research article ...ocean.otr.usm.edu/~w777157/dytiscid symposium/kuechler_kehl_dettner... · from...
TRANSCRIPT

R E S E A R C H A R T I C L E
Characterizationand localizationofRickettsia sp. inwater beetles ofgenusDeronectes (Coleoptera:Dytiscidae)Stefan Martin Kuchler, Siegfried Kehl & Konrad Dettner
Department of Animal Ecology II, University of Bayreuth, Bayreuth, Germany
Correspondence: Stefan Martin Kuchler,
Department of Animal Ecology II, University of
Bayreuth, Universitatsstraße 30, 95440
Bayreuth, Germany. Tel.: 10049 0921 55
2733; fax: 10049 0921 55 2743; e-mail:
Received 22 September 2008; revised 3
February 2009; accepted 3 February 2009.
First published online 16 March 2009.
DOI:10.1111/j.1574-6941.2009.00665.x
Editor: Michael Wagner
Keywords
Rickettsia; Deronectes; Dytiscidae; citrate
synthase; electron microscopy.
Abstract
In the present study, Rickettsia sp. was detected in four water beetles of the genus
Deronectes (Dytiscidae) for the first time. Rickettsiae were found in 100% of
examined specimens of Deronectes platynotus (45/45), 39.4% of Deronectes aubei
(28/71), 40% of Deronectes delarouzei (2/5) and 33.3% of Deronectes semirufus
(1/3). Analysis of 16S rRNA gene sequences revealed a phylogenetic relationship
with rickettsial isolates of Limonia chorea (Diptera), tentatively classified as
members of the basal ancestral group. Phylogenetic analysis of the gltA (citrate
synthase) gene sequences showed that Deronectes symbionts were closest to
bacterial symbionts from spiders. Ultrastructural examinations revealed typical
morphological features and intracellular arrangements of rickettsiae. The distribu-
tion, transmission and localization of Rickettsia sp. in D. platynotus were studied
using a diagnostic PCR assay and FISH. Eggs from infected females of D. platynotus
were all Rickettsia-positive, indicative of a vertical transmission.
Introduction
Gram-negative bacteria of the genus Rickettsia belong to the
family Rickettsiaceae in the subdivision of the monophyletic
order Rickettsiales (Alphaproteobacteria). All members of the
genus Rickettsia are obligate intracellular bacteria that reside
free in the cytoplasm of eukaryotic cells. Mostly known
as pathogenic endosymbionts of arthropods (ticks, mites,
fleas and lice), rickettsiae cause human and animal diseases
with a worldwide distribution (Eremeeva & Dasch, 2000).
Some species are also capable of infecting and causing
disease in plants (Davis et al., 1998). Rickettsiae are usually
transmitted by arthropods to vertebrates via feces or salivary
secretions. Because of their considerable medical impor-
tance, the genus Rickettsia was traditionally divided into two
groups (Weiss & Moulder, 1984): the spotted fever group,
including Rickettsia rickettsii and allied species, causative
agents of spotted fever; and the typhus group, including
Rickettsia prowazekii and Rickettsia typhi, the agents of
murine and endemic typhus, respectively. In recent years,
new rickettsiae have been discovered that do not belong to
the previously described subgroups. A third group named
‘ancestral group’, partially subdivided into the bellii, leech
and limoniae groups (Perlman et al., 2006; Perotti et al.,
2006), was established (Stothard & Fuerst, 1995; Fournier &
Raoult, 2007), and consists of Rickettsia bellii, Rickettsia
canadensis and additional numerous rickettsial isolates that
have been isolated from different terrestrial arthropods such
as whiteflies, flies, aphids and beetles (Chen et al., 1996;
Fukatsu et al., 2000; Campbell et al., 2004; Gottlieb et al.,
2006; Zchori-Fein et al., 2006). The ancestral group is still an
initial theoretical classification that is not based on
morphological criteria or biochemical features concerning
basal standing rickettsial isolates in the genus. Rickettsiae of
the ancestral group were not shown to be pathogens for
humans or their animal hosts. However, insects are not the
only vector of rickettsiae. Recently, rickettsiae have been
found in spider mite Tetranychus urticae (Hoy & Jeyapra-
kash, 2005), various spider species (Goodacre et al., 2006),
amoeba Nuclearia pattersoni (Dykova et al., 2003), marine
ciliate Diophrys appendiculata (Vannini et al., 2005) and
several leeches from freshwater environments (Kikuchi et al.,
2002; Kikuchi & Fukatsu, 2005), which also cluster into the
ancestral group. Analyses of 16S rRNA genes suggest that
many of these rickettsial isolates could represent new genera
within the order Rickettsiales. As already supposed (Yu &
Walker, 2006), the family Rickettsiaceae could be more
widespread than observed so far.
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

The purpose of this study was to investigate and identify
symbiotic bacteria in selected water beetles of the palaearctic
genus Deronectes. The association of endosymbiotic bacteria
with water insects, especially water beetles, was already
presumed in previous studies (Maddison et al., 1999; Shull
et al., 2001). Representatives of the genus Deronectes are
predacious and colored uniformly black or brown. Dero-
nectes species usually live among gravel in small, swift or
strongly flowing streams with sparse vegetation. Most
species prefer mountainous regions, some occurring at a
high altitude. To date 55 species have been described (Fery &
Brancucci, 1997; Fery et al., 2001).
In the present molecular study, we report the first finding
of a coccobacillus bacterium in Deronectes, which could be
identified as a member of the genus Rickettsia. A total of 136
specimens from seven Deronectes species were examined for
the existence of Rickettsia sp. using a diagnostic PCR assay.
Phylogenetic analysis inferred by 16S rRNA gene and
particularly the rickettsial gltA (citrate synthase) gene
disclosed the phylogenetic position of beetle Rickettsia in
the existing phylogenetic system of Rickettsia endosym-
bionts. FISH was used for the investigation of rickettsial
distribution and vertical transmission rates. The morpholo-
gical characteristics of the bacterium were analyzed by
electron microscopic observations.
Materials and methods
Samples
A total of 136 Deronectes specimens were collected between
2003 and 2008 (Table 1) by kick-sampling (Schwoerbel,
1994). Deronectes platynotus and Deronectes latus were
obtained from Fichtelgebirge, a mountain range in north-
eastern Bavaria, Germany. Deronectes aubei was collected
from Black Forest (Germany), Italy and French alps. Dero-
nectes delarouzei, Deronectes semirufus, D. aubei sanfilippoi
and Deronectes moestus inconspectus came from Spain, Italy
and France. Beetles were brought to laboratory alive and
embedded for histology/FISH or dissected for diagnostic
PCR immediately.
