regulation of urea cycle enzymes in - cancer research
TRANSCRIPT

[CANCER RESEARCH 41. 2692-2699, July 1981]0008-5472/81 /0041-OOOOS02.00
Regulation of Urea Cycle Enzymes in Transplantable Hepatomas and inthe Livers of Tumor-bearing Rats and Humans1
Linda D. Brebnor, Johanna Grimm, and John B. Balinsky2
Department of Biochemistry. University of the Witwatersrand, Johannesburg 2001, South Africa
ABSTRACT
The levels of the five enzymes of the urea cycle were measured in normal 5-week-old rats, in a transplantable hepatoma,and in the livers of tumor-bearing rats (host livers). The levels
of all five enzymes were much lower in the hepatoma, althoughthere was no exact correlation of the decrease in levels. In hostlivers, the levels were higher than in the tumors, but lower thanin normal liver. The levels of all five urea cycle enzymes werepositively correlated with dietary protein content in normallivers, in hepatomas, and in host livers. In fact, the hepatomasshowed the greatest changes in response to diet. On all diets,the levels in host liver remained below those in normal liver,indicating that the decreased level was probably not due topreferential utilization of nutrients by the tumor.
The levels of urea cycle enzymes in normal liver were notaltered by a single injection of glucocorticoid, glucagon, ordibutyryl cyclic adenosine 3':5'-monophosphate. By contrast,
in hepatoma, the levels were usually significantly elevated bythe same treatment. In addition, the levels in host livers werealways significantly elevated and were usually above those innormal animals, whether the latter were hormone treated ornot.
Injection of plasma from tumor-bearing rats into normal ani
mals produced a decrease in the levels of all five enzymes; ifglucagon was injected together with the plasma, large increases in levels were observed. This result supports theconcept of a humoral factor produced by the tumor whichaffects the levels and the inducibility of urea cycle enzymes inhost livers.
Autopsied human primary hepatomas also showed levels ofurea cycle enzymes below those in normal livers with hostlivers having intermediate values. A cell line derived from ahuman hepatoma showed induction of arginase by glucocorticoid in culture; in this, it resembled a cell line of the rathepatoma. Tyrosine aminotransferase in human hepatoma cellswas not induced by glucocorticoid; in this, it differed from therat hepatoma cells where induction of this enzyme was observed.
INTRODUCTION
Urea cycle enzymes in mammalian liver serve the function ofdetoxification of ammonia produced by the degradation ofamino acids. They are therefore associated with protein catab-
olism and gluconeogenesis from amino acids.
Mammalian urea cycle enzymes are susceptible to regulation. Schimke (25) has shown a positive correlation betweenall 5 enzymes of the urea cycle and dietary protein content.These enzymes are also inducible by glucagon (15) and glucocorticoid (26). In adults, repeated injections of hormone over2 to 7 days are usually utilized to raise the levels of urea cycleenzymes, although Snodgrass et al. (29) in one experimentreport significant effects for 4 of the enzymes after a singleinjection of glucagon. In neonatal rats, urea cycle enzymes areinitially low and increase during the first weeks of life. At thattime, they are much more sensitive to hormonal control, andinduction can be obtained by a single dose of glucagon orglucocorticoid (10, 27).
High levels of urea cycle enzymes are characteristic of fullydifferentiated adult rat liver. Extensive work on chemicallyinduced rat hepatomas has shown that they have altered metabolism, frequently with an enhanced glycolysis and decreased gluconeogenesis (31 ). Moreover, susceptibility in hepatomas is different from that in normal liver (28). It might beexpected that the behavior of urea cycle enzymes could resemble that of gluconeogenic enzymes. Alterations in regulatoryproperties might also be expected. Levels of GPS3 and OTC
had been measured previously in hepatomas (13, 18), but acomplete investigation of all enzymes of the urea cycle had notso far been made. The urea cycle enzymes are a group of 5functionally related enzymes. In normal tissues, they frequentlyshow correlated regulation (1), and their levels are frequentlycorrelated with one another in different species (16). It seemedof interest, therefore, to investigate the levels and regulation ofthese enzymes in hepatomas and the extent to which theirbehavior in tumors is correlated, both qualitatively and quantitatively.
The results described below represent a study of urea cycleenzymes in a chemically induced rat hepatoma (2). They showdecreased levels but increased inducibility. Moreover, the experiments demonstrate a clear-cut effect of the tumor on thelivers of tumor-bearing rats (host livers). This effect includes
decreased levels and increased inducibility. The results alsopresent evidence for the humoral transmission of these effects.
Samples of autopsied human primary hepatoma were available, as well as a cell line derived from a human hepatoma.While it was not possible to repeat on the human hepatoma thefull range of experiments carried out on rat tissues, sufficientwork on these was possible to indicate the likelihood that fairlysimilar controls operate in human tumors. This supports therelevance of the work to problems of human disease. A preliminary report on this work has been published (5).
' This investigation was supported in part by a grant from the South African
Council for Scientific and Industrial Research. Most of the work is taken from thePh.D. Dissertation of Linda D. Brebnor.
2 Present address: Department of Zoology, Iowa State University, Ames, Iowa
50011. To whom requests for reprints should be addressed.Received August 4, 1980; accepted April 7, 1981.
3 The abbreviations used are: CPS, carbamoylphosphate synthetase; OTC,ornithine carbamoyltransferase; dibutyryl cyclic AMP, AV6-O2'-dibutyryl cyclicadenosine 3':5'-monophosphate; ASL, argininosuccinate lyase; TAT, tyrosine
aminotransferase.
