modulated acute inflammatory responses inducedby zymosan reducing IL-1120573 and TNF-120572 levels and the num-ber of peritoneal cells possibly due to a decrease in cellmigra-tion Moreover ouabain inhibited NF-120581B activation whichis related to the production of TNF-120572 and IL-1 cytokinesimportant for leucocyte extravasation Taken together ourresults provide new evidences for the mechanisms related tothe anti-inflammatory effects of ouabain in vivo
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by CNPq (Conselho Nacionalde Desenvolvimento Cientıfico e Tecnologico) CAPES(Coordenacao de Aperfeicoamento de Pessoal de Nıvel Supe-rior) and FAPERJ (Fundacao de Amparo a Pesquisa doEstado do Rio de Janeiro)
References
[1] M P Blaustein J Zhang L Chen et al ldquoThe pump theexchanger and endogenous ouabain signaling mechanismsthat link salt retention to hypertensionrdquo Hypertension vol 53no 2 pp 291ndash298 2009
[2] A Y Bagrov and J I Shapiro ldquoEndogenous digitalis patho-physiologic roles and therapeutic applicationsrdquo Nature ClinicalPractice Nephrology vol 4 no 7 pp 378ndash392 2008
[3] W Schoner and G Scheiner-Bobis ldquoEndogenous and exoge-nous cardiac glycosides their roles in hypertension saltmetabolism and cell growthrdquoAmerican Journal of PhysiologymdashCell Physiology vol 293 no 2 pp 509ndash536 2007
[4] J Laredo B P Hamilton and J M Hamlyn ldquoOuabain issecreted by bovine adrenocortical cellsrdquo Endocrinology vol 135no 2 pp 794ndash797 1994
[5] N Bauer J Muller-Ehmsen U Kramer et al ldquoOuabain-likecompound changes rapidly on physical exercise in humans anddogs effects of 120573-blockade and angiotensin-converting enzymeinhibitionrdquo Hypertension vol 45 no 5 pp 1024ndash1028 2005
[6] A Goto K Yamada H Nagoshi Y Terano and M OmataldquoStress-induced elevation of ouabainlike compound in ratplasma and adrenalrdquoHypertension vol 26 no 6 pp 1173ndash11761995
[7] W Schoner ldquoOuabain a new steroid hormone of adrenal glandand hypothalamusrdquo Experimental and Clinical Endocrinology ampDiabetes vol 108 no 7 pp 449ndash454 2000
[8] S Rodrigues-Mascarenhas A D S de Oliveira N D AmoedoO R Affonso-Mitidieri F D Rumjanek and V M RumjanekldquoModulation of the immune system by ouabainrdquo Annals of theNew York Academy of Sciences vol 1153 pp 153ndash163 2009
[9] B Olej N F dos Santos L Leal and VM Rumjanek ldquoOuabaininduces apoptosis on PHA-activated lymphocytesrdquo BioscienceReports vol 18 no 1 pp 1ndash7 1998
[10] V L G deMoraes B Olej L de la Rocque andVM RumjanekldquoLack of sensitivity to ouabain in natural killer activityrdquo TheFASEB Journal vol 3 no 12 pp 2425ndash2429 1989
[11] M Szamel and K Resch ldquoInhibition of lymphocyte activationby ouabain Interference with the early activation of membranephospholipid metabolismrdquo Biochimica et Biophysica Acta vol647 no 2 pp 297ndash301 1981
[12] S R Mascarenhas J Echevarria-Lima N F dos Santos andV M Rumjanek ldquoCD69 expression induced by thapsigarginphorbol ester and ouabain on thymocytes is dependent onexternal Ca2+ entryrdquo Life Sciences vol 73 no 8 pp 1037ndash10512003
[13] S Rodrigues-Mascarenhas F F Bloise J Moscat and V MRumjanek ldquoOuabain inhibits p38 activation in thymocytesrdquoCell Biology International vol 32 no 10 pp 1323ndash1328 2008
[14] S Rodrigues-Mascarenhas N F D Santos and V M Rum-janek ldquoSynergistic effect between ouabain and glucocorticoidsfor the induction of thymic atrophyrdquo Bioscience Reports vol 26no 2 pp 159ndash169 2006
[15] M P Teixeira and V M Rumjanek ldquoOuabain affects theexpression of activation markers cytokine production andendocytosis of human monocytesrdquo Mediators of Inflammationvol 2014 Article ID 760368 10 pages 2014
