research article gab1 is modulated by chronic...
TRANSCRIPT

Research ArticleGab1 Is Modulated by Chronic Hypoxia in Childrenwith Cyanotic Congenital Heart Defect and Its OverexpressionReduces Apoptosis in Rat Neonatal Cardiomyocytes
Myriam Cherif1 Massimo Caputo12 Yoshikazu Nakaoka3
Gianni D Angelini1 and Mohamed T Ghorbel1
1 Bristol Heart Institute School of Clinical Sciences University of Bristol Research Level 7 Bristol Royal InfirmaryUpper Maudlin Street Bristol BS2 8HW UK
2RUSH University Medical Center Chicago IL 60612 USA3Department of Cardiovascular Medicine Osaka University Graduate School of Medicine 2-2 YamadaokaSuita Osaka 565-0871 Japan
Correspondence should be addressed to Mohamed T Ghorbel mghorbelbristolacuk
Received 25 July 2014 Revised 1 October 2014 Accepted 28 October 2014
Academic Editor Hajime Imura
Copyright copy 2015 Myriam Cherif et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Gab1 (Grb2 associated binding protein 1) is a member of the scaffoldingdocking proteins (Gab1 Gab2 and Gab3) It is required forfibroblast cell survival and maintaining cardiac function Very little is known about human Gab1 expression in response to chronichypoxia The present study examined the hypothesis that hypoxia regulates Gab1 expression in human paediatric myocardium andcultured rat cardiomyocytes Here we showed that Gab1 is expressed in myocardial tissue in acyanotic and cyanotic children withcongenital heart defects Gab1 protein was upregulated in cyanotic compared to acyanotic hearts suggesting that Gab1 upregulationis a component of the survival program initiated by hypoxia in cyanotic childrenThe expression of other Gab1 interacting partnerswas not affected by hypoxia and Gab1 regulation Additionally using an in vitromodel we demonstrated that overexpressing Gab1in neonatal cardiomyocytes under hypoxic condition resulted in the reduction of apoptosis suggesting a role for this protein incardiomyocyte survival Altogether our data provide strong evidence thatGab1 is important for heart cell survival following hypoxicstress
1 Introduction
Heart malformation during embryonic development cancause congenital heart diseases (CHD)These affect one babyin 125 live births and tetralogy of Fallot (TOF) representsthe most common form of the ldquoblue baby syndromerdquo InUK one baby in 3600 is born with TOF malformation[1] TOF malformation exhibits four abnormalities Theseinclude a ventricular septal defect (VSD) right ventricularhypertrophy overriding of the aorta and pulmonary stenosis(PS) [2]The causes that induce TOF are not fully understoodbut the aetiology is thought to be multifactorial Somestudies associated TOF with untreated maternal diabetesphenylketonuria and intake of retinoic acid In additionchromosomal abnormalities (such as trisomies 21 18 and 13)
have been shown to exhibit a higher TOF incidence [3] Thedegree of stenosis varies between individuals with TOF and isthe primary determinant of symptoms and severity IndeedTOF is divided into two categories acyanotic (pink) andcyanotic (blue) depending on the blood oxygen saturationAlthough successful corrective surgery of heart defects existsthere is an increased risk of morbidity and mortality incyanotic children compared with acyanotic [4] There isevidence that an unintended reoxygenation injury occursin myocardium of cyanotic patients due to the delivery ofhigh levels of oxygen during cardiopulmonary bypass (CPB)used in surgery which does not match preoperative levels ofoxygen in these children Reoxygenation injury produces anincrease in free radical production which may result in celldamage [5]
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 718492 8 pageshttpdxdoiorg1011552015718492
2 BioMed Research International
In previous study we have shown that chronic hypoxiain pediatric patients with TOF triggered the expression ofnetwork of genes associated with apoptosis and reduced theexpression of genes involved in myocyte contractility andfunction [6] This state of hypoxia in TOF children maybe responsible for the susceptibility of cyanotic children toreoxygenation injury during and after surgery We have alsoshown that concomitant with the initiation of the injuriousprogram a protective program is triggered by cyanosis Gab1shown to significantly increase at messenger level in cyanoticcompared to acyanotic patients [6] could be an importantplayer in this protective program
Grb2 associated binding protein 1 (Gab1) is a memberof the scaffoldingdocking proteins (Gab1 Gab2 and Gab3)[7 8] Gab1 knockout mice are not viable and displayimpaired development of heart placenta skin and muscle[9] In cultured cardiomyocytes Gab1 is shown to interactwith tyrosine phosphatase SHP2 and to promote cardiachypertrophy [10] There is evidence that Gab1 is essentialfor cardiac function in the postnatal heart in vivo [11] Inaddition Gab1 has been shown to exert an antiapoptotic roleinmouse embryonic fibroblasts and is activated through tyro-sine phosphorylation following oxidative treatment (H
2O2)
[12] In their investigation Holgado-Madruga and Wongidentified Gab1 as an important component in oxidativestress signalling with an essential role in the activation ofc-Jun NH(2)-terminal kinase (JNK) and the influencing ofcell survival [12] This Gab1 antiapoptotic role in fibroblastsfollowing oxidative treatment [12] has led us to hypothesisethat Gab1 may play similar role in cardiac tissue and cardiacmyocytes subjected to hypoxia
In this study we investigated the effects of cyanosison Gab1 in myocardium samples from paediatric patientssuffering from TOF and we examined the effects of hypoxiain primary cultures of rat neonatal cardiomyocytes on Gab1and its possible role in cell survival
2 Materials and Methods
21 Reagents All reagents were from Sigma (UK) exceptthose stated otherwise Gab1 antibody was from MilliporeAntibodies against SHP2 and p85 were from Cell SignallingTechnology (UK) GAPDH antibody was from ResearchDiagnostics Inc (UK)
22 Cardiac Biopsies The collection of human right ventriclespecimens used in this study was approved by the NorthSomerset and South Bristol Research Ethics Committee(REC reference 07H0106172) the National Research EthicsService England Parental informed written consent wasgained for all patients Patients with a diagnosis of cyanotic(O2saturation 796plusmn 75 age 106plusmn 55months) or acyanotic
(O2saturation 942 plusmn 35 age 95 plusmn 23months) tetralogy of
Fallot undergoing surgical repair at the Bristol Royal Hospitalfor Children were studied
Five ventricular biopsy specimens per group were col-lected from the right ventricle of acyanotic and cyanoticTOF patients by using ldquoTrue-Cutrdquo needle immediately after
institution of cardiopulmonary bypass (CPB) Each specimenwas immediately put in liquid nitrogen for protein extraction
23 Immunohistochemistry Right ventricular specimenswere fixed in 4 paraformaldehyde washed in PBS andembedded in paraffin and 4120583msectionswere obtained Imm-unohistochemistry was performed using the ABC-Kit fromDakocytomation Slides were observed with anOlympus B40microscope Pictures were taken using a Media Cyberneticscamera (Bethesda MD USA) and analysed with proimageplus software (Bethesda MD USA)
24 RatNeonatal PrimaryCulture All the procedures involv-ing laboratory animals conformed to the guidelines and reg-ulations of the University of Bristol and the United KingdomHome Office Neonatal rat cardiomyocytes were cultured aspreviously described with minor modifications [13 14] Onecell culture preparation was used for each experiment andeach experiment was repeated three times using different cellcultures prepared at different time from pooled neonatal rathearts For each cell culture 14ndash24 neonatal rats were usedto harvest hearts Hearts were quickly removed from one-to three-day-old Wistar rats and only ventricles were keptThey were washed with PBS three times and incubated with005 trypsin and 002 EDTA for 30 minutes They werethen enzymatically digested six times for fifteen minutes in01 trypsin and 002 EDTA in PBS Digestion was stoppedby addition of foetal calf serum at a final concentration of30 Cells were then centrifuged at 400 g for fiveminutes andresuspended in DMEM supplemented with 10 FBS and 1PS Cells were then incubated one to two hours in a T75 flaskto allow noncardiac myocytes (mainly cardiac fibroblasts) toadhere to plastic They were then plated in gelatin-coatedplates at a density of 128 times 105 cells per cm2 After 40 hoursof culture themediumwas changed inDMEMsupplementedwith 20 M-199 and 1 PS All treatments were performedon three-day-old cultures Hypoxia was induced by placingthe cells in a hypoxic chamber where oxygen levels could bemonitored (Biospherix ProOx C21 Lacona NY USA) TheProOx is connected to a sensor which monitors the oxygenconcentration within the host chamber The ProOx quicklyinfuses nitrogen (hypoxia) in the chamber and reaches a setpoint In this study the chamber was set to 02 O
2and 5
CO2in a humid atmosphere for 24 hours
25 Immunocytochemistry Cells were grown on gelatin cov-erslips (Nunc UK) at a density of 132 times 105 cells percm2 Immunocytochemistry was performed on normoxicand hypoxic (24 hours 02 O
2 5 CO
2) cells Cells were
washed twice with PBS fixed with 4 paraformaldehyde for15 minutes washed 3 times with PBS and washed with 01Triton X-100 3 times for 5 minutesThey were then incubatedwithNH
4Cl for 10minutes washed three times with PBS and
blocked using 5 goat serum (Dakocytomation Dako UK)for 45ndash60minutes at room temperature Cells were incubatedin Gab1 overnight at 4∘C washed three times incubatedin anti-rabbit conjugated with Alexa-488 (Invitrogen UK)for 1 hour at room temperature washed again and incu-bated with mouse antisarcomeric actin (Dakocytomation
BioMed Research International 3
Gab1
Acyanotic myocardium Cyanotic myocardium
Negative
control
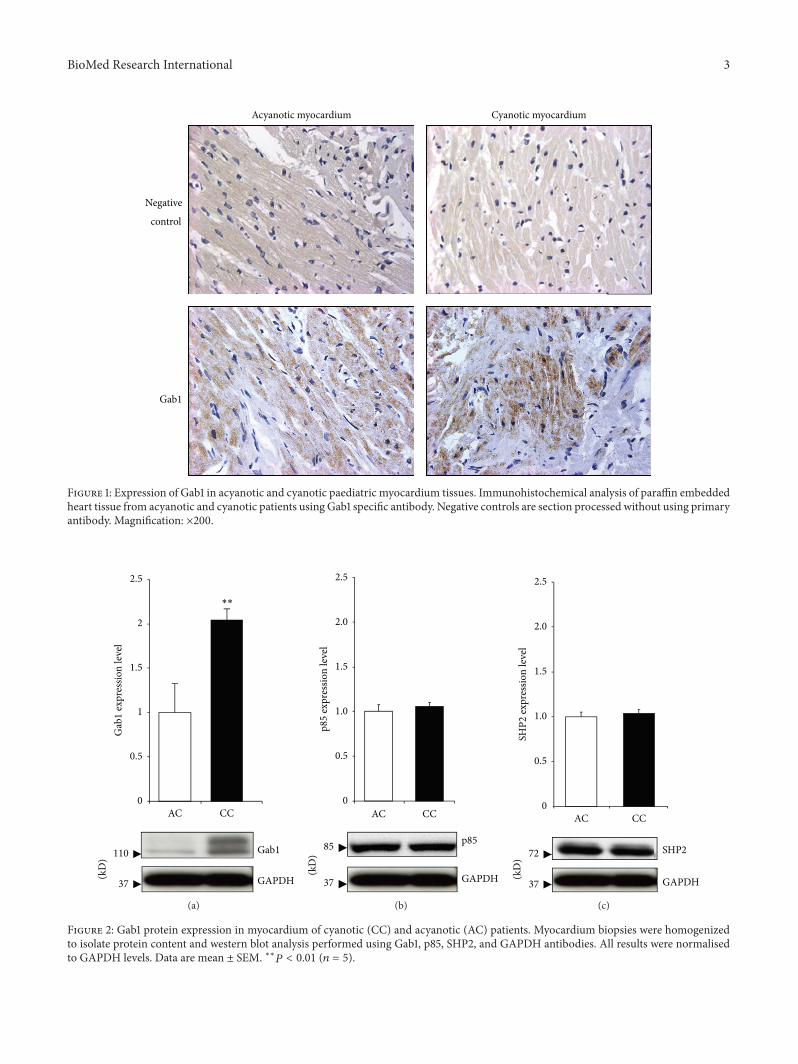
Figure 1 Expression of Gab1 in acyanotic and cyanotic paediatric myocardium tissues Immunohistochemical analysis of paraffin embeddedheart tissue from acyanotic and cyanotic patients using Gab1 specific antibody Negative controls are section processed without using primaryantibody Magnification times200
0
05
1
15
2
25
AC CC
Gab
1 ex
pres
sion
leve
l
110 Gab1
37 GAPDH(kD
)
lowastlowast
(a)
0
05
10
15
20
25
AC CC
p85
expr
essio
n le
vel
p8585
37 GAPDH(kD
)
(b)
0
05
10
15
20
25
AC CC
SHP2
expr
essio
n le
vel
SHP272
37 GAPDH(kD
)
(c)
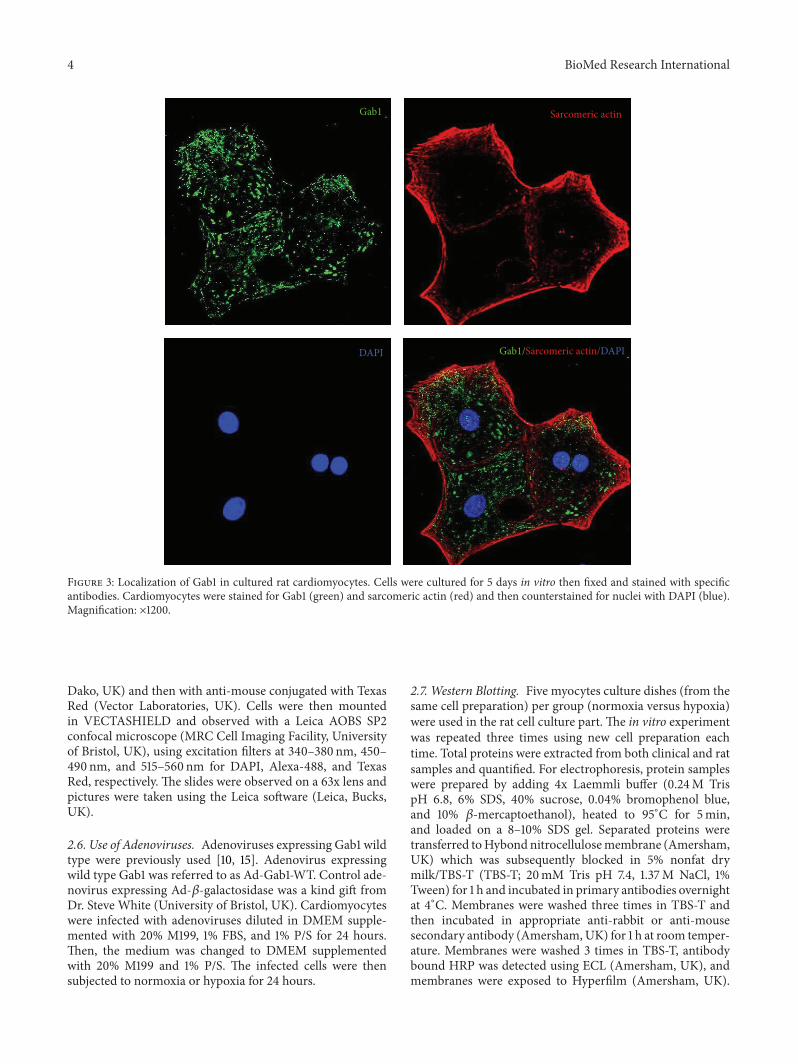
Figure 2 Gab1 protein expression in myocardium of cyanotic (CC) and acyanotic (AC) patients Myocardium biopsies were homogenizedto isolate protein content and western blot analysis performed using Gab1 p85 SHP2 and GAPDH antibodies All results were normalisedto GAPDH levels Data are mean plusmn SEM lowastlowast119875 lt 001 (119899 = 5)
4 BioMed Research International
DAPI
Gab1 Sarcomeric actin
Gab1Sarcomeric actinDAPI
Figure 3 Localization of Gab1 in cultured rat cardiomyocytes Cells were cultured for 5 days in vitro then fixed and stained with specificantibodies Cardiomyocytes were stained for Gab1 (green) and sarcomeric actin (red) and then counterstained for nuclei with DAPI (blue)Magnification times1200
Dako UK) and then with anti-mouse conjugated with TexasRed (Vector Laboratories UK) Cells were then mountedin VECTASHIELD and observed with a Leica AOBS SP2confocal microscope (MRC Cell Imaging Facility Universityof Bristol UK) using excitation filters at 340ndash380 nm 450ndash490 nm and 515ndash560 nm for DAPI Alexa-488 and TexasRed respectively The slides were observed on a 63x lens andpictures were taken using the Leica software (Leica BucksUK)
26 Use of Adenoviruses Adenoviruses expressing Gab1 wildtype were previously used [10 15] Adenovirus expressingwild type Gab1 was referred to as Ad-Gab1-WT Control ade-novirus expressing Ad-120573-galactosidase was a kind gift fromDr Steve White (University of Bristol UK) Cardiomyocyteswere infected with adenoviruses diluted in DMEM supple-mented with 20 M199 1 FBS and 1 PS for 24 hoursThen the medium was changed to DMEM supplementedwith 20 M199 and 1 PS The infected cells were thensubjected to normoxia or hypoxia for 24 hours
27 Western Blotting Five myocytes culture dishes (from thesame cell preparation) per group (normoxia versus hypoxia)were used in the rat cell culture part The in vitro experimentwas repeated three times using new cell preparation eachtime Total proteins were extracted from both clinical and ratsamples and quantified For electrophoresis protein sampleswere prepared by adding 4x Laemmli buffer (024M TrispH 68 6 SDS 40 sucrose 004 bromophenol blueand 10 120573-mercaptoethanol) heated to 95∘C for 5minand loaded on a 8ndash10 SDS gel Separated proteins weretransferred toHybond nitrocellulosemembrane (AmershamUK) which was subsequently blocked in 5 nonfat drymilkTBS-T (TBS-T 20mM Tris pH 74 137M NaCl 1Tween) for 1 h and incubated in primary antibodies overnightat 4∘C Membranes were washed three times in TBS-T andthen incubated in appropriate anti-rabbit or anti-mousesecondary antibody (AmershamUK) for 1 h at room temper-ature Membranes were washed 3 times in TBS-T antibodybound HRP was detected using ECL (Amersham UK) andmembranes were exposed to Hyperfilm (Amersham UK)
BioMed Research International 5
37 GAPDH(kD
) Gab1110
0
05
10
15
N H
Gab
1 ex
pres
sion
leve
l
lowast
(a)
(kD
)0
05
10
15
N H
SHP2
expr
essio
n le
vel
72
37
SHP2
GAPDH
lowast
(b)
(kD
) 80 p85
37 GAPDH
0
05
10
15
N H
p85
expr
essio
n le
vel
(c)
Figure 4 Gab1 protein expression in rat cardiomyocytes following 24 h of hypoxic treatment Cells were treated with hypoxic conditions (H)by incubating in a chamber with 5 CO
202 O
2or normoxia (N) by leaving cells in normal CO2 incubator After 24 h cells were lysed and
western blot analysis performed using GAB1 (a) SHP2 (b) and p85 (c) antibodies All results were normalized to GAPDH levels Data aremean plusmn SEM lowast119875 lt 005 (119899 = 5)
Protein bands were quantified using NIH Image J softwareThe expression level of each protein was normalized to theloading control GAPDH and then the obtained values forexperimental (cyanotic or hypoxic) were divided per controlvalue (acyanotic or normoxia) Representative blots werepresented in the western blotting figures
28 Apoptosis Assay Cell death was investigated with theldquoin situ cell death detectionrdquo kit from Roche Rat neona-tal cardiomyocytes were cultured in 4-well chamber slides(Nunc UK) at a density of 15 times 105 cells per cm2 Cellswere washed twice with PBS and permeabilized for 1 hourat room temperature with 01 Triton (vv) in 01 sodiumcitrate (vv) They were then washed three times in PBS andincubated with the labelled enzyme for 1 hour at 37∘C Theywere washed twice mounted in VECTASHIELD containingDAPI and observed with an Olympus B40 microscopeApoptosis was quantified by counting apoptotic cells in 25different fields or 400 cells Pictures were taken using aMediaCybernetics camera (Bethesda MD USA) and analysed withproimage plus software (Bethesda MD USA)
29 Statistical Analysis All data were analyzed using thesoftware Instat 31 (GraphPad) Results are expressed as plusmnstandard error of mean (plusmnSEM) Statistical significance wasassessed by one-way ANOVA or Studentrsquos t-test A value of119875 lt 005 was considered to be statistically significant
3 Results
31 Gab1 Protein Is Expressed in Human Heart Sections ofBoth Cyanotic and Acyanotic Congenital Heart Patients Theanalysis of fixed paediatric heart biopsy sections by immuno-histochemistry showed expression of GAB1 in heart tissuetaken from both acyanotic and cyanotic patients undergoingcorrective surgery for congenital heart defects (Figure 1)
32 Gab1 Protein Expression Is Upregulated in CyanoticPatients Compared to Acyanotic Western blot analysis ofproteins extracted from children myocardium biopsiesrevealed a significant upregulation ofGab1 protein expressionin cyanotic patients (Figure 2(a)) Additionally we investi-gated the protein expression of Gab binding partners SHP2and the regulatory subunit of PI3K (p85) The levels of bothGab binding partners proteins were unaffected (Figure 2(b))
33 Gab1 Shows a Cytoplasmic Localization in NeonatalHeart Cardiomyocytes Gab1 appeared to be expressed in ratneonatal cardiomyocytes (Figure 3) It is interesting to notethat Gab1 is locatedmainly in the cytoplasm of normoxic cellswith some nuclear expression
34 Hypoxia Produce a Downregulation of GAB1 in CulturedRat Cardiomyocytes As there is a limit to what can be doneusing human biopsies an in vitro model of cyanosis wouldbe advantageous Both GAB1 and SHP2 were downregulated
6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
In previous study we have shown that chronic hypoxiain pediatric patients with TOF triggered the expression ofnetwork of genes associated with apoptosis and reduced theexpression of genes involved in myocyte contractility andfunction [6] This state of hypoxia in TOF children maybe responsible for the susceptibility of cyanotic children toreoxygenation injury during and after surgery We have alsoshown that concomitant with the initiation of the injuriousprogram a protective program is triggered by cyanosis Gab1shown to significantly increase at messenger level in cyanoticcompared to acyanotic patients [6] could be an importantplayer in this protective program
Grb2 associated binding protein 1 (Gab1) is a memberof the scaffoldingdocking proteins (Gab1 Gab2 and Gab3)[7 8] Gab1 knockout mice are not viable and displayimpaired development of heart placenta skin and muscle[9] In cultured cardiomyocytes Gab1 is shown to interactwith tyrosine phosphatase SHP2 and to promote cardiachypertrophy [10] There is evidence that Gab1 is essentialfor cardiac function in the postnatal heart in vivo [11] Inaddition Gab1 has been shown to exert an antiapoptotic roleinmouse embryonic fibroblasts and is activated through tyro-sine phosphorylation following oxidative treatment (H
2O2)
[12] In their investigation Holgado-Madruga and Wongidentified Gab1 as an important component in oxidativestress signalling with an essential role in the activation ofc-Jun NH(2)-terminal kinase (JNK) and the influencing ofcell survival [12] This Gab1 antiapoptotic role in fibroblastsfollowing oxidative treatment [12] has led us to hypothesisethat Gab1 may play similar role in cardiac tissue and cardiacmyocytes subjected to hypoxia
In this study we investigated the effects of cyanosison Gab1 in myocardium samples from paediatric patientssuffering from TOF and we examined the effects of hypoxiain primary cultures of rat neonatal cardiomyocytes on Gab1and its possible role in cell survival
2 Materials and Methods
21 Reagents All reagents were from Sigma (UK) exceptthose stated otherwise Gab1 antibody was from MilliporeAntibodies against SHP2 and p85 were from Cell SignallingTechnology (UK) GAPDH antibody was from ResearchDiagnostics Inc (UK)
22 Cardiac Biopsies The collection of human right ventriclespecimens used in this study was approved by the NorthSomerset and South Bristol Research Ethics Committee(REC reference 07H0106172) the National Research EthicsService England Parental informed written consent wasgained for all patients Patients with a diagnosis of cyanotic(O2saturation 796plusmn 75 age 106plusmn 55months) or acyanotic
(O2saturation 942 plusmn 35 age 95 plusmn 23months) tetralogy of
Fallot undergoing surgical repair at the Bristol Royal Hospitalfor Children were studied
Five ventricular biopsy specimens per group were col-lected from the right ventricle of acyanotic and cyanoticTOF patients by using ldquoTrue-Cutrdquo needle immediately after
institution of cardiopulmonary bypass (CPB) Each specimenwas immediately put in liquid nitrogen for protein extraction
23 Immunohistochemistry Right ventricular specimenswere fixed in 4 paraformaldehyde washed in PBS andembedded in paraffin and 4120583msectionswere obtained Imm-unohistochemistry was performed using the ABC-Kit fromDakocytomation Slides were observed with anOlympus B40microscope Pictures were taken using a Media Cyberneticscamera (Bethesda MD USA) and analysed with proimageplus software (Bethesda MD USA)
24 RatNeonatal PrimaryCulture All the procedures involv-ing laboratory animals conformed to the guidelines and reg-ulations of the University of Bristol and the United KingdomHome Office Neonatal rat cardiomyocytes were cultured aspreviously described with minor modifications [13 14] Onecell culture preparation was used for each experiment andeach experiment was repeated three times using different cellcultures prepared at different time from pooled neonatal rathearts For each cell culture 14ndash24 neonatal rats were usedto harvest hearts Hearts were quickly removed from one-to three-day-old Wistar rats and only ventricles were keptThey were washed with PBS three times and incubated with005 trypsin and 002 EDTA for 30 minutes They werethen enzymatically digested six times for fifteen minutes in01 trypsin and 002 EDTA in PBS Digestion was stoppedby addition of foetal calf serum at a final concentration of30 Cells were then centrifuged at 400 g for fiveminutes andresuspended in DMEM supplemented with 10 FBS and 1PS Cells were then incubated one to two hours in a T75 flaskto allow noncardiac myocytes (mainly cardiac fibroblasts) toadhere to plastic They were then plated in gelatin-coatedplates at a density of 128 times 105 cells per cm2 After 40 hoursof culture themediumwas changed inDMEMsupplementedwith 20 M-199 and 1 PS All treatments were performedon three-day-old cultures Hypoxia was induced by placingthe cells in a hypoxic chamber where oxygen levels could bemonitored (Biospherix ProOx C21 Lacona NY USA) TheProOx is connected to a sensor which monitors the oxygenconcentration within the host chamber The ProOx quicklyinfuses nitrogen (hypoxia) in the chamber and reaches a setpoint In this study the chamber was set to 02 O