2692 CANCER RESEARCH VOL. 41
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

Urea Cycle Enzyme Regulation in Tumor-bearing Rats and Humans
MATERIALS AND METHODS
Materials. ATP, A/-acetylglutamate, casein (light, white, vitamin free), dilithium carbamyl phosphate, L-ornithine, urease,argininosuccinate, L-arginine-HCI, bovine serum albumin, so
dium potassium tartrate, cornstarch, cortisol hemisuccinate,glucagon, and dibutyryl cyclic AMP were obtained from SigmaChemical Co., St. Louis, Mo. yS-Methasone was obtained asBetasolan, soluble, for injection (2 mg/ml) from Glaxo-Allen-
bury, Johannesburg, South Africa. Gevral vitamin:mineral nutritional supplement was supplied by Lederle Laboratories,Isando, South Africa, and food yeast was from Anchor, Industria, South Africa. Tissue culture medium ingredients (Eagle's
minimal essential medium and fetal calf serum) were obtainedfrom Grand Island Biological Co., Grand Island, N. Y. All otherreagents were obtained from British Drughouses Ltd., Poole,England, or from E. Merck Co., Darmstadt, Germany.
Experimental Animals. Albino Sprague-Dawley rats werebred and supplied for this investigation by the animal facility ofthe University of the Witwatersrand. All animals were maintained on the "8 + 16" controlled feeding schedule of Potter
et al. (20). Rats were killed when they were 35 days old.To ensure that normal and tumor-bearing rats consumed the
same amount of food, both groups were given quantities offood which were equal and slightly less than the maximumamount which the animals would eat. All food given was alwaysconsumed.
The diets used were as described by us elsewhere (6).Transplantable Rat Hepatomas. The liver tumor was origi
nally induced by Dr. C. F. de V. Albrecht and Dr. L. D. Nourse(2) of the National Chemical Research Laboratories, Pretoria,South Africa. It will hereafter be referred to as the hepatomaDAB-1, the name currently used by these investigators.
The hepatoma, derived from the Wistar strain, was shown byrepeated trials not to be directly transplantable into Sprague-
Dawley rats. A cell culture of the hepatoma was carried out inthis laboratory as described previously (11 ). After the cells hadbeen in culture for about 4 weeks, it was found that the culturedhepatoma line was transplantable into Sprague-Dawley rats.This line was used for all the experiments described in thepresent paper.
Three-week-old rats were lightly anesthetized with ether. The
neoplastic tissue from a freshly sacrificed animal was coveredwith 0.9% NaCI solution and cut very finely with a sharp pair ofscissors. Approximately 1 to 2 ml of tissue was sucked up intoa clean tuberculin syringe fitted with a 15-gauge needle. The
injections were made into the muscle on the inside of the hindleg. The tumors were allowed to grow for 2 weeks, reaching aweight of about 2 g. After this, the animals were sacrificed, andthe hepatomas were dissected out. Only growing hepatomatissue was used, care being taken to avoid necrotic tissue.
Cells cultured in vitro were scraped off, suspended in 0.15M sodium chloride containing 10 rtiM sodium phosphate buffer,pH 7.4, and counted in a hemocytometer. They were storedfrozen in liquid nitrogen. For enzyme assay, the cells werethawed; to complete enzyme extraction, they were blended inthe same way as other tissues, using an Ultra-Turrax homog-enizer.
Human Hepatoma and Normal Human Liver. Normal humanliver tissue, removed at autopsy from accident victims, wasprovided by Dr. M. G. G. Maier, South African Institute for
Medical Research, Johannesburg, South Africa. Livers andhepatomas from patients with primary hepatocellular carcinoma were provided by Dr. E. M. Geddes of Crown MinesHospital and Professor I. Webster and staff at the South AfricanInstitute for Medical Research. The cancerous regions of thelivers were dissected from the regions which appeared to bemorphologically normal. Only hepatoma tissue in which therewas less than 15% necrosis (as determined histologically bythe Histopathology Department of the South African Institutefor Medical Research) was used for assay.
A human hepatoma cell line derived from a patient withprimary liver cancer ("Mahlavu culture") was obtained from
Dr. O. W. Prozesky of the University of Pretoria (23). The cellswere cultured by methods similar to those for the rat hepatoma,except that supplementation was with 15% fetal calf serum.
Method of Obtaining Rat Plasma. Both normal and tumor-bearing rats were fatally anesthetized by injecting sodiumpentabarbitone (15 mg/animal) i.p. The thoracic cavity wasopened, and 5 ml of blood were withdrawn from the posteriorvena cava. The blood was dispensed immediately into EDTA-treated centrifuge tubes, prepared by transferring into them 1ml of 10% EDTA by pipet and drying in a hot oven. The bloodcells were removed by centrifugation at 500 x g for 10 min.The supernatant plasma was then injected i.p. into normal 35-day-old non-tumor-bearing animals (2 ml/animal).
Preparation of Homogenates and Assay of Urea CycleEnzymes. Homogenates of livers, hepatomas, and culturedcells in 20 mw potassium phosphate buffer, pH 7.4, anddilutions in 10 mw manganese sulfate were prepared as described previously (3). CPS (EC 6.3.4.16), OTC (carbamoyl-phosphate-L-ornithine carbamoyltransferase) (EC 2.1.3.3), argininosuccinate synthetase (L-citrulline:L-aspartate ligase(AMP)) (EC 6.3.4.5), ASL (L-argininosuccinate arginineilyase)(EC 4.3.2.1), and arginase (L-arginine ureohydrolase) (EC
3.5.3.1) were assayed as described previously (4). For assaysof arginase in rat and human cell lines, the incubation periodwas extended to 1 hr. The reaction was shown to be linear forat least 90 min.
TAT (u-tyrosine:2-oxoglutarate aminotransferase) (EC
2.6.1.5) was assayed by the method of Diamondstone (9).All enzyme activities were expressed in units defined as
utilizing 1 ftmol of substrate or producing 1 /xmol of product permin at 37° under the assay conditions described. Specific
activity is defined as the activity per mg of protein, the latterbeing measured by the method of Lowry ef al. (14), usingbovine serum albumin as standard.