Histology
Before all the beetles were fixed in 4% paraformaldehyde
overnight, the elytra were carefully removed. The fixed
beetles were washed in 1� phosphate-buffered saline and
96% ethanol (1 : 1), dehydrated serially in ethanol (70%,
90%, 2� 100%) and embedded in UNICRYLTM (Plano
GmbH, Germany). Serial sections (2 mm) were cut using a
Leica Jung RM2035 rotary microtome (Leica Instruments
GmbH, Germany), mounted on epoxy-coated glass slides
and subjected to FISH.
DNA extraction, cloning and sequencing
DNA was extracted using a QUIAGEN DNeasys Tissue Kit
(QUIAGEN GmbH, Germany) following the protocol for
animal tissue. The eubacterial 16S rRNA gene was PCR
amplified using the primer set 07F (50-AGAGTTT
GATCMTGGCTCAG-30) and 1507R (50-TACCTTGTTAC
GACTTCAC-30) (Lane, 1991). All PCR reactions were
performed in a Biometra thermal cycler with the following
program: an initial denaturating step at 94 1C for 3 min,
followed by 34 cycles of 94 1C for 30 s, 50 1C for 2 min and
72 1C for 1 min. A final extension step of 72 1C for 10 min
was included. PCR products of the expected sizes were
cloned using the TOPO TA Clonings Kit (Invitrogen, CA).
Suitable clones for sequencing were selected by restriction
fragment length polymorphism (RFLP). Inserts were di-
gested by restriction endonucleases HaeIII and TaqI. Plas-
mids containing the DNA inserts of the expected sizes were
sequenced at the DNA analytics core facility of the
Table 1. Infection rates of Rickettsia sp. in natural populations of Deronectes spp.
Species Locality Date of collection
No. of
individuals
% of infection
(infected/total)
D. platynotus Fichtelgebirge, Bavaria, Germany 2007/2008 45 100 (45/45)
D. aubei Black Forest, Germany, Italy, France 2003–2008 71 39.4 (28/71)
D. latus Fichtelgebirge, Bavaria, Germany 17.10.2007 7 0 (0/7)
D. delarouzei Bonansa, El pont de Suert, Spain 05.10.2007 2
Llesp, El pont de Suert, Spain 10.08.2004 1 40 (2/5)
05.10.2007 1
Saldes, Spain 05.10.2007 1
D. semirufus Grotta all’Onda, Casoli, Lucca, Tuscany, Italy 09.10.2003 1
Sospel, Moulinet, Apuane-Maritimes, France 07.10.2003 1 33.3 (1/3)
Fociomboli, Apuane Alps, Italy 07.09.2007 1
D. aubei sanfilippoi Vernet les Bains, Pyr. Orientales, France 07.10.2007 3 0 (0/3)
D. moestus inconspectus Saldes, Spain 17.10.2007 2 0 (0/2)
FEMS Microbiol Ecol 68 (2009) 201–211c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
202 S.M. Kuchler et al.

University of Bayreuth with M13 forward and M13 reverse
sequencing primers (Invitrogen).
For diagnostic PCR, the 16S rRNA gene was amplified using
the Rickettsia-specific primers Ri_170F (50-GGGCTTGCTCTA
AATTAGTTAGT-30) and Ri_1500R (50-ACGTTAGCTCACC
ACCTTCAGG-30). The citrate synthase gene (gltA) was de-
tected using the primers CS5_F (50-TCTTTATGGGGACC
AGCC-30) and CS4_R (50-TTTCCATTGTGCCATCCAG-30).
PCR primers were designed using the probe design tool of the
ARB software package (Ludwig et al., 2004). Diagnostic PCR
was performed under the temperature profile described above.
FISH
The following probes were used for FISH targeted to the 16S
rRNA gene: eubacterial probe EUB338 [50-(Cy5)-GCTGCCT
CCCGTAGGAGT-30] (Amann et al., 1995), EUB388 II [50-
(Cy3)-GCAGCCACCCGTAGGTGT-30], EUB338 III [50-(Cy3)-
GCTGCCACCCGTAGGTGT-30] (Daims et al., 1999) and
Rickettsia-specific probe Rick_B1 [50-(Cy3)-CCATCATCCC
CTACTACA-30] (Perotti et al., 2006). Additionally, a nonsense
probe complementary to EUB338, NON338 [50-(Cy3)-
ACTCCTACGGGAGGCAGC-30] (Manz et al., 1992) was
used as a negative control of the hybridization protocol.
Slides were hybridized in hybridization buffer [20 mM Tris-
HCl (pH 8.0), 0.9 M NaCl, 0.01% sodium dodecyl sulfate
(SDS), and 20% formamide] containing 10 pmol of fluor-
escent probes per milliliter, incubated at 46 1C for 90 min,
rinsed in washing buffer [20 mM Tris-HCl (pH 8.0),
225 mM NaCl, 5 mM EDTA, and 0.01% SDS], mounted
with antibleaching solution (VECTASHIELDs Mounting
Medium; Vector Laboratories, UK) and viewed under a
fluorescent microscope.
Electron microscopy
Male accessory glands of D. platynotus were fixed in 2.5%
glutaraldehyde in 0.1 M cacodylate buffer (pH 7.3) for 1 h,
embedded in 2% agarose and fixed again in 2.5% glutar-
aldehyde in 0.1 M cacodylate buffer (pH 7.3) overnight.
Specimens were washed in 0.1 M cacodylate three times for
20 min. Following postfixation in 2% osmium tetroxide for
2 h, the specimens were washed and stained en bloc in 2%
uranyl acetate for 90 min. After fixation, specimens were
dehydrated serially in ethanol (30%, 50%, 70%, 95% and
3� 100%), transferred to propylene oxide and embedded in
Epon. Ultrathin sections (70 nm) were cut with a diamond
knife (MicroStar, Huntsville, TX) on a Leica Ultracut UCT
microtome (Leica Microsystems, Vienna, Austria). Ultra-
thin sections were mounted on pioloform-coated copper
grids, and stained with saturated uranyl acetate, followed by
lead citrate. The sections were viewed with a Zeiss CEM 902
A transmission electron microscope (Carl Zeiss, Oberko-
chen, Germany) at 80 kV. Micrographs were taken using an
SO-163 EM film (Eastman Kodak, Rochester, NY).