2and 5
CO2in a humid atmosphere for 24 hours
25 Immunocytochemistry Cells were grown on gelatin cov-erslips (Nunc UK) at a density of 132 times 105 cells percm2 Immunocytochemistry was performed on normoxicand hypoxic (24 hours 02 O
2 5 CO
2) cells Cells were
washed twice with PBS fixed with 4 paraformaldehyde for15 minutes washed 3 times with PBS and washed with 01Triton X-100 3 times for 5 minutesThey were then incubatedwithNH
4Cl for 10minutes washed three times with PBS and
blocked using 5 goat serum (Dakocytomation Dako UK)for 45ndash60minutes at room temperature Cells were incubatedin Gab1 overnight at 4∘C washed three times incubatedin anti-rabbit conjugated with Alexa-488 (Invitrogen UK)for 1 hour at room temperature washed again and incu-bated with mouse antisarcomeric actin (Dakocytomation
BioMed Research International 3
Gab1
Acyanotic myocardium Cyanotic myocardium
Negative
control
Figure 1 Expression of Gab1 in acyanotic and cyanotic paediatric myocardium tissues Immunohistochemical analysis of paraffin embeddedheart tissue from acyanotic and cyanotic patients using Gab1 specific antibody Negative controls are section processed without using primaryantibody Magnification times200
0
05
1
15
2
25
AC CC
Gab
1 ex
pres
sion
leve
l
110 Gab1
37 GAPDH(kD
)
lowastlowast
(a)
0
05
10
15
20
25
AC CC
p85
expr
essio
n le
vel
p8585
37 GAPDH(kD
)
(b)
0
05
10
15
20
25
AC CC
SHP2
expr
essio
n le
vel
SHP272
37 GAPDH(kD
)
(c)
Figure 2 Gab1 protein expression in myocardium of cyanotic (CC) and acyanotic (AC) patients Myocardium biopsies were homogenizedto isolate protein content and western blot analysis performed using Gab1 p85 SHP2 and GAPDH antibodies All results were normalisedto GAPDH levels Data are mean plusmn SEM lowastlowast119875 lt 001 (119899 = 5)
4 BioMed Research International
DAPI
Gab1 Sarcomeric actin
Gab1Sarcomeric actinDAPI
Figure 3 Localization of Gab1 in cultured rat cardiomyocytes Cells were cultured for 5 days in vitro then fixed and stained with specificantibodies Cardiomyocytes were stained for Gab1 (green) and sarcomeric actin (red) and then counterstained for nuclei with DAPI (blue)Magnification times1200
Dako UK) and then with anti-mouse conjugated with TexasRed (Vector Laboratories UK) Cells were then mountedin VECTASHIELD and observed with a Leica AOBS SP2confocal microscope (MRC Cell Imaging Facility Universityof Bristol UK) using excitation filters at 340ndash380 nm 450ndash490 nm and 515ndash560 nm for DAPI Alexa-488 and TexasRed respectively The slides were observed on a 63x lens andpictures were taken using the Leica software (Leica BucksUK)
26 Use of Adenoviruses Adenoviruses expressing Gab1 wildtype were previously used [10 15] Adenovirus expressingwild type Gab1 was referred to as Ad-Gab1-WT Control ade-novirus expressing Ad-120573-galactosidase was a kind gift fromDr Steve White (University of Bristol UK) Cardiomyocyteswere infected with adenoviruses diluted in DMEM supple-mented with 20 M199 1 FBS and 1 PS for 24 hoursThen the medium was changed to DMEM supplementedwith 20 M199 and 1 PS The infected cells were thensubjected to normoxia or hypoxia for 24 hours
27 Western Blotting Five myocytes culture dishes (from thesame cell preparation) per group (normoxia versus hypoxia)were used in the rat cell culture part The in vitro experimentwas repeated three times using new cell preparation eachtime Total proteins were extracted from both clinical and ratsamples and quantified For electrophoresis protein sampleswere prepared by adding 4x Laemmli buffer (024M TrispH 68 6 SDS 40 sucrose 004 bromophenol blueand 10 120573-mercaptoethanol) heated to 95∘C for 5minand loaded on a 8ndash10 SDS gel Separated proteins weretransferred toHybond nitrocellulosemembrane (AmershamUK) which was subsequently blocked in 5 nonfat drymilkTBS-T (TBS-T 20mM Tris pH 74 137M NaCl 1Tween) for 1 h and incubated in primary antibodies overnightat 4∘C Membranes were washed three times in TBS-T andthen incubated in appropriate anti-rabbit or anti-mousesecondary antibody (AmershamUK) for 1 h at room temper-ature Membranes were washed 3 times in TBS-T antibodybound HRP was detected using ECL (Amersham UK) andmembranes were exposed to Hyperfilm (Amersham UK)
BioMed Research International 5
37 GAPDH(kD
) Gab1110
0
05
10
15
N H
Gab
1 ex
pres
sion
leve
l
lowast
(a)
(kD
)0
05
10
15
N H
SHP2
expr
essio
n le
vel
72
37
SHP2
GAPDH
lowast
(b)
(kD
) 80 p85
37 GAPDH
0
05
10
15
N H
p85
expr
essio
n le
vel
(c)
Figure 4 Gab1 protein expression in rat cardiomyocytes following 24 h of hypoxic treatment Cells were treated with hypoxic conditions (H)by incubating in a chamber with 5 CO
202 O
2or normoxia (N) by leaving cells in normal CO2 incubator After 24 h cells were lysed and
western blot analysis performed using GAB1 (a) SHP2 (b) and p85 (c) antibodies All results were normalized to GAPDH levels Data aremean plusmn SEM lowast119875 lt 005 (119899 = 5)
Protein bands were quantified using NIH Image J softwareThe expression level of each protein was normalized to theloading control GAPDH and then the obtained values forexperimental (cyanotic or hypoxic) were divided per controlvalue (acyanotic or normoxia) Representative blots werepresented in the western blotting figures
28 Apoptosis Assay Cell death was investigated with theldquoin situ cell death detectionrdquo kit from Roche Rat neona-tal cardiomyocytes were cultured in 4-well chamber slides(Nunc UK) at a density of 15 times 105 cells per cm2 Cellswere washed twice with PBS and permeabilized for 1 hourat room temperature with 01 Triton (vv) in 01 sodiumcitrate (vv) They were then washed three times in PBS andincubated with the labelled enzyme for 1 hour at 37∘C Theywere washed twice mounted in VECTASHIELD containingDAPI and observed with an Olympus B40 microscopeApoptosis was quantified by counting apoptotic cells in 25different fields or 400 cells Pictures were taken using aMediaCybernetics camera (Bethesda MD USA) and analysed withproimage plus software (Bethesda MD USA)
29 Statistical Analysis All data were analyzed using thesoftware Instat 31 (GraphPad) Results are expressed as plusmnstandard error of mean (plusmnSEM) Statistical significance wasassessed by one-way ANOVA or Studentrsquos t-test A value of119875 lt 005 was considered to be statistically significant
3 Results
31 Gab1 Protein Is Expressed in Human Heart Sections ofBoth Cyanotic and Acyanotic Congenital Heart Patients Theanalysis of fixed paediatric heart biopsy sections by immuno-histochemistry showed expression of GAB1 in heart tissuetaken from both acyanotic and cyanotic patients undergoingcorrective surgery for congenital heart defects (Figure 1)
32 Gab1 Protein Expression Is Upregulated in CyanoticPatients Compared to Acyanotic Western blot analysis ofproteins extracted from children myocardium biopsiesrevealed a significant upregulation ofGab1 protein expressionin cyanotic patients (Figure 2(a)) Additionally we investi-gated the protein expression of Gab binding partners SHP2and the regulatory subunit of PI3K (p85) The levels of bothGab binding partners proteins were unaffected (Figure 2(b))
33 Gab1 Shows a Cytoplasmic Localization in NeonatalHeart Cardiomyocytes Gab1 appeared to be expressed in ratneonatal cardiomyocytes (Figure 3) It is interesting to notethat Gab1 is locatedmainly in the cytoplasm of normoxic cellswith some nuclear expression
34 Hypoxia Produce a Downregulation of GAB1 in CulturedRat Cardiomyocytes As there is a limit to what can be doneusing human biopsies an in vitro model of cyanosis wouldbe advantageous Both GAB1 and SHP2 were downregulated
6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
BioMed Research International 3
Gab1
Acyanotic myocardium Cyanotic myocardium
Negative
control
Figure 1 Expression of Gab1 in acyanotic and cyanotic paediatric myocardium tissues Immunohistochemical analysis of paraffin embeddedheart tissue from acyanotic and cyanotic patients using Gab1 specific antibody Negative controls are section processed without using primaryantibody Magnification times200
0
05
1
15
2
25
AC CC
Gab
1 ex
pres
sion
leve
l
110 Gab1
37 GAPDH(kD
)
lowastlowast
(a)
0
05
10
15
20
25
AC CC
p85
expr
essio
n le
vel
p8585
37 GAPDH(kD
)
(b)
0
05
10
15
20
25
AC CC
SHP2
expr
essio
n le
vel
SHP272
37 GAPDH(kD
)
(c)
Figure 2 Gab1 protein expression in myocardium of cyanotic (CC) and acyanotic (AC) patients Myocardium biopsies were homogenizedto isolate protein content and western blot analysis performed using Gab1 p85 SHP2 and GAPDH antibodies All results were normalisedto GAPDH levels Data are mean plusmn SEM lowastlowast119875 lt 001 (119899 = 5)
4 BioMed Research International
DAPI
Gab1 Sarcomeric actin
Gab1Sarcomeric actinDAPI
Figure 3 Localization of Gab1 in cultured rat cardiomyocytes Cells were cultured for 5 days in vitro then fixed and stained with specificantibodies Cardiomyocytes were stained for Gab1 (green) and sarcomeric actin (red) and then counterstained for nuclei with DAPI (blue)Magnification times1200
Dako UK) and then with anti-mouse conjugated with TexasRed (Vector Laboratories UK) Cells were then mountedin VECTASHIELD and observed with a Leica AOBS SP2confocal microscope (MRC Cell Imaging Facility Universityof Bristol UK) using excitation filters at 340ndash380 nm 450ndash490 nm and 515ndash560 nm for DAPI Alexa-488 and TexasRed respectively The slides were observed on a 63x lens andpictures were taken using the Leica software (Leica BucksUK)
26 Use of Adenoviruses Adenoviruses expressing Gab1 wildtype were previously used [10 15] Adenovirus expressingwild type Gab1 was referred to as Ad-Gab1-WT Control ade-novirus expressing Ad-120573-galactosidase was a kind gift fromDr Steve White (University of Bristol UK) Cardiomyocyteswere infected with adenoviruses diluted in DMEM supple-mented with 20 M199 1 FBS and 1 PS for 24 hoursThen the medium was changed to DMEM supplementedwith 20 M199 and 1 PS The infected cells were thensubjected to normoxia or hypoxia for 24 hours
27 Western Blotting Five myocytes culture dishes (from thesame cell preparation) per group (normoxia versus hypoxia)were used in the rat cell culture part The in vitro experimentwas repeated three times using new cell preparation eachtime Total proteins were extracted from both clinical and ratsamples and quantified For electrophoresis protein sampleswere prepared by adding 4x Laemmli buffer (024M TrispH 68 6 SDS 40 sucrose 004 bromophenol blueand 10 120573-mercaptoethanol) heated to 95∘C for 5minand loaded on a 8ndash10 SDS gel Separated proteins weretransferred toHybond nitrocellulosemembrane (AmershamUK) which was subsequently blocked in 5 nonfat drymilkTBS-T (TBS-T 20mM Tris pH 74 137M NaCl 1Tween) for 1 h and incubated in primary antibodies overnightat 4∘C Membranes were washed three times in TBS-T andthen incubated in appropriate anti-rabbit or anti-mousesecondary antibody (AmershamUK) for 1 h at room temper-ature Membranes were washed 3 times in TBS-T antibodybound HRP was detected using ECL (Amersham UK) andmembranes were exposed to Hyperfilm (Amersham UK)
BioMed Research International 5
37 GAPDH(kD
) Gab1110
0
05
10
15
N H
Gab
1 ex
pres
sion
leve
l
lowast
(a)
(kD
)0
05
10
15
N H
SHP2
expr
essio
n le
vel
72
37
SHP2
GAPDH
lowast
(b)
(kD
) 80 p85
37 GAPDH
0
05
10
15
N H
p85
expr
essio
n le
vel
(c)
Figure 4 Gab1 protein expression in rat cardiomyocytes following 24 h of hypoxic treatment Cells were treated with hypoxic conditions (H)by incubating in a chamber with 5 CO
202 O
2or normoxia (N) by leaving cells in normal CO2 incubator After 24 h cells were lysed and
western blot analysis performed using GAB1 (a) SHP2 (b) and p85 (c) antibodies All results were normalized to GAPDH levels Data aremean plusmn SEM lowast119875 lt 005 (119899 = 5)
Protein bands were quantified using NIH Image J softwareThe expression level of each protein was normalized to theloading control GAPDH and then the obtained values forexperimental (cyanotic or hypoxic) were divided per controlvalue (acyanotic or normoxia) Representative blots werepresented in the western blotting figures
28 Apoptosis Assay Cell death was investigated with theldquoin situ cell death detectionrdquo kit from Roche Rat neona-tal cardiomyocytes were cultured in 4-well chamber slides(Nunc UK) at a density of 15 times 105 cells per cm2 Cellswere washed twice with PBS and permeabilized for 1 hourat room temperature with 01 Triton (vv) in 01 sodiumcitrate (vv) They were then washed three times in PBS andincubated with the labelled enzyme for 1 hour at 37∘C Theywere washed twice mounted in VECTASHIELD containingDAPI and observed with an Olympus B40 microscopeApoptosis was quantified by counting apoptotic cells in 25different fields or 400 cells Pictures were taken using aMediaCybernetics camera (Bethesda MD USA) and analysed withproimage plus software (Bethesda MD USA)
29 Statistical Analysis All data were analyzed using thesoftware Instat 31 (GraphPad) Results are expressed as plusmnstandard error of mean (plusmnSEM) Statistical significance wasassessed by one-way ANOVA or Studentrsquos t-test A value of119875 lt 005 was considered to be statistically significant
3 Results
31 Gab1 Protein Is Expressed in Human Heart Sections ofBoth Cyanotic and Acyanotic Congenital Heart Patients Theanalysis of fixed paediatric heart biopsy sections by immuno-histochemistry showed expression of GAB1 in heart tissuetaken from both acyanotic and cyanotic patients undergoingcorrective surgery for congenital heart defects (Figure 1)
32 Gab1 Protein Expression Is Upregulated in CyanoticPatients Compared to Acyanotic Western blot analysis ofproteins extracted from children myocardium biopsiesrevealed a significant upregulation ofGab1 protein expressionin cyanotic patients (Figure 2(a)) Additionally we investi-gated the protein expression of Gab binding partners SHP2and the regulatory subunit of PI3K (p85) The levels of bothGab binding partners proteins were unaffected (Figure 2(b))
33 Gab1 Shows a Cytoplasmic Localization in NeonatalHeart Cardiomyocytes Gab1 appeared to be expressed in ratneonatal cardiomyocytes (Figure 3) It is interesting to notethat Gab1 is locatedmainly in the cytoplasm of normoxic cellswith some nuclear expression
34 Hypoxia Produce a Downregulation of GAB1 in CulturedRat Cardiomyocytes As there is a limit to what can be doneusing human biopsies an in vitro model of cyanosis wouldbe advantageous Both GAB1 and SHP2 were downregulated
6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
4 BioMed Research International
DAPI
Gab1 Sarcomeric actin
Gab1Sarcomeric actinDAPI
Figure 3 Localization of Gab1 in cultured rat cardiomyocytes Cells were cultured for 5 days in vitro then fixed and stained with specificantibodies Cardiomyocytes were stained for Gab1 (green) and sarcomeric actin (red) and then counterstained for nuclei with DAPI (blue)Magnification times1200
Dako UK) and then with anti-mouse conjugated with TexasRed (Vector Laboratories UK) Cells were then mountedin VECTASHIELD and observed with a Leica AOBS SP2confocal microscope (MRC Cell Imaging Facility Universityof Bristol UK) using excitation filters at 340ndash380 nm 450ndash490 nm and 515ndash560 nm for DAPI Alexa-488 and TexasRed respectively The slides were observed on a 63x lens andpictures were taken using the Leica software (Leica BucksUK)
26 Use of Adenoviruses Adenoviruses expressing Gab1 wildtype were previously used [10 15] Adenovirus expressingwild type Gab1 was referred to as Ad-Gab1-WT Control ade-novirus expressing Ad-120573-galactosidase was a kind gift fromDr Steve White (University of Bristol UK) Cardiomyocyteswere infected with adenoviruses diluted in DMEM supple-mented with 20 M199 1 FBS and 1 PS for 24 hoursThen the medium was changed to DMEM supplementedwith 20 M199 and 1 PS The infected cells were thensubjected to normoxia or hypoxia for 24 hours
27 Western Blotting Five myocytes culture dishes (from thesame cell preparation) per group (normoxia versus hypoxia)were used in the rat cell culture part The in vitro experimentwas repeated three times using new cell preparation eachtime Total proteins were extracted from both clinical and ratsamples and quantified For electrophoresis protein sampleswere prepared by adding 4x Laemmli buffer (024M TrispH 68 6 SDS 40 sucrose 004 bromophenol blueand 10 120573-mercaptoethanol) heated to 95∘C for 5minand loaded on a 8ndash10 SDS gel Separated proteins weretransferred toHybond nitrocellulosemembrane (AmershamUK) which was subsequently blocked in 5 nonfat drymilkTBS-T (TBS-T 20mM Tris pH 74 137M NaCl 1Tween) for 1 h and incubated in primary antibodies overnightat 4∘C Membranes were washed three times in TBS-T andthen incubated in appropriate anti-rabbit or anti-mousesecondary antibody (AmershamUK) for 1 h at room temper-ature Membranes were washed 3 times in TBS-T antibodybound HRP was detected using ECL (Amersham UK) andmembranes were exposed to Hyperfilm (Amersham UK)
BioMed Research International 5
37 GAPDH(kD
) Gab1110
0
05
10
15
N H
Gab
1 ex
pres
sion
leve
l
lowast
(a)
(kD
)0
05
10
15
N H
SHP2
expr
essio
n le
vel
72
37
SHP2
GAPDH
lowast
(b)
(kD
) 80 p85
37 GAPDH
0
05
10
15
N H
p85
expr
essio
n le
vel
(c)
Figure 4 Gab1 protein expression in rat cardiomyocytes following 24 h of hypoxic treatment Cells were treated with hypoxic conditions (H)by incubating in a chamber with 5 CO
202 O
2or normoxia (N) by leaving cells in normal CO2 incubator After 24 h cells were lysed and
western blot analysis performed using GAB1 (a) SHP2 (b) and p85 (c) antibodies All results were normalized to GAPDH levels Data aremean plusmn SEM lowast119875 lt 005 (119899 = 5)
Protein bands were quantified using NIH Image J softwareThe expression level of each protein was normalized to theloading control GAPDH and then the obtained values forexperimental (cyanotic or hypoxic) were divided per controlvalue (acyanotic or normoxia) Representative blots werepresented in the western blotting figures
28 Apoptosis Assay Cell death was investigated with theldquoin situ cell death detectionrdquo kit from Roche Rat neona-tal cardiomyocytes were cultured in 4-well chamber slides(Nunc UK) at a density of 15 times 105 cells per cm2 Cellswere washed twice with PBS and permeabilized for 1 hourat room temperature with 01 Triton (vv) in 01 sodiumcitrate (vv) They were then washed three times in PBS andincubated with the labelled enzyme for 1 hour at 37∘C Theywere washed twice mounted in VECTASHIELD containingDAPI and observed with an Olympus B40 microscopeApoptosis was quantified by counting apoptotic cells in 25different fields or 400 cells Pictures were taken using aMediaCybernetics camera (Bethesda MD USA) and analysed withproimage plus software (Bethesda MD USA)
29 Statistical Analysis All data were analyzed using thesoftware Instat 31 (GraphPad) Results are expressed as plusmnstandard error of mean (plusmnSEM) Statistical significance wasassessed by one-way ANOVA or Studentrsquos t-test A value of119875 lt 005 was considered to be statistically significant
3 Results
31 Gab1 Protein Is Expressed in Human Heart Sections ofBoth Cyanotic and Acyanotic Congenital Heart Patients Theanalysis of fixed paediatric heart biopsy sections by immuno-histochemistry showed expression of GAB1 in heart tissuetaken from both acyanotic and cyanotic patients undergoingcorrective surgery for congenital heart defects (Figure 1)
32 Gab1 Protein Expression Is Upregulated in CyanoticPatients Compared to Acyanotic Western blot analysis ofproteins extracted from children myocardium biopsiesrevealed a significant upregulation ofGab1 protein expressionin cyanotic patients (Figure 2(a)) Additionally we investi-gated the protein expression of Gab binding partners SHP2and the regulatory subunit of PI3K (p85) The levels of bothGab binding partners proteins were unaffected (Figure 2(b))
33 Gab1 Shows a Cytoplasmic Localization in NeonatalHeart Cardiomyocytes Gab1 appeared to be expressed in ratneonatal cardiomyocytes (Figure 3) It is interesting to notethat Gab1 is locatedmainly in the cytoplasm of normoxic cellswith some nuclear expression
34 Hypoxia Produce a Downregulation of GAB1 in CulturedRat Cardiomyocytes As there is a limit to what can be doneusing human biopsies an in vitro model of cyanosis wouldbe advantageous Both GAB1 and SHP2 were downregulated
6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
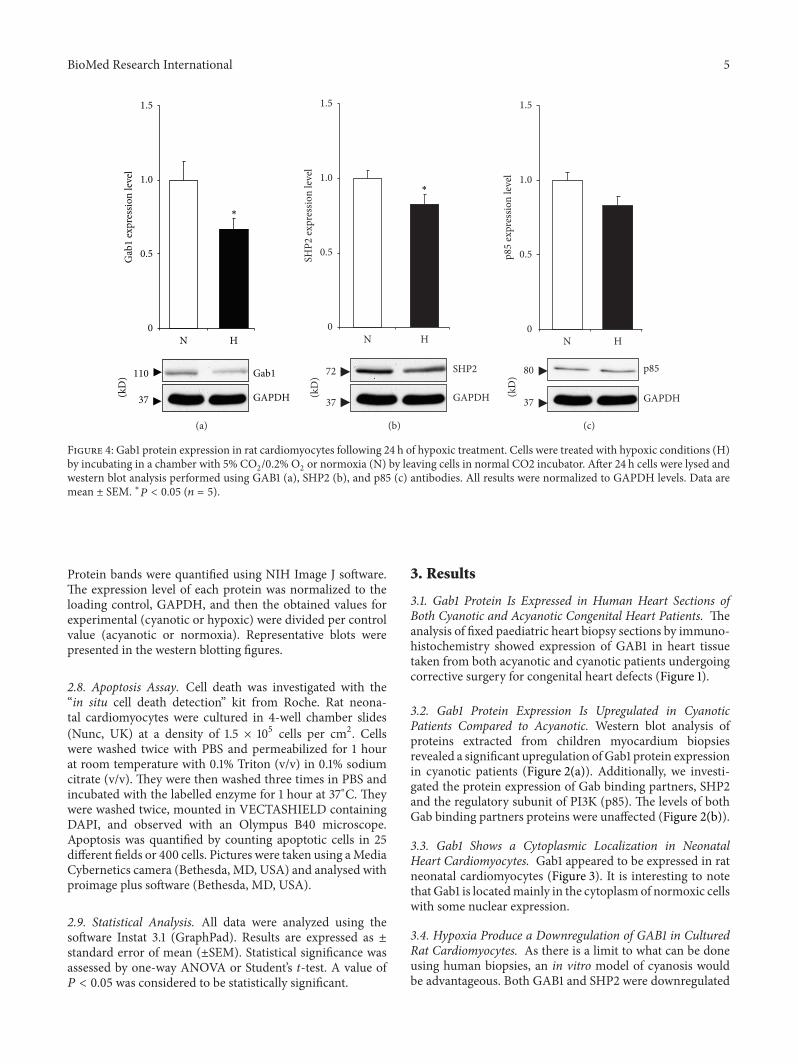
BioMed Research International 5
37 GAPDH(kD
) Gab1110
0
05
10
15
N H
Gab
1 ex
pres
sion
leve
l
lowast
(a)
(kD
)0
05
10
15
N H
SHP2
expr
essio
n le
vel
72
37
SHP2
GAPDH
lowast
(b)
(kD
) 80 p85
37 GAPDH
0
05
10
15
N H
p85
expr
essio
n le
vel
(c)
Figure 4 Gab1 protein expression in rat cardiomyocytes following 24 h of hypoxic treatment Cells were treated with hypoxic conditions (H)by incubating in a chamber with 5 CO
202 O
2or normoxia (N) by leaving cells in normal CO2 incubator After 24 h cells were lysed and
western blot analysis performed using GAB1 (a) SHP2 (b) and p85 (c) antibodies All results were normalized to GAPDH levels Data aremean plusmn SEM lowast119875 lt 005 (119899 = 5)
Protein bands were quantified using NIH Image J softwareThe expression level of each protein was normalized to theloading control GAPDH and then the obtained values forexperimental (cyanotic or hypoxic) were divided per controlvalue (acyanotic or normoxia) Representative blots werepresented in the western blotting figures
28 Apoptosis Assay Cell death was investigated with theldquoin situ cell death detectionrdquo kit from Roche Rat neona-tal cardiomyocytes were cultured in 4-well chamber slides(Nunc UK) at a density of 15 times 105 cells per cm2 Cellswere washed twice with PBS and permeabilized for 1 hourat room temperature with 01 Triton (vv) in 01 sodiumcitrate (vv) They were then washed three times in PBS andincubated with the labelled enzyme for 1 hour at 37∘C Theywere washed twice mounted in VECTASHIELD containingDAPI and observed with an Olympus B40 microscopeApoptosis was quantified by counting apoptotic cells in 25different fields or 400 cells Pictures were taken using aMediaCybernetics camera (Bethesda MD USA) and analysed withproimage plus software (Bethesda MD USA)
29 Statistical Analysis All data were analyzed using thesoftware Instat 31 (GraphPad) Results are expressed as plusmnstandard error of mean (plusmnSEM) Statistical significance wasassessed by one-way ANOVA or Studentrsquos t-test A value of119875 lt 005 was considered to be statistically significant
3 Results
31 Gab1 Protein Is Expressed in Human Heart Sections ofBoth Cyanotic and Acyanotic Congenital Heart Patients Theanalysis of fixed paediatric heart biopsy sections by immuno-histochemistry showed expression of GAB1 in heart tissuetaken from both acyanotic and cyanotic patients undergoingcorrective surgery for congenital heart defects (Figure 1)
32 Gab1 Protein Expression Is Upregulated in CyanoticPatients Compared to Acyanotic Western blot analysis ofproteins extracted from children myocardium biopsiesrevealed a significant upregulation ofGab1 protein expressionin cyanotic patients (Figure 2(a)) Additionally we investi-gated the protein expression of Gab binding partners SHP2and the regulatory subunit of PI3K (p85) The levels of bothGab binding partners proteins were unaffected (Figure 2(b))
33 Gab1 Shows a Cytoplasmic Localization in NeonatalHeart Cardiomyocytes Gab1 appeared to be expressed in ratneonatal cardiomyocytes (Figure 3) It is interesting to notethat Gab1 is locatedmainly in the cytoplasm of normoxic cellswith some nuclear expression
34 Hypoxia Produce a Downregulation of GAB1 in CulturedRat Cardiomyocytes As there is a limit to what can be doneusing human biopsies an in vitro model of cyanosis wouldbe advantageous Both GAB1 and SHP2 were downregulated
6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 BioMed Research International
SHP2
GAPDH
p85
Gab1
(kD
)
110
80
72
37
No
viru
s
Ad-G
ab1
-WT10
MO
I
Ad-120573
-Gal50
MO
I
(a)Ad-Gab1-WT
Apop
totic
cells
()
0
5
10
15
Normoxia Hypoxia
lowast
Ad-120573-Gal
(b)
Normoxia
Ad-Gab1-WT
DAPI TUNEL
Hypoxia
DAPI TUNEL
Ad-120573-Gal
(c)
Figure 5 (a) Protein expression level of Gab1 p85 and SHP2 in infected rat neonatal cardiomyocytes Cardiac myocytes were infectedwith Ad-120573-Gal (50 MOI) and Ad-Gab1-WT (10 MOI) for 24 hours GAPDH was used as a loading control (b) Apoptosis quantification ofinfected rat neonatal cardiomyocytes subjected to normoxia and hypoxia Cardiac myocytes were infected with Ad-120573-galactosidase (Ad-120573-Gal) and Ad-Gab1-WT for 24 hours Data are mean plusmn SEM lowast119875 lt 005 (c) Representative pictures of the TUNEL assay performed on ratneonatal cardiomyocytes infected by Ad-120573-Gal or Ad-Gab1-WT and subjected to normoxia or hypoxia Arrows show apoptotic nuclei (greenfluorescence) Cell nuclei were stained by DAPI (blue fluorescence) Magnification times400
at the protein level following hypoxia when compared to thenormoxic control (Figures 4(a) and 4(b)) However hypoxiadid not affect p85 protein expression (Figure 4(c))
35 Gab1 Overexpression Reduces Apoptosis during Hypoxiain Rat Cardiomyocytes We first examined the efficiency ofcell infection by Ad-Gab1-WT (Figure 5(a)) Infection of ratneonatal cardiomyocytes with Ad-Gab1-WT was successfulas demonstrated by the increase of Gab1 protein expressioncompared to cells infected with Ad-120573-Gal (Figure 5(a)) Inaddition the infection with both viruses did not affect theprotein expression of p85 SHP2 and GAPDH
During normoxia the overexpression of Gab1 in car-diomyocytes did not alter the percentage of apoptotic cellssignificantly However during hypoxia Gab1 overexpressionreduced significantly apoptosis in rat neonatal cardiomy-ocytes (Figures 5(b) and 5(c))
4 Discussion
Our study revealed a significant upregulation of Gab1 proteinexpression in cyanotic TOF patients This result confirmedour previous findings by microarray analysis [6] The upreg-ulation of Gab1 protein expression in cyanotic patients may
BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 7
suggest an increase of survival signalling mediated throughGab1 in cyanotic patients independently of SHP2 and p85These data are the first to implicate Gab1 in cardioprotectivesignalling in cyanotic patients in response to chronic hypoxiastress
Gab1 protein levels were downregulated during hypoxiain neonatal rat cardiomyocytesThese are also the first resultsto implicate Gab1 in the cardiomyocytes response to hypoxiaThe difference between the in vivo and in vitro data regardingthe response to hypoxia can be explained by the complexity ofthe in vivo situation in comparison to the relative simplicityof the in vitro model Furthermore it can be explainedby the short time of hypoxia protocol (24 h) used for ratmyocytes compared to the patients that stayed cyanotic formonths before surgery Additionally this may be attributedto the difference between TOF patientsrsquo tissue specimens andneonatal rat ventricle Neonatal rat ventricle includes LV andRV tissues whereas TOF patientsrsquo tissue was only from RVIt is well known that there is a difference in cardiac tissuebetween the two ventricles [16]
We have previously shown that chronic hypoxia inducesboth cytoprotective and injury related transcriptomic repro-gramming [6]The protective program induces survival path-ways and the deleterious program triggers cell death signaling[6] Any imbalance between these two programs would resultin either cell survival or death Therefore modulating thebalance between these two programs offers the potential todevelop strategies for cardioprotectionOur data suggest thatin cyanotic pediatric heart the increase in Gab1 expressionis part of the survival pathway Additionally the reductionof apoptosis observed following the overexpression of Gab1suggests a critical and prosurvival role for Gab1 in ratneonatal cardiomyocytesThis is in line with previous reportsdescribing an antiapoptotic function for Gab1 [12] It has alsobeen shown that SHP2 can help to promote cell survival bythe activation of the RafMEKERK signalling pathway [17]
Gab1 has been shown to play an antiapoptotic role inoxidative condition [12] It would be interesting to see howGab1 expression levels would be following corrective surgeryfor cyanotic congenital heart disease Similarly it would beinteresting to examine the removal of hypoxic stress in anexperimental model One can speculate that the removal ofhypoxic stress would result in the return of Gab1 expressionto normoxic condition levels Additionally we would expectthatan abrupt reoxygenation could modulate the levels ofthis unique protein Furthermore the normal transition fromfoetal to neonatal circulation may affect Gab1 expressionlevels
Gab1 expression changes following hypoxia may be con-troversial considering the observed difference between thein vivo and in vitro situations However it has previouslybeen shown that oxygen availability can play a critical rolein defining the cellular responses to stimuli [18] Comparedto the cell culture system the in vivo situation adds anotherlevel of complexity It is likely that other in vivo signallingpathways come into play that could result in a differentresponse to hypoxia as compared to culture system In theabove-mentioned study it has been shown that whereasIGF signaling promotes muscle cell differentiation under
normoxia it stimulates proliferation under hypoxia by dif-ferentially regulating multiple signaling pathways [18] Apossible mechanism involved in modulating Gab1 expressioncould be the HIF1 alpha-signaling pathway
SHP2 and p85 expressions showed no differencebetween cyanotic and acyanotic TOF patients however theydecreased although it was not significant for p85 in hypoxicmyocytes A plausible explanation is the difference betweenRV tissue collected from TOF patients and the mixed cellpopulation harvested from LV and RV of neonatal ratsThere is evidence that cardiac tissue obtained from the twoventricles has different expression profile and can responddifferently to stimuli [16]
Altogether our data provide strong evidence that Gab1is important for cardiomyocytes survival following hypoxicstress Gab1 represents a potential target for cardioprotection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors would like to thank Dr Amir Mokhtari and DrAndrew Parry for their surgical involvement This researchwas funded by the BritishHeart Foundation GarfieldWestonTrust andNIHRBristol Biomedical ResearchUnit inCardio-vascular Medicine Mohamed T Ghorbel was supported byan Intermediate Research Fellowship from the British HeartFoundation
References
[1] E A Shinebourne S V Babu-Narayan and J S CarvalholdquoTetralogy of fallot from fetus to adultrdquo Heart vol 92 no 9pp 1353ndash1359 2006
[2] S Bharati and M Lev The Pathology of Congenital Heart Dis-ease A Personal Experience with More than 6300 CongenitallyMalformed Hearts Futura Armonk NY USA 1996
[3] F Bailliard and R H Anderson ldquoTetralogy of FallotrdquoOrphanetJournal of Rare Diseases vol 4 no 1 article 2 2009
[4] K H Teoh D A G Mickle R D Weisel et al ldquoEffect ofoxygen tension and cardiovascular operations on the myocar-dial antioxidant enzyme activities in patients with tetralogyof Fallot and aorta- coronary bypassrdquo Journal of Thoracic andCardiovascular Surgery vol 104 no 1 pp 159ndash164 1992
[5] K J Davies ldquoOxidative stress the paradox of aerobic liferdquo inProceedings of the Biochemical Society Symposia vol 61 pp 1ndash31 1995
[6] M T Ghorbel M Cherif E Jenkins et al ldquoTranscriptomicanalysis of patients with tetralogy of Fallot reveals the effect ofchronic hypoxia on myocardial gene expressionrdquoThe Journal ofThoracic andCardiovascular Surgery vol 140 no 2 pp 337e26ndash345e26 2010
[7] M Hibi and T Hirano ldquoGab-family adapter molecules in signaltransduction of cytokine and growth factor receptors andT andB cell antigen receptorsrdquo Leukemia and Lymphoma vol 37 no3-4 pp 299ndash307 2000
8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
[8] Y Liu and L R Rohrschneider ldquoThe gift of Gabrdquo FEBS Lettersvol 515 no 1ndash3 pp 1ndash7 2002
[9] M Itoh Y Yoshida K Nishida M Narimatsu M Hibi and THirano ldquoRole of Gab l in heart placenta and skin developmentand growth factor- and cytokine-induced extracellular signal-regulated kinase mitogen-activated protein kinase activationrdquoMolecular and Cellular Biology vol 20 no 10 pp 3695ndash37042000
[10] Y Nakaoka K Nishida Y Fujio et al ldquoActivation of gp130transduces hypertrophic signal through interaction of scaffold-ingdocking protein Gab1 with tyrosine phosphatase SHP2 incardiomyocytesrdquo Circulation Research vol 93 no 3 pp 221ndash229 2003
[11] Y Nakaoka K Nishida M Narimatsu et al ldquoGab familyproteins are essential for postnatalmaintenance of cardiac func-tion via neuregulin-1ErbB signalingrdquo The Journal of ClinicalInvestigation vol 117 no 7 pp 1771ndash1781 2007
[12] M Holgado-Madruga and A J Wong ldquoGab1 is an integratorof cell death versus cell survival signals in oxidative stressrdquoMolecular and Cellular Biology vol 23 no 13 pp 4471ndash44842003
[13] S Matoba T Tatsumi N Keira et al ldquoCardioprotectiveeffect of angiotensin-converting enzyme inhibition againsthypoxiareoxygenation injury in cultured rat cardiacmyocytesrdquoCirculation vol 99 no 6 pp 817ndash822 1999
[14] G B Sala-Newby N V E Freeman M A Curto and A CNewby ldquoMetabolic and functional consequences of cytosolic51015840 -nucleotidase-IA overexpression in neonatal rat cardiomy-ocytesrdquo American Journal of Physiology Heart and CirculatoryPhysiology vol 285 no 3 pp H991ndashH998 2003
[15] T Koyama Y Nakaoka Y Fujio et al ldquoInteraction of scaf-folding adaptor protein Gab1 with tyrosine phosphatase SHP2negatively regulates IGF-I-dependent myogenic differentiationvia the ERK12 signaling pathwayrdquo The Journal of BiologicalChemistry vol 283 no 35 pp 24234ndash24244 2008
[16] B Littlejohns K Heesom G D Angelini and M S SuleimanldquoThe effect of disease on human cardiac protein expressionprofiles in paired samples from right and left ventriclesrdquoClinicalProteomics vol 11 no 1 article 34 2014
[17] K Nishida and T Hirano ldquoThe role of Gab family scaffoldingadapter proteins in the signal transduction of cytokine andgrowth factor receptorsrdquo Cancer Science vol 94 no 12 pp1029ndash1033 2003
[18] H RenDAccili andCDuan ldquoHypoxia converts themyogenicaction of insulin-like growth factors into mitogenic action bydifferentially regulating multiple signaling pathwaysrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 107 no 13 pp 5857ndash5862 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom