research article gardenia jasminoides extract attenuates the...
TRANSCRIPT

Research ArticleGardenia jasminoides Extract Attenuates the UVB-InducedExpressions of Cytokines in Keratinocytesand Indirectly Inhibits Matrix Metalloproteinase-1 Expressionin Human Dermal Fibroblasts
Jiaa Park1 Jin Kyung Seok1 Hwa-Jin Suh2 and Yong Chool Boo13
1 Department of Molecular Medicine Cell and Matrix Research Institute BK21 Plus KNU Biomedical Convergence ProgramKyungpook National University School of Medicine Daegu 700-422 Republic of Korea
2 Gyeongbuk Natural Color Industry Institute Gyeongbuk Republic of Korea3 Ruby Crown Co Ltd Daegu Republic of Korea
Correspondence should be addressed to Yong Chool Boo ycbooknuackr
Received 4 November 2013 Revised 28 January 2014 Accepted 3 February 2014 Published 11 March 2014
Academic Editor Ki-Wan Oh
Copyright copy 2014 Jiaa Park et alThis is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Ultraviolet radiation (UV) is a major cause of photoaging which also involves inflammatory cytokines and matrix metallopro-teinases (MMP) The present study was undertaken to examine the UVB-protecting effects of yellow-colored plant extracts incell-based assays HaCaT keratinocytes were exposed to UVB in the absence or presence of plant extracts and resulting changesin cell viability and inflammatory cytokine expression were measured Of the plant extracts tested Gardenia jasminoides extractshowed the lowest cytotoxicity and dose-dependently enhanced the viabilities of UVB-exposed cells Gardenia jasminoides extractalso attenuated the mRNA expressions of interleukin-1120573 (IL-1120573) and tumor necrosis factor-120572 (TNF-120572) in HaCaT cells stimulatedby UVB Conditioned medium from UVB-exposed HaCaT cells was observed to stimulate MMP-1 protein expression in humandermal fibroblasts and this effect was much smaller for the conditioned medium of HaCaT cells exposed to UVB in the presenceof Gardenia jasminoides extract Gardenia jasminoides extract also exhibited antioxidative and antiapoptotic effects in HaCaT cellsexposed to UVB These results indicated that UVB-induced injury and inflammatory responses of skin cells can be attenuated byyellow-colored plant extracts such as Gardenia jasminoides extract
1 Introduction
Cosmetics have become essential products as people pursueesthetic desires in modern society and their role has beenextended to the retardation of skin aging caused by physio-logical and environmental factors Ultraviolet radiation (UV)is a major cause of skin aging which is characterized bywrinkles laxity blister formation roughness and loss of skintone [1ndash3] Therefore there is considerable interest in thedevelopment of safer more effective ingredients that mitigatethe harmful effects of UV
Photoaging involves changes in dermal extracellular ma-trix composition and collagen loss Matrix metalloproteinas-es (MMPs) a group of zinc endopeptidases play a key role
in the turnover of extracellular matrix macromoleculesincluding type I collagen [4 5] Furthermore the expressionsof MMPs such as MMP-1 -2 -3 and -9 are known tobe upregulated in UV-exposed human dermal fibroblasts[6 7]TheseMMPs can regulate the remodeling of connectivetissues associated with the formation of wrinkles and otherphenotypes of photoagingThus the pharmacological target-ing ofMMPs is considered a promising strategy to reduce thephotoaging process [8]
The UVB-induced upregulation of MMPs is mainlymediated by signal transduction pathways involving the UV-induced activation of cytokine receptors and the subse-quent activations of mitogen-activated protein kinases suchas extracellular signal-regulated kinase c-Jun-N-terminal
Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2014 Article ID 429246 10 pageshttpdxdoiorg1011552014429246
2 Evidence-Based Complementary and Alternative Medicine
Wavelength (nm)
Gardenia jasminoides
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(a)
Phellodendron amurense
Wavelength (nm)
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(b)
Rheum rhabarbarum
Wavelength (nm)
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(c)
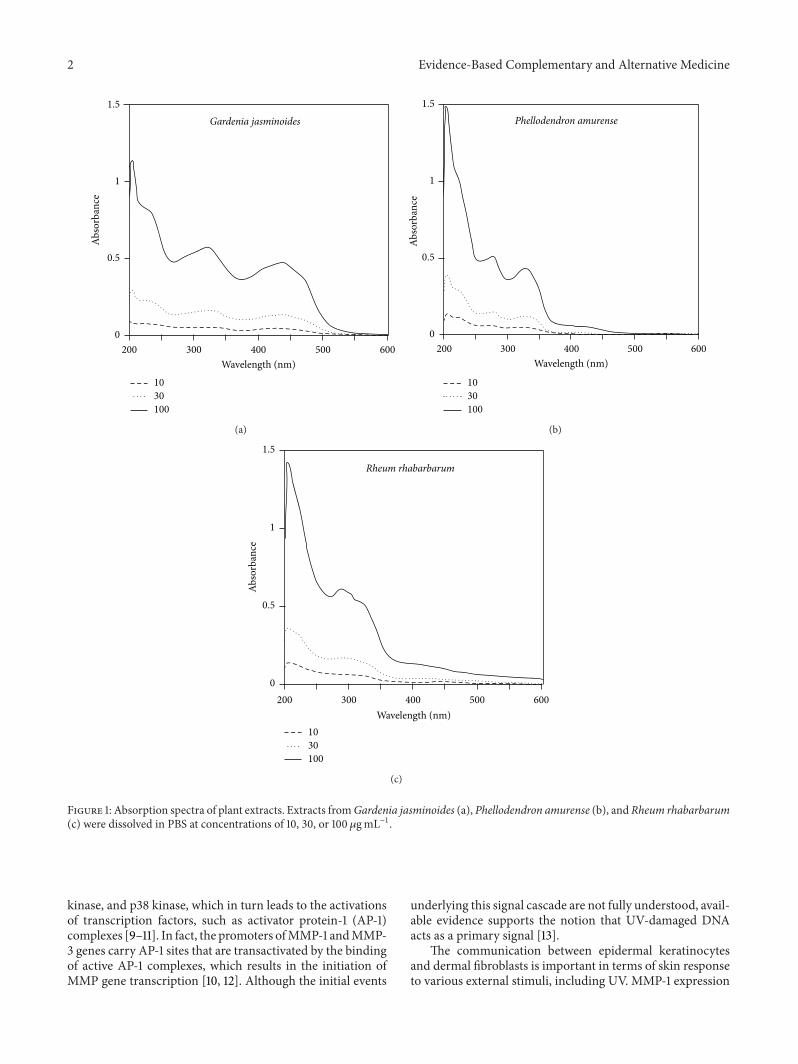
Figure 1 Absorption spectra of plant extracts Extracts fromGardenia jasminoides (a) Phellodendron amurense (b) and Rheum rhabarbarum(c) were dissolved in PBS at concentrations of 10 30 or 100 120583gmLminus1
kinase and p38 kinase which in turn leads to the activationsof transcription factors such as activator protein-1 (AP-1)complexes [9ndash11] In fact the promoters ofMMP-1 andMMP-3 genes carry AP-1 sites that are transactivated by the bindingof active AP-1 complexes which results in the initiation ofMMP gene transcription [10 12] Although the initial events
underlying this signal cascade are not fully understood avail-able evidence supports the notion that UV-damaged DNAacts as a primary signal [13]
The communication between epidermal keratinocytesand dermal fibroblasts is important in terms of skin responseto various external stimuli including UV MMP-1 expression
Evidence-Based Complementary and Alternative Medicine 3
Vehicle10 30 100 10 30 100 10 30 100
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
0255075
100125
Cel
l via
bilit
y(
of c
ontro
l)
Figure 2 Effects of plant extracts derived from Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarum on theviability of HaCaT keratinocytes Cells were treated with plantextracts at the indicated concentration for 24 h and cell viabilitieswere determined using an MTT assay Cell viabilities are presentedas percentages of the vehicle control (means plusmn SEs 119899 = 3) 119875 lt 005versus the vehicle control
in dermal fibroblasts can be stimulated by direct expo-sure of the fibroblasts to UV In addition MMP-1 expres-sion in dermal fibroblasts can be stimulated indirectly bythe cytokines or other factors secreted by epidermal ker-atinocytes exposed to UV Previous studies have shown thatthe conditioned medium from UV irradiated keratinocyteseffectively increases MMP-1 expression in fibroblasts viaparacrine effects [14 15] Thus MMP-1 expression in dermalfibroblasts may be regulated indirectly by inhibiting theactivation of epidermal keratinocytes in response to UV Inthis regard plant extracts with UV-shielding antioxidativeand anti-inflammatory properties are an attractive prospect[16 17]
The purpose of the present study was to examine theprotective effects of plant extracts derived fromGardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumagainst skin cell responses to UVThese yellow plant extractswere chosen because of their ability to absorb UV effectively
2 Materials and Methods
21 Plant Materials Extracts of Gardenia jasminoides Ellis(seed) Phellodendron amurense Rupr (root) and Rheumrhabarbarum L (root) were prepared at Gyeongbuk NaturalColor Industry Institute (httpwwwgncrekr) Voucherspecimens of the plants and other information regarding theextracts are available at this institute Plant materials were ex-tracted with hot water 80∘C for 2 h and extracts were evap-orated to dryness under reduced pressure
22 UV Spectrophotometry Test materials were dissolved inphosphate buffered saline (PBS) to final concentrations of10 30 and 100 120583gmLminus1 Absorption spectra were recordedin the 200sim600 nm range using a Shimadzu UV-1650PCspectrophotometer (Shimadzu Corporation Kyoto Japan)
23 HPLC Analysis HPLC analysis of extracts was doneusing a Gilson HPLC system (Gilson Inc Middleton WIUSA) equipped with a 321 pump and UVVIS 151 detectorAqueous solution of test materials (10mgmLminus1) was injected
at 10 120583L Separation was done on a 5 120583m Hector-M C18 col-umn (46mm times 250mm) (RS Tech Co Daejeon Korea)The mobile phase consisted of 05 formic acid (A) and ac-etonitrile (B) The gradient was programmed as follows 0ndash20min a linear gradient from20 to 80B 20ndash35min 80BThe flow rate was 05mLminminus1 The UV detector was setat 280 nm and 440 nm Purified crocin was purchased fromSigma-Aldrich (St Louis MO USA) and its purity was deter-mined spectrophotometrically using the extinction coeffi-cient 120576
443
= 89000Mminus1 cmminus1 [18]
24 HaCaT Cell Culture HaCaT cells (a human immortal-ized keratinocyte cell-line) were grown in DMEMF-12 me-dium (GIBCO-BRL Grand Island NY USA) supplementedwith a 10 fetal bovine serum 100UmLminus1 penicillin01mgmLminus1 streptomycin 025 120583gmLminus1 amphotericin B and10 120583gmLminus1 hydrocortisone Cells were cultured at 37∘C in ahumidified atmosphere containing 5 CO
2
and 95 air
25 UVB-Exposure of HaCaT Cells HaCaT cells were seededon a six-well plate at a density of 2 times 105 cells per well andgrown in a culture medium for 48 h to reach 80 confluencyThe cells were then washed twice with PBS and exposed toUVB in PBS containing the test material UVB treatmentwas conducted under a cell culture hood using a UVB lamp(Model UVB-18 ULTRAlowastLUM Inc Claremont CA USA)that emitted radiation in the wavelength range of 280 to340 nm with maximum intensity at 300 nm The intensity ofradiation was determined using a UV light meter (ModelUV 340A Lutron Electronic Enterprise Co Taipei Taiwan)and the UV was administered to cells in culture plates atan intensity of 80 120583Wcmminus2 However durations of treatmentvaried to provide specific doses of UVB (5 10 or 15mJ cmminus2)Following irradiation PBS was replaced by growth mediumand cells were incubated for 1 day Cells were subjected tovarious tests as detailed below Culture conditioned mediumwas harvested and used to treat human dermal fibroblasts