RESULTS
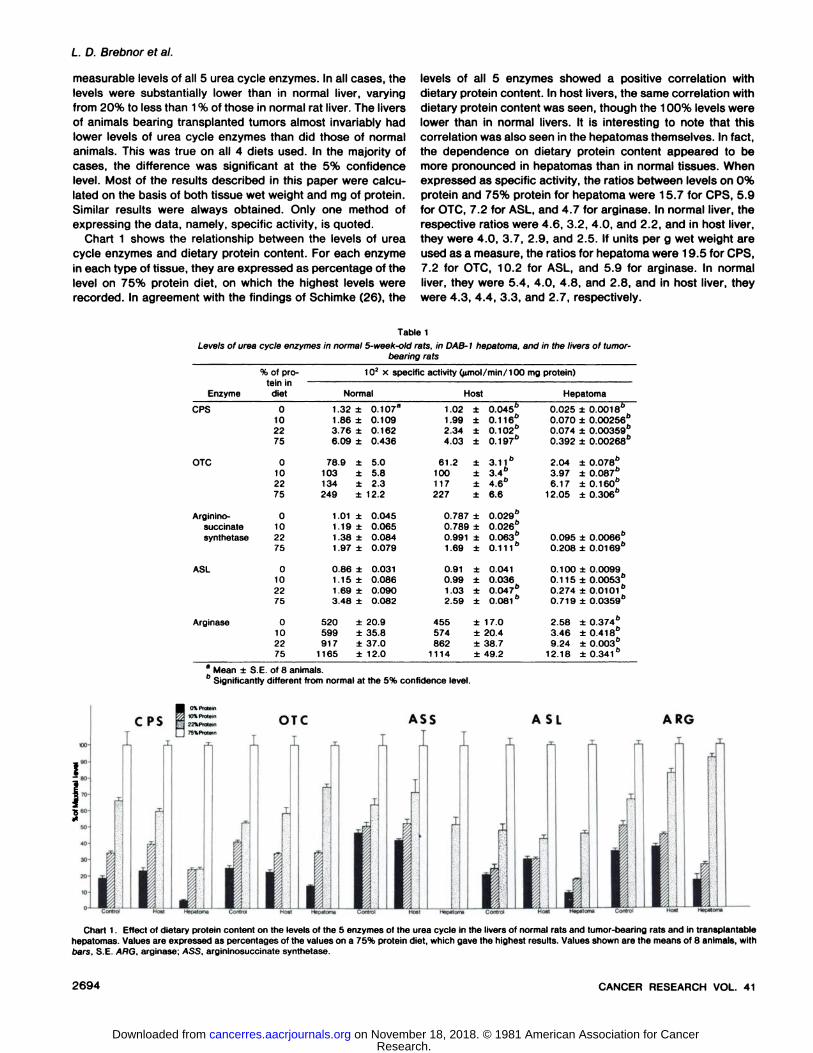
Levels of Urea Cycle Enzymes in Normal Liver, Host Liver,and Hepatoma. In Table 1, giving the results of the firstexperiments, are shown the levels of urea cycle enzymes inthe livers of normal rats, expressed as units per mg protein(specific activity). Table 1 also shows measurements of thelevels of urea cycle enzymes in hepatomas implanted intoanimals from the same group and in the livers of the animalswhich received implants. The experiment was carried out onanimals fed diets which varied in protein content. The 4 dietswere standard rat diet, containing 22% protein, and syntheticdiets containing 0, 10, and 75% protein.
Table 1 shows that the transplantable hepatomas contained
JULY 1981 2693
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from
L. D. Brebnor et al.
measurable levels of all 5 urea cycle enzymes. In all cases, thelevels were substantially lower than in normal liver, varyingfrom 20% to less than 1% of those in normal rat liver. The liversof animals bearing transplanted tumors almost invariably hadlower levels of urea cycle enzymes than did those of normalanimals. This was true on all 4 diets used. In the majority ofcases, the difference was significant at the 5% confidencelevel. Most of the results described in this paper were calculated on the basis of both tissue wet weight and mg of protein.Similar results were always obtained. Only one method ofexpressing the data, namely, specific activity, is quoted.
Chart 1 shows the relationship between the levels of ureacycle enzymes and dietary protein content. For each enzymein each type of tissue, they are expressed as percentage of thelevel on 75% protein diet, on which the highest levels wererecorded. In agreement with the findings of Schimke (26), the
levels of all 5 enzymes showed a positive correlation withdietary protein content. In host livers, the same correlation withdietary protein content was seen, though the 100% levels werelower than in normal livers. It is interesting to note that thiscorrelation was also seen in the hepatomas themselves. In fact,the dependence on dietary protein content appeared to bemore pronounced in hepatomas than in normal tissues. Whenexpressed as specific activity, the ratios between levels on 0%protein and 75% protein for hepatoma were 15.7 for CPS, 5.9for OTC, 7.2 for ASL, and 4.7 for arginase. In normal liver, therespective ratios were 4.6, 3.2, 4.0, and 2.2, and in host liver,they were 4.0, 3.7, 2.9, and 2.5. If units per g wet weight areused as a measure, the ratios for hepatoma were 19.5 for CPS,7.2 for OTC, 10.2 for ASL, and 5.9 for arginase. In normalliver, they were 5.4, 4.0, 4.8, and 2.8, and in host liver, theywere 4.3, 4.4, 3.3, and 2.7, respectively.