Phylogenetic analysis
High-quality sequences of the 16S rRNA gene and the gltA
gene for citrate synthase were aligned with the CLUSTALW
software in BIOEDIT (Hall, 1999) and transferred to the ARB
software package and edited manually. Phylogenetic ana-
lyses using maximum parsimony were performed using
PAUP� version 4.0b10 (Swofford, 2000). The heuristic
search included 10 000 random addition replicates and tree
bissection–reconnection (TBR) branch swapping with the
Multrees option. A 50% majority-rule consensus tree of the
most parsimonious tree was constructed and exported to
ARB program package, where branch lengths were calculated.
Bootstrap analyses were performed with 500 replicates
with TBR branch swapping, 25 random addition repli-
cates and the Multrees option in PAUP�. The gltA citrate
synthase sequences were checked for chimeras using
CCODE (Gonzalez et al., 2005).
Sequence data
Clone sequences of 16S rRNA gene and gltA sequences for the
D. platynotus-, D. aubei-, D. semirufus- and D. delarouzei-
associated Rickettsia sp. were deposited in the DDBJ/EMBL/
GenBank nucleotide sequence databases with the accession
numbers FM177868–FM177878 and FM955310–FM955315,
respectively.
Results
Identification of bacterial symbiont
A 1.5-kb segment of the eubacterial 16S rRNA gene was
amplified by PCR and was subjected to cloning and RFLP
typing. Overall, 19 clones were sequenced and compared with
other sequences found in GenBank. The nucleotide sequences
of clones exhibited 99% similarity to the sequence of Rick-
ettsia limoniae (AF322442, AF322443) from cranefly L. chorea
(Diptera, Limoniidae). The validity of the sequences was
confirmed by FISH performed with probe Rick_B1. Further-
more, we sequenced a 416-bp fragment of the gltA gene
(citrate synthase) from four Rickettsia-positive D. platynotus
beetles as well as from D. aubei (404 bp), D. semirufus
(367 bp) and D. delarouzei (401 bp). The gltA sequences from
D. platynotus rickettsiae were 100% identical to rickettsial
isolate (DQ 231491) from spider Pityophantes phrygianus
(Goodacre et al., 2006). The gltA gene sequences from
rickettsial symbionts of D. aubei and D. delarouzei exhibited
99% similarity to the sequence of P. phrygianus, whereas
rickettsial gltA sequences from D. semirufus showed 98%
similarity to Rickettsia endosymbionts of spider Meta menegi.
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
203Rickettsia endosymbiont in water beetles

Screening for the presence of Rickettsia sp.
Six Deronectes species and one subspecies were examined
using Rickettsia-specific primers in a diagnostic PCR assay
(Table 1). A total of 45 D. platynotus were screened. Rickettsia
sp. could be detected in 100% of all tested D. platynotus. In
contrast, only 39.4% of 71 examined individuals of D. aubei
were infected. Both species were collected in different seasons
and populations. Just two individuals of D. delarouzei and
one individual of D. semirufus were found to be positive for
the Rickettsia symbiont. For elaboration of infection rates in
these two species, more specimens from different populations
have to be examined for rickettsiae. No rickettsiae were
detected in seven individuals of D. latus. In the same way,
three surveys of D. aubei sanfilippoi indicated a negative signal
for Rickettsia. Two examined individuals of D. moestus
inconspectus also did not show any infection of Rickettsia.
However, only a few individuals of D. latus, D. aubei
sanfilippoi and D. moestus inconspectus could be examined
for the presence of rickettsiae. All examined species of
D. platynotus and D. aubei exhibited a well-balanced sex ratio.
Furthermore, there was no evidence for male killing, parthe-
nogenesis or other sex ratio distortion in numerically exam-
ined species of D. platynotus and D. aubei. Current studies
indicated that further water beetles are associated with
Rickettsia sp. too (data not shown). Examined species of
Agabus melanarius, Agabus wasastjernae, Agabus guttatus,
Hydroporus gyllenhalii, Hydroporus tristis, Hydroporus umbro-
sus and Hydroporus obscurus also exhibited Rickettsia infec-
tions.
In situ hybridization of Rickettsia symbiont
FISH revealed a remarkable affection of the Rickettsia
symbiont (Fig. 1a). All tissues that are in contact with the
hemolymph are affected by rickettsiae. The symbionts could
be identified in all compartments of Deronectes, including
the head (ommatidia) (Fig. 1b) and legs. Actually, internal
parts of elytra with soft tissue exhibited bacteria. Especially,
numerous rickettsiae could be detected in tissue of active
metabolism, such as the fatbody and internal reproduction
organs. A lower number of rickettsiae was found in muscles.
In addition, Rickettsia sp. is more abundant in females than
males. Accessory glands of males will be affected predomi-
nantly (Fig. 1c). The strongest signals of a Rickettsia-specific
probe were found in females of D. platynotus. Minor signals
of symbionts were detected in D. aubei, D. semirufus and
D. delarouzei (data not shown).
The distribution of the Rickettsia was also investigated in
eggs and larvae of D. platynotus. The presence of large
numbers of symbionts in oocytes and follicle cells (Fig. 1d),
and the detection of Rickettsia in eggs (Fig. 1e) are indicative
of the vertical transmission of symbionts. In the second and
third larval stages of Deronectes, rickettsiae could be detected
in the entire body. The number of symbionts increased from
stage to stage.
Fig. 1. FISH with specific probe Rick_B1 (Cy3) of
square sections of water beetle Deronectes
platynotus adults. (a) Rickettsia sp. infection of
fatbody in a female abdomen. Scale bar = 40mm.
(b) Square section of head; numerous Rickettsia
signals from ommatidia (arrows). Rickettsia sp.
clustered around the cerebral ganglia. Scale
bar = 40 mm. (c) Male accessory glands infected
by intracytosolic Rickettsia sp. Bacteria are
located near the connective tissue (arrow). Scale
bar = 20 mm. (d) Ovariole infected by Rickettsia
sp. symbiont. Scale bar = 20 mm. (e) Bacterial
symbionts are also present within the egg
(arrow). Surrounding tissue of ovarioles habour-
ing Rickettsia. Scale bar = 50mm. FB, fat body;
B, brain; E, egg; AGT, accessory glandular tissue;
M, muscle; O, ovariole; OM, ommatidia. (a), (b)
and (c) were taken by confocal microscope.
(d) and (e) were taken by a conventional
fluorescence microscope.
FEMS Microbiol Ecol 68 (2009) 201–211c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
204 S.M. Kuchler et al.

Double hybridization performed with Rick_B1, together
with the universal probe EUB338, showed the absence of any
other kind of endosymbiotic bacteria (Fig. 2), with the
exception of gut bacteria inside adult and larval specimens
of Deronectes. FISH accomplished with EUB388 II and
EUB338 III did not show any positive signals (data not
shown).
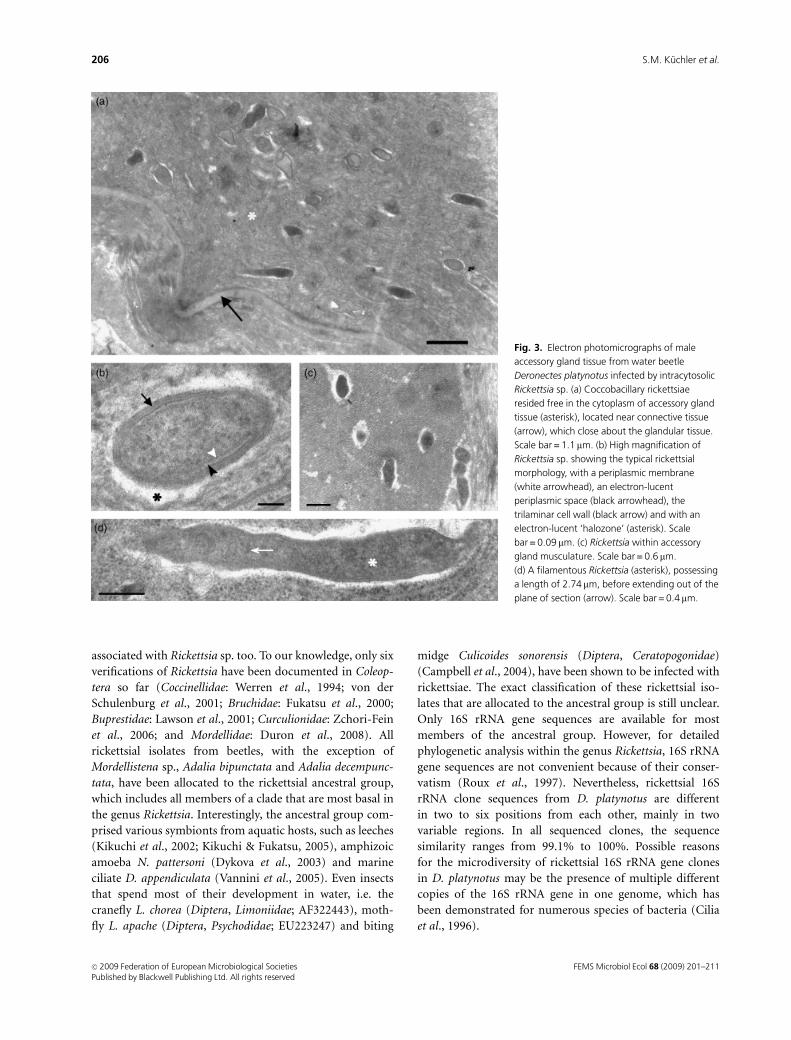
Electron microscopy of the Rickettsia symbiont
Ultrastructural examinations of male accessory glands of
D. platynotus revealed coccobacillary rickettsiae that were
observed free in the cytoplasm (Fig. 3a). Most rickettsiae
ranged from 0.35 mm in width to 0.65 mm in length. Rick-
ettsiae were delineated by an inner periplasmatic membrane,
a periplasmatic space and a trilaminar cell wall with a thicker
inner leaflet, characteristic of the genus Rickettsia. A Rick-
ettsia-typical slime layer or an electron-translucent zone, up
to 60 nm thick, surrounded the cell wall and separated the
bacterium from the cytoplasm of the host cell (Fig. 3b).
Isolated bacteria were also found in the musculature,
enclosing the male accessory gland (Fig. 3c). Some of the
symbionts were much longer and had a ‘filamentous’
appearance. They could be as long as 2.74 mm (Fig. 3d).
Phylogenetic analysis of the Rickettsia symbiont
Phylogenetic analyses on the basis of the 16S rRNA gene
sequence revealed that Rickettsia sp. from Deronectes belong
neither to the ‘spotted fever group’ nor the ‘typhus group,’
but was placed within the ancestral group (100% bootstrap
support) (Fig. 4). Clones of the endosymbiotic bacteria of
D. platynotus, D. aubei and D. delarouzei were placed in the
clade with R. limoniae, Rickettsia endosymbionts of Ceroba-
sis guestifalica (Psocoptera, Trogiidae) and Lutzomyia apache
(Diptera, Psychodinae), whereas Rickettsia sp. of D. semirufus
cluster basal with rickettsiae from leeches. By phylogenetic
analysis based on the gltA gene (Fig. 5), the Rickettsia strain
from Deronectes clustered with Rickettsia endosymbionts of
L. apache and different spider species in a terminal group
(99% bootstrap support) that is distinctly separated from
other Rickettsia species of the spotted fever, typhus and bellii
groups. The phylogenetic position of Deronectes rickettsiae
was the same in the 16S rRNA gene and gltA trees when the
maximum-likelihood, parsimony and distance methods
were used (data not shown). Only the position of the
Rickettsia endosymbiont of D. semirufus varied with differ-
ent methods (MrBayes, data not shown), and was some-
times added to the leech group or to the other rickettsial
endosymbionts of the Deronectes species of the limoniae
group. This explains the low bootstrap support values
in the ancestral group in the phylogenetic tree of 16S rRNA
gene.
Discussion
The present study reports the first molecular identification
of Rickettsia in four water beetle species of the genus
Deronectes and generally for adephagan beetles. Evaluable
results have been obtained of D. platynotus and D. aubei.
Rickettsia infection was detected in 100% (45/45) of
D. platynotus and 39.4% (28/71) of D. aubei. The frequen-
cies of Rickettsia infection were maintained constant in
different seasons. Individuals of D. delarouzei and D. semi-
rufus were also tested positive for rickettsiae, whereas
individuals of D. latus, D. aubei sanfilippoi and D. moestus
inconspectus did not show any signal for symbiotic bacteria.
Remarkably, Rickettsia-positive species of Deronectes do not
cluster in a single group within the phylogenetic tree of
Deronectes. In addition, the current analysis indicated six
more species of Agabus and Hydroporus (Dytiscidae) that are
Fig. 2. FISH double hybridization of the
Rickettsia symbiont on tissue (fatbody) cross-
section from Deronectes platynotus (a–c, scale
bar = 40 mm) and Deronectes aubei (d–f, scale
bar = 20 mm). The left panels (a, d) show FISH
with the probe Cy3-Rick_B1, which visualizes the
Rickettsia symbiont, while middle panels (b, e)
show FISH with the probe Cy5-EUB338, which
recognizes diverse eubacteria universally. The
overlay (c, f) does not show other bacteria.
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
205Rickettsia endosymbiont in water beetles
associated with Rickettsia sp. too. To our knowledge, only six
verifications of Rickettsia have been documented in Coleop-
tera so far (Coccinellidae: Werren et al., 1994; von der
Schulenburg et al., 2001; Bruchidae: Fukatsu et al., 2000;
Buprestidae: Lawson et al., 2001; Curculionidae: Zchori-Fein
et al., 2006; and Mordellidae: Duron et al., 2008). All
rickettsial isolates from beetles, with the exception of
Mordellistena sp., Adalia bipunctata and Adalia decempunc-
tata, have been allocated to the rickettsial ancestral group,
which includes all members of a clade that are most basal in
the genus Rickettsia. Interestingly, the ancestral group com-
prised various symbionts from aquatic hosts, such as leeches
(Kikuchi et al., 2002; Kikuchi & Fukatsu, 2005), amphizoic
amoeba N. pattersoni (Dykova et al., 2003) and marine
ciliate D. appendiculata (Vannini et al., 2005). Even insects
that spend most of their development in water, i.e. the
cranefly L. chorea (Diptera, Limoniidae; AF322443), moth-
fly L. apache (Diptera, Psychodidae; EU223247) and biting
midge Culicoides sonorensis (Diptera, Ceratopogonidae)
(Campbell et al., 2004), have been shown to be infected with
rickettsiae. The exact classification of these rickettsial iso-
lates that are allocated to the ancestral group is still unclear.
Only 16S rRNA gene sequences are available for most
members of the ancestral group. However, for detailed
phylogenetic analysis within the genus Rickettsia, 16S rRNA
gene sequences are not convenient because of their conser-
vatism (Roux et al., 1997). Nevertheless, rickettsial 16S
rRNA clone sequences from D. platynotus are different
in two to six positions from each other, mainly in two
variable regions. In all sequenced clones, the sequence
similarity ranges from 99.1% to 100%. Possible reasons
for the microdiversity of rickettsial 16S rRNA gene clones
in D. platynotus may be the presence of multiple different
copies of the 16S rRNA gene in one genome, which has
been demonstrated for numerous species of bacteria (Cilia
et al., 1996).
Fig. 3. Electron photomicrographs of male
accessory gland tissue from water beetle
Deronectes platynotus infected by intracytosolic
Rickettsia sp. (a) Coccobacillary rickettsiae
resided free in the cytoplasm of accessory gland
tissue (asterisk), located near connective tissue
(arrow), which close about the glandular tissue.
Scale bar = 1.1 mm. (b) High magnification of
Rickettsia sp. showing the typical rickettsial
morphology, with a periplasmic membrane
(white arrowhead), an electron-lucent
periplasmic space (black arrowhead), the
trilaminar cell wall (black arrow) and with an
electron-lucent ‘halozone’ (asterisk). Scale
bar = 0.09 mm. (c) Rickettsia within accessory
gland musculature. Scale bar = 0.6 mm.
(d) A filamentous Rickettsia (asterisk), possessing
a length of 2.74mm, before extending out of the
plane of section (arrow). Scale bar = 0.4 mm.
FEMS Microbiol Ecol 68 (2009) 201–211c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
206 S.M. Kuchler et al.

Fig. 4. Phylogenetic position of different clones of the endosymbiotic bacteria of water beetles Deronectes platynotus, Deronectes aubei,
Deronectes semirufus and Deronectes delarouzei (bold) based on 55 16S rRNA gene sequences (1350 bp, maximum parsimony heuristic search, 50%
majority-rule consensus tree of 840 most parsimonious trees, tree length = 497, CI = 0.71, RI = 0.86). The tree has been rooted with Orientia
tsutsugamushi as an outgroup. Only bootstrap values of at least 50% are shown at nodes (n = 500).
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
207Rickettsia endosymbiont in water beetles
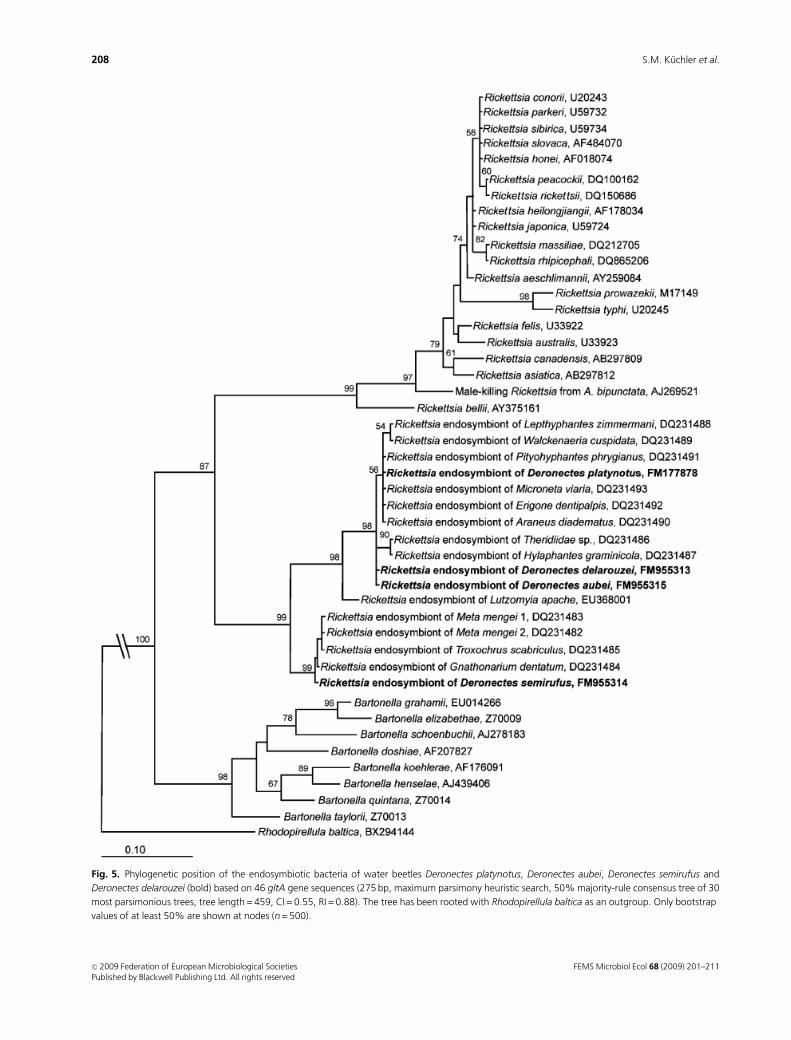
Fig. 5. Phylogenetic position of the endosymbiotic bacteria of water beetles Deronectes platynotus, Deronectes aubei, Deronectes semirufus and
Deronectes delarouzei (bold) based on 46 gltA gene sequences (275 bp, maximum parsimony heuristic search, 50% majority-rule consensus tree of 30
most parsimonious trees, tree length = 459, CI = 0.55, RI = 0.88). The tree has been rooted with Rhodopirellula baltica as an outgroup. Only bootstrap
values of at least 50% are shown at nodes (n = 500).