26 Cell Viability Test Cell viability was assayed using 3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazolium bromide(MTT) This assay method is based on the ability of mito-chondrial dehydrogenase in viable cells to reduce pale yellowMTT to dark blue formazan crystals that accumulate withincells Briefly cells were washed with PBS and incubated in1mL of culturemedia supplementedwith 1mgmLminus1MTT for3 h The medium was then discarded and cells were treatedwith isopropanol to solubilize the formazan Solutions werethen transferred to microplates and formazan was quantifiedby measuring absorbance at 595 nm
27 Quantitative Polymerase Chain Reaction (qRT-PCR)Anal-ysis Total cellular RNA was extracted using the RNeasy kit(Qiagen Valencia CA USA) To prepare cDNA one 120583g ofcellular mRNA was reverse transcribed using the High Ca-pacity cDNA Archive Kit (Applied Biosystems Foster CityCA USA) This kit utilizes random hexamers primers andMultiScribe Reverse Transcriptase PCRwas conducted using
4 Evidence-Based Complementary and Alternative Medicine
0 5 10 15
0
25
50
75
100
125
Cel
l via
bilit
y (
of c
ontro
l)
UVB (mJ cmminus2)
(a)
0255075
100125
Cel
l via
bilit
y (
of c
ontro
l)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
UVB (10mJ cmminus2)
lowast lowast
lowast
lowast lowast
lowast
(b)
Figure 3 Effects of plant extracts on UVB cytotoxicity in HaCaT cells In (a) HaCaT cells were irradiated with 5sim15mJ cmminus2 UVB In (b)cells were irradiated with 10mJ cmminus2 UVB in PBS in the absence or presence of plant extracts (10 30 or 100120583gmLminus1) Irradiated cells weresubsequently incubated for 24 h and cell viabilities were determined using an MTT assay Cell viabilities are presented as percentages of thevehicle control (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
0 5 10 15
0
5
10
15
20
UVB (mJ cmminus2)
IL-1120573
mRN
A (f
old)
(a)
0 5 10 15
0
5
10
15
20
TNF-120572
mRN
A (f
old)
UVB (mJ cmminus2)
(b)
05
101520
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
IL-1120573
mRN
A(fo
ld) lowast
lowast
lowast
UVB (10mJ cmminus2)
(c)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
02468
10
TNF-120572
mRN
A(fo
ld)
lowast lowast lowast
UVB (10mJ cmminus2)
(d)
Figure 4 Effects of plant extracts on mRNA expressions of inflammatory cytokines induced by UVB in HaCaT cells In (a) and (b) HaCaTcells were irradiated with 5sim15mJ cmminus2 UVB In (c) and (d) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence of plantextracts (10 30 or 100120583gmLminus1) and then incubated for 24 h before mRNA analysis by qRT-PCR GAPDH was used as the reference Dataare expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiatedwith UVB
the StepOnePlus Real-Time PCR System (Applied Biosys-tems) in reaction mixtures (20120583L) containing SYBR GreenPCRMasterMix (Applied Biosystems) 60 ng of cDNA and 2picomole of gene-specific primer sets (Macrogen Seoul)Theprimers used for PCR analysis were as follows interleukin-1120573 (IL-1120573) (GeneBank accession number NM 0005762)
51015840-CCT GTC CTG CGT GTT GAA AGA-31015840 (forward)and 51015840-GGG AAC TGG GCA GAC TCA AA-31015840 (reverse)tumor necrosis factor-120572 (TNF-120572) (NM 0005943) 51015840-TGCTCC TCA CCC ACA CCA T-31015840 (forward) and 51015840-GAGATA GTC GGG CCG ATT GA-31015840 (reverse) and GAPDH(NM 0020463) 51015840-ATG GGG AAG GTG AAG GTC G-31015840
Evidence-Based Complementary and Alternative Medicine 5
MMP-1
0
5
10
15
MM
P-1
prot
ein
(fold
)
0 5 10 15
UVB (mJ cmminus2)
120573-Actin
(a)
0
1
2
3
4
MM
P-1
prot
ein
(fold
)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV UVB (10mJ cmminus2)
MMP-1120573-Actin
lowastlowast
lowast
(b)
Figure 5 Expression levels of MMP-1 protein in human dermal fibroblasts stimulated by the conditioned media of HaCaT cells irradiatedwith UVB in the absence or presence of plant extracts In (a) HaCaT cells were irradiated with UVB at 5sim15mJ cmminus2 and then incubatedfor 24 h The conditioned medium from the UVB-irradiated HaCaT cells was used to treat human dermal fibroblasts for 24 h In (b) HaCaTcells were irradiated with UVB at 10mJ cmminus2 in PBS containing plant extracts at various concentrations 120583gmLminus1 and then incubated for 24 hConditionedmedia from these UVB-irradiated HaCaT cells were used to treat human dermal fibroblasts for 24 hMMP-1 protein expressionsin fibroblasts were analyzed by Western blot using 120573-actin as a loading control Typical blot images are shown Data are expressed as foldchanges (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
(forward) and 51015840-GGG GTC ATT GAT GGC AAC AA-31015840(reverse) Reactions were performed using the following con-ditions 50∘C for 2min 95∘C for 10min and 40 amplificationcycles (95∘C for 15 s and 60∘C for 1min) followed by adissociation stepMelting curve analysis showed single peakswhich supported the homogeneity of amplicons The mRNAexpression levels of IL-1120573 and TNF-120572 relative to that ofinternal control glyceraldehyde 3-phosphate dehydrogenase(GAPDH) were calculated using the comparative thresholdcycle (CT) method
28 Culture of HumanDermal Fibroblasts Human dermal fi-broblasts isolated from adult skin were obtained from Cas-cade Biologics (Portland OR USA) The cells were culturedat 37∘C in a humidified atmosphere containing 5 CO
2
and95 air in growth medium (Iscoversquos Modified Dulbeccorsquosmedium (GIBCO-BRL Grand Island NY USA) containing10 fetal bovine serum 100UmLminus1 penicillin 01mgmLminus1streptomycin and 025 120583gmLminus1 amphotericin B) In exper-iments on the paracrine effects of factors secreted by kerat-inocytes the medium was completely replaced by the condi-tioned medium of UVB-irradiated HaCaT cells and fibrob-lasts were cultivated for 24 h before analysis
29 Western Blotting Whole cell lysates were prepared usinga lysis buffer (10mM Tris-Cl pH 74 120mM NaCl 25mMKCl 2mM EGTA 1mM EDTA 05 Triton X-100 and pro-tease inhibitor cocktail) Aliquots of lysates were subjectedto sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) under protein denaturing conditions Proteinsseparated were transferred to polyvinylidene fluoride mem-branes which were incubated with an appropriate primaryantibody overnight at 4∘C and then with a secondary anti-body conjugated to horseradish peroxidase for 1 h at room
temperature Immunoreactive bands were detected usinga picoEPD Western Reagent kit (ELPIS-Biotech DaejeonKorea) and subjected to densitometric analysis The primaryantibody for MMP-1 was purchased from Calbiochem (SanDiego CA USA) Rabbit polyclonal caspase-3 antibodyand rabbit polyclonal caspase-9 were purchased from CellSignaling (Danvers MA USA) The mouse monoclonal 120573-actin antibody was from Sigma-Aldrich (St Louis MOUSA)
210 Analysis of Lipid Peroxidation As a marker of lipid per-oxidation 2-thiobarbituric acid-reactive substances (TBARS)were quantified [19] Briefly cells were treated in a lysisbuffer (20mM Tris-Cl 25mM EDTA 10 SDS pH 75)Cell lysates (200mg protein in 100 120583L) were mixed with900 120583L of 10 phosphoric acid and 10mL of 09 2-thiobarbituric acid (Sigma-Aldrich) and then heated on aboiling water bath for 45min Standard solutions of 1133-tetramethoxypropane (Sigma-Aldrich) a precursor of mal-ondialdehyde were treated in the same way as cell lysatesAfter cooling 15mL of 1-butanol was added and the mixturewas centrifuged at 13000 rpm for 15min to separate into twolayers The fluorescence intensity of the 1-butanol layer wasmeasured at an emission wavelength of 590 nm (excitationat 540 nm) using the Gemini EM fluorescence microplatereader (Molecular Devices Sunnyvale CA USA)
211 Statistical Analysis Data are presented as the means plusmnSEs of three or more independent experiments The signif-icance of differences between groups was determined usingthe Studentrsquos 119905-test Duncanrsquos multiple-range test was appliedwhen differences were significant (119875 lt 005) by the Studentrsquos119905-test Statistical significance was accepted for119875 valueslt 005
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
2 Evidence-Based Complementary and Alternative Medicine
Wavelength (nm)
Gardenia jasminoides
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(a)
Phellodendron amurense
Wavelength (nm)
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(b)
Rheum rhabarbarum
Wavelength (nm)
0
05
1
15
200 300 400 500 600
Abso
rban
ce
1030100
(c)
Figure 1 Absorption spectra of plant extracts Extracts fromGardenia jasminoides (a) Phellodendron amurense (b) and Rheum rhabarbarum(c) were dissolved in PBS at concentrations of 10 30 or 100 120583gmLminus1
kinase and p38 kinase which in turn leads to the activationsof transcription factors such as activator protein-1 (AP-1)complexes [9ndash11] In fact the promoters ofMMP-1 andMMP-3 genes carry AP-1 sites that are transactivated by the bindingof active AP-1 complexes which results in the initiation ofMMP gene transcription [10 12] Although the initial events
underlying this signal cascade are not fully understood avail-able evidence supports the notion that UV-damaged DNAacts as a primary signal [13]
The communication between epidermal keratinocytesand dermal fibroblasts is important in terms of skin responseto various external stimuli including UV MMP-1 expression
Evidence-Based Complementary and Alternative Medicine 3
Vehicle10 30 100 10 30 100 10 30 100
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
0255075