[16] C R Nascimento R C Valente J Echevarria-Lima et al ldquoTheinfluence of Ouabain on human dendritic cells maturationrdquoMediators of Inflammation vol 2014 Article ID 494956 15pages 2014
[17] P L Jacob J A Leite A K A Alves et al ldquoImmunomodulatoryactivity of ouabain in Leishmania leishmania amazonensis-infected Swiss micerdquo Parasitology Research vol 112 no 3 pp1313ndash1321 2013
[18] D I B De Vasconcelos J A Leite L T Carneiro et al ldquoAnti-inflammatory and antinociceptive activity of ouabain in micerdquoMediators of Inflammation vol 2011 Article ID 912925 11 pages2011
[19] J Bereta M C Cohen and M Bereta ldquoStimulatory effect ofouabain on VCAM-1 and iNOS expression in murine endothe-lial cells involvement of NF-120581Brdquo FEBS Letters vol 377 no 1 pp21ndash25 1995
[20] G Sowa and R Przewłocki ldquoOuabain enhances thelipopolysaccharide-induced nitric oxide production by ratperitoneal macrophagesrdquo Immunopharmacology vol 36 no 1pp 95ndash100 1997
[21] AMatsumori K Ono R Nishio et al ldquoModulation of cytokineproduction and protection against lethal endotoxemia by thecardiac glycoside ouabainrdquo Circulation vol 96 no 5 pp 1501ndash1506 1997
[22] A Matsumori K Ono R Nishio Y Nose and S SasayamaldquoAmlodipine inhibits the production of cytokines induced byouabainrdquo Cytokine vol 12 no 3 pp 294ndash297 2000
[23] RMedzhitov ldquoOrigin and physiological roles of inflammationrdquoNature vol 454 no 7203 pp 428ndash435 2008
[24] J L CashG EWhite andDRGreaves ldquoChapter 17 Zymosan-induced peritonitis as a simple experimental system for thestudy of inflammationrdquo Methods in Enzymology vol 461 pp379ndash396 2009
[25] M N Ajuebor L Virag R J Flower M Perretti and CSzabo ldquoRole of inducible nitric oxide synthase in the regulation
12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Mediators of Inflammation
of neutrophil migration in zymosan-induced inflammationrdquoImmunology vol 95 no 4 pp 625ndash630 1998
[26] N S Doherty P Poubelle P Borgeat T H Beaver G LWestrich and N L Schrader ldquoIntraperitoneal injection ofzymosan in mice induces pain inflammation and the synthesisof peptidoleukotrienes and prostaglandin E2rdquo Prostaglandinsvol 30 no 5 pp 769ndash789 1985
[27] M N Ajuebor A M Das L Virag R J Flower C Szabo andM Perretti ldquoRole of resident peritoneal macrophages and mastcells in chemokine production and neutrophil migration inacute inflammation evidence for an inhibitory loop involvingendogenous IL-10rdquo Journal of Immunology vol 162 no 3 pp1685ndash1691 1999
[28] E Kolaczkowska R Seljelid and B Plytycz ldquoRole of mast cellsin zymosan-induced peritoneal inflammation in Balbc andmast cell-deficientWBB6F1micerdquo Journal of Leukocyte Biologyvol 69 no 1 pp 33ndash42 2001
[29] E Kolaczkowska ldquoShedding light on vascular permeability dur-ing peritonitis role of mast cell histamine versus macrophagecysteinyl leukotrienesrdquo InflammationResearch vol 51 no 11 pp519ndash521 2002
[30] E Kolaczkowska S Shahzidi R Seljelid N Van Rooijen andB Plytycz ldquoEarly vascular permeability inmurine experimentalperitonitis is comediated by resident peritoneal macrophagesand mast cells crucial involvement of macrophage-derivedcysteinyl-leukotrienesrdquo Inflammation vol 26 no 2 pp 61ndash712002
[31] E Kolaczkowska B Arnold and G Opdenakker ldquoGelati-nase BMMP-9 as an inflammatory marker enzyme in mousezymosan peritonitis comparison of phase-specific and cell-specific production by mast cells macrophages and neu-trophilsrdquo Immunobiology vol 213 no 2 pp 109ndash124 2008
