regulation of kinase and intermolecular bonding in intact and
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1986 by The American Society of Biological Chemists, Inc.
Vol. 261. No. 27, Issue of September 25, pp. 12879-12882,1986 Printed in U.S.A.
Regulation of Kinase and Intermolecular Bonding in Intact and Truncated Epidermal Growth Factor Receptor*
(Received for publication, April 22, 1986)
Mitali Basu, Anis Sen-Majumdar, Aruna Basu, Uma Murthy, and Manjusri Das From the Department of Biochemistry and Biophysics, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania 19104
Tyrosine kinase activity of the epidermal growth factor (EGF) receptor can be regulated by its state of association. Studies done with the purified receptor solubilized in Triton X-100 indicate that dimer for- mation results in negative regulation of kinase, whereas successive binding of EGF and ATP shift the association equilibrium toward the catalytically active monomeric form. The promotion of the monomeric state by ATP can be mimicked by various nonphos- phorylating analogs indicating that nucleotide binding rather than autophosphorylation is responsible for sta- bilizing the monomeric receptor form. Truncated re- ceptor forms, lacking either the external EGF-binding domain or the internal kinase (ATP-binding) domain, are unable to form stable dimers. These results suggest that both intra- and extracellular domains of the re- ceptor act to stabilize the kinase-regulatory dimer.
The epidermal growth factor (EGF’) receptor is a trans- membrane mitogenic glycoprotein with a cytoplasmically fac- ing tyrosine kinase site that is activated by growth factor binding to the external domain (Cohen et al., 1982). This receptor represents the product of the c-erb-B proto-oncogene (Ullrich et al., 1984). Its cytoplasmic domain shows extensive homology with the tyrosine kinase family of proteins, espe- cially the neu oncogene protein (Bargmann et al., 1986). Tyrosine kinases are intimately involved in growth control and cellular proliferation (Hunter and Cooper, 1985). In the case of EGF receptor and other growth factor receptors, the expres- sion of kinase activity is dependent upon the presence of specific growth factors, whereas no such regulation is seen with the viral oncogene-encoded tyrosine kinases.
Our previous in vitro studies suggested that the 170,000 dalton EGF receptor behaves like an allosteric enzyme that can interconvert between active (monomeric) and inactive (dimeric) forms, and that EGF activates the kinase by shifting the equilibrium toward the catalytically active monomeric form (Biswas et al. 1985). Here we report further experiments concerning the role of ATP in the control of receptor associ- ation-dissociation, the additivity of ATP and EGF effects, and the behavior of truncated receptor forms lacking either the external EGF-binding domain or the internal ATP-bind- ing domain.
* This work was supported by National Institutes of Health Re- search Grants CA-43787 and HD-17896. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduerttsernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The abbreviations used are: EGF, epidermal growth factor; BSA, bovine serum albumin; TLCK, l-chloro-3-tosylamido-7-amino-2- heptanone; Hepes, 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid.
MATERIALS AND METHODS AND RESULTS’
DISCUSSION
The experiments described here have produced the follow- ing results: (a ) In the detergent preparations tested, ATP controls the association state of the EGF receptor by stabiliz- ing the monomeric form (Fig. 1); (b) autophosphorylation does not appear to be necessary for this process (Fig. 2); (c) the effect of ATP is seen over and above that seen with EGF alone (Fig. 3), suggesting the involvement of both external and internal domains of the receptor in inter-receptor bond- ing; ( d ) loss of either the external EGF-binding domain or the internal kinase domain results in a loss of inter-receptor bonding stability (Figs. 4-6).
The simplest interpretation of our results is pictured in Fig. 7. The EGF receptor pre-exists mainly as a dormant dimer, in which the kinase site (dotted in Fig. 7 ) is inaccessible for binding to ATP, and the EGF-binding site (hatched in the figure) has only reduced affinity for binding to EGF. The key to opening this dimer lies not with ATP but with EGF (see below). One can picture that, in the presence of EGF (Fig. 7), a half-opened receptor form with the external domains sepa- rated is generated. This form can reversibly equilibrate with the fully open monomeric receptor form. The monomer form is then stabilized by ATP binding, which tends to block reassociation of the intracellular domains. The active mon- omer form can revert to the dormant dimer after loss of EGF and ATP. We have documented the conversion of the mon- omer to the dimer in vitro with EGF-affinity-purified receptor. Even in this detergent-solubilized system where the receptor concentration is relatively low-far lower effective concentra- tion than that in the two-dimensional structure of plasma membranes and endosomes-the receptor dimerizes within a few hours. Although kinetics measured in solution (three- dimensional) are not directly translatable to the in vivo mem- brane system (two-dimensional), the rate of dimerization is likely to be much faster in the membrane (see Adam and Delbruck, 1968).
Earlier work in other laboratories has shown that pretreat- ment of various tyrosine kinases with ATP results in an enhanced activity toward exogenous substrates. Three differ- ent mechanisms have been proposed to explain this phenom-
Portions of this paper (including “Materials and Methods,” “Re- sults,” and Figs. 1-7) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biolog- ical Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 86M-1323, cite the authors, and include a check or money order for $4.00 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
12879

12880 Dimerization of EGF Receptor
enon: ( a ) The catalytic efficiency of the kinase is greater in the autophosphorylated form than in the nonphosphorylated form (Rosen et al., 1983). (b ) The self-phosphorylation site in the kinase acts as a competitive inhibitor (alternate substrate) against exogenous substrates, and prior self-phosphorylation with ATP removes this competitive constraint (Bertics and Gill, 1985). (c) ATP/nucleotide binding to the kinase protects the catalytically active site (Sugimoto et al., 1985), and sta- bilizes the active form of the kinase. The results presented here on the EGF receptor kinase are most consistent with the last alternative. In our system, however, the effect of ATP is not due to direct stabilization of the kinase site itself but, at the first level, to stabilization of the monomeric receptor form. The monomeric form, for reasons yet to be found (perhaps due to unmasking of the kinase site), promotes kinase activity.
Results available indicate that ATP can exert its effect only after the receptor has seen EGF, and that the key to opening the dormant dimer lies not with ATP but with EGF. Our affinity labeling studies with A431 membranes indicate that the ATP analog fluorosulfonylbenzoyl ['4C]adenosine (Buhrow et al., 1982; Basu et al., 1984) binds to and labels the receptor only after EGF addition, and that there is negligible labeling in the absence of EGF.3 A similar result can be seen in data published by Buhrow et al. (1982).
It is of interest to compare our results on the effect of receptor association on kinase catalytic activity with the results of Yarden and Schlessinger (1985). Their studies on the effect of receptor aggregation on kinase catalytic activity were, as in our case, conducted using detergent-solubilized receptor, and not with whole cells or membranes. Yarden and Schlessinger (1985) found the autokinase activity of solubi- lized EGF receptor to be stimulated by various bivalent anti- receptor antibodies (directed to either the external or the internal domain) but not by their monovalent counterparts. The authors concluded that receptor aggregation can activate its kinase site. It should be appreciated that, although mono- meric receptors can be cross-linked into aggregates by anti- bodies and other polyvalent reagents, these associations must be quite different from the specific dimer discussed here, and in general will be inconsistent with such structures. It seems to us that the activating effect of antibodies in the Yarden and Schlessinger experiments, rather than reflecting an acti- vation dependent upon nonspecific aggregation (held together by intervening antibody), may quite possibly have operated by physically blocking the binding necessary for forming the highly specific dimer structure discussed here.
When one considers that in our system the intact EGF receptor readily dimerizes in solution, the lack of this ability in the two truncated receptor forms is interesting. The results suggest that for stable bonding to occur, both external and internal domains must be present as parts of the same mole- cule. Thus, loss of the EGF-binding domain can not only
M. Basu and M. Das, unpublished experiments.
abolish positive regulation by EGF, but may in itself abrogate the structural basis for negative regulation of kinase activity. It can be noted that another tyrosine kinase, pp60"", which lacks any extracellular ligand-regulated domain, appears to sediment only as a monomer (Sugimoto et al., 1985). It will be of interest to determine whether the v-erb-B kinase, the truncated and oncogenic homolog of EGF receptor, can form stable dimers, and if not, whether the lack of self-association ability is responsible for the constitutive activity and un- regulated nature of its kinase site.
Acknowledgments-We thank Laura Mattes for preparing A431 plasma membranes and Dr. Walter Englander for helpful discussions.
REFERENCES Adam, G., and Delbruck, M. (1968) in Structural and Moleculur
Biology (Rich, A., and Davidson, N., eds) pp. 198-215, W. H. Freeman & Co., San Francisco
Bargmann, C. I., Huang, M. C., and Weinberg, R. A. (1986) Nature
Basu, M., Biswas, R., and Das, M. (1984) Nature 311 , 477-480 Bertics, P. J., and Gill, G. N. (1985) J. Biol. Chem. 260,14642-14647 Biswas, R., Basu, M., Sen-Majumdar, A., and Das, M. (1985) Bio-
Buhrow, S. A., Cohen, S., and Staros, J. V. (1982) J. Biol. Chem.
Cohen, S., Ushiro, H., Stoscheck, C., and Chinkers, M. (1982) J. Biol. Chem. 257,1523-1539
Das, M., Miyakawa, T., Fox, C. F., Pruss, R. M., Aharonov, A., and Herschman, H. R. (1977) Proc. Natl. Acad. Sci. U. S. A. 74,2790- 2794
Das, M., Biswas, R., Basu, M., and Bishayee, S.. (1984a) Cancer Res. 44,3539-3545
Das, M., Knowles, B., Biswas, R., and Bishayee, S. (1984b) Eur. J. Biochen. 14 1,429-434
Hunter, T., and Cooper, J. A. (1985) Annu. Reu. Biochem. 5 4 , 897- 930
Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol. Chem. 193,265-275
Martin, R. G., and Ames, B. N. (1961) J. Biol. Chem. 2 3 6 , 1372- 1379
Murthy, U., Basu, M., Sen-Majumdar, A., and Das, M. (1986) J. Cell. Bid . 103,333-342
Pike, L. J., Gallis, B., Casnellie, J. E., Bornstein, P., and Krebs, E. G. (1982) Proc. Natl. Acad. Sci. U. S. A. 79, 1443-1447
Rosen, D. M., Herrera, R., Olowe, Y., Petruzzelli, L. M., and Cobh,
Savage, C. R., Jr., Cohen, S. (1972) J. Biol. Chem. 247,7609-7611 M. (1983) Proc. Natl. Acad. Sci. U. S. A. 80,3237-3240
Sen-Majumdar, A., Murthy, U., Chianese, D., and Das, M. (1986) Biochemistry 25,634-640
Sugimoto, Y., Erikson, E., Graziani, Y., and Erikson, R. L. (1985) J. Biol. Chem. 260,13838-13843
Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci. U. S. A. 7 6 , 4350-4354
Ullrich, A., Coussens, L., Hayflick, J. S., Dull, T. J., Gray, A., Tam, A. W., Lee, J., Yarden, Y., Lieberman, T. A., Schlessinger, J.,
Seeburg, P. H. (1984) Nature 309 , 418-425 Downward, J., Mayes, E. L. V., Whittle, N., Waterfield, M. D., and
Yarden, Y., and Schlessinger, J. (1985) in Growth Factors in Biology & Medicine, Ciba Foundation Symposium 116, pp. 23-45, Pitman Press, London
319,226-230
chemistry 24,3795-3802
257,4019-4022
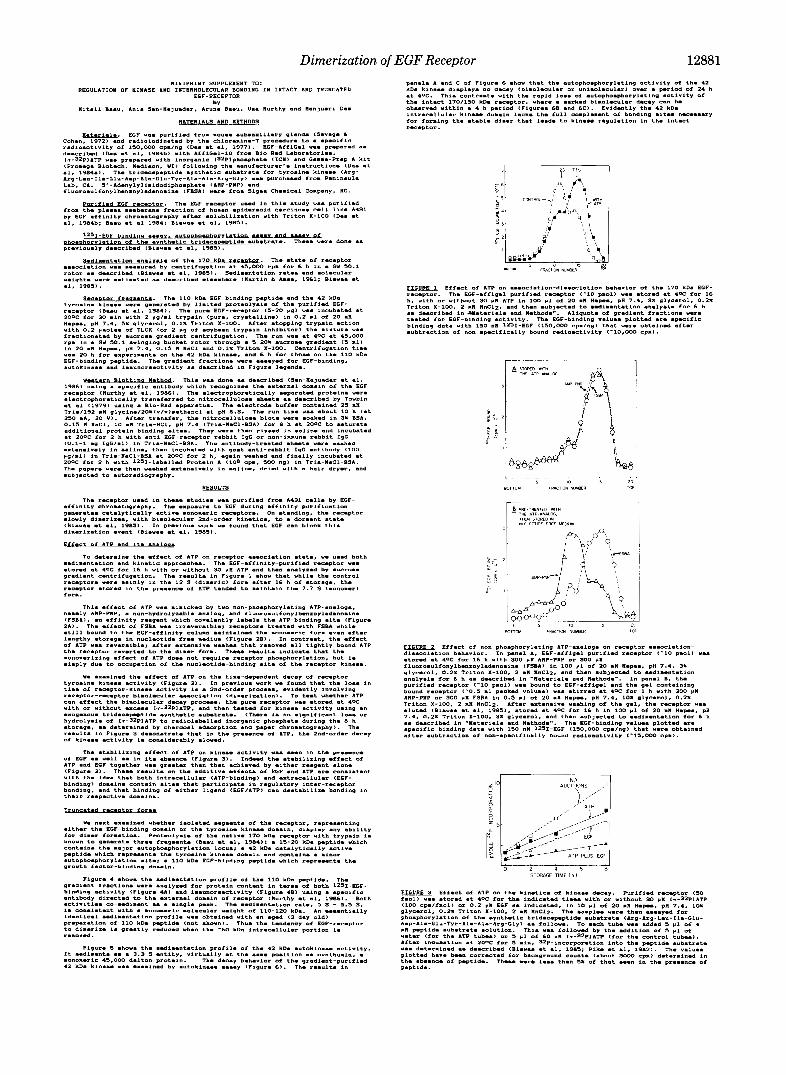
Dimerization of EGF Receptor 12881

12882 Dimerization of EGF Receptor
B. 15
(1
m
a
5
j
6
5
9
42K- 45
e9
A.
HOURS- o 2 4 8 24
- 205
-116 - 93
- 67
42K- - - - I -45
-29
B. HOURS- 0 2 4
170K, 150K ' - - -116
- 93
- 6 7
-205
- 45
-29