Table 1Levels of urea cycle enzymes in normal 5-week-old rats, in DAB-1 hepatoma. and in the livers of tumor-
bearing rats
EnzymeCPSOTCArginino-succinatesynthetaseASLArginase%
of protein indiet01022750102275010227501022750102275102
X specific activity (fimol/min/100 mgprotein)Normal1.32
±0.107a1.86
±0.1093.76±0.1626.09±0.43678.9
±5.0103±5.8134±2.3249±12.21.01
±0.0451.19±0.0651.38 ±0.0841.97
±0.0790.86
±0.0311.15 ±0.0861.69
±0.0903.48±0.082520
±20.9599±35.8917
±37.01165±12.0Host1.02
±0.04561.99±0.116"2.34±0.10264.03±0.197o61.2
±3.116100±3.4b117±4.6b227
±6.60.787
±0.029"0.789±0.026"0.991±0.063"1.69
±0.1110.91
±0.0410.99±0.0361.03±0.047o2.59±0.08l"455
±17.0574±20.4862±38.71114±49.2Hepatoma0.025
±0.001860.070±0.00256"0.074±0.00359"0.392±0.00268"2.04
±0.078"3.97±0.087"6.17
±0.160"12.05±0.306"0.095
±0.0066"0.208±0.0169"0.100
±0.00990.115±0.00530.274±0.0101"0.719±0.035962.58
±0.374"3.46±0.418"9.24±0.003"12.18±0.341 "
a Mean ±S.E. of 8 animals.' Significantly different from normal at the 5% confidence level.
ASL ARG
Chart 1. Effect of dietary protein content on the levels of the 5 enzymes of the urea cycle in the livers of normal rats and tumor-bearing rats and in transplantablehepatomas. Values are expressed as percentages of the values on a 75% protein diet, which gave the highest results. Values shown are the means of 8 animals, withoars, S.E. ARG, arginase. ASS, argininosuccinate synthetase.
2694 CANCER RESEARCH VOL. 41
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

Urea Cycle Enzyme Regulation in Tumor-bearing Rats and Humans
Hormonal Induction of the Urea Cycle. In the next series ofexperiments, the susceptibility of the urea cycle enzymes toinduction by hormones was tested. The hormones used were/?-methasone (2 fig/g body weight), glucagon (5 /¿g/gbody
weight), and dibutyryl cyclic AMP (70 jug/g body weight), given¡.p.as single doses 24 hr prior to killing the animals. The dosesand time interval correspond in general to those used earlieron neonatal rats (27). The dose of dibutyryl cyclic AMP is onewhich produced maximum induction of TAT (32). As in theprevious experiments, rats kept on 4 different diets were used.
The results are shown in Table 2, expressed as percentagesof the specific activity of corresponding control levels. A singleinjection of glucagon or dibutyryl cyclic AMP into normal animals gave no significant increase in the level of any of the 5urea cycle enzymes. This was true on all 4 diets used. A singleinjection of /8-methasone, in general, also failed to result in
increases of urea cycle enzymes, although on a 10% proteindiet, significant increases were seen in CPS, argininosuccinatesynthetase, and arginase. These were, however, substantiallysmaller than those seen in the corresponding host livers.
The levels of urea cycle enzymes were usually raised in thehepatomas of hormone-injected animals. The increases were
usually significant at the 5% confidence level. In some groups,they fell short of significance; in other groups, they were verylarge indeed. In particular, the greatest hormone effects wereseen on the 10% protein diet, with approximately 2-fold in
creases in argininosuccinate synthetase and arginase by both/8-methasone and glucagon. It is important to note also that, on
the 10% protein diet, all 3 hormones produced significantincreases in hepatomas of all 5 urea cycle enzymes.
A very important result is the response to hormones in hostlivers. All 5 enzymes showed substantial increases in all hostlivers. The increases ranged from 20 to over 200% and werealways significant at the 5% confidence level. This was true for
all 3 hormones and for all 4 diets. It is in striking contrast withthe response of normal livers, which almost always showed nosignificant effect.
Table 3 represents a further analysis of the data in whichenzyme levels in hormone-treated host livers are expressed as
Table 3Levels of urea cycle enzymes after hormone injections in the livers of tumor-
bearing animals compared to normal
Animals were given injections of hormones 24 hr before sacrifice.
CPSOTCArginino
succinatesynthetaseASLArginase%of
proteinindiet01022750102275010227501022750102275/3-Methasone9385929110476124108119122140120111991449011088123122±±±±±±±±±±±±±±±±±±+±7"359426°25"9*l?"3b5762456"4"Glucagon1161081241051129713812210119616013914015317399111107112131±±±±±±±±±±±±±±±+±±±±722"1074<3"e11"14"2b3Ì9"7"3653*6"Dibutyryl
cyclicAMP12314011710010514014112997188157141136166159103112135127142±±±±±±±±±±±±±±±±±±±±128*8837*7"e"4K910b4b4"12611"63"K56Ö5Ò
8 Mean ±S.E. of 8 animals, expressed in percentages of the specific activities
in the livers of normal rats from the same population receiving concurrentinjections of the same hormone, calculated to 3 significant figures, and thenrounded.
0 Significantly above hormone-treated controls at the 5% confidence level.
Table 2Effect of single doses of hormones on the levels of urea cycle enzymes in normal 5-week-old rats, in DAB-1 hepatomas, and livers of tumor-bearing rats
Animals were given injections of hormones 24 hr before sacrifice.
EnzymeCPSOTCArgininoProteinindiet010227501022750succinate
10synthetase22ASLArginase7501022750102275Normal113
±9a139
±3105±4109±591
±5112±4107±4113±691
±4127±86103
±6104±2106
±5119±598±3107±2104
±6112±3698
±4108±2/3-MethasoneHost138
±106170±6°155±9°150±14b122±
46133±4s151
±8b134±46138
±66271
±21200±146144±96113±
36137±7"150±76129±3b131
±5*165±7Ö128±
5b138 ± 5ÖHepatoma99
±3156 ±16138±
5b118
±8109
±6150±8b113
±5106±3148
±18Ö188 ±13b146
±12"169±7b100
±9133 ±10115±
26116±36112
±17227±266105
±8118±36Normal114
±6104±5100±495±8110
±592±2102±398±4109
±795±696±4100±2105
±893±590±597±4110
±492±5105±199±1QlucagonHost172
±106163±46200
±34°150±146159±
96138±56160±76132±36139±
96323±186213
±18162±2°142
±36166±10Ö165±7b128±
46139±
7b165±8b125±46136±
6Dibutyryl
cyclicAMPHepatoma1
56 ±56159
±19160 ±13109
±6124
±8b167±106109±2b103
±2125
±76242±17b143
±14154±36125
±10b160
±17116±3fi111
±495
±14208±206105
±5118±3Normal88
±494±4104±3102±498
±387±399±396±2105
±6100±699±4100±5112
±496±497±6100±2110
±393±2100±499±5Host140
±13"191±1161
96 ±156155±12fi128±
4b190±1061
59 ±86136±6b130±
56327
±16217±14U163±5b146
±56184±136163±11Ò1
39 ±8°141
±46210±8Ò135±7*146±
5Hepatoma124
±4Ò124±4b138±
86135±Bb99
±11138 ±6b110
±4109±6133±
56139±10b1
74 ±20"165±5699
±7146±8°135±76126±1*108
±11172±7b113±46125± 46
* Mean ±S.E. of 8 animals, expressed as percentage of corresponding values in untreated animals from the same population studied concurrently, calculated to
3 significant figures, and then rounded.Significantly above the untreated control at the 5% confidence level.