[32] E Kolaczkowska A Koziol B Plytycz and B Arnold ldquoInflam-matorymacrophages andnot only neutrophils die by apoptosisduring acute peritonitisrdquo Immunobiology vol 215 no 6 pp492ndash504 2010
[33] Y Rong and M Baudry ldquoSeizure activity results in a rapidinduction of nuclear factor-120581B in adult but not juvenile ratlimbic structuresrdquo Journal of Neurochemistry vol 67 no 2 pp662ndash668 1996
[34] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[35] T Okazaki V S Ilea A Okazaki K Wicher R E Reismanand C E Arbesman ldquoInhibition of antigen induced his-tamine release by ouabainrdquo The Journal of Allergy and ClinicalImmunology vol 57 no 5 pp 454ndash462 1976
[36] E Ray and A K Samanta ldquoReceptor-mediated endocytosis ofIL-8 a fluorescent microscopic evidence and implication ofthe process in ligand-induced biological response in humanneutrophilsrdquo Cytokine vol 9 no 8 pp 587ndash596 1997
[37] S K Manna Y Sreenivasan and A Sarkar ldquoCardiac glycosideinhibits IL-8-induced biological responses by downregulatingIL-8 receptors through altering membrane fluidityrdquo Journal ofCellular Physiology vol 207 no 1 pp 195ndash207 2006
[38] Y Takada K Matsuo H Ogura et al ldquoOdoroside A andouabain inhibit Na+K+-ATPase and prevent NF-120581B-inducibleprotein expression by blocking Na+-dependent amino acidtransportrdquo Biochemical Pharmacology vol 78 no 9 pp 1157ndash1166 2009
[39] G D Brown J Herre D L Williams J A Willment A SJ Marshall and S Gordon ldquoDectin-1 mediates the biologicaleffects of 120573-glucansrdquoThe Journal of Experimental Medicine vol197 no 9 pp 1119ndash1124 2003
[40] V le Cabec S Carreno A Moisand C Bordier and IMaridonneau-Parini ldquoComplement receptor 3 (CD11bCD18)mediates type I and type II phagocytosis during nonopsonic andopsonic phagocytosis respectivelyrdquoThe Journal of Immunologyvol 169 no 4 pp 2003ndash2009 2002
[41] AOzinskyDMUnderhill J D Fontenot et al ldquoThe repertoirefor pattern recognition of pathogens by the innate immunesystem is defined by cooperation between toll-like receptorsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 97 no 25 pp 13766ndash13771 2000
[42] M N Ajuebor R J Flower R Hannon et al ldquoEndogenousmonocyte chemoattractant protein-1 recruits monocytes in thezymosan peritonitis modelrdquo Journal of Leukocyte Biology vol63 no 1 pp 108ndash116 1998
[43] T Kawai and S Akira ldquoSignaling to NF-120581B by Toll-likereceptorsrdquoTrends inMolecularMedicine vol 13 no 11 pp 460ndash469 2007
[44] Q Yang W Huang C Jozwik et al ldquoCardiac glycosidesinhibit TNF-120572NF-120581B signaling by blocking recruitment ofTNF receptor-associated death domain to the TNF receptorrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 102 no 27 pp 9631ndash9636 2005
[45] J Forshammar L Block C Lundborg B Biber and EHanssonldquoNaloxone and ouabain in ultralow concentrations restoreNa+K+-ATPase and cytoskeleton in lipopolysaccharide-treatedastrocytesrdquo Journal of Biological Chemistry vol 286 no 36 pp31586ndash31597 2011
[46] Z Wang M Zheng Z Li et al ldquoCardiac glycosides inhibit p53synthesis by a mechanism relieved by Src or MAPK inhibitionrdquoCancer Research vol 69 no 16 pp 6556ndash6564 2009
[47] J M Hallett A E Leitch N A Riley R Duffin C Haslettand A G Rossi ldquoNovel pharmacological strategies for drivinginflammatory cell apoptosis and enhancing the resolution ofinflammationrdquo Trends in Pharmacological Sciences vol 29 no5 pp 250ndash257 2008
[48] S P Hart I Dransfield and A G Rossi ldquoPhagocytosis ofapoptotic cellsrdquoMethods vol 44 no 3 pp 280ndash285 2008
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom