regulation of gene expression: this is material for the final exam 2013 the following sections of...
TRANSCRIPT

Regulation of Gene Expression:
• This is material for the final exam 2013
• The following sections of your textbook apply to this material:
• A. chapter 18-Sections 18.1-18.3 (351-366)
• B. chapter 12.3 (238-243)
• C. Chapter 18-Section 18.5 (373-77)

Overview: Conducting the Genetic Orchestra
• Prokaryotes and eukaryotes alter gene expression in response to their changing environment
• In multicellular eukaryotes, gene expression also regulates development and is responsible for differences in cell types
• RNA molecules play many roles in regulating gene expression in eukaryotes
• Mistakes in the regulation of gene expression can lead to disease (Cancer)

Bacteria often respond to environmental change by regulating transcription
• Natural selection has favored bacteria that produce only the products needed by that cell
• A cell can regulate the production of enzymes by feedback inhibition or by gene regulation
• Which of these forms of regulation saves more energy? Which can respond to changes more quickly?

LE 18-20
Regulation of enzymeactivity
Regulation of enzymeproduction
Enzyme 1
Regulation of gene expression
Enzyme 2
Enzyme 3
Enzyme 4
Enzyme 5
Gene 2
Gene 1
Gene 3
Gene 4
Gene 5
Tryptophan
Precursor
Feedbackinhibition

The Operon Model
• Gene expression in bacteria is controlled by the operon model
• Jacob and Monod (1961)
• Operon elements-repressor gene; operator; structural gene(s)
• Inducible and repressible operons

Inducible operon (lac operon)
• Activates breakdown of a substrate when the substrate becomes available.
• Normal status-transcription does not occurs because the operator is blocked by the repressor protein.
• Substrate (inducer) alters shape of the repressor protein so that does not bind to the operator allowing transcription of the structural genes to occur
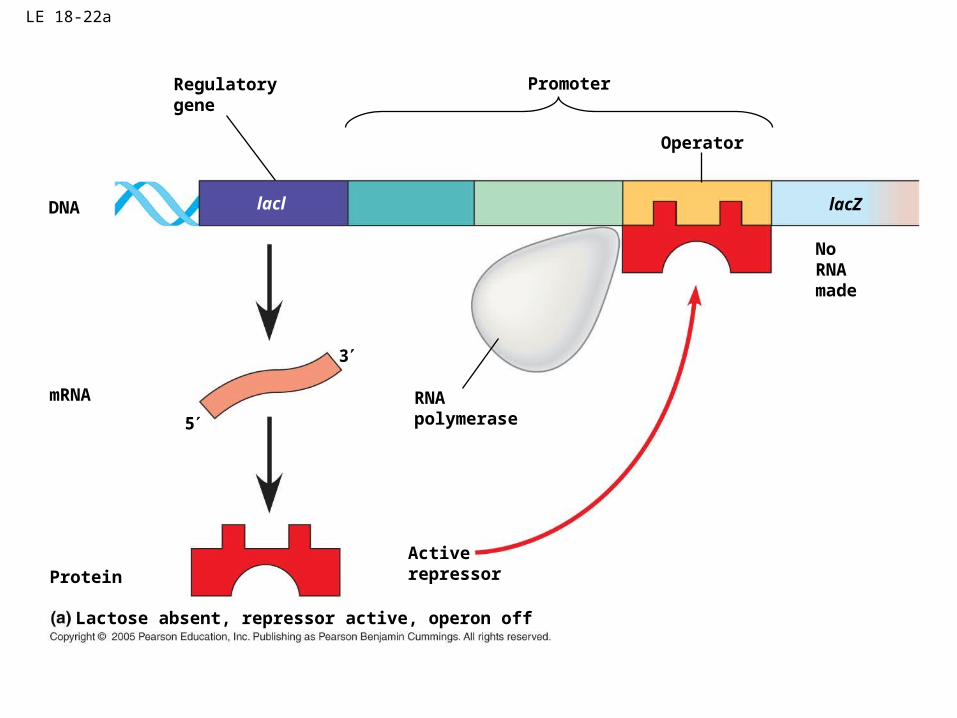
LE 18-22a
DNA lacl
Regulatorygene
mRNA
5
3
RNApolymerase
ProteinActiverepressor
NoRNAmade
lacZ
Promoter
Operator
Lactose absent, repressor active, operon off

LE 18-22b
DNA lacl
mRNA5
3
lac operon
Lactose present, repressor inactive, operon on
lacZ lacY lacA
RNApolymerase
mRNA 5
Protein
Allolactose(inducer)
Inactiverepressor
-Galactosidase Permease Transacetylase

Another form of regulation of the LAC operon
• The lac operon can be turned on or off by the presence/absence of the inducer molecule (lactose)
• CAP and CAMP can also regulate the rate of transcription of the lac operon (volume control versus on/off control)
• If both glucose and lactose are present, the bacteria will preferentially use glucose

LE 18-23a
DNA
cAMP
lacl
CAP-binding site
Promoter
ActiveCAP
InactiveCAP
RNApolymerasecan bindand transcribe
Operator
lacZ
Inactive lacrepressor
Lactose present, glucose scarce (cAMP level high): abundant lacmRNA synthesized

LE 18-23b
DNA lacl
CAP-binding site
Promoter
RNApolymerasecan’t bind
Operator
lacZ
Inactive lacrepressor
InactiveCAP
Lactose present, glucose present (cAMP level low): little lacmRNA synthesized

The repressible operon (trp operon)
• Shuts down a biochemical pathway when the end product of the pathway build up.
• Normal status-transcription occurs because the operator is unblocked.
• End product of the pathway (co-repressor) alters shape of the repressor protein so that it binds to the operator blocking transcription of the structural genes

LE 18-21a
Promoter Promoter
DNA trpR
Regulatorygene
RNApolymerase
mRNA
3
5
Protein Inactiverepressor
Tryptophan absent, repressor inactive, operon on
mRNA 5
trpE trpD trpC trpB trpA
OperatorStart codonStop codon
trp operon
Genes of operon
E
Polypeptides that make upenzymes for tryptophan synthesis
D C B A

LE 18-21b_1
DNA
Protein
Tryptophan(corepressor)
Tryptophan present, repressor active, operon off
mRNA
Activerepressor

LE 18-21b_2
DNA
Protein
Tryptophan(corepressor)
Tryptophan present, repressor active, operon off
mRNA
Activerepressor
No RNA made

Differential Gene Expression in Eukaryotes
• Almost all the cells in an multicellular organism are genetically identical
• Differences between cell types result from differential gene expression, the expression of different genes by cells with the same genome
• In a given cell, usually about 20% of its genes are active at a given time (rest turned off)
• Errors in gene expression can lead to diseases including cancer
• Gene expression is regulated at many stages in eukaryotes (in addition to regulation at the transcriptional level which bacteria also do)
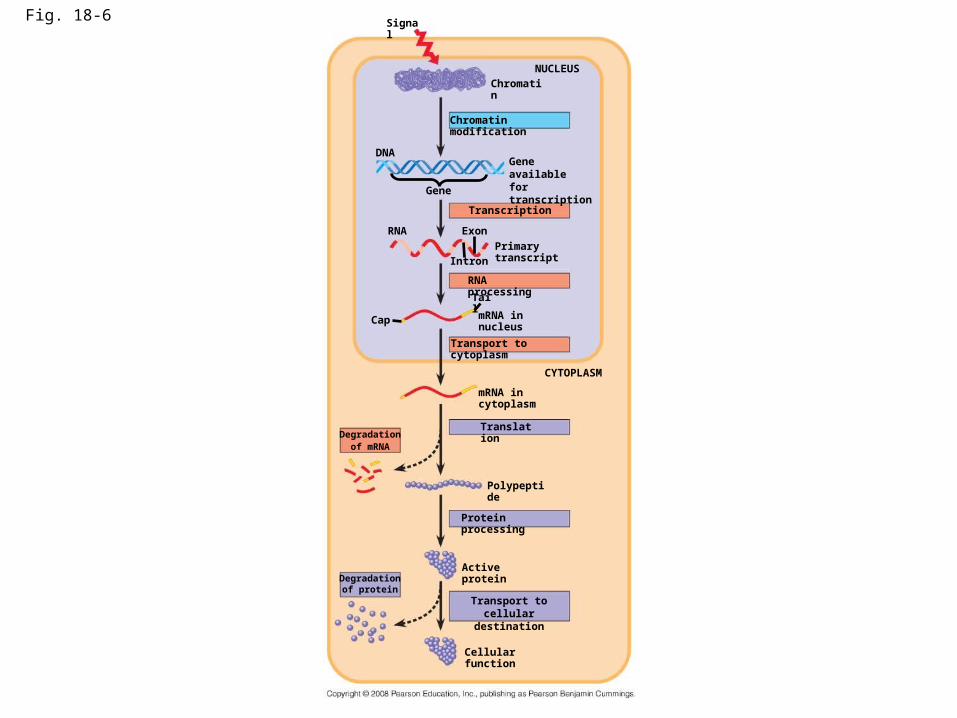
Fig. 18-6
DNA
Signal
Gene
NUCLEUS
Chromatin modification
Chromatin
Gene availablefor transcription
Exon
Intron
Tail
RNA
Cap
RNA processing
Primary transcript
mRNA in nucleus
Transport to cytoplasm
mRNA in cytoplasm
Translation
CYTOPLASM
Degradationof mRNA
Protein processing
Polypeptide
Active protein
Cellular function
Transport to cellulardestination
Degradationof protein
Transcription

Regulation of chromatin Structure:
• DNA is packed into an elaborate complex known as chromatin
• The basic unit of chromtain is the nucleosome• Histone proteins help maintain nucleosome
structure

Fig. 16-21a
DNA double helix (2 nm in diameter)
Nucleosome(10 nm in diameter)
Histones Histone tailH1
DNA, the double helix Histones Nucleosomes, or “beads on a string” (10-nm fiber)

Fig. 16-21b
30-nm fiber
Chromatid (700 nm)
Loops Scaffold
300-nm fiber
Replicated chromosome (1,400 nm)
30-nm fiber Looped domains (300-nm fiber)
Metaphase chromosome

Chromatin Packing in Eukaryotes
• Loosely packed DNA (euchromatin) is usually transcribed
• Tightly packed DNA (heterochromatin) is not usually transcribed

Regulation of Chromatin Structure
• Chemical modifications to histones and DNA of chromatin influence both chromatin structure (tightly packed versus loosely packed) and gene expression
• Chemical modification of histones include acetylation and methylation
• Chemical modification of DNA include methylation

Histone Modifications
• In histone acetylation, acetyl groups are attached to positively charged lysines in histone tails
• This process loosens chromatin structure, thereby promoting the initiation of transcription
• The addition of methyl groups (methylation) can condense chromatin;

Fig. 18-7
Histonetails
DNAdouble helix
(a) Histone tails protrude outward from a nucleosome
Acetylated histones
Aminoacidsavailablefor chemicalmodification
(b) Acetylation of histone tails promotes loose chromatin structure that permits transcription
Unacetylated histones

DNA Methylation
• DNA methylation, the addition of methyl groups to certain bases in DNA, is associated with reduced transcription in some species
• DNA methylation can cause long-term inactivation of genes in cellular differentiation
• Barr Bodies

Regulation of Transcription Initiation
• Chromatin-modifying enzymes provide initial control of gene expression by making a region of DNA either more or less able to bind the transcription machinery
• Regulation of transcription by enhancers (and their associated control elements) is another important mechanism by which initiation of transcription can be regulated.

Organization of a Typical Eukaryotic Gene
• Associated with most eukaryotic genes are control elements, segments of noncoding DNA that help regulate transcription by binding certain proteins
• Control elements and the proteins they bind are critical to the precise regulation of gene expression in different cell types

Fig. 18-8-1
Enhancer(distal control elements)
Proximalcontrol elements
Poly-A signalsequence
Terminationregion
DownstreamPromoter
UpstreamDNA
ExonExon ExonIntron Intron

The Roles of Transcription Factors
• To initiate transcription, eukaryotic RNA polymerase requires the assistance of proteins called transcription factors
• General transcription factors are essential for the transcription of all protein-coding genes
• If a gene is going to be transcribed at anything but a slow rate, specific transcription factor must also be involved.
• In eukaryotes, high levels of transcription of particular genes depend on control elements interacting with specific transcription factors

Fig. 18-8-1
Enhancer(distal control elements)
Proximalcontrol elements
Poly-A signalsequence
Terminationregion
DownstreamPromoter
UpstreamDNA
ExonExon ExonIntron Intron

• An activator is a protein that binds to a control element of an enhancer and stimulates transcription of a gene
• Bound activators cause mediator proteins to interact with proteins at the promoter
Enhancers and Specific Transcription Factors

Fig. 18-9-3
Enhancer TATAbox
PromoterActivators
DNAGene
Distal controlelement
Group ofmediator proteins
DNA-bendingprotein
Generaltranscriptionfactors
RNApolymerase II
RNApolymerase II
Transcriptioninitiation complex RNA synthesis

• Some transcription factors function as repressors, inhibiting expression of a particular gene
• Some activators and repressors act indirectly by influencing chromatin structure to promote or silence transcription

Why does a liver cell produce albumin and a lens cell crystalline protein?
• All activators needed for high-level expression of the albumin gene are present in liver cells but not lens cells.
• All activators needed for high-level expression of the crystalline gene are present in lens cells but not liver cells.
• The presence of activators in cells may occur at precise times during development or in a particular cell type

Fig. 18-10
Controlelements
Enhancer
Availableactivators
Albumin gene
(b) Lens cell
Crystallin geneexpressed
Availableactivators
LENS CELLNUCLEUS
LIVER CELLNUCLEUS
Crystallin gene
Promoter
(a) Liver cell
Crystallin genenot expressed
Albumin geneexpressed
Albumin genenot expressed

Noncoding RNAs play multiple roles in controlling gene expression
• Only a small fraction of DNA codes for proteins, rRNA, and tRNA (less than 2%)
• A significant amount of the genome may be transcribed into noncoding RNAs
• Noncoding RNAs regulate gene expression at the level of translation by degrading M-rna or blocking its translation
• 2 important types of noncoding RNA include MicrorNA (miRNA) and small interfering RNA (siRNA).

Fig. 18-13
miRNA-proteincomplex(a) Primary miRNA transcript
Translation blocked
Hydrogenbond
(b) Generation and function of miRNAs
Hairpin miRNA
miRNA
Dicer
3
mRNA degraded
5

Effects on mRNAs by MicroRNAs and Small Interfering RNAs
• MicroRNAs (miRNAs) are small single-stranded RNA molecules that can bind to mRNA
• These can degrade mRNA or block its translation

• The phenomenon of inhibition of gene expression by RNA molecules is called RNA interference (RNAi)
• RNAi is caused by small interfering RNAs (siRNAs) which are a type of miRNA
• RNAi may represent the evolution of a genetic control system from a defense mechanism against double stranded RNA virus infection

What are two different methods that bacteria use to regulate gene expression?
• Availability of sigma factors (genes that encode sigma factors can be induced by environmental signals)
• DNA binding proteins-repressor protein in lac operon; activator protein (auto-inducer in quorum sensing)

What happens if a bacterium receives an environmental signal for motility?
• Environmental signal activates gene for the motility associated sigma factor.
• Motility sigma factors interact with RNA polymerase and help them bind to the promotor of the 25-30 motility proteins
• Flagella are produced.

What is quorum sensing?
• What advantage doe the luminescent bacteria get from its symbiotic association with its host?
• What advantage doe the host obtain from its symbiotic association with the luminescent bacteria?
• Why shouldn’t the symbiotic bacteria express the luminescence gene until there is a critical number of luminescent bacteria present?
• How do luminescent bacteria “know” that there are a critical number of bacteria present so they should all begin expressing the luminescing gene?

The Molecular Basis of cancer

The eukaryotic cell cycle is regulated by a molecular control system
• The frequency of cell division varies with the type of cell
• These cell cycle differences result from regulation at the molecular level
• Understanding the regulation of the cell cycle at the molecular level helps us to understand how cancer cells escape the usual controls and divide indefinitely
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

The Cell Cycle Control System
• The sequential events of the cell cycle are directed by a distinct cell cycle control system, which is similar to a clock
• The clock has specific checkpoints where the cell cycle stops until a go-ahead signal is received
• Checkpoints can be influenced by signals from outside the cell as well as inside the cell.
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-14
SG1
M checkpoint
G2M
Controlsystem
G1 checkpoint
G2 checkpoint

• For many cells, the G1 checkpoint seems to be the most important one
• If a cell receives a go-ahead signal at the G1 checkpoint, it will usually complete the S, G2, and M phases and divide
• If the cell does not receive the go-ahead signal, it will exit the cycle, switching into a nondividing state called the G0 phase
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-15
G1
G0
G1 checkpoint
(a) Cell receives a go-ahead signal
G1
(b) Cell does not receive a go-ahead signal

The Cell Cycle Clock: Cyclins and Cyclin-Dependent Kinases
• Two types of regulatory proteins are involved in cell cycle control: cyclins and cyclin-dependent kinases (Cdks)
• The activity of cyclins and Cdks fluctuates during the cell cycle
• MPF (maturation-promoting factor) is a cyclin-Cdk complex that triggers a cell’s passage past the G2 checkpoint into the M phase
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-17
M G1S G2
M G1S G2
M G1
MPF activity
Cyclinconcentration
Time(a) Fluctuation of MPF activity and cyclin concentration during the cell cycle
Degradedcyclin
Cdk
G 1S
G 2
M
CdkG2
checkpointCyclin isdegraded
CyclinMPF
(b) Molecular mechanisms that help regulate the cell cycle
Cy
clin
ac
cu
mu
latio
n

Stop and Go Signs: Internal and External Signals at the Checkpoints
• An example of an internal signal is that kinetochores not attached to spindle microtubules send a molecular signal that delays anaphase
• Some external signals are growth factors, proteins released by certain cells that stimulate other cells to divide
• For example, platelet-derived growth factor (PDGF) stimulates the division of human fibroblast cells in culture
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-18
Petriplate
Scalpels
Cultured fibroblasts
Without PDGFcells fail to divide
With PDGFcells prolifer-ate
10 µm

• Another example of external signals is density-dependent inhibition, in which crowded cells stop dividing
• Most animal cells also exhibit anchorage dependence, in which they must be attached to a substratum in order to divide
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-19
Anchorage dependence
Density-dependent inhibition
Density-dependent inhibition
(a) Normal mammalian cells (b) Cancer cells25 µm25 µm

• Cancer cells exhibit neither density-dependent inhibition nor anchorage dependence
• This is why they can grow out of control to form a mass and invade surrounding tissue
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Loss of Cell Cycle Controls in Cancer Cells
• Cancer cells do not respond normally to the body’s control mechanisms
• Cancer cells may not need growth factors to grow and divide. Why?– They may make their own growth factor– They may convey a growth factor’s signal
without the presence of the growth factor– They may have an abnormal cell cycle control
system
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

• A normal cell is converted to a cancerous cell by a process called transformation
• Cancer cells form tumors, masses of abnormal cells within otherwise normal tissue
• If abnormal cells remain at the original site, the lump is called a benign tumor
• Malignant tumors invade surrounding tissues and can metastasize, exporting cancer cells to other parts of the body, where they may form secondary tumors
Copyright © 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings

Fig. 12-20
Tumor
A tumor growsfrom a singlecancer cell.
Glandulartissue
Lymphvessel
Bloodvessel
Metastatictumor
Cancercell
Cancer cellsinvade neigh-boring tissue.
Cancer cells spreadto other parts ofthe body.
Cancer cells maysurvive andestablish a newtumor in anotherpart of the body.
1 2 3 4

Cancer results from genetic changes that affect cell cycle control
• In cancer cells, checkpoints that normally stop cell division no longer function effectively.
• Cancer cells in tissue culture can essentially grow forever (immortal). Normal cells only divide 20-50 times in culture before they stop
• HeLa cells (growing continuously in culture since 1951)

Review-The Normal Cell Cycle-
• The Cell cycle starts and stops at appropriate time for a given cell type. However, the cell cycle can be altered by stimuli from the environment.
• The molecular basis that controls the cell cycle and determines whether the cycle moves past key checkpoints (G-1 for example) is determined by the rising and falling level of key proteins including cyclin, cyclin dependent kinases (cdk’s) and MPF ( a complex of cyclin and cdk’s). The expression of the proteins that regulate the cell cycle is subject to factors that regulate the expression of these genes (including signals from outside the cell such as growth factors).

An example of changes in the Cell Cycle:
• Assume a skin cell is not dividing (it hasn’t made it past the G-1 checkpoint in the cell cycle). Why?
• The levels of cyclin/cdk/MPF are not correct to move the cell past that checkpoint.
• Now-suppose the skin is damaged.
• Platelets in the blood release PDGF (platelet derived growth factor). PGDF binds to a receptor on skin cells, this sends a signal to the cell’s DNA to alter the expression of cell cycle regulating proteins (cyclins/cdk’s/MPF), and now the cell cycle moves past the G-1 checkpoint allowing cell division to occur. Healing begins.

continued
• Once skin healed, platelets no longer secrete PDGF. Thus skin cell are no longer signaled to express the proteins that move the cell cycle past the G-1 checkpoint. Cell division stops as the cell cycle can no longer move past the G-1 checkpoint.

Types of Genes Associated with Cancer
• Cancer can be caused by mutations to genes that regulate cell growth and division
• Tumor viruses can cause cancer in animals including humans
• A study of tumor virus have led to a better understanding of the molecular basis of cancer.

Oncogenes and Proto-Oncogenes
• Oncogenes are cancer-causing genes
• Proto-oncogenes are the corresponding normal cellular genes that are responsible for normal cell growth and division
• Conversion of a proto-oncogene to an oncogene can lead to abnormal stimulation of the cell cycle

• Proto-oncogenes can be converted to oncogenes by– Movement of DNA within the genome: if it ends
up near an active promoter, transcription may increase
– Amplification of a proto-oncogene: increases the number of copies of the gene
– Point mutations in the proto-oncogene or its control elements: causes an increase in gene expression
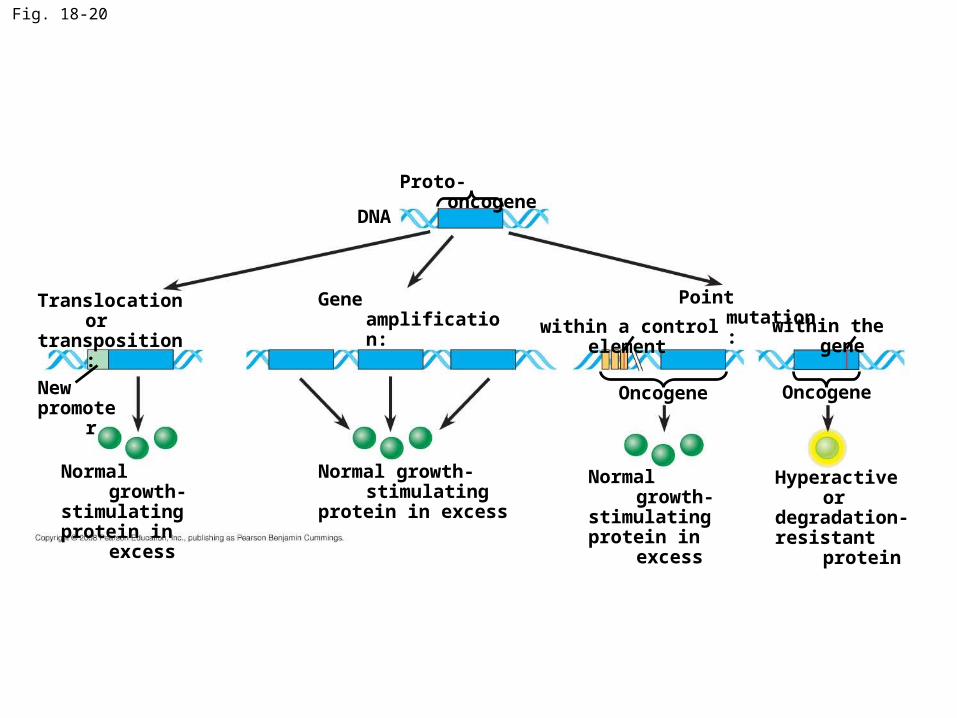
Fig. 18-20
Normal growth-stimulatingprotein in excess
Newpromoter
DNA
Proto-oncogene
Gene amplification:Translocation ortransposition:
Normal growth-stimulatingprotein in excess
Normal growth-stimulatingprotein in excess
Hyperactive ordegradation-resistant protein
Point mutation:
Oncogene Oncogene
within a control element within the gene

Cancer caused by conversion of protooncogenes to oncogenes can cause two problematic molecular events
• Interference with normal signaling pathways that control cell division (stuck gas pedal-allows cell cycle to move past checkpoints when it shouldn’t ).
• Inactivation of tumor suppressor genes (defective brakes-you can’t stop cell division in damaged/mutated cells)
• Mutations in the ras proto-oncogene and p53 tumor-suppressor gene are common in human cancers

Tumor-Suppressor Genes
• Tumor-suppressor genes help prevent uncontrolled cell growth. P53-”guardian angel of the genome”
• Mutations that decrease protein products of tumor-suppressor genes may contribute to cancer onset
• Tumor-suppressor proteins (usually activated by damaged DNA)
– Repair damaged DNA
– Inhibit the cell cycle so the cell doesn’t divide until the damaged DNA is repaired
– Activates “suicide genes” when DNA damage is irreparable and causes programmed cell death (apoptosis)

• Suppression of the cell cycle can be important in the case of damage to a cell’s DNA; p53 prevents a cell from passing on mutations due to DNA damage
• Mutations in the p53 gene prevent suppression of the cell cycle (knocks out the brakes that stop down cell division)

Fig. 18-21b
MUTATIONProtein kinases
DNA
DNA damagein genome
Defective ormissingtranscriptionfactor, suchas p53, cannotactivatetranscription
Protein thatinhibitsthe cell cycle
Activeformof p53
UVlight
(b) Cell cycle–inhibiting pathway
2
3
1

Interference with Normal Cell-Signaling Pathways
• Mutations in the ras gene can lead to production of a hyperactive Ras protein and increased cell division
• It’s like a stuck gas pedal in your car. Cell Division keeps going at a faster rate.

Fig. 18-21a
Receptor
Growthfactor
G protein GTP
Ras
GTP
Ras
Protein kinases(phosphorylationcascade)
Transcriptionfactor (activator)
DNA
HyperactiveRas protein(product ofoncogene)issuessignalson its own
MUTATION
NUCLEUS
Gene expression
Protein thatstimulatesthe cell cycle
(a) Cell cycle–stimulating pathway
11
3
4
5
2

Fig. 18-21
Receptor
Growthfactor
G protein
GTP
Ras
GTP
Ras
Protein kinases(phosphorylationcascade)
Transcriptionfactor (activator)
DNA
HyperactiveRas protein(product ofoncogene)issuessignalson its own
MUTATION
NUCLEUS
Gene expression
Protein thatstimulatesthe cell cycle
(a) Cell cycle–stimulating pathway
MUTATIONProtein kinases
DNA
DNA damagein genome
Defective ormissingtranscriptionfactor, suchas p53, cannotactivatetranscription
Protein thatinhibitsthe cell cycle
Activeformof p53
UVlight
(b) Cell cycle–inhibiting pathway
(c) Effects of mutations
EFFECTS OF MUTATIONS
Cell cycle notinhibited
Protein absent
Increased celldivision
Proteinoverexpressed
Cell cycleoverstimulated
1
2
3
4
5
2
1
3

The Multistep Model of Cancer Development
• Multiple mutations are generally needed for full-fledged cancer; thus the incidence increases with age
• At the DNA level, a cancerous cell is usually characterized by at least one active oncogene and the mutation of several tumor-suppressor genes

Fig. 18-22
EFFECTS OF MUTATIONS
Malignant tumor(carcinoma)
Colon
Colon wall
Loss of tumor-suppressor geneAPC (or other)
Activation ofras oncogene
Loss oftumor-suppressorgene DCC
Loss oftumor-suppressorgene p53
Additionalmutations
Larger benigngrowth (adenoma)
Small benigngrowth (polyp)
Normal colonepithelial cells
5
42
3
1

Inherited Predisposition and Other Factors Contributing to Cancer
• Individuals can inherit oncogenes or mutant alleles of tumor-suppressor genes (some cancers run in families)
• Inherited mutations in the tumor-suppