renal lysozyme levels animals developing sterile ...gressive chronic pyelonephritis has a bacterial...
TRANSCRIPT

INFECTION AND IMMUNITY, Sept. 1971, p. 269-273Copyright ( 1971 American Society for Microbiology
Vol. 4, No. 3Prinzted in U.S.A
Renal Lysozyme Levels in Animals Developing"Sterile Pyelonephritis"
W. W. EUDY, S. E. BURROUS, AND F. W. SIGLER
Research and Developmenit Department, The Norwich Phlarmacal Company, Norwich, New York 13815
Received for publication 4 May 1971
The induction of sterile unilateral pyelonephritis in rats with heat-killed Proteusmirabilis cells is described. The lesions were identical to those produced with viablebacteria. Lysozyme levels in both kidneys of rats developing unilateral sterile pyelo-nephritis underwent biphasic elevations similar to those produced with viable bac-teria. In the injected kidney, the first elevation, associated with the trauma of injec-tion, could be produced by injecting sterile saline. The second elevation was asso-ciated with the onset of chronicity in the injected kidney. The nonmanipulated,contralateral kidney showed a similar biphasic elevation, of equal duration but ofgreater magnitude.
Previous work in our laboratory indicated thattrauma to a kidney causes an increase in its solu-ble lysozyme content (10). This observation agreeswith the results ofHamdy (11), who demonstratedthat trauma causes elevated levels of solublelysozyme in poultry tissue. However, in unilateralrenal trauma, a simultaneous elevation of thesoluble lysozyme content of the contralateralkidney was also observed which was of greatermagnitude than and equalin duration to that oftheinjured kidney (10). Furthermore, in rats develop-ing chronic, unilateral Proteus mirabilis-inducedpyelonephritis (7), the levels of renal lysozyme inboth kidneys exhibit a second elevation with thecontralateral kidney showing the higher lysozymeactivity (10).
Incidence of infection in the experimentally un-infected kidney is low (7), and pathological indi-cations of infection are absent (10). These findingsdo not eliminate the possibility that viable bac-teria in the experimentally uninfected kidney areresponsible for the observed results. To investigatethis possibility and furthermore to investigate therole of the inflammatory response in the tissuedestruction characteristic of chronic pyelonephri-tis, a sterile pyelonephritis model induced withheat-killed bacteria has been developed. Thiscommunication concerns the description of thismodel and the levels of soluble renal lysozyme inthe kidneys of rats developing experimentallyinduced, sterile, unilateral, chronic pyelonephritis.
MATERIALS AND METHODS
Experimental animals. CFN rats (Wistar derived,Carworth Farms) of either sex weighing 160 to 184 gwere used. Animals were housed five per wire mesh
cage (38 by 35 by 18 cm) and maintained on Purinalaboratory chow and water. The cage rooms werelighted from 6 AM to 6 PM. Animals were sacrificedbetween 9:00 and 11:00 AM.
Injection of dead cells into the kidney. P. mirabilis(Pr-91) was grown overnight in Brain Heart Infusionbroth at 37 C on a shaker. The cells were harvested bycentrifugation (12,000 X g for 10 min), washed twicewith sterile 0.85% saline, suspended in saline at threetimes the original concentration, and autoclaved at121 C for 15 min. The dead cells were cooled to roomtemperature and immediately injected into the kid-ney cortex by using a previously described technique(7, 10). Kidneys were titered for viable bacteriaas previously described (7).
Urease assays. P. mirabilis cells were prepared asdescribed above, and the suspension was divided intotwo samples. One sample was autoclaved for 15 min at121 C. Both live and dead bacteria were assayed forurease activity as follows. The cells were harvested bycentrifugation at 3,000 X g for 15 min, washed, andsuspended in 0.1 M tris(hydroxymethyl)aminomethanebuffer (pH 8.0) that was 0.1 M in sodium chloride. Thereaction mixture contained 2.4 ml of the cell suspen-sion and 0.1 ml of urea (final concentration, 0.15 M).The reaction mixture was incubated for 10 min at 37 C,and ammonia nitrogen was determined by the methodof Seligson and Seligson (18). Activity was calculatedas micrograms of ammonia nitrogen liberated perminute, and specific activity was calculated as activityper milligram of cells.
Preparation of renal lysozyme. Kidneys (approxi-mate weight, 1 g) were removed immediately fromCO2-killed rats and, after the pelvis was teased away,were placed in 9.0 ml of 0.85% saline at 0 C. All sub-sequent procedures were carried out at 0 to 4 C. Thekidneys were homogenized in a Potter-Elvehjem tissuehomogenizer, and the homogenate was centrifuged at48,200 X g for 10 min. The resulting supernatant fluidserved as the source of lysozyme.
269
on June 3, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
EUDY, BURROUS, AND SIGLER
41 4v t1* o,n"I'M~~
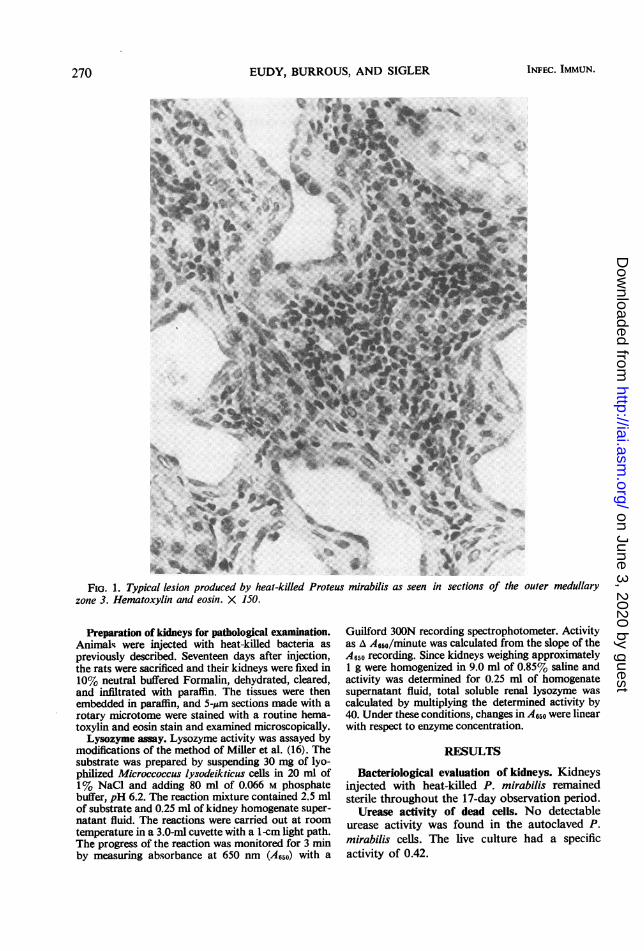
FIG. 1. Typical lesion produced by heal-killed Proteus mirabilis as seen in sections of the outer medullaryzone 3. Hematoxylin and eosin. X 150.
Preparation of kidneys for pathological examination.Animals were injected with heat-killed bacteria as
previously described. Seventeen days after injection,the rats were sacrificed and their kidneys were fixed in10% neutral buffered Formalin, dehydrated, cleared,and infiltrated with paraffin. The tissues were thenembedded in paraffin, and 5-IAm sections made with arotary microtome were stained with a routine hema-toxylin and eosin stain and examined microscopically.Lysozyme assay. Lysozyme activity was assayed by
modifications of the method of Miller et al. (16). Thesubstrate was prepared by suspending 30 mg of lyo-philized Microccoccus lysodeikticus cells in 20 ml of1% NaCl and adding 80 ml of 0.066 M phosphatebuffer, pH 6.2. The reaction mixture contained 2.5 mlof substrate and 0.25 ml of kidney homogenate super-natant fluid. The reactions were carried out at roomtemperature in a 3.0-ml cuvette with a 1-cm light path.The progress of the reaction was monitored for 3 minby measuring absorbance at 650 nm (A650) with a
Guilford 300N recording spectrophotometer. Activityas A A650/minute was calculated from the slope of theA650 recording. Since kidneys weighing approximately1 g were homogenized in 9.0 ml of 0.85% saline andactivity was determined for 0.25 ml of homogenatesupernatant fluid, total soluble renal lysozyme wascalculated by multiplying the determined activity by40. Under these conditions, changes in A650 were linearwith respect to enzyme concentration.
RESULTS
Bacteriological evaluation of kidneys. Kidneysinjected with heat-killed P. mirabilis remainedsterile throughout the 17-day observation period.
Urease activity of dead cells. No detectableurease activity was found in the autoclaved P.mirabilis cells. The live culture had a specificactivity of 0.42.
INFEC. IMMUN.270
on June 3, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
STERILE PYELONEPHRITIS 271
4/
o2 4 6 8 111 1'2 14 16 18
Day, after injection
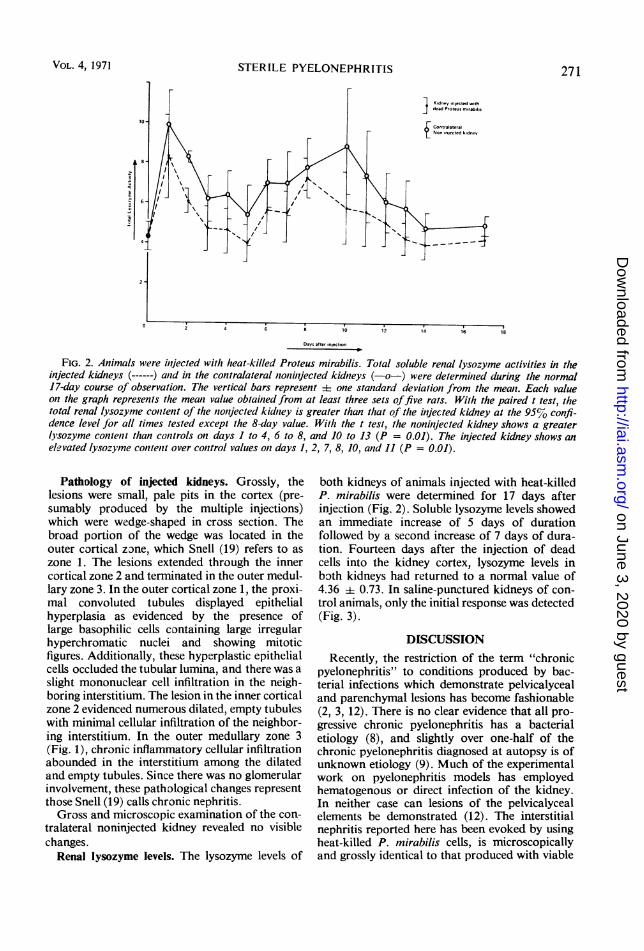
FIG. 2. Animals were inijected with heat-killed Proteus mirabilis. Total soluble renal lysozyme activities in t(leinjected kidneys (------) atid in the conitralateral zonizit/ected kidneys (-o-) were determinied during thle normal17-day course of observation. The vertical bars represent 4 one standard devialion from the meaw. Each valueon the graph represents tlhe mean value obtained from at least three sets offive rats. With the paired t test, thetotal renal lysozyme conztent of the zoiijected kidntey is greater than that of the inijected kidney at thle 95% confi-dence level for all times tested except the 8-day value. With the t test, the noninjected kidney shows a greaterlysozyme contenit than conttrols on days I to 4, 6 to 8, and 10 to 13 (P = 0.01). The injected kidney shiows anelevated lysozyme contenit over control values onz days 1, 2, 7, 8, 10, and 11 (P = 0.01).
Pathology of injected kidneys. Grossly, thelesions were small, pale pits in the cortex (pre-sumably produced by the multiple injections)which were wedge-shaped in cross section. Thebroad portion of the wedge was located in theouter cortical zone, which Snell (19) refers to aszone 1. The lesions extended through the innercortical zone 2 and terminated in the outer medul-lary zone 3. In the outer cortical zone 1, the proxi-mal convoluted tubules displayed epithelialhyperplasia as evidenced by the presence oflarge basophilic cells containing large irregularhyperchromatic nuclei and showing mitoticfigures. Additionally, these hyperplastic epithelialcells occluded the tubular lumina, and there was aslight mononuclear cell infiltration in the neigh-boring interstitium. The lesion in the inner corticalzone 2 evidenced numerous dilated, empty tubuleswith minimal cellular infiltration of the neighbor-ing interstitium. In the outer medullary zone 3(Fig. 1), chronic inflammatory cellular infiltrationabounded in the interstitium among the dilatedand empty tubules. Since there was no glomerularinvolvement, these pathological changes representthose Snell (19) calls chronic nephritis.Gross and microscopic examination of the con-
tralateral noninjected kidney revealed no visiblechanges.
Renal lysozyme levels. The lysozyme levels of
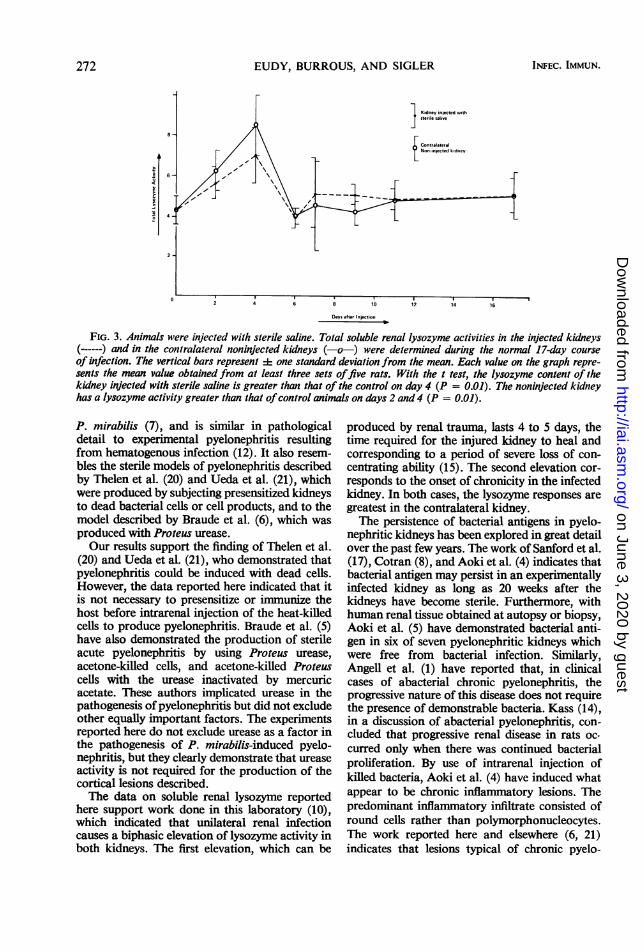
both kidneys of animals injected with heat-killedP. mirabilis were determined for 17 days afterinjection (Fig. 2). Soluble lysozyme levels showedan immediate increase of 5 days of durationfollowed by a second increase of 7 days of dura-tion. Fourteen days after the injection of deadcells into the kidney cortex, lysozyme levels inboth kidneys had returned to a normal value of4.36 + 0.73. In saline-punctured kidneys of con-trol animals, only the initial response was detected(Fig. 3).
DISCUSSIONRecently, the restriction of the term "chronic
pyelonephritis" to conditions produced by bac-terial infections which demonstrate pelvicalycealand parenchymal lesions has become fashionable(2, 3, 12). There is no clear evidence that all pro-gressive chronic pyelonephritis has a bacterialetiology (8), and slightly over one-half of thechronic pyelonephritis diagnosed at autopsy is ofunknown etiology (9). Much of the experimentalwork on pyelonephritis models has employedhematogenous or direct infection of the kidney.In neither case can lesions of the pelvicalycealelements be demonstrated (12). The interstitialnephritis reported here has been evoked by usingheat-killed P. mirabilis cells, is microscopicallyand grossly identical to that produced with viable
VOL. 4, 1971
on June 3, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
EUDY, BURROUS, AND SIGLER
6-~~~~~-
4 6 8 '10 12 14 16
Days after Injection
FIG. 3. Animals were injected with sterile saline. Total soluble renal lysozyme activities in the injected kidneys(------) and in the contralateral noninjected kidneys (-o-) were determined during the normal 17-day courseof infection. The vertical bars represent :1: one standard deviation from the mean. Each value on the graph repre-sents the mean value obtained from at least three sets offive rats. With the t test, the lysozyme content of thekidney injected with sterile saline is greater than that of the control on day 4 (P = 0.01). The noninjected kidneyhas a lysozyme activity greater than that ofcontrol animals on days 2 and 4 (P = 0.01).
P. mirabilis (7), and is similar in pathologicaldetail to experimnental pyelonephritis resultingfrom hematogenous infection (12). It also resem-bles the sterile models of pyelonephritis describedby Thelen et al. (20) and Ueda et al. (21), whichwere produced by subjecting presensitized kidneysto dead bacterial cells or cell products, and to themodel described by Braude et al. (6), which wasproduced with Proteus urease.Our results support the finding of Thelen et al.
(20) and Ueda et al. (21), who demonstrated thatpyelonephritis could be induced with dead cells.However, the data reported here indicated that itis not necessary to presensitize or immunize thehost before intrarenal injection of the heat-killedcells to produce pyelonephritis. Braude et al. (5)have also demonstrated the production of sterileacute pyelonephritis by using Proteus urease,acetone-killed cells, and acetone-killed Proteuscells with the urease inactivated by mercuricacetate. These authors implicated urease in thepathogenesis of pyelonephritis but did not excludeother equally important factors. The experimentsreported here do not exclude urease as a factor inthe pathogenesis of P. mirabilis-induced pyelo-nephritis, but they clearly demonstrate that ureaseactivity is not required for the production of thecortical lesions described.The data on soluble renal lysozyme reported
here support work done in this laboratory (10),which indicated that unilateral renal infectioncauses a biphasic elevation of lysozyme activity inboth kidneys. The first elevation, which can be
produced by renal trauma, lasts 4 to 5 days, thetime required for the injured kidney to heal andcorresponding to a period of severe loss of con-centrating ability (15). The second elevation cor-responds to the onset of chronicity in the infectedkidney. In both cases, the lysozyme responses aregreatest in the contralateral kidney.The persistence of bacterial antigens in pyelo-
nephritic kidneys has been explored in great detailover the past few years. The work of Sanford et al.(17), Cotran (8), and Aoki et al. (4) indicates thatbacterial antigen may persist in an experimentallyinfected kidney as long as 20 weeks after thekidneys have become sterile. Furthermore, withhuman renal tissue obtained at autopsy or biopsy,Aoki et al. (5) have demonstrated bacterial anti-gen in six of seven pyelonephritic kidneys whichwere free from bacterial infection. Similarly,Angell et al. (1) have reported that, in clinicalcases of abacterial chronic pyelonephritis, theprogressive nature of this disease does not requirethe presence of demonstrable bacteria. Kass (14),in a discussion of abacterial pyelonephritis, con-cluded that progressive renal disease in rats oc-curred only when there was continued bacterialproliferation. By use of intrarenal injection ofkilled bacteria, Aoki et al. (4) have induced whatappear to be chronic inflammatory lesions. Thepredominant inflammatory infiltrate consisted ofround cells rather than polymorphonucleocytes.The work reported here and elsewhere (6, 21)indicates that lesions typical of chronic pyelo-
272 INFEC. IMMUN.
on June 3, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

STERILE PYELONEPHRITIS
nephritis can be produced in the rat in the absenceof viable bacteria.
In conclusion, the work reported here indicatesthat chronic interstitial nephritis with renal lesionssimilar to those of chronic pyelonephritis andidentical to those of experimental pyelonephritisin animal models produced by hematogenous anddirect renal puncture can be produced by heat-killed bacteria. The pathogenesis of the diseasedoes not depend on viable bacteria nor on thepresence of active bacterial urease. Soluble renallysozyme in these injected kidneys is elevated bi-phasically with the second elevation in lysozymecorresponding to the onset of chronicity in theinjected kidney. The soluble renal lysozyme ac-tivities of the contralateral kidney show a similarincrease, of equal duration but of greater magni-tude.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the expert assistance,patience, and attention to detail of Roberta Alexander, BarbaraBensko, Carol Dubiel, and Nancy Gallo. Urease assays werekindly provided by Frank Kopko and Jon Andersen. Statisticalevaluations were conducted by C. T. Tu.
LITERATURE CITED
1. Angell, M. E., A. S. Relman, and S. L. Robbins. 1968."Active" chronic pyelonephritis without evidence of bac-terial infection. N. Engl. J. Med. 278:1303-1308.
2. Anonymous. 1968. Pyelonephritis without bacteria (editorial).Lancet 2:1125-1126.
3. Anonymous. 1970. Markers for chronic pyelonephritis (edi-torial). Lancet 1:758-759.
4. Aoki, S., M. Merkel, M. Aoki, and W. R. McCabe. 1967.Immunofluorescent localization of bacterial antigen inpyelonephritis. 1. The use of antisera against the commonenterobacterial antigen in experimental renal lesions. J.Lab. Clin. Med. 70:204-212.
5. Aoki, S., S. Imamura, M. Aoki, and W. R. McCabe. 1969."Abacterial" and bacterial pyelonephritis. Immunofluo-rescent localization of bacterial antigen. N. Engl. J.-Med.281:1376-1382.
6. Braude, A. I., J. Siemienski, and A. P. Shapiro. 1960.The role of bacterial urease in the pathogenesis of pyelo-nephritis, p. 69-88. In E. L. Quinn and E. H. Kass (ed.),Biology of pyelonephritis. Little, Brown and Co., Boston.
7. Burrous, S. E., and J. B. Cawein. 1969. Rat pyelonephritismodel suitable for primary or secondary screening. Appl.Microbiol. 18:448-451.
8. Cotran, R. S. 1963. Retrograde Proteus pyelonephritis inrats. Localization of antigen and antibody in treatedsterile pyelonephritic kidneys. J. Exp. Med. 117:813-822.
9. Cotran, R. S. 1969. The renal lesion of chronic pyelonephritis:immunofluorescent and ultrastructural studies. J. Infec.Dis. 120:109-118.
10. Eudy, W. W., and S. E. Burrous. 1971. Renal lysozyme levelsin animals developing Proteus mirabilis-induced pyelone-phritis. Appl. Microbiol. 21:300-305.
11. Hamdy, M. K. 1969. Effect of trauma and infection on lyso-zyme in poultry tissue. Proc. Soc. Exp. Biol. Med. 131:409-415.
12. Heptinstall, R. H. 1969. The enigma of chronic pyelonephritis.J. Infec. Dis. 120:104-108.
13. Kalmanson, G. M., and L. B. Guze. 1963. Pyelonephritis. Anattempt to demonstrate anti-kidney antibody in the sera ofpatients with chronic bacteriuria. Amer. J. Med. Sci. 246:532-536.
14. Kass, E. H. 1969. Bacterial antigen in the kidney. N. Engl.J. Med. 281:1420-1421.
15. Kaye, D., and H. Rocha. 1970. Urinary concentrating abilityin early experimental pyelonephritis. J. Clin. Invest. 49:1427-1437.
16. Miller, T. E., C. M. Cameron, and J. D. K. North. 1968.Distribution of lysozyme in the rat kidney and the roleof this enzyme in experimental pyelonephritis. Proc. Soc.Exp. Biol. Med. 128:749-752.
17. Sanford, J. P., B. W. Hunter, and L. L. Souda. 1962. Therole of immunity in the pathogenesis of experimentalhematogenous pyelonephritis. J. Exp. Med. 115:383-410.
18. Seligson, D., and H. Seligson. 1951. A microdiffusion methodfor the determination of nitrogen liberated as ammonia. J.Lab. Clin. Med. 38:324-330.
19. Snell, K. C. 1967. Renal disease of the rat, p. 105-147. In E.Cotchin and J. C. F. Roe (ed.), Pathology of laboratoryrats and mice. F. A. Davis Co., Philadelphia.
20. Thelen, A., K. Rother, and H. Sarre. 1956. ExperimentelleUntersuchungen zur Pathogenese der pyelonephritischenSchrumpfniere. Urol. Int. 3:359-389.
21. Ueda, Y., 0. Sakai, and T. Takasu. 1967. A study on experi-mental pyelonephritis: effect of E. coli endotoxin in chronicpyelonephritis, p. 140-149. Iln R. H. Heptinstall (ed.),Morphology, immunology, urology. S. Karger, Basel.
VOL. 4, 1971 273
on June 3, 2020 by guesthttp://iai.asm
.org/D
ownloaded from