JULY 1981 2695
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

L. D. Brebnor et al.
percentages of the levels in the livers of normal rats similarlytreated with hormone injections. It shows that treatment oftumor-bearing animals with dibutyryl cyclic AMP did not merely
restore the levels of urea cycle enzymes to the same levels asthe controls. It actually produced levels which were higher thanin hormone-treated controls, although the starting levels were
lower. Thus, the presence of the tumor allowed higher levels tobe attained than resulted from the same treatment of normalanimals. The same conclusion is generally true for the effect ofglucagon. /S-Methasone treatment resulted in levels of urea
cycle enzymes which were sometimes significantly above thosein similarly treated controls, sometimes comparable to them,and sometimes below.
The results of the above experiments indicate that the presence of the tumor affected urea cycle enzymes in the host liverin 2 ways: (a) the levels were lowered; and (o) the enzymesbecame more sensitive to hormonal induction so that a singledose of hormone could produce levels above those in normalanimals.
Effect of Plasma from Tumor-bearing Animals. It seems
likely that this effect was exercised through some factor produced by the tumor. In order to confirm this hypothesis and totest the means by which such a factor is conveyed from tumorto host liver, normal rats were given injections of plasma fromtumor-bearing animals (2 ml/rat). Control animals received a
similar volume of plasma from normal rats. Some animals fromboth groups also received a simultaneous injection of glucagon(5 ng/Q body weight). All animals were killed 24 hr after theinjections.
The results are shown in Chart 2. Injection of normal plasmahad no significant effect on the levels of any of the urea cycleenzymes. Similarly, an injection of glucagon with normal serumhad no effect. Injection of plasma from tumor-bearing animals
resulted in a lowering of specific activity of all 5 enzymes by 15to 40%. Simultaneous injection of glucagon together with theserum of tumor-bearing animals resulted in highly significant
Control
Normal Plasma
Hepatoma Plasma
Normal Plasma+Glucagon
Hepatoma Plasma +Glucagon
Chart 2. Effect of injection of plasma (2 ml/rat) from normal and tumor-bearing rats, with and without glucagon (5 /ig/g body weight) on the levels ofurea cycle enzymes in 5-week-old rats. Injections were given 24 hr beforesacrifice. Values shown are means of 8 animals with bars, S.E. They areexpressed as percentages of specific activities of uninjected controls. The valuesof 102 x specific activities (fimol/ min 7100 mg protein) of controls are shown
underneath the appropriate bars. Arg, arginase; ASS. argininosuccinate synthetase.
Table 4
Levels of urea cycle enzymes in normal human liver, primary hepatoma, and thenormal portion of the liver of hepatoma patients (host liver)
Values shown are 102 times the means of the specific activities of the number
of patients. All measurements were made on autopsied material.
CPSOTCASLArginaseNormal
(4)a3.17
±1.24o164
±4.100.32
±0.04265
±24.2Host
(4)1.84
±0.3476.3
±26.20.24
±0.12103
±23.7Hepatoma
(8)0.32
±0.1063.4
±12.40.21
±0.0516.5
± 3.8
Numbers in parentheses, number of patients.' Mean ±S.E.
8r
N
LU
O 1O 2O 3OTIME OF HORMONE TREATMENT^ hr)
Chart 3. Effect of Cortisol hemisuccinate (100 /ÃŒM)on the specific activities ofarginase and TAT in human and rat hepatoma cells in culture. Means of duplicatemeasurements are shown. A and A, 10 ' x arginase in rat hepatoma, Experiments1 and 2; D and •¿�,103 x arginase in human hepatoma. Experiments 1 and 2;3, C, and •¿�,105 x TAT in rat hepatoma. Experiments 1,2, and 3; O and O, 104
x TAT in human hepatoma, Experiments 1 and 2.
increases in the levels of all 5 urea cycle enzymes. Comparedto controls, the increases ranged from 27 to 50%. However, ifthe levels of animals given injections of tumor serum only areused for comparison, the increases in levels are much larger:79% for CPS; 78% for OTC; 156% for argininosuccinatesynthetase; 82% for ASL; and 99% for arginase. The results ofthe experiment are thus very similar to those in animals withtransplanted tumors. Urea cycle enzymes were lowered andrendered inducible by a single injection of glucagon, resultingin levels higher than controls.
2696 CANCER RESEARCH VOL. 41
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

Urea Cycle Enzyme Regulation in Tumor-bearing Rats and Humans
Regulation in Human Hepatoma and Host Liver. In order totest whether the phenomena seen in rat liver have a counterpartin human primary hepatoma, we measured the levels of ureacycle enzymes in autopsy livers from normal patients who diedof diseases not involving liver damage, in primary hepatomas,and in pieces of adjacent apparently uninvolved liver tissue.The results are shown in Table 4. They are, qualitatively, verysimilar to the results for tumor-bearing rats. The levels of urea
cycle enzymes in the hepatomas were measurable but lowerthan in normal liver. The levels in the host livers were significantly below those in normal liver and intermediate betweenthose of normal liver and hepatomas.
In order to study the inducibility of urea cycle enzymes inhuman hepatomas, it was necessary to obtain them in tissueculture. A tissue culture of the rat hepatoma was also testedfor comparison. Chart 3 shows the effect of addition of 100/¿Mcortisol hemisuccinate to the medium in which cells werecultured. In agreement with the results in vivo, arginase wasfound to be induced by glucocorticoid in the rat hepatoma. Inhuman hepatoma, too, arginase induction was observed. A lagin induction of 12 hr was seen in the rat hepatoma culture butnot in the human. The reason for this difference is unexplained.There is, however, a qualitative similarity between human andrat tumor. Surprisingly, TAT did not follow a similar pattern.While the enzyme was induced in rat hepatoma, it failed toshow any effect in the human tumor. The level of TAT in thehuman tumor was much higher than in the rat tissue. However,since units per 10s cells were used to express activity, this
comparison can only be approximate.
DISCUSSION
The present results are in agreement with earlier work (25)on the dietary and hormonal regulation of urea cycle enzymes.Though single doses of glucocorticoid or glucagon elevate thelevels of urea cycle enzymes in neonatal rats (10, 27), inductionof these enzymes in adult animals usually requires repeatedinjection for 2 to 7 days (15, 26, 29).
The regulatory behavior of the urea cycle enzymes in hepatoma DAB-1 is interesting because it presents a coherent
pattern which contrasts with the random diversity of levels andcontrols reported in many other tumors and stresses furtherthe correlation that exists among the 5 enzymes studied. Chemically induced rat hepatomas vary widely in their enzyme complements. Thus, a large range has been reported in levels ofTAT in Morris hepatomas (22, 24, 28), from very low to wellabove the uninduced normal level. CPS levels in rat hepatomasshow a similar variation and also include values which are at orabove the normal level (13, 18). OTC was also found to belower than in normal liver (18), but its level was not quantitatively correlated with that of CPS. The present work representsthe first study of the entire urea cycle. A functional urea cyclein mammals is specific for liver cells. Our present finding inhepatomas DAB-1 of all 5 urea cycle enzymes thus representsretention of a specific liver-like property.
The activities of the enzymes relative to each other weremarkedly altered in the hepatoma. For example, the ratio ofspecific activity levels of the 5 enzymes (CPS:OTC:arginino-succinate synthetase:ASL:arginase) in the liver of a rat eatinga 75% protein diet was 3.09:126:1.00:1.76:591. In the hepatoma of animals on a similar diet, the ratio was 1.88:57.9:1.00:
3.45:58.6. The comparatively low level of arginase in thehepatoma raises the possibility that the levels of ASL measuredin the hepatomas in the present experiments may be underestimates. The assay for ASL is a coupled one, involving endogenous arginase and the measurement of urea. The arginasewas present in 17- to 33-fold excess. Nuzum and Snodgrass
(17) found activity of ASL to be suboptimal under these conditions. It is possible, therefore, that the level of ASL reportedhere for hepatoma was lower than maximal. It seems veryunlikely, however, that, qualitatively, the regulatory effects ofdiet and hormones on ASL were artifacts due to differences inarginase concentration, since the 2 activities were not strictlycorrelated in different situations. Thus, the level of ASL on a75% protein diet, where the least excess of arginase over ASLwas present, was 2.6-fold higher compared to 22% protein;arginase was only 1.3-fold higher (Table 1). On the other hand,on a 10% protein diet, /S-methasone induced arginase 2.3-fold,but ASL was increased only 1.3-fold (Table 2). The responses
of ASL in hepatoma are fairly consistent in different experiments and on different diets, and they are probably at leastqualitatively correct.
The levels of CPS in host livers of other hepatomas studied(13, 18) were frequently, but not always, lower than in normallivers, and in many instances, they were below the levels in thehepatomas themselves. In fact, in Morris hepatomas 21, 28A,and 47C, Lawson ef al. (13) found a reciprocal relationshipbetween host and hepatoma levels of CPS, so that hepatomaswith high CPS produced low levels in host liver. The totalactivity per animal remained approximately constant, suggesting a possible homeostatic mechanism. No evidence of such asituation is seen in the present results, although it should bestressed that the levels of urea cycle enzymes are much lowerin DAB-1 than in the minimal deviation hepatomas studied by
Lawson era/. (13). All the OTC levels in the host livers examinedby Ono ef al. (18) were intermediate between those of normalliver and hepatoma.
An important observation in the present work is that thedifference in levels between normal and host liver was seen onall diets used. There was no correlation between the dietaryprotein content and the difference in level between normal andhost. This result suggests strongly that the effect of the tumorwas not through reduced availability of amino acids. If a deficiency of one or several amino acids, caused by their rapidutilization by the growing tumor, were involved in the effect, itshould have been diet dependent, being greatest on a low-protein diet and least on a high-protein diet.
Despite the lower levels, dietary regulation of urea cycleenzymes persists in hepatoma DAB-1. The dependence of TAT
and of serine dehydratase on dietary protein in the Morrishepatomas examined by Potter ef al. (21,22) varied from tumorto tumor. The majority of tumors showed no correlation withdietary protein; this was irrespective of whether the levels werehigh or low. A few tumors (9108, 9121) showed positivecorrelation for TAT (though not for serine dehydratase); onehepatoma (7793) showed a negative correlation at low dietaryprotein levels. Our results on DAB-1 suggest that the normal
mechanism of dietary regulation of the urea cycle is intact.Whatever control points are responsible for the lower levels ofthese enzymes in the tumor, they appear to be different fromthe dietary control points.
The sensitivity of urea cycle enzymes in hepatoma DAB-1 to
JULY 1981 2697
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

L. D. Brebnor et al.
induction by glucocorticoids, glucagon, and dibutyryl cyclicAMP was not only retained but enhanced, compared to normalliver. Most of the Morris hepatomas studied by Scott ef al. (28)had decreased sensitivity of TAT to glucagon; 9121, however,had enhanced sensitivity. Results most closely analogous tothe present ones were observed for glutamate:oxaloacetateaminotransferase in hepatoma 5123B (19) and for glutaminesynthetase in hepatoma 7800 (33), in that sensitivity to induction by cortisol was seen in the hepatoma and not in normalliver.
The present results on humans represent the first measurements of urea cycle enzymes in primary human hepatomas. Aserious shortcoming of the measurements is that they weremade on autopsy material taken some hr after death and storedfrozen. Urea cycle enzymes are fairly labile on storage; thereported values are therefore probably underestimates. Compared to the values published for biopsied and autopsied liver(17), our values for OTC are higher, those for CPS are higherthan for autopsied tissue (but equal to those in biopsied liver),the values for arginase are comparable, and those for ASL arelower. A further complication is that the terminal hepatomapatients were virtually unable to eat; this could have had theeffect of raising the levels of urea cycle enzymes, as occurs instarving rats (25). Possible cross-contamination of tumor and
host liver can also not be ruled out as the tissues were adjacentto one another. Despite the above reservations, it seems probable that, qualitatively, the differences between normal liver,host, and hepatoma are valid.
While nothing is known about hormonal regulation of theurea cycle in humans, our results are consistent with therebeing a similarity in regulation between rat and human. Theconclusions drawn from our results on the rat hepatoma aretherefore likely to have significance in relation to humans.
The induction of arginase in human hepatoma cells suggeststhat receptors for glucocorticoid were present. The failure toinduce TAT must therefore be due to an effect at some otherpoint. It should be pointed out here that the behavior of thehuman hepatoma cell lines does not necessarily indicate thatthe original hepatoma from which it was derived had also lostthe regulation of TAT. Cultured cells can differ in propertiesfrom their parent tumors, and the loss of control may haveoccurred after explantation.
The most interesting observation made in the course of thiswork was that the tumor caused the host liver to become moresensitive to induction by hormones. This result is unusual andshould be compared with some others. In host liver of ratsbearing hepatoma 7800, the inducibility of serine dehydratasewas decreased (8); that of TAT showed a temporary increasefollowed by a drop. Neither for glutamateioxaloacetate trans-
aminase in 5123B (19) nor for glutamine synthetase in 7800(32) was increased inducibility transmitted to the host liver.
This effect of increasing inducibility makes it unlikely that theeffect of tumor on host was mediated by increased productionof glucocorticoid by the host, which is known to be the effectof many tumors (7). A decreased level of enzymes coupled withincreased sensitivity to glucocorticoids is hardly consistent withsuch an assumption. Since it is known that greater sensitivityof some at least of the urea cycle enzymes to the hormones weused exists in neonatal and perinatal rats (10, 27), it is not animplausible view that this inducibility, like the lower levels,represents a reversion to a situation which existed at an earlier
developmental stage.Considerable evidence has accumulated which indicates that
the effect of the tumor on host is mediated by a humoral factor(12, 30). Earlier work has shown that such a factor may eitherraise or lower enzyme levels in host liver to values intermediatebetween normal liver and tumor. The present work shows thatit may also impart greater sensitivity to hormonal induction.The results are consistent with the view that its effect on theurea cycle enzymes represents a specific regulatory effectrather than a generalized depression of liver function.
ACKNOWLEDGMENTSI
The assistance of Ann Hartley in handling the animals is gratefully acknowledged.
REFERENCES
1. Aebi, J. Coordinated changes in enzymes of the ornithine cycle and responseto dietary conditions. In: S. Grisolia, R. Baguena, and F. Mayor (eds.). TheUrea Cycle, pp. 275-299. New York: Wiley & Sons. Inc., 1970.
2. Albrecht, C. F., Liebenberg. N. van de W.. Nourse, L. D., and Seegers, J. C.Some characteristics of a cell line derived from a fast-growing rat hepatoma.S. Afr. J. Med. Sci., 40: 35-42. 1975.
3. Balinsky, J. B., Chemaly, S. M., Currin, A. E., Thompson, R. L., and Van derWesthuizen, D. R. A comparative study of enzymes of urea and uric acidmetabolism in different species of amphibia and the adaptation to theenvironment of the tree frog Chiromantis xerampelina Peters. Comp. Bio-chem. Physiol.. 54B: 549-555. 1976.
4. Balinsky. J. B., Coetzer. T. L.. and Mattheyse, F. J. The effect of thyroxineand hypertonic environment on the enzymes of the urea cycle in Xenopuslaevis. Comp. Biochem. Physiol., 43B: 83-95, 1972.
5. Brebnor. L. D.. and Balinsky. J. B. Hormonal and nutritional effects on ureacycle enzymes in transplantable hepatomas and in the livers of tumor-bearing rats. Fed. Proc., 38: 504. 1979.
6. Christowitz, D., Mattheyse. F. J., and Balinsky, J. B. Dietary and hormonalregulation of urea cycle enzymes in rat liver. Enzyme (Basel), 26. in press,1981.
7. Costa, G. Cachexia: the metabolic component of neoplastic diseases. Prog.Exp. Tumor Res., 3. 321-369, 1963.
8. de Rosa, G., and Pilot, H. C. Alterations in enzymes of amino acid catabolismin livers of rats bearing the Morris 7800 hepatoma. Cancer Res., 38. 950-
954, 1978.9. Diamondstone. T. I. Assay of tyrosine transaminase activity by conversion of
p-hydroxyphenylpyruvate to p-hydroxybenzaldehyde. Anal. Biochem.. 16:395-401, 1966.
10. Greengard, O., Sahib, M. K., and Knox. W. E. Developmental formation anddistribution of arginase in rat tissues. Arch. Biochem. Biophys., J37. 477-
482. 1970.11. Grimm, J. The influence of insulin on various enzyme activities in human and
rat hepatoma cells. Eur. J. Biochem., 64: 249-253, 1976.12. Ishikawa, E., and Suda. M. Blood of tumor-bearing animals as a cause of
metabolic deviations. Gann Monogr.. )3. 289-299, 1972.13. Lawson, D., Paik, W. K.. Morris, H. P., and Weinhouse, S. Carbamyl
phosphate synthetases in rat liver neoplasms. Cancer Res.. 35. 156-163,
1975.14. Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall. R. J. Protein
measurement with the Folin phenol reagent. J. Biol. Chem.. J93. 265-275,
1951.15. McLean, P.. and Novello, F. Influence of pancreatic hormones on enzymes
concerned with urea synthesis in rat liver. Biochem. J., 94. 410-422, 1965.16. Mora, J., Tarrab, R., Martuscelli, *(., Ortiz-Pineda, J., and Soberón, G. The
regulation of urea-biosynthesis enzymes in vertebrates. Biochem. J. 96. 28-
35, 1965.17. Nuzum. A. T.. and Snodgrass, P. J. Multiple assays of the five urea-cycle
enzymes in human liver homogenates. In: S. Grisolia, R. Baguena, and F.Mayor (eds.). The Urea Cycle, pp. 325-349. New York: Wiley & Sons. Inc.,1970.
18. Ono, T.. Blair, G. R., Potter, V. R., and Morris, H. P. The comparativeenzymology and cell origin of rat hepatomas. IV. Pyrimidine metabolism inminimal-deviation tumors. Cancer Res.. 23. 240-249. 1963.
19. Otani, T. T., and Morris, H. P. The effect of glucocorticoids on the relativedistribution of aspartate aminotransferase isozymes of Morris hepatoma5123B. Cancer Res., 28. 2092-2097, 1968.
20. Potter, V. R.. Baril, E. F., Watanabe, M., and Whittle, E. D. Systematicoscillations in metabolic functions in liver from rats adapted to controlledfeeding schedules. Fed. Proc.. 27 1238-1244. 1968.
2698 CANCER RESEARCH VOL. 41
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

Urea Cycle Enzyme Regulation in Tumor-bearing Rats and Humans
21. Potter, V. R., Gebert, R. A., Pilot, H. C., Peraino, C., Lámar, C.. Jr., Leshner,S., and Morris, H. P. Systematic oscillations in metabolic activity in rat liverand in hepatomas. I. Morris hepatoma no. 7793. Cancer Res., 26. 1547-
1560, 1966.22. Potter. V. R.. Watanabe, M., Pilot, H. C.. and Morris. H. P. Systematic
oscillations in metabolic activity in rat liver and hepatomas. Survey of normaldiploid and other hepatoma lines. Cancer Res., 29. 55-78. 1969.
23. Prozesky, O. W., Brits, C.. and Grabow, W. O. K. In vitro culture of cell linesfrom Australia antigen positive and negative hepatoma patients. In: S. J.Saunders and J. Terblanche (eds.), Liver: Proceedings of the InternationalLiver Conference with Special Reference to Africa, pp. 358-360. London:Pitman Medical Publishing Co.. Ltd.. 1973.
24. Reynolds, R. D., Scott, D. F., Potter, V. R., and Morris, H. P. Induction oftyrosine aminotransferase and amino acid transport in Morris hepatomasand in adult and neonatal rat liver. Cancer Res., 31: 1580-1589, 1971.
25. Schimke, R. T. Adaptive characteristics of urea cycle enzymes in the rat. J.Biol. Chem.. 237 459-468, 1962.
26. Schimke, R. T. Studies on factors affecting the levels of urea cycle enzymesin rat liver. J. Biol. Chem., 238. 1012-1018, 1963.
27. Schwartz, A. L. Influence of glucagon, 6-/V,2'-O-dibutyryladenosine 3':5'-
cyclic monophosphate and triamcinolone on the arginine synthetase systemin perinatal rat liver. Biochem. J., r 26 89-98, 1972.
28. Scott, D. F., Butcher, F. R.. Potter. V. R., and Morris. H. P. Naturallyoccurring and induced levels of amino acid transport and tyrosine aminotransferase in rats bearing Morris hepatomas. Cancer Res., 32. 2127-2134.1972.
29. Snodgrass, P. J., Lin, R. C., Muller. W. A., and Aoki, T. J. Induction of ureacycle enzymes of rat liver by glucagon. J. Biol. Chem., 253: 2748-2753,1978.
30. Suda. M., Tanaka. T.. Sue, F.. Kuroda. K., and Morimura, H. Rapid increaseof pyruvate kinase (M type) and hexokinase in normal rat liver by perfusionof the blood of tumor bearing rat. Gann Monogr., 4: 103-112, 1968.
31. Weber. G. The molecular correlation concept. Studies on the metabolicpattern of hepatomas. Gann Monogr.. 7. 151-178, 1966.
32. Wicks, W. D., Kenney, F. T.. and Lee. K-L. Induction of hepatic enzymesynthesis in vivo by adenosine 3'.5'-monophosphate. J. Biol. Chem., 244.6008-6013, 1969.
33. Wu, C., and Morris. H. P. Responsiveness of glutamine-metabolizing enzymes in Morris hepatomas to metabolic modulations. Cancer Res., 30:2675-2684, 1970.
JULY 1981 2699
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from

1981;41:2692-2699. Cancer Res Linda D. Brebnor, Johanna Grimm and John B. Balinsky HumansHepatomas and in the Livers of Tumor-bearing Rats and Regulation of Urea Cycle Enzymes in Transplantable
Updated version
http://cancerres.aacrjournals.org/content/41/7/2692
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/41/7/2692To request permission to re-use all or part of this article, use this link
Research. on November 18, 2018. © 1981 American Association for Cancercancerres.aacrjournals.org Downloaded from