100125
Cel
l via
bilit
y(
of c
ontro
l)
Figure 2 Effects of plant extracts derived from Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarum on theviability of HaCaT keratinocytes Cells were treated with plantextracts at the indicated concentration for 24 h and cell viabilitieswere determined using an MTT assay Cell viabilities are presentedas percentages of the vehicle control (means plusmn SEs 119899 = 3) 119875 lt 005versus the vehicle control
in dermal fibroblasts can be stimulated by direct expo-sure of the fibroblasts to UV In addition MMP-1 expres-sion in dermal fibroblasts can be stimulated indirectly bythe cytokines or other factors secreted by epidermal ker-atinocytes exposed to UV Previous studies have shown thatthe conditioned medium from UV irradiated keratinocyteseffectively increases MMP-1 expression in fibroblasts viaparacrine effects [14 15] Thus MMP-1 expression in dermalfibroblasts may be regulated indirectly by inhibiting theactivation of epidermal keratinocytes in response to UV Inthis regard plant extracts with UV-shielding antioxidativeand anti-inflammatory properties are an attractive prospect[16 17]
The purpose of the present study was to examine theprotective effects of plant extracts derived fromGardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumagainst skin cell responses to UVThese yellow plant extractswere chosen because of their ability to absorb UV effectively
2 Materials and Methods
21 Plant Materials Extracts of Gardenia jasminoides Ellis(seed) Phellodendron amurense Rupr (root) and Rheumrhabarbarum L (root) were prepared at Gyeongbuk NaturalColor Industry Institute (httpwwwgncrekr) Voucherspecimens of the plants and other information regarding theextracts are available at this institute Plant materials were ex-tracted with hot water 80∘C for 2 h and extracts were evap-orated to dryness under reduced pressure
22 UV Spectrophotometry Test materials were dissolved inphosphate buffered saline (PBS) to final concentrations of10 30 and 100 120583gmLminus1 Absorption spectra were recordedin the 200sim600 nm range using a Shimadzu UV-1650PCspectrophotometer (Shimadzu Corporation Kyoto Japan)
23 HPLC Analysis HPLC analysis of extracts was doneusing a Gilson HPLC system (Gilson Inc Middleton WIUSA) equipped with a 321 pump and UVVIS 151 detectorAqueous solution of test materials (10mgmLminus1) was injected
at 10 120583L Separation was done on a 5 120583m Hector-M C18 col-umn (46mm times 250mm) (RS Tech Co Daejeon Korea)The mobile phase consisted of 05 formic acid (A) and ac-etonitrile (B) The gradient was programmed as follows 0ndash20min a linear gradient from20 to 80B 20ndash35min 80BThe flow rate was 05mLminminus1 The UV detector was setat 280 nm and 440 nm Purified crocin was purchased fromSigma-Aldrich (St Louis MO USA) and its purity was deter-mined spectrophotometrically using the extinction coeffi-cient 120576
443
= 89000Mminus1 cmminus1 [18]
24 HaCaT Cell Culture HaCaT cells (a human immortal-ized keratinocyte cell-line) were grown in DMEMF-12 me-dium (GIBCO-BRL Grand Island NY USA) supplementedwith a 10 fetal bovine serum 100UmLminus1 penicillin01mgmLminus1 streptomycin 025 120583gmLminus1 amphotericin B and10 120583gmLminus1 hydrocortisone Cells were cultured at 37∘C in ahumidified atmosphere containing 5 CO
2
and 95 air
25 UVB-Exposure of HaCaT Cells HaCaT cells were seededon a six-well plate at a density of 2 times 105 cells per well andgrown in a culture medium for 48 h to reach 80 confluencyThe cells were then washed twice with PBS and exposed toUVB in PBS containing the test material UVB treatmentwas conducted under a cell culture hood using a UVB lamp(Model UVB-18 ULTRAlowastLUM Inc Claremont CA USA)that emitted radiation in the wavelength range of 280 to340 nm with maximum intensity at 300 nm The intensity ofradiation was determined using a UV light meter (ModelUV 340A Lutron Electronic Enterprise Co Taipei Taiwan)and the UV was administered to cells in culture plates atan intensity of 80 120583Wcmminus2 However durations of treatmentvaried to provide specific doses of UVB (5 10 or 15mJ cmminus2)Following irradiation PBS was replaced by growth mediumand cells were incubated for 1 day Cells were subjected tovarious tests as detailed below Culture conditioned mediumwas harvested and used to treat human dermal fibroblasts
26 Cell Viability Test Cell viability was assayed using 3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazolium bromide(MTT) This assay method is based on the ability of mito-chondrial dehydrogenase in viable cells to reduce pale yellowMTT to dark blue formazan crystals that accumulate withincells Briefly cells were washed with PBS and incubated in1mL of culturemedia supplementedwith 1mgmLminus1MTT for3 h The medium was then discarded and cells were treatedwith isopropanol to solubilize the formazan Solutions werethen transferred to microplates and formazan was quantifiedby measuring absorbance at 595 nm
27 Quantitative Polymerase Chain Reaction (qRT-PCR)Anal-ysis Total cellular RNA was extracted using the RNeasy kit(Qiagen Valencia CA USA) To prepare cDNA one 120583g ofcellular mRNA was reverse transcribed using the High Ca-pacity cDNA Archive Kit (Applied Biosystems Foster CityCA USA) This kit utilizes random hexamers primers andMultiScribe Reverse Transcriptase PCRwas conducted using
4 Evidence-Based Complementary and Alternative Medicine
0 5 10 15
0
25
50
75
100
125
Cel
l via
bilit
y (
of c
ontro
l)
UVB (mJ cmminus2)
(a)
0255075
100125
Cel
l via
bilit
y (
of c
ontro
l)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
UVB (10mJ cmminus2)
lowast lowast
lowast
lowast lowast
lowast
(b)
Figure 3 Effects of plant extracts on UVB cytotoxicity in HaCaT cells In (a) HaCaT cells were irradiated with 5sim15mJ cmminus2 UVB In (b)cells were irradiated with 10mJ cmminus2 UVB in PBS in the absence or presence of plant extracts (10 30 or 100120583gmLminus1) Irradiated cells weresubsequently incubated for 24 h and cell viabilities were determined using an MTT assay Cell viabilities are presented as percentages of thevehicle control (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
0 5 10 15
0
5
10
15
20
UVB (mJ cmminus2)
IL-1120573
mRN
A (f
old)
(a)
0 5 10 15
0
5
10
15
20
TNF-120572
mRN
A (f
old)
UVB (mJ cmminus2)
(b)
05
101520
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
IL-1120573
mRN
A(fo
ld) lowast
lowast
lowast
UVB (10mJ cmminus2)
(c)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
02468
10
TNF-120572
mRN
A(fo
ld)
lowast lowast lowast
UVB (10mJ cmminus2)
(d)
Figure 4 Effects of plant extracts on mRNA expressions of inflammatory cytokines induced by UVB in HaCaT cells In (a) and (b) HaCaTcells were irradiated with 5sim15mJ cmminus2 UVB In (c) and (d) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence of plantextracts (10 30 or 100120583gmLminus1) and then incubated for 24 h before mRNA analysis by qRT-PCR GAPDH was used as the reference Dataare expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiatedwith UVB
the StepOnePlus Real-Time PCR System (Applied Biosys-tems) in reaction mixtures (20120583L) containing SYBR GreenPCRMasterMix (Applied Biosystems) 60 ng of cDNA and 2picomole of gene-specific primer sets (Macrogen Seoul)Theprimers used for PCR analysis were as follows interleukin-1120573 (IL-1120573) (GeneBank accession number NM 0005762)
51015840-CCT GTC CTG CGT GTT GAA AGA-31015840 (forward)and 51015840-GGG AAC TGG GCA GAC TCA AA-31015840 (reverse)tumor necrosis factor-120572 (TNF-120572) (NM 0005943) 51015840-TGCTCC TCA CCC ACA CCA T-31015840 (forward) and 51015840-GAGATA GTC GGG CCG ATT GA-31015840 (reverse) and GAPDH(NM 0020463) 51015840-ATG GGG AAG GTG AAG GTC G-31015840
Evidence-Based Complementary and Alternative Medicine 5
MMP-1
0
5
10
15
MM
P-1
prot
ein
(fold
)
0 5 10 15
UVB (mJ cmminus2)
120573-Actin
(a)
0
1
2
3
4
MM
P-1
prot
ein
(fold
)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV UVB (10mJ cmminus2)
MMP-1120573-Actin
lowastlowast
lowast
(b)
Figure 5 Expression levels of MMP-1 protein in human dermal fibroblasts stimulated by the conditioned media of HaCaT cells irradiatedwith UVB in the absence or presence of plant extracts In (a) HaCaT cells were irradiated with UVB at 5sim15mJ cmminus2 and then incubatedfor 24 h The conditioned medium from the UVB-irradiated HaCaT cells was used to treat human dermal fibroblasts for 24 h In (b) HaCaTcells were irradiated with UVB at 10mJ cmminus2 in PBS containing plant extracts at various concentrations 120583gmLminus1 and then incubated for 24 hConditionedmedia from these UVB-irradiated HaCaT cells were used to treat human dermal fibroblasts for 24 hMMP-1 protein expressionsin fibroblasts were analyzed by Western blot using 120573-actin as a loading control Typical blot images are shown Data are expressed as foldchanges (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
(forward) and 51015840-GGG GTC ATT GAT GGC AAC AA-31015840(reverse) Reactions were performed using the following con-ditions 50∘C for 2min 95∘C for 10min and 40 amplificationcycles (95∘C for 15 s and 60∘C for 1min) followed by adissociation stepMelting curve analysis showed single peakswhich supported the homogeneity of amplicons The mRNAexpression levels of IL-1120573 and TNF-120572 relative to that ofinternal control glyceraldehyde 3-phosphate dehydrogenase(GAPDH) were calculated using the comparative thresholdcycle (CT) method