FEMS Microbiol Ecol 68 (2009) 201–211c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
208 S.M. Kuchler et al.

Referred to rickettsiae of the ancestral group, the citrate
synthase-coding gltA gene sequences, also used for phyloge-
netic analysis, are only available for Rickettsia sp. of L. apache
(EU368001) and rickettsial isolates from spiders (Goodacre
et al., 2006). Sequences of further protein-coding genes,
mostly those belonging to the surface cell antigen (sca)
family, which have been used for the classification of
rickettsial isolates over the past few years (Fournier et al.,
1998; Roux & Raoult, 2000; Sekeyova et al., 2001; Ngwami-
diba et al., 2006), are not available for rickettsiae of the
ancestral group (Fournier et al., 2003).
Phylogenetic analysis based on the 16S rRNA gene
showed that Rickettsia sp. from Deronectes cluster into the
‘limoniae group’, the basal subgroup of the ancestral group.
Furthermore, we succeeded in partial sequencing of the gltA
gene of these rickettsiae. Sequences from Deronectes sym-
bionts resulted in a high concordance with rickettsial
isolates from spiders P. phrygianus and M. mengei (Goodacre
et al., 2006). However, 16S rRNA gene sequences are not
available for spider rickettsiae in GenBank. It is interesting
that sequences of the gltA gene of Rickettsia sp. isolated from
water beetles are quite divergent from all previous valid
Rickettsia strains from the spotted fever, typhus and bellii
groups. Furthermore, gltA sequences of Rickettsia sp. show a
high similarity to the related genus Bartonella, whose
members are known as emerging human pathogens (Min-
nick & Anderson, 2006), but do not cluster with Bartonella
sequences. Goodacre et al. (2006) presumed that spider-
specific Rickettsia form an isolated, terminal group that
contains no symbionts of other arthropod hosts. They
further assumed that frequent horizontal transmission of
rickettsiae between spiders and their insect prey is unlikely
because this would produce a heterogeneous assortment of
non-spider-specific Rickettsia in spiders. In contrast, we were
able to connect molecular data of the 16S rRNA gene
sequences from basal rickettsiae strains and spider-specific
Rickettsia by sequencing of 16S rRNA and gltA genes of the
Deronectes symbiont. We assume that other Rickettsia strains
of the ancestral group could exhibit comparable gltA
sequences. Based on this assumption, Rickettsia from water
beetles and spiders could belong to a new taxon, which might
represent a near relative of the genus Rickettsia. It can be
speculated that sequences of the ompA, ompB genes and gene
D will show appreciable deviations like gltA, if these genes are
present in rickettsiae of the ancestral group in general.
However, ultrastructural observations revealed typical
morphological features and intracellular arrangements of
rickettsiae, such as a thickened inner leaflet or electron-
lucent ‘halos’ (Silverman, 1991). Additionally, some rick-
ettsiae had a ‘filamentous’ appearance, similar to R. bellii
from Amblyomma cooperi ticks (Labruna et al., 2004). The
actual length of these bacteria cells could not be revealed by
transmission electron microscopical sections.
Furthermore, Rickettsia could be detected in all tissues of
all investigated Rickettsia-positive Deronectes species. Pre-
sumably, symbionts have been transported by the hemo-
lymph, comparable to observations in aphids (Chen et al.,
1996). This would support their appearance in the eyes and
legs. However, there is no evidence of harboring of Rickettsia
in specific sheath cells, secondary mycetocytes or mycetome-
like structures in aphids or booklice (Sakurai et al., 2005;
Perotti et al., 2006). Likewise, Rickettsia could not be
detected in high densities close to midgut epithelia or
malpighian tubules (Perotti et al., 2006). In contrast, the
internal genitals and fatbody exhibited numerous sym-
bionts. Rickettsiae depend on precursors of amino acids
from their host. Equally, the de novo synthesis of purines as
well as pyrimidine is not achieved by rickettsiae (Renesto
et al., 2005). Hence, tissue of high metabolic activity, such as
the fatbody, could be preferably infested with Rickettsia. The
presence of bacteria within ovariols and eggs also indicates
strong evidence for transovarial transmission, which must
contribute to the maintenance of Rickettsia sp. infection in
Deronectes populations (Whitworth et al., 2003). This is
supported by the existence of Rickettsia sp. in the second and
third larval stages of Deronectes, whereas the quantity of
symbionts increased gradually in each stage.
Thus, the amount of bacteria seems to be determined by
several factors: (1) fidelity of vertical transmission, (2)
frequency of horizontal transmission (Fukatsu et al., 2000),
(3) fitness effect on the host and (4) selfish manipulation of
the host reproduction (Tsuchida et al., 2002). Moreover,
interactions between the endosymbionts and their hosts are
complicated due to subpopulation structures and fluctuat-
ing environmental conditions. The resulting bottleneck
during vertical transmission could be a reason for the
fluctuated rate of rickettsial affection in D. aubei. More
individuals of D. delarouzei and D. semirufus have to be
investigated to verify the infection rates. Because of the
predatory nature of Deronectes (Dettner et al., 1986), a
horizontal transmission of Rickettsia sp. seems to be possi-
ble. Current research should demonstrate whether aquatic
prey of Deronectes is also associated with Rickettsia sp.
On the basis of the present results, there are no indica-
tions that Rickettsia sp. affection has an effect on the fitness
of Deronectes. Neither reduced body weight and fecundity
like in Rickettsia-infected host aphids (Sakurai et al., 2005)
nor remarkable increases in host size as observed in leeches
(Kikuchi & Fukatsu, 2005) could be observed. However, the
research of fitness effects is quite difficult, because Dero-
nectes cannot be bred under laboratory conditions.
Parasitic living bacteria, such as Rickettsia, Spiroplasma,
Cardinium and especially Wolbachia, have a tendency to
manipulate host reproduction for their own benefit. Repro-
ductive phenomena such as parthenogenesis, cytoplasmatic
incompatibility, feminization and male killing have been
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
209Rickettsia endosymbiont in water beetles

documented (O’Neill et al., 1997). Male killing caused by
Rickettsia was described until now in ladybird beetles A.
bipunctata and A. decempunctata (Werren et al., 1994; von
der Schulenburg et al., 2001) and buprestid beetle Brachys
tessellates (Lawson et al., 2001). Probably, rickettsiae also
influence oogenesis in bark beetle Coccotrypes dactyliperda
(Zchori-Fein et al., 2006). Rickettsia-induced parthenogenesis
was documented from eulophid parasitoid Neochrysocharis
formosa (Hagimori et al., 2006). In contrast, all investigated
species of Deronectes indicated a balanced sex ratio.
In this context, further research is required to investigate
in detail more about the influence of Rickettsia on Deronectes
species, especially at the biochemical level. Furthermore,
transmission and maintenance of Rickettsia in different
populations of Deronectes species must be examined. Future
studies will show, whether other Dytiscidae as well as their
aquatic prey are also infected by Rickettsia sp. symbiont.
Acknowledgements
We thank M. Meyering-Vos, G. Rambold and D. Persoh for
providing equipment and assistance in cloning, sequencing
and phylogenetic analysis. S. Geimer and R. Grotjahn are
gratefully acknowledged for assistance in electron micro-
scopy analysis, as well as S. Heidmann for the opportunity
to use the confocal microscope, and for providing help. We
also thank A. Kirpal for technical assistance.
References
Amann RI, Ludwig W & Schleifer KH (1995) Phylogenetic
identification and in situ detection of individual microbial cells
without cultivation. Microbiol Rev 59: 143–169.
Campbell CL, Mummey DL, Schmidtmann ET & Wilson WC
(2004) Culture-independent analysis of midgut microbiota in
the arbovirus vector Culicoides sonorensis (Diptera:
Ceratopogonidae). J Med Entomol 41: 340–348.
Chen DQ, Campbell BC & Purcell AH (1996) A new rickettsia
from a herbivorous insect, the pea aphid Acyrthosiphon pisum
(Harris). Curr Microbiol 33: 123–128.
Cilia V, Lafay B & Christen R (1996) Sequence heterogeneities
among 16S ribosomal RNA sequences, and their effect on
phylogenetic analyses at the species level. Mol Biol Evol 13:
451–461.
Daims H, Bruhl A, Amann R, Schleifer K & Wagner M (1999) The
domain-specific probe EUB338 is insufficient for the detection
of all Bacteria: development and evaluation of a more
comprehensive probe set. Syst Appl Microbiol 22: 434–444.
Davis MJ, Ying Z, Brunner BR, Pantoja A & Ferwerda FH (1998)
Rickettsial relative associated with papaya bunchy top disease.
Curr Microbiol 36: 80–84.
Dettner K, Hubner M & Classen R (1986) Age structure,
phenology and prey of some rheophilic Dytiscidae
(Coleoptera). Ent Basil 11: 343–370.
Duron O, Bouchon D, Boutin S, Bellamy L, Zhou L, Engelstadter
J & Hurst G (2008) The diversity of reproductive parasites
among arthropodes: Wolbachia do not walk alone. BMC Biol
6: 27.
Dykova I, Veverkova M, Fiala I, Machackova B & Peckova H
(2003) Nuclearia pattersoni sp. n. (Filosea), a new species of
amphizoic amoeba isolated from gills of roach (Rutilus
rutilus), and its rickettsial endosymbiont. Folia Parasit 50:
161–170.
Eremeeva ME & Dasch GA (2000) Rickettsiae. Encyclopedia of
Microbiology, Vol. 4 (Lederberg J, ed.), pp. 140–179. Academic
Press, New York.
Fery H & Brancucci M (1997) A taxonomic revision of Deronectes
(Sharp), 1882 (Insecta: Coleoptera: Dytiscidae) (partI). Ann
Naturhist Mus Wien 99B: 17–302.
Fery H, Koksal Erman O & Hosseinie S (2001) Two new
Deronectes (Sharp), 1882 (Insecta: Coleoptera: Dytiscidae) and
notes on other species of the genus. Ann Naturhist Mus Wien
103B: 341–351.
Fournier P & Raoult D (2007) Identification of rickettsial isolates
at the species level using multi-spacer typing. BMC Microbiol
7: 72.
Fournier PE, Roux V & Raoult D (1998) Phylogenetic analysis of
spotted fever group rickettsiae by study of the outer surface
protein rOmpA. Int J Syst Bacteriol 48: 839–849.
Fournier PE, Dumler JS, Greub G, Zhang J, Wu Y & Raoult D
(2003) Gene sequence-based criteria for identification of new
rickettsia isolates and description of Rickettsia heilongjiangensis
sp. nov. J Clin Microbiol 41: 5456–5465.
Fukatsu T, Nikoh N, Kawai R & Koga R (2000) The secondary
endosymbiotic bacterium of the pea aphid Acyrthosiphon
pisum (Insecta: Homoptera). Appl Environ Microbiol 66:
2748–2758.
Gonzalez J, Zimmermann J & Saiz-Jimenez C (2005) Evaluating
putative chimeric sequences from PCR-amplified products.
Bioinformatics 21: 333–337.
Goodacre SL, Martin OY, Thomas CF & Hewitt GM (2006)
Wolbachia and other endosymbiont infections in spiders. Mol
Ecol 15: 517–527.
Gottlieb Y, Ghanim M, Chiel E et al. (2006) Identification and
localization of a Rickettsia sp. in Bemisia tabaci (Homoptera:
Aleyrodidae). Appl Environ Microbiol 72: 3646–3652.
Hagimori T, Abe Y, Date S & Miura K (2006) The first finding of a
Rickettsia bacterium associated with parthenogenesis
induction among insects. Curr Microbiol 52: 97–101.
Hall TA (1999) BioEdit. A user-friendly biological sequence
alignment editor analysis program for Windows 95/98/NT.
Nucl Acids Symp Ser 41: 95–98.
Hoy MA & Jeyaprakash A (2005) Microbial diversity in the
predatory mite Metaseiulus occidentalis (Acari: Phytoseiidae)
and its prey, Tetranychus urticae (Acari: Tetranychidae). Biol
Control 32: 427–441.
Kikuchi Y & Fukatsu T (2005) Rickettsia infection in natural leech
populations. Microb Ecol 49: 265–271.
FEMS Microbiol Ecol 68 (2009) 201–211c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
210 S.M. Kuchler et al.

Kikuchi Y, Sameshima S, Kitade O, Kojima J & Fukatsu T (2002)
Novel clade of Rickettsia spp. from leeches. Appl Environ
Microbiol 68: 999–1004.
Labruna MB, Whitworth T, Horta MC, Bouyer DH, McBride JW,
Pinter A, Popov V, Gennari SM & Walker DH (2004) Rickettsia
species infecting Amblyomma cooperi ticks from an area in the
state of Sao Paulo, Brazil, where Brazilian spotted fever is
endemic. J Clin Microbiol 42: 90–98.
Lane DJ (1991) 16S and 23S rRNA sequencing. Nucleic Acid
Techniques in Bacterial Systematics (Stackenbrandt E &
Goodfellow M, eds), pp. 115–148. John Wiley and Sons,
New York.
Lawson ET, Mousseau TA, Klaper R, Hunter MD & Werren JH
(2001) Rickettsia associated with male-killing in a buprestid
beetle. Heredity 86: 497–505.
Ludwig W, Strunk O, Westram R et al. (2004) ARB: a software
environment for sequence data. Nucleic Acids Res 32:
1363–1371.
Maddison D, Baker M & Ober K (1999) Phylogeny of Carabid
beetles as inferred from 18S ribosomal DNA (Coleoptera:
Carabidae). Syst Entomol 24: 1–36.
Manz W, Amann R, Ludwig W, Wagner M & Schleifer K-H
(1992) Phylogenetic oligodeoxynucleotide probes for the
major subclasses of proteobacteria: problems and solutions.
Syst Appl Microbiol 15: 593–600.
Minnick MF & Anderson BE (2006) The genus Bartonella.
Prokaryotes, Vol. 5 (Dworkin M, Falkow S, Rosenberg E,
Schleifer K-H & Stackebrandt E, eds), pp. 467–492. Springer,
New York.
Ngwamidiba M, Blanc G, Raoult D & Fournier PE (2006) Sca1, a
previously undescribed paralog from autotransporter protein-
encoding genes in Rickettsia species. BMC Microbiol 6: 12.
O’Neill SL, Hoffmann AA & Werren JH (1997) Influential
Passengers: Inherited Microorganisms and Arthropod
Reproduction. Oxford University Press, Oxford.
Perlman SJ, Hunter MS & Zchori-Fein E (2006) The emerging
diversity of Rickettsia. Proc Biol Sci 273: 2097–2106.
Perotti MA, Clarke HK, Turner BD & Braig HR (2006) Rickettsia
as obligate and mycetomic bacteria. FASEB J 20: 2372–2374.
Renesto P, Ogata H, Audic S, Claverie JM & Raoult D (2005)
Some lessons from Rickettsia genomics. FEMS Microbiol Rev
29: 99–117.
Roux V & Raoult D (2000) Phylogenetic analysis of members of
the genus Rickettsia using the gene encoding the outer-
membrane protein rOmpB (ompB). Int J Syst Evol Microbiol
50: 1449–1455.
Roux V, Rydkina E, Eremeeva M & Raoult D (1997) Citrate
synthase gene comparison, a new tool for phylogenetic
analysis, and its application for the rickettsiae. Int J Syst
Bacteriol 47: 252–261.
Sakurai M, Koga R, Tsuchida T, Meng XY & Fukatsu T (2005)
Rickettsia symbiont in the pea aphid Acyrthosiphon pisum:
novel cellular tropism, effect on host fitness, and interaction
with the essential symbiont Buchnera. Appl Environ Microbiol
71: 4069–4075.
Schwoerbel J (1994) Methoden der Hydrobiologie,
Sußwasserbiologie. Gustav Fischer, Stuttgart.
Sekeyova Z, Roux V & Raoult D (2001) Phylogeny of Rickettsia
spp. inferred by comparing sequences of ‘gene D’, which
encodes an intracytoplasmic protein. Int J Syst Evol Microbiol
51: 1353–1360.
Shull VL, Vogler AP, Baker MD, Maddison DR & Hammond PM
(2001) Sequence alignment of 18S ribosomal RNA and the
basal relationships of Adephagan beetles: evidence for
monophyly of aquatic families and the placement of
Trachypachidae. Syst Biol 50: 945–969.
Silverman DJ (1991) Some contributions of electron microscopy
to the study of rickettsiae. Eur J Epidemiol 7: 200–206.
Stothard DR & Fuerst PA (1995) Evolutionary analysis of the
spotted fever and typhus groups of Rickettsia using 16 rRNA
gene sequences. Syst Appl Microbiol 18: 52–61.
Swofford DL (2000) PAUP�. Phylogenetic Analysis Using
Parsimony (�and Other Methods). Sinauer Associates,
Sunderland, MA.
Tsuchida T, Koga R, Shibao H, Matsumoto T & Fukatsu T (2002)
Diversity and geographic distribution of secondary
endosymbiotic bacteria in natural populations of the pea
aphid, Acyrthosiphon pisum. Mol Ecol 11: 2123–2135.
Vannini C, Petroni G, Verni F & Rosati G (2005) A bacterium
belonging to the Rickettsiaceae family inhabits the cytoplasm
of the marine ciliate Diophrys appendiculata (Ciliophora,
Hypotrichia). Microb Ecol 49: 434–442.
von der Schulenburg JH, Habig M, Sloggett JJ, Webberley KM,
Bertrand D, Hurst GD & Majerus ME (2001) Incidence of
male-killing Rickettsia spp. (alpha-proteobacteria) in the ten-
spot ladybird beetle Adalia decempunctata L. (Coleoptera:
Coccinellidae). Appl Environ Microbiol 67: 270–277.
Weiss E & Moulder JW (1984) Order I. Rickettsiales
Gieszczkiewicz 1939, 25AL. Bergey’s Manual of Systematic
Bacteriology, Vol. 1 (Krieg NR & Holt JG, eds), pp. 687–701.
Williams and Wilkins, Baltimore, MD.
Werren JH, Hurst GD, Zhang W, Breeuwer JA, Stouthamer R &
Majerus ME (1994) Rickettsial relative associated with male
killing in the ladybird beetle (Adalia bipunctata). J Bacteriol
176: 388–394.
Whitworth T, Popov V, Han V, Bouyer D, Stenos J, Graves S, Ndip
L & Walker D (2003) Ultrastructural and genetic evidence of a
reptilian tick, Aponomma hydrosauri, as a host of Rickettsia
honei in Australia: possible transovarial transmission. Ann NY
Acad Sci 990: 67–74.
Yu X-J & Walker DH (2006) The order Rickettsiales.
Prokaryotes, Vol. 5 (Dworkin M, Falkow S, Rosenberg E,
Schleifer K-H & Stackebrandt E, eds), pp. 493–528. Springer,
New York.
Zchori-Fein E, Borad C & Harari A (2006) Oogenesis in the date
stone beetle, Coccotrypes dactyliperda, depends on symbiotic
bacteria. Phy Entomol 31: 164–169.
FEMS Microbiol Ecol 68 (2009) 201–211 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
211Rickettsia endosymbiont in water beetles