28 Culture of HumanDermal Fibroblasts Human dermal fi-broblasts isolated from adult skin were obtained from Cas-cade Biologics (Portland OR USA) The cells were culturedat 37∘C in a humidified atmosphere containing 5 CO
2
and95 air in growth medium (Iscoversquos Modified Dulbeccorsquosmedium (GIBCO-BRL Grand Island NY USA) containing10 fetal bovine serum 100UmLminus1 penicillin 01mgmLminus1streptomycin and 025 120583gmLminus1 amphotericin B) In exper-iments on the paracrine effects of factors secreted by kerat-inocytes the medium was completely replaced by the condi-tioned medium of UVB-irradiated HaCaT cells and fibrob-lasts were cultivated for 24 h before analysis
29 Western Blotting Whole cell lysates were prepared usinga lysis buffer (10mM Tris-Cl pH 74 120mM NaCl 25mMKCl 2mM EGTA 1mM EDTA 05 Triton X-100 and pro-tease inhibitor cocktail) Aliquots of lysates were subjectedto sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) under protein denaturing conditions Proteinsseparated were transferred to polyvinylidene fluoride mem-branes which were incubated with an appropriate primaryantibody overnight at 4∘C and then with a secondary anti-body conjugated to horseradish peroxidase for 1 h at room
temperature Immunoreactive bands were detected usinga picoEPD Western Reagent kit (ELPIS-Biotech DaejeonKorea) and subjected to densitometric analysis The primaryantibody for MMP-1 was purchased from Calbiochem (SanDiego CA USA) Rabbit polyclonal caspase-3 antibodyand rabbit polyclonal caspase-9 were purchased from CellSignaling (Danvers MA USA) The mouse monoclonal 120573-actin antibody was from Sigma-Aldrich (St Louis MOUSA)
210 Analysis of Lipid Peroxidation As a marker of lipid per-oxidation 2-thiobarbituric acid-reactive substances (TBARS)were quantified [19] Briefly cells were treated in a lysisbuffer (20mM Tris-Cl 25mM EDTA 10 SDS pH 75)Cell lysates (200mg protein in 100 120583L) were mixed with900 120583L of 10 phosphoric acid and 10mL of 09 2-thiobarbituric acid (Sigma-Aldrich) and then heated on aboiling water bath for 45min Standard solutions of 1133-tetramethoxypropane (Sigma-Aldrich) a precursor of mal-ondialdehyde were treated in the same way as cell lysatesAfter cooling 15mL of 1-butanol was added and the mixturewas centrifuged at 13000 rpm for 15min to separate into twolayers The fluorescence intensity of the 1-butanol layer wasmeasured at an emission wavelength of 590 nm (excitationat 540 nm) using the Gemini EM fluorescence microplatereader (Molecular Devices Sunnyvale CA USA)
211 Statistical Analysis Data are presented as the means plusmnSEs of three or more independent experiments The signif-icance of differences between groups was determined usingthe Studentrsquos 119905-test Duncanrsquos multiple-range test was appliedwhen differences were significant (119875 lt 005) by the Studentrsquos119905-test Statistical significance was accepted for119875 valueslt 005
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 3
Vehicle10 30 100 10 30 100 10 30 100
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
0255075
100125
Cel
l via
bilit
y(
of c
ontro
l)
Figure 2 Effects of plant extracts derived from Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarum on theviability of HaCaT keratinocytes Cells were treated with plantextracts at the indicated concentration for 24 h and cell viabilitieswere determined using an MTT assay Cell viabilities are presentedas percentages of the vehicle control (means plusmn SEs 119899 = 3) 119875 lt 005versus the vehicle control
in dermal fibroblasts can be stimulated by direct expo-sure of the fibroblasts to UV In addition MMP-1 expres-sion in dermal fibroblasts can be stimulated indirectly bythe cytokines or other factors secreted by epidermal ker-atinocytes exposed to UV Previous studies have shown thatthe conditioned medium from UV irradiated keratinocyteseffectively increases MMP-1 expression in fibroblasts viaparacrine effects [14 15] Thus MMP-1 expression in dermalfibroblasts may be regulated indirectly by inhibiting theactivation of epidermal keratinocytes in response to UV Inthis regard plant extracts with UV-shielding antioxidativeand anti-inflammatory properties are an attractive prospect[16 17]
The purpose of the present study was to examine theprotective effects of plant extracts derived fromGardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumagainst skin cell responses to UVThese yellow plant extractswere chosen because of their ability to absorb UV effectively
2 Materials and Methods
21 Plant Materials Extracts of Gardenia jasminoides Ellis(seed) Phellodendron amurense Rupr (root) and Rheumrhabarbarum L (root) were prepared at Gyeongbuk NaturalColor Industry Institute (httpwwwgncrekr) Voucherspecimens of the plants and other information regarding theextracts are available at this institute Plant materials were ex-tracted with hot water 80∘C for 2 h and extracts were evap-orated to dryness under reduced pressure
22 UV Spectrophotometry Test materials were dissolved inphosphate buffered saline (PBS) to final concentrations of10 30 and 100 120583gmLminus1 Absorption spectra were recordedin the 200sim600 nm range using a Shimadzu UV-1650PCspectrophotometer (Shimadzu Corporation Kyoto Japan)
23 HPLC Analysis HPLC analysis of extracts was doneusing a Gilson HPLC system (Gilson Inc Middleton WIUSA) equipped with a 321 pump and UVVIS 151 detectorAqueous solution of test materials (10mgmLminus1) was injected
at 10 120583L Separation was done on a 5 120583m Hector-M C18 col-umn (46mm times 250mm) (RS Tech Co Daejeon Korea)The mobile phase consisted of 05 formic acid (A) and ac-etonitrile (B) The gradient was programmed as follows 0ndash20min a linear gradient from20 to 80B 20ndash35min 80BThe flow rate was 05mLminminus1 The UV detector was setat 280 nm and 440 nm Purified crocin was purchased fromSigma-Aldrich (St Louis MO USA) and its purity was deter-mined spectrophotometrically using the extinction coeffi-cient 120576
443
= 89000Mminus1 cmminus1 [18]
24 HaCaT Cell Culture HaCaT cells (a human immortal-ized keratinocyte cell-line) were grown in DMEMF-12 me-dium (GIBCO-BRL Grand Island NY USA) supplementedwith a 10 fetal bovine serum 100UmLminus1 penicillin01mgmLminus1 streptomycin 025 120583gmLminus1 amphotericin B and10 120583gmLminus1 hydrocortisone Cells were cultured at 37∘C in ahumidified atmosphere containing 5 CO
2
and 95 air
25 UVB-Exposure of HaCaT Cells HaCaT cells were seededon a six-well plate at a density of 2 times 105 cells per well andgrown in a culture medium for 48 h to reach 80 confluencyThe cells were then washed twice with PBS and exposed toUVB in PBS containing the test material UVB treatmentwas conducted under a cell culture hood using a UVB lamp(Model UVB-18 ULTRAlowastLUM Inc Claremont CA USA)that emitted radiation in the wavelength range of 280 to340 nm with maximum intensity at 300 nm The intensity ofradiation was determined using a UV light meter (ModelUV 340A Lutron Electronic Enterprise Co Taipei Taiwan)and the UV was administered to cells in culture plates atan intensity of 80 120583Wcmminus2 However durations of treatmentvaried to provide specific doses of UVB (5 10 or 15mJ cmminus2)Following irradiation PBS was replaced by growth mediumand cells were incubated for 1 day Cells were subjected tovarious tests as detailed below Culture conditioned mediumwas harvested and used to treat human dermal fibroblasts
26 Cell Viability Test Cell viability was assayed using 3-[45-dimethylthiazol-2-yl]-25-diphenyltetrazolium bromide(MTT) This assay method is based on the ability of mito-chondrial dehydrogenase in viable cells to reduce pale yellowMTT to dark blue formazan crystals that accumulate withincells Briefly cells were washed with PBS and incubated in1mL of culturemedia supplementedwith 1mgmLminus1MTT for3 h The medium was then discarded and cells were treatedwith isopropanol to solubilize the formazan Solutions werethen transferred to microplates and formazan was quantifiedby measuring absorbance at 595 nm
27 Quantitative Polymerase Chain Reaction (qRT-PCR)Anal-ysis Total cellular RNA was extracted using the RNeasy kit(Qiagen Valencia CA USA) To prepare cDNA one 120583g ofcellular mRNA was reverse transcribed using the High Ca-pacity cDNA Archive Kit (Applied Biosystems Foster CityCA USA) This kit utilizes random hexamers primers andMultiScribe Reverse Transcriptase PCRwas conducted using
4 Evidence-Based Complementary and Alternative Medicine
0 5 10 15
0
25
50
75
100
125
Cel
l via
bilit
y (
of c
ontro
l)
UVB (mJ cmminus2)
(a)
0255075
100125
Cel
l via
bilit
y (
of c
ontro
l)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
UVB (10mJ cmminus2)
lowast lowast
lowast
lowast lowast
lowast
(b)
Figure 3 Effects of plant extracts on UVB cytotoxicity in HaCaT cells In (a) HaCaT cells were irradiated with 5sim15mJ cmminus2 UVB In (b)cells were irradiated with 10mJ cmminus2 UVB in PBS in the absence or presence of plant extracts (10 30 or 100120583gmLminus1) Irradiated cells weresubsequently incubated for 24 h and cell viabilities were determined using an MTT assay Cell viabilities are presented as percentages of thevehicle control (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
0 5 10 15
0
5
10
15
20
UVB (mJ cmminus2)
IL-1120573
mRN
A (f
old)
(a)
0 5 10 15
0
5
10
15
20
TNF-120572
mRN
A (f
old)
UVB (mJ cmminus2)
(b)
05
101520
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
IL-1120573
mRN
A(fo
ld) lowast
lowast
lowast
UVB (10mJ cmminus2)
(c)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
02468
10
TNF-120572
mRN
A(fo
ld)
lowast lowast lowast
UVB (10mJ cmminus2)
(d)
Figure 4 Effects of plant extracts on mRNA expressions of inflammatory cytokines induced by UVB in HaCaT cells In (a) and (b) HaCaTcells were irradiated with 5sim15mJ cmminus2 UVB In (c) and (d) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence of plantextracts (10 30 or 100120583gmLminus1) and then incubated for 24 h before mRNA analysis by qRT-PCR GAPDH was used as the reference Dataare expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiatedwith UVB
the StepOnePlus Real-Time PCR System (Applied Biosys-tems) in reaction mixtures (20120583L) containing SYBR GreenPCRMasterMix (Applied Biosystems) 60 ng of cDNA and 2picomole of gene-specific primer sets (Macrogen Seoul)Theprimers used for PCR analysis were as follows interleukin-1120573 (IL-1120573) (GeneBank accession number NM 0005762)
51015840-CCT GTC CTG CGT GTT GAA AGA-31015840 (forward)and 51015840-GGG AAC TGG GCA GAC TCA AA-31015840 (reverse)tumor necrosis factor-120572 (TNF-120572) (NM 0005943) 51015840-TGCTCC TCA CCC ACA CCA T-31015840 (forward) and 51015840-GAGATA GTC GGG CCG ATT GA-31015840 (reverse) and GAPDH(NM 0020463) 51015840-ATG GGG AAG GTG AAG GTC G-31015840
Evidence-Based Complementary and Alternative Medicine 5
MMP-1
0
5
10
15
MM
P-1
prot
ein
(fold
)
0 5 10 15
UVB (mJ cmminus2)
120573-Actin
(a)
0
1
2
3
4
MM
P-1
prot
ein
(fold
)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV UVB (10mJ cmminus2)
MMP-1120573-Actin
lowastlowast
lowast
(b)
Figure 5 Expression levels of MMP-1 protein in human dermal fibroblasts stimulated by the conditioned media of HaCaT cells irradiatedwith UVB in the absence or presence of plant extracts In (a) HaCaT cells were irradiated with UVB at 5sim15mJ cmminus2 and then incubatedfor 24 h The conditioned medium from the UVB-irradiated HaCaT cells was used to treat human dermal fibroblasts for 24 h In (b) HaCaTcells were irradiated with UVB at 10mJ cmminus2 in PBS containing plant extracts at various concentrations 120583gmLminus1 and then incubated for 24 hConditionedmedia from these UVB-irradiated HaCaT cells were used to treat human dermal fibroblasts for 24 hMMP-1 protein expressionsin fibroblasts were analyzed by Western blot using 120573-actin as a loading control Typical blot images are shown Data are expressed as foldchanges (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
(forward) and 51015840-GGG GTC ATT GAT GGC AAC AA-31015840(reverse) Reactions were performed using the following con-ditions 50∘C for 2min 95∘C for 10min and 40 amplificationcycles (95∘C for 15 s and 60∘C for 1min) followed by adissociation stepMelting curve analysis showed single peakswhich supported the homogeneity of amplicons The mRNAexpression levels of IL-1120573 and TNF-120572 relative to that ofinternal control glyceraldehyde 3-phosphate dehydrogenase(GAPDH) were calculated using the comparative thresholdcycle (CT) method
28 Culture of HumanDermal Fibroblasts Human dermal fi-broblasts isolated from adult skin were obtained from Cas-cade Biologics (Portland OR USA) The cells were culturedat 37∘C in a humidified atmosphere containing 5 CO
2
and95 air in growth medium (Iscoversquos Modified Dulbeccorsquosmedium (GIBCO-BRL Grand Island NY USA) containing10 fetal bovine serum 100UmLminus1 penicillin 01mgmLminus1streptomycin and 025 120583gmLminus1 amphotericin B) In exper-iments on the paracrine effects of factors secreted by kerat-inocytes the medium was completely replaced by the condi-tioned medium of UVB-irradiated HaCaT cells and fibrob-lasts were cultivated for 24 h before analysis
29 Western Blotting Whole cell lysates were prepared usinga lysis buffer (10mM Tris-Cl pH 74 120mM NaCl 25mMKCl 2mM EGTA 1mM EDTA 05 Triton X-100 and pro-tease inhibitor cocktail) Aliquots of lysates were subjectedto sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) under protein denaturing conditions Proteinsseparated were transferred to polyvinylidene fluoride mem-branes which were incubated with an appropriate primaryantibody overnight at 4∘C and then with a secondary anti-body conjugated to horseradish peroxidase for 1 h at room
temperature Immunoreactive bands were detected usinga picoEPD Western Reagent kit (ELPIS-Biotech DaejeonKorea) and subjected to densitometric analysis The primaryantibody for MMP-1 was purchased from Calbiochem (SanDiego CA USA) Rabbit polyclonal caspase-3 antibodyand rabbit polyclonal caspase-9 were purchased from CellSignaling (Danvers MA USA) The mouse monoclonal 120573-actin antibody was from Sigma-Aldrich (St Louis MOUSA)
210 Analysis of Lipid Peroxidation As a marker of lipid per-oxidation 2-thiobarbituric acid-reactive substances (TBARS)were quantified [19] Briefly cells were treated in a lysisbuffer (20mM Tris-Cl 25mM EDTA 10 SDS pH 75)Cell lysates (200mg protein in 100 120583L) were mixed with900 120583L of 10 phosphoric acid and 10mL of 09 2-thiobarbituric acid (Sigma-Aldrich) and then heated on aboiling water bath for 45min Standard solutions of 1133-tetramethoxypropane (Sigma-Aldrich) a precursor of mal-ondialdehyde were treated in the same way as cell lysatesAfter cooling 15mL of 1-butanol was added and the mixturewas centrifuged at 13000 rpm for 15min to separate into twolayers The fluorescence intensity of the 1-butanol layer wasmeasured at an emission wavelength of 590 nm (excitationat 540 nm) using the Gemini EM fluorescence microplatereader (Molecular Devices Sunnyvale CA USA)
211 Statistical Analysis Data are presented as the means plusmnSEs of three or more independent experiments The signif-icance of differences between groups was determined usingthe Studentrsquos 119905-test Duncanrsquos multiple-range test was appliedwhen differences were significant (119875 lt 005) by the Studentrsquos119905-test Statistical significance was accepted for119875 valueslt 005
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Evidence-Based Complementary and Alternative Medicine
0 5 10 15
0
25
50
75
100
125
Cel
l via
bilit
y (
of c
ontro
l)
UVB (mJ cmminus2)
(a)
0255075
100125
Cel
l via
bilit
y (
of c
ontro
l)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
UVB (10mJ cmminus2)
lowast lowast
lowast
lowast lowast
lowast
(b)
Figure 3 Effects of plant extracts on UVB cytotoxicity in HaCaT cells In (a) HaCaT cells were irradiated with 5sim15mJ cmminus2 UVB In (b)cells were irradiated with 10mJ cmminus2 UVB in PBS in the absence or presence of plant extracts (10 30 or 100120583gmLminus1) Irradiated cells weresubsequently incubated for 24 h and cell viabilities were determined using an MTT assay Cell viabilities are presented as percentages of thevehicle control (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
0 5 10 15
0
5
10
15
20
UVB (mJ cmminus2)
IL-1120573
mRN
A (f
old)
(a)
0 5 10 15
0
5
10
15
20
TNF-120572
mRN
A (f
old)
UVB (mJ cmminus2)
(b)
05
101520
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
IL-1120573
mRN
A(fo
ld) lowast
lowast
lowast
UVB (10mJ cmminus2)
(c)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV
02468
10
TNF-120572
mRN
A(fo
ld)
lowast lowast lowast
UVB (10mJ cmminus2)
(d)
Figure 4 Effects of plant extracts on mRNA expressions of inflammatory cytokines induced by UVB in HaCaT cells In (a) and (b) HaCaTcells were irradiated with 5sim15mJ cmminus2 UVB In (c) and (d) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence of plantextracts (10 30 or 100120583gmLminus1) and then incubated for 24 h before mRNA analysis by qRT-PCR GAPDH was used as the reference Dataare expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiatedwith UVB
the StepOnePlus Real-Time PCR System (Applied Biosys-tems) in reaction mixtures (20120583L) containing SYBR GreenPCRMasterMix (Applied Biosystems) 60 ng of cDNA and 2picomole of gene-specific primer sets (Macrogen Seoul)Theprimers used for PCR analysis were as follows interleukin-1120573 (IL-1120573) (GeneBank accession number NM 0005762)
51015840-CCT GTC CTG CGT GTT GAA AGA-31015840 (forward)and 51015840-GGG AAC TGG GCA GAC TCA AA-31015840 (reverse)tumor necrosis factor-120572 (TNF-120572) (NM 0005943) 51015840-TGCTCC TCA CCC ACA CCA T-31015840 (forward) and 51015840-GAGATA GTC GGG CCG ATT GA-31015840 (reverse) and GAPDH(NM 0020463) 51015840-ATG GGG AAG GTG AAG GTC G-31015840
Evidence-Based Complementary and Alternative Medicine 5
MMP-1
0
5
10
15
MM
P-1
prot
ein
(fold
)
0 5 10 15
UVB (mJ cmminus2)
120573-Actin
(a)
0
1
2
3
4
MM
P-1
prot
ein
(fold
)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV UVB (10mJ cmminus2)
MMP-1120573-Actin
lowastlowast
lowast
(b)
Figure 5 Expression levels of MMP-1 protein in human dermal fibroblasts stimulated by the conditioned media of HaCaT cells irradiatedwith UVB in the absence or presence of plant extracts In (a) HaCaT cells were irradiated with UVB at 5sim15mJ cmminus2 and then incubatedfor 24 h The conditioned medium from the UVB-irradiated HaCaT cells was used to treat human dermal fibroblasts for 24 h In (b) HaCaTcells were irradiated with UVB at 10mJ cmminus2 in PBS containing plant extracts at various concentrations 120583gmLminus1 and then incubated for 24 hConditionedmedia from these UVB-irradiated HaCaT cells were used to treat human dermal fibroblasts for 24 hMMP-1 protein expressionsin fibroblasts were analyzed by Western blot using 120573-actin as a loading control Typical blot images are shown Data are expressed as foldchanges (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
(forward) and 51015840-GGG GTC ATT GAT GGC AAC AA-31015840(reverse) Reactions were performed using the following con-ditions 50∘C for 2min 95∘C for 10min and 40 amplificationcycles (95∘C for 15 s and 60∘C for 1min) followed by adissociation stepMelting curve analysis showed single peakswhich supported the homogeneity of amplicons The mRNAexpression levels of IL-1120573 and TNF-120572 relative to that ofinternal control glyceraldehyde 3-phosphate dehydrogenase(GAPDH) were calculated using the comparative thresholdcycle (CT) method
28 Culture of HumanDermal Fibroblasts Human dermal fi-broblasts isolated from adult skin were obtained from Cas-cade Biologics (Portland OR USA) The cells were culturedat 37∘C in a humidified atmosphere containing 5 CO
2
and95 air in growth medium (Iscoversquos Modified Dulbeccorsquosmedium (GIBCO-BRL Grand Island NY USA) containing10 fetal bovine serum 100UmLminus1 penicillin 01mgmLminus1streptomycin and 025 120583gmLminus1 amphotericin B) In exper-iments on the paracrine effects of factors secreted by kerat-inocytes the medium was completely replaced by the condi-tioned medium of UVB-irradiated HaCaT cells and fibrob-lasts were cultivated for 24 h before analysis
29 Western Blotting Whole cell lysates were prepared usinga lysis buffer (10mM Tris-Cl pH 74 120mM NaCl 25mMKCl 2mM EGTA 1mM EDTA 05 Triton X-100 and pro-tease inhibitor cocktail) Aliquots of lysates were subjectedto sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) under protein denaturing conditions Proteinsseparated were transferred to polyvinylidene fluoride mem-branes which were incubated with an appropriate primaryantibody overnight at 4∘C and then with a secondary anti-body conjugated to horseradish peroxidase for 1 h at room
temperature Immunoreactive bands were detected usinga picoEPD Western Reagent kit (ELPIS-Biotech DaejeonKorea) and subjected to densitometric analysis The primaryantibody for MMP-1 was purchased from Calbiochem (SanDiego CA USA) Rabbit polyclonal caspase-3 antibodyand rabbit polyclonal caspase-9 were purchased from CellSignaling (Danvers MA USA) The mouse monoclonal 120573-actin antibody was from Sigma-Aldrich (St Louis MOUSA)
210 Analysis of Lipid Peroxidation As a marker of lipid per-oxidation 2-thiobarbituric acid-reactive substances (TBARS)were quantified [19] Briefly cells were treated in a lysisbuffer (20mM Tris-Cl 25mM EDTA 10 SDS pH 75)Cell lysates (200mg protein in 100 120583L) were mixed with900 120583L of 10 phosphoric acid and 10mL of 09 2-thiobarbituric acid (Sigma-Aldrich) and then heated on aboiling water bath for 45min Standard solutions of 1133-tetramethoxypropane (Sigma-Aldrich) a precursor of mal-ondialdehyde were treated in the same way as cell lysatesAfter cooling 15mL of 1-butanol was added and the mixturewas centrifuged at 13000 rpm for 15min to separate into twolayers The fluorescence intensity of the 1-butanol layer wasmeasured at an emission wavelength of 590 nm (excitationat 540 nm) using the Gemini EM fluorescence microplatereader (Molecular Devices Sunnyvale CA USA)
211 Statistical Analysis Data are presented as the means plusmnSEs of three or more independent experiments The signif-icance of differences between groups was determined usingthe Studentrsquos 119905-test Duncanrsquos multiple-range test was appliedwhen differences were significant (119875 lt 005) by the Studentrsquos119905-test Statistical significance was accepted for119875 valueslt 005
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
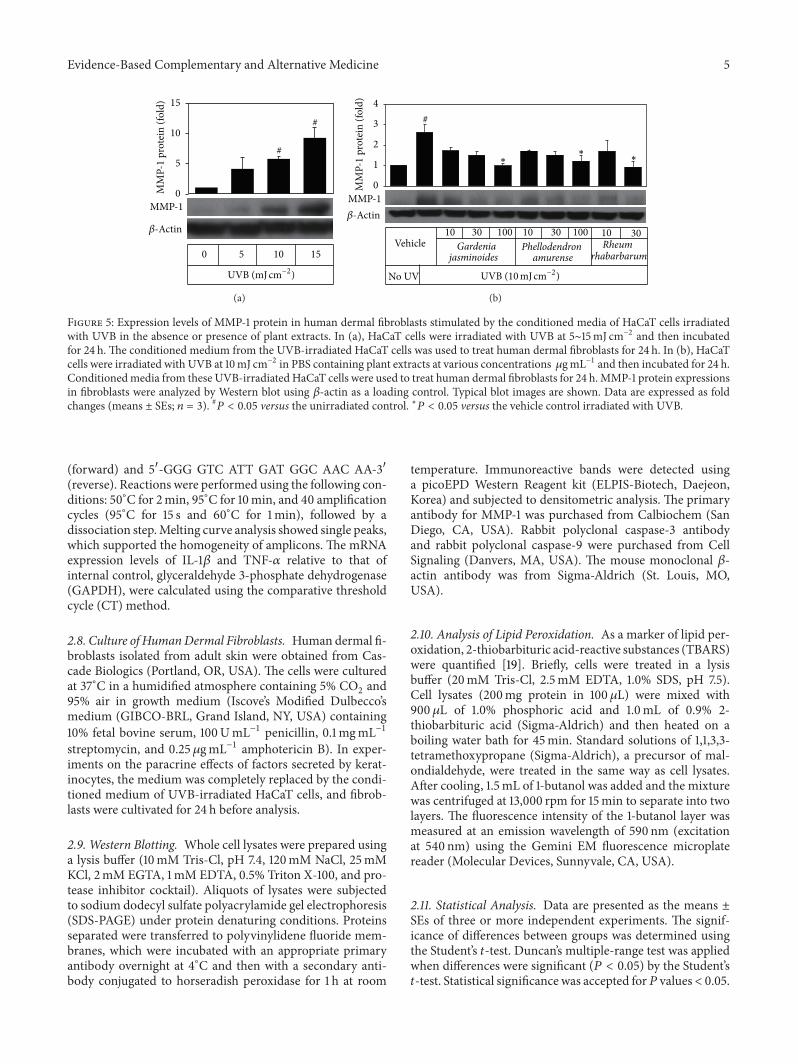
Evidence-Based Complementary and Alternative Medicine 5
MMP-1
0
5
10
15
MM
P-1
prot
ein
(fold
)
0 5 10 15
UVB (mJ cmminus2)
120573-Actin
(a)
0
1
2
3
4
MM
P-1
prot
ein
(fold
)
Vehicle10 30 100 10 30 100 10 30
Gardenia jasminoides
Phellodendronamurense
Rheum rhabarbarum
No UV UVB (10mJ cmminus2)
MMP-1120573-Actin
lowastlowast
lowast
(b)
Figure 5 Expression levels of MMP-1 protein in human dermal fibroblasts stimulated by the conditioned media of HaCaT cells irradiatedwith UVB in the absence or presence of plant extracts In (a) HaCaT cells were irradiated with UVB at 5sim15mJ cmminus2 and then incubatedfor 24 h The conditioned medium from the UVB-irradiated HaCaT cells was used to treat human dermal fibroblasts for 24 h In (b) HaCaTcells were irradiated with UVB at 10mJ cmminus2 in PBS containing plant extracts at various concentrations 120583gmLminus1 and then incubated for 24 hConditionedmedia from these UVB-irradiated HaCaT cells were used to treat human dermal fibroblasts for 24 hMMP-1 protein expressionsin fibroblasts were analyzed by Western blot using 120573-actin as a loading control Typical blot images are shown Data are expressed as foldchanges (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus the vehicle control irradiated with UVB
(forward) and 51015840-GGG GTC ATT GAT GGC AAC AA-31015840(reverse) Reactions were performed using the following con-ditions 50∘C for 2min 95∘C for 10min and 40 amplificationcycles (95∘C for 15 s and 60∘C for 1min) followed by adissociation stepMelting curve analysis showed single peakswhich supported the homogeneity of amplicons The mRNAexpression levels of IL-1120573 and TNF-120572 relative to that ofinternal control glyceraldehyde 3-phosphate dehydrogenase(GAPDH) were calculated using the comparative thresholdcycle (CT) method
28 Culture of HumanDermal Fibroblasts Human dermal fi-broblasts isolated from adult skin were obtained from Cas-cade Biologics (Portland OR USA) The cells were culturedat 37∘C in a humidified atmosphere containing 5 CO
2
and95 air in growth medium (Iscoversquos Modified Dulbeccorsquosmedium (GIBCO-BRL Grand Island NY USA) containing10 fetal bovine serum 100UmLminus1 penicillin 01mgmLminus1streptomycin and 025 120583gmLminus1 amphotericin B) In exper-iments on the paracrine effects of factors secreted by kerat-inocytes the medium was completely replaced by the condi-tioned medium of UVB-irradiated HaCaT cells and fibrob-lasts were cultivated for 24 h before analysis
29 Western Blotting Whole cell lysates were prepared usinga lysis buffer (10mM Tris-Cl pH 74 120mM NaCl 25mMKCl 2mM EGTA 1mM EDTA 05 Triton X-100 and pro-tease inhibitor cocktail) Aliquots of lysates were subjectedto sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE) under protein denaturing conditions Proteinsseparated were transferred to polyvinylidene fluoride mem-branes which were incubated with an appropriate primaryantibody overnight at 4∘C and then with a secondary anti-body conjugated to horseradish peroxidase for 1 h at room
temperature Immunoreactive bands were detected usinga picoEPD Western Reagent kit (ELPIS-Biotech DaejeonKorea) and subjected to densitometric analysis The primaryantibody for MMP-1 was purchased from Calbiochem (SanDiego CA USA) Rabbit polyclonal caspase-3 antibodyand rabbit polyclonal caspase-9 were purchased from CellSignaling (Danvers MA USA) The mouse monoclonal 120573-actin antibody was from Sigma-Aldrich (St Louis MOUSA)
210 Analysis of Lipid Peroxidation As a marker of lipid per-oxidation 2-thiobarbituric acid-reactive substances (TBARS)were quantified [19] Briefly cells were treated in a lysisbuffer (20mM Tris-Cl 25mM EDTA 10 SDS pH 75)Cell lysates (200mg protein in 100 120583L) were mixed with900 120583L of 10 phosphoric acid and 10mL of 09 2-thiobarbituric acid (Sigma-Aldrich) and then heated on aboiling water bath for 45min Standard solutions of 1133-tetramethoxypropane (Sigma-Aldrich) a precursor of mal-ondialdehyde were treated in the same way as cell lysatesAfter cooling 15mL of 1-butanol was added and the mixturewas centrifuged at 13000 rpm for 15min to separate into twolayers The fluorescence intensity of the 1-butanol layer wasmeasured at an emission wavelength of 590 nm (excitationat 540 nm) using the Gemini EM fluorescence microplatereader (Molecular Devices Sunnyvale CA USA)
211 Statistical Analysis Data are presented as the means plusmnSEs of three or more independent experiments The signif-icance of differences between groups was determined usingthe Studentrsquos 119905-test Duncanrsquos multiple-range test was appliedwhen differences were significant (119875 lt 005) by the Studentrsquos119905-test Statistical significance was accepted for119875 valueslt 005
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
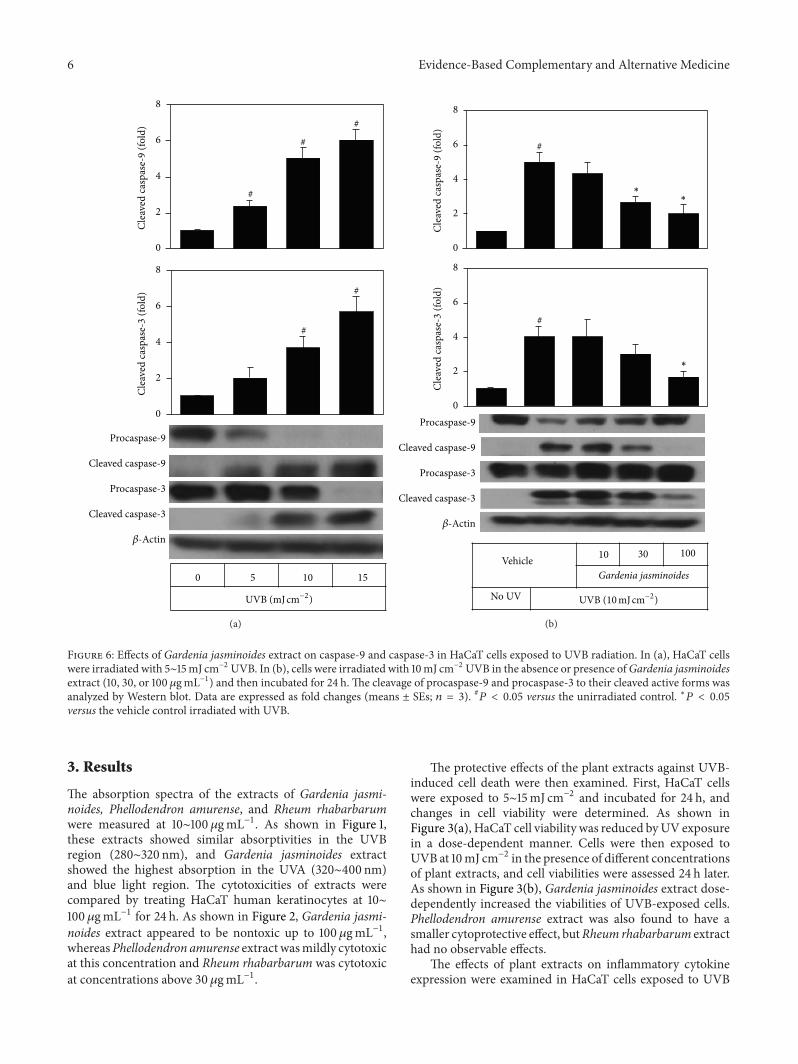
6 Evidence-Based Complementary and Alternative Medicine
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
0 5 10 15
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
UVB (mJ cmminus2)
(a)
0
2
4
6
8
Clea
ved
casp
ase-
9 (fo
ld)
0
2
4
6
8
Clea
ved
casp
ase-
3 (fo
ld)
Vehicle 10 30 100
Gardenia jasminoides
No UV
Procaspase-9
Cleaved caspase-9
Procaspase-3
Cleaved caspase-3
120573-Actin
lowastlowast
lowast
UVB (10mJ cmminus2)
(b)
Figure 6 Effects of Gardenia jasminoides extract on caspase-9 and caspase-3 in HaCaT cells exposed to UVB radiation In (a) HaCaT cellswere irradiated with 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoidesextract (10 30 or 100120583gmLminus1) and then incubated for 24 h The cleavage of procaspase-9 and procaspase-3 to their cleaved active forms wasanalyzed by Western blot Data are expressed as fold changes (means plusmn SEs 119899 = 3) 119875 lt 005 versus the unirradiated control lowast119875 lt 005versus the vehicle control irradiated with UVB
3 Results
The absorption spectra of the extracts of Gardenia jasmi-noides Phellodendron amurense and Rheum rhabarbarumwere measured at 10sim100 120583gmLminus1 As shown in Figure 1these extracts showed similar absorptivities in the UVBregion (280sim320 nm) and Gardenia jasminoides extractshowed the highest absorption in the UVA (320sim400 nm)and blue light region The cytotoxicities of extracts werecompared by treating HaCaT human keratinocytes at 10sim100 120583gmLminus1 for 24 h As shown in Figure 2 Gardenia jasmi-noides extract appeared to be nontoxic up to 100 120583gmLminus1whereasPhellodendron amurense extract wasmildly cytotoxicat this concentration and Rheum rhabarbarum was cytotoxicat concentrations above 30 120583gmLminus1
The protective effects of the plant extracts against UVB-induced cell death were then examined First HaCaT cellswere exposed to 5sim15mJ cmminus2 and incubated for 24 h andchanges in cell viability were determined As shown inFigure 3(a) HaCaT cell viability was reduced byUV exposurein a dose-dependent manner Cells were then exposed toUVB at 10mJ cmminus2 in the presence of different concentrationsof plant extracts and cell viabilities were assessed 24 h laterAs shown in Figure 3(b) Gardenia jasminoides extract dose-dependently increased the viabilities of UVB-exposed cellsPhellodendron amurense extract was also found to have asmaller cytoprotective effect butRheum rhabarbarum extracthad no observable effects
The effects of plant extracts on inflammatory cytokineexpression were examined in HaCaT cells exposed to UVB
Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 7
06
08
1
12
14
0 5 10 15
Lipi
d pe
roxi
datio
n (fo
ld)
UVB (mJ cmminus2)
(a)
06
08
1
12
14
Lipi
d pe
roxi
datio
n (fo
ld)
Vehicle 10 30 100Gardenia jasminoides
No UV
lowast
UVB (10mJ cmminus2)
(b)
Figure 7 Effects of Gardenia jasminoides extract on UVB-induced lipid peroxidation in HaCaT cells In (a) HaCaT cells were irradiatedwith 5sim15mJ cmminus2 UVB In (b) cells were irradiated with 10mJ cmminus2 UVB in the absence or presence ofGardenia jasminoides extract (10 30or 100120583gmLminus1) and then incubated for 24 h Lipid peroxidation was determined by quantifying TBARS Data are expressed as fold changes(means plusmn SEs 119899 = 3) Control value was 033 nmol TBARS per mg protein 119875 lt 005 versus the unirradiated control lowast119875 lt 005 versus thevehicle control irradiated with UVB
at 5sim15mJ cmminus2 and incubated for 24 h Total cellular mRNAwas extracted and the mRNA levels of cytokines weredetermined by quantitative Real-Time PCR As shown inFigure 4(a) UVB increased the mRNA expressions of IL-1120573and TNF-120572 markedly Cells were then exposed to UVB at10mJ cmminus2 in the presence of plant extracts at different con-centrations It was found that Gardenia jasminoides extractat 30sim100 120583gmLminus1 attenuated inflammatory cytokine expres-sion in HaCaT cells exposed to UVB and Phellodendronamurense extract showed a smaller effect
Because UVB increased inflammatory cytokine expres-sion in HaCaT cells we examined whether the conditionedmedium of UVB-exposed HaCaT keratinocytes which con-tained inflammatory cytokines stimulated MMP-1 expres-sion in fibroblasts via paracrine effects When conditionedmedium fromHaCaT cells exposed to different doses of UVBwas added to human dermal fibroblasts cultures and culturedfor 24 h MMP-1 expressions in fibroblasts were found toincrease in a UVB dose-dependent manner (Figure 5(a)) Wethen examined the effects of plant extracts on the paracrineeffects of cytokines secreted by UVB-exposed HaCaT ker-atinocytes on MMP-1 expression in fibroblasts As shown inFigure 5(b) the presence of plant extracts such as Gardeniajasminoides extract during UVB exposure of HaCaT cellsrendered the conditioned medium less effective in increasingMMP-1 expression in fibroblasts
Apoptotic cell death is also known as one of the majormechanisms of photoaging of the skin [20] Because caspase-9 and caspase-3 are known to be critical players in UV-induced apoptosis in keratinocytes [21] we examined theeffect of Gardenia jasminoides extract on the activation ofthese apoptosis mediator enzymes in HaCaT cells exposedto UVB As expected UVB irradiation increased cleavageof procaspase-9 and procaspase-3 to their cleaved activeforms in a UVB dose-dependent manner (Figure 6(a)) Itwas also observed that the UVB-induced increases of cleavedcaspase-9 and cleaved caspase-3 were markedly prevented
when Gardenia jasminoides extract was included in themedium (Figure 6(b))
UV can also induce oxidative stress in cells by stimu-lating overproduction of reactive oxygen species (ROS) anddepleting antioxidants [22 23] In the current study wedetermined the levels of TBARS tomonitor lipid peroxidation[19] The results showed that lipid peroxidation increased inHaCaT cells exposed to UVB (Figure 7(a)) and this changewas attenuated by Gardenia jasminoides extract in a dose-dependent manner (Figure 7(b))
Typical HPLC patterns of the extracts of Gardenia jas-minoides Phellodendron amurense and Rheum rhabarbarumare shown in Figure 8 Commercially sourced crocinwas usedas a reference compound because it is known as a majorpigment in Gardenia jasminoides [24] The content of crocinin Gardenia jasminoides extract was estimated to be 17 Itwas not detected in the extract of Phellodendron amurense orRheum rhabarbarum
4 Discussion
UV is a major cause of skin photoaging and photocarcino-genesis [25] UV consists of UVA (320sim400 nm) UVB (280sim320 nm) and UVC (200sim280 nm) and overexposure to UVparticularly the UVB component causes erythema edemahyperplasia hyperpigmentation photoaging immunosup-pression and skin cancer [2 26] Overexposure to UValso causes oxidative stress as evidenced by increased lipidperoxidation and by the depletion of cutaneous antioxidants[27 28] Previous studies have demonstrated the preventiveeffects of exogenous antioxidants on photocarcinogenesis[29] Plant extracts are attractive cosmeceuticals becausesome are rich in secondary metabolites that provide UVB-screening antioxidative and anti-inflammatory activitiesThe results of the current study indicate that yellow-coloredplant extracts likeGardenia jasminoides extract have potential
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
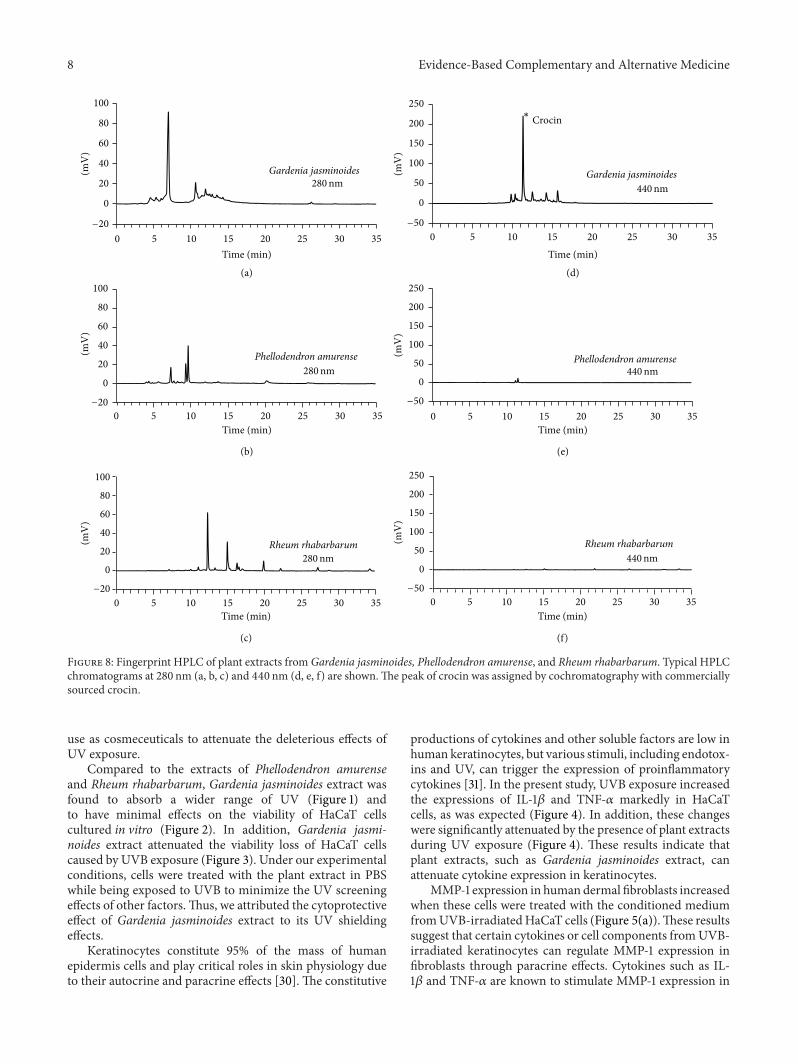
8 Evidence-Based Complementary and Alternative Medicine
0
20
40
60
80
100
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
minus20
280nm
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nmPhellodendron amurense
Time (min)
20
0 5 10 15 20 25 30 35
0
40
60
80
100
(mV
)
minus20
280nm
Time (min)
Rheum rhabarbarum
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
Gardenia jasminoides
Crocin
minus50
440nm
lowast
Time (min)
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)minus50
440nm
Time (min)
Phellodendron amurense
0
50
100
150
200
250
0 5 10 15 20 25 30 35
(mV
)
minus50
440nm
Time (min)
Rheum rhabarbarum
(a)
(b)
(c)
(d)
(e)
(f)
Figure 8 Fingerprint HPLC of plant extracts from Gardenia jasminoides Phellodendron amurense and Rheum rhabarbarum Typical HPLCchromatograms at 280 nm (a b c) and 440 nm (d e f) are shownThe peak of crocin was assigned by cochromatography with commerciallysourced crocin
use as cosmeceuticals to attenuate the deleterious effects ofUV exposure
Compared to the extracts of Phellodendron amurenseand Rheum rhabarbarum Gardenia jasminoides extract wasfound to absorb a wider range of UV (Figure 1) andto have minimal effects on the viability of HaCaT cellscultured in vitro (Figure 2) In addition Gardenia jasmi-noides extract attenuated the viability loss of HaCaT cellscaused by UVB exposure (Figure 3) Under our experimentalconditions cells were treated with the plant extract in PBSwhile being exposed to UVB to minimize the UV screeningeffects of other factorsThus we attributed the cytoprotectiveeffect of Gardenia jasminoides extract to its UV shieldingeffects
Keratinocytes constitute 95 of the mass of humanepidermis cells and play critical roles in skin physiology dueto their autocrine and paracrine effects [30] The constitutive
productions of cytokines and other soluble factors are low inhuman keratinocytes but various stimuli including endotox-ins and UV can trigger the expression of proinflammatorycytokines [31] In the present study UVB exposure increasedthe expressions of IL-1120573 and TNF-120572 markedly in HaCaTcells as was expected (Figure 4) In addition these changeswere significantly attenuated by the presence of plant extractsduring UV exposure (Figure 4) These results indicate thatplant extracts such as Gardenia jasminoides extract canattenuate cytokine expression in keratinocytes
MMP-1 expression in human dermal fibroblasts increasedwhen these cells were treated with the conditioned mediumfromUVB-irradiatedHaCaT cells (Figure 5(a))These resultssuggest that certain cytokines or cell components fromUVB-irradiated keratinocytes can regulate MMP-1 expression infibroblasts through paracrine effects Cytokines such as IL-1120573 and TNF-120572 are known to stimulate MMP-1 expression in
Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 9
fibroblasts [32] other factors such as stratifin also could play arole [15 33] NonethelessGardenia jasminoides extract atten-uated these paracrine effects in a dose-dependent manner(Figure 5(b)) which was in line with its effects on cytokineexpressions in HaCaT cells (Figure 4)
UV can induce apoptosis of keratinocytes via intrinsicpathways involving direct DNA damage extrinsic pathwaysinvolving activated cell membrane death receptors andgeneration of ROS [20 21] Occurrence of apoptosis canbe monitored by using various markers including DNAladdering changes of proapoptotic (Bax Bak and Bid) andantiapoptotic (Bcl-2 and Bcl-x) members of the Bcl-2 proteinfamily activation of caspases and so on In the presentstudy we observedUVB dose-dependent increases in cleavedactive forms of caspase-9 and caspase-3 and attenuation ofthese changes by Gardenia jasminoides extract (Figure 6)In addition Gardenia jasminoides extract was observed toattenuate UVB-induced oxidative stress as monitored bylipid peroxidation (Figure 7) Therefore Gardenia jasmi-noides extract was considered to be useful in protecting skincells from UV-induced oxidative stress inflammation andapoptotic death
Gardenia jasminoides belongs to the botanical familyRubiaceae and is found in South Asia Gardenia jasminoidesfruits have been traditionally used in oriental medicine totreat jaundice fever hypertension and skin ulcers Its yellowpigment has been used as a natural food colorant and dyeThemajor constituent of this yellow pigment is crocin a watersoluble carotenoid derivative [24] and the pharmacologicaleffects of crocin and of its aglycone crocetin have beenwidely investigated [34 35]Theprotective effects ofGardeniajasminoides extract against UVB-induced injury and inflam-matory responses of skin cells might be attributed to the UVshielding effect of crocin (Figures 1 and 8)
5 Conclusion
In conclusion this study suggests that UVB-induced oxida-tive stress apoptotic cell death and inflammatory responsesof skin cells can be attenuated by yellow-colored plantextracts such as Gardenia jasminoides extract that absorbUV effectively and these extracts are useful cosmetic ingre-dients
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This study was supported by a grant (Grant no A103017)from the Korea Health Technology RampD Project Ministry ofHealth andWelfare and by a grant (Grant no R0002039) fromthe Regional Specialized Technology Convergence ProjectMinistry of Trade Industry and Energy Republic of Korea
References
[1] C Antoniou M G Kosmadaki A J Stratigos and A DKatsambas ldquoPhotoaging prevention and topical treatmentsrdquoAmerican Journal of Clinical Dermatology vol 11 no 2 pp 95ndash102 2010
[2] G-E Costin and V J Hearing ldquoHuman skin pigmentationmelanocytes modulate skin color in response to stressrdquo FASEBJournal vol 21 no 4 pp 976ndash994 2007
[3] K Scharffetter-Kochanek P Brenneisen J Wenk et al ldquoPho-toaging of the skin from phenotype to mechanismsrdquo Experi-mental Gerontology vol 35 no 3 pp 307ndash316 2000
[4] V-M Kahari and U Saarialho-Kere ldquoMatrix metallopro-teinases in skinrdquo Experimental Dermatology vol 6 no 5 pp199ndash213 1997
[5] W Bode C Fernandez-Catalan H Tschesche F Grams HNagase and K Maskos ldquoStructural properties of matrix metal-loproteinasesrdquo Cellular and Molecular Life Sciences vol 55 no4 pp 639ndash652 1999
[6] K Scharffetter M Wlaschek A Hogg et al ldquoUVA irradiationinduces collagenase in human dermal fibroblasts in vitro and invivordquo Archives of Dermatological Research vol 283 no 8 pp506ndash511 1991
[7] P Brenneisen J Oh M Wlaschek et al ldquoUltraviolet B wave-length dependence for the regulation of two major matrix-metalloproteinases and their inhibitor TIMP-1 in human der-mal fibroblastsrdquo Photochemistry and Photobiology vol 64 no5 pp 877ndash885 1996
[8] S Pillai C Oresajo and J Hayward ldquoUltraviolet radiationand skin aging roles of reactive oxygen species inflamma-tion and protease activation and strategies for prevention ofinflammation-induced matrix degradationmdasha reviewrdquo Interna-tional Journal of Cosmetic Science vol 27 no 1 pp 17ndash34 2005
[9] G J Fisher S Kang J Varani et al ldquoMechanisms of photoagingand chronological skin agingrdquoArchives of Dermatology vol 138no 11 pp 1462ndash1470 2002
[10] G J Fisher H S Talwar J Lin et al ldquoRetinoic acid inhibitsinduction of c-Jun protein by ultraviolet radiation that occurssubsequent to activation of mitogen-activated protein kinasepathways in human skin in vivordquo Journal of Clinical Investiga-tion vol 101 no 6 pp 1432ndash1440 1998
[11] P Brenneisen H Sies and K Scharffetter-KochanekldquoUltraviolet-B irradiation and matrix metalloproteinasesfrom induction via signaling to initial eventsrdquo Annals of theNew York Academy of Sciences vol 973 pp 31ndash43 2002
[12] J Westermarck and V-M Kahari ldquoRegulation of matrix metal-loproteinase expression in tumor invasionrdquo FASEB Journal vol13 no 8 pp 781ndash792 1999
[13] K K Dong N Damaghi S D Picart et al ldquoUV-induced DNAdamage initiates release of MMP-1 in human skinrdquo Experimen-tal Dermatology vol 17 no 12 pp 1037ndash1044 2008
[14] D Fagot D Asselineau and F Bernerd ldquoMatrix metallopro-teinase-1 production observed after solar-simulated radiationexposure is assumed by dermal fibroblasts but involves aparacrine activation through epidermal keratinocytesrdquo Photo-chemistry and Photobiology vol 79 no 6 pp 499ndash505 2004
[15] D Fagot D Asselineau and F Bernerd ldquoDirect role of humandermal fibroblasts and indirect participation of epidermalkeratinocytes in MMP-1 production after UV-B irradiationrdquoArchives of Dermatological Research vol 293 no 11 pp 576ndash583 2002
10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Evidence-Based Complementary and Alternative Medicine
[16] S K Katiyar N J Korman H Mukhtar and R Agarwal ldquoPro-tective effects of silymarin against photocarcinogenesis in amouse skin modelrdquo Journal of the National Cancer Institute vol89 no 8 pp 556ndash566 1997
[17] N Cole PW Sou ANgo et al ldquoTopical ldquoSydneyrdquo propolis pro-tects against UV-radiation-induced inflammation lipid perox-idation and immune suppression in mouse skinrdquo InternationalArchives of Allergy and Immunology vol 152 no 2 pp 87ndash972010
[18] L V Joslashrgensen H J Andersen and L H Skibsted ldquoKineticsof reduction of hypervalent iron in myoglobin by crocin inaqueous solutionrdquo Free Radical Research vol 27 no 1 pp 73ndash871997
[19] M Uchiyama and M Mihara ldquoDetermination of malonalde-hyde precursor in tissues by thiobarbituric acid testrdquo AnalyticalBiochemistry vol 86 no 1 pp 271ndash278 1978
[20] C H Lee S B Wu C H Hong H S Yu and Y HWei ldquoMolecular mechanisms of UV-induced apoptosis and itseffects on skin residential cells the implication in UV-basedphototherapyrdquo International Journal of Molecular Sciences vol14 no 3 pp 6414ndash6435 2013
[21] L A Sitailo S S Tibudan and M F Denning ldquoActivationof caspase-9 is required for UV-induced apoptosis of humankeratinocytesrdquo Journal of Biological Chemistry vol 277 no 22pp 19346ndash19352 2002
[22] G R Aitken J R Henderson S-C Chang C J McNeil andMA Birch-Machin ldquoDirect monitoring of UV-induced free rad-ical generation in HaCaT keratinocytesrdquo Clinical and Experi-mental Dermatology vol 32 no 6 pp 722ndash727 2007
[23] M PoddaMG Traber CWeber L-J Yan and L Packer ldquoUV-irradiation depletes antioxidants and causes oxidative damagein a model of human skinrdquo Free Radical Biology and Medicinevol 24 no 1 pp 55ndash65 1998
[24] T Watanabe and S Terabe ldquoAnalysis of natural food pigmentsby capillary electrophoresisrdquo Journal of Chromatography A vol880 no 1-2 pp 311ndash322 2000
[25] J H Epstein ldquoPhotocarcinogenesis skin cancer and agingrdquoJournal of the American Academy of Dermatology vol 9 no 4pp 487ndash502 1983
[26] F Afaq and H Mukhtar ldquoBotanical antioxidants in the pre-vention of photocarcinogenesis and photoagingrdquo ExperimentalDermatology vol 15 no 9 pp 678ndash684 2006
[27] N A Soter ldquoAcute effects of ultraviolet radiation on the skinrdquoSeminars in Dermatology vol 9 no 1 pp 11ndash15 1990
[28] F Afaq andHMukhtar ldquoEffects of solar radiation on cutaneousdetoxification pathwaysrdquo Journal of Photochemistry and Photo-biology B vol 63 no 1ndash3 pp 61ndash69 2001
[29] F DrsquoAgostini R M Balansky A Camoirano and S de FloraldquoModulation of light-induced skin tumors by N-acetylcysteineandor ascorbic acid in hairless micerdquo Carcinogenesis vol 26no 3 pp 657ndash664 2005
[30] B J Nickoloff and L A Turka ldquoKeratinocytes key immuno-cytes of the integumentrdquoAmerican Journal of Pathology vol 143no 2 pp 325ndash331 1993
[31] M Yoshizumi T Nakamura M Kato et al ldquoRelease ofcytokineschemokines and cell death inUVB-irradiated humankeratinocytes HaCaTrdquo Cell Biology International vol 32 no 11pp 1405ndash1411 2008
[32] K Ohguchi T Itoh Y Akao H Inoue Y Nozawa and M ItoldquoSIRT1 modulates expression of matrix metalloproteinases inhuman dermal fibroblastsrdquo British Journal of Dermatology vol163 no 4 pp 689ndash694 2010
[33] AGhahary F Karimi-Busheri YMarcoux et al ldquoKeratinocyte-releasable stratifin functions as a potent collagenase-stimulatingfactor in fibroblastsrdquo Journal of Investigative Dermatology vol122 no 5 pp 1188ndash1197 2004
[34] J-D Hsu F-P I Chou M-J Lee et al ldquoSuppression of theTPA-induced expression of nuclear-protooncogenes in mouseepidermis by crocetin via antioxidant activityrdquo Anticancer Re-search vol 19 no 5 pp 4221ndash4227 1999
[35] T Q Pham F Cormier E Farnworth H van Tong and M-Rvan Calsteren ldquoAntioxidant properties of crocin from Gardeniajasminoides Ellis and study of the reactions of crocin withlinoleic acid and crocin with oxygenrdquo Journal of Agriculturaland Food Chemistry vol 48 no 5 pp 1455ndash1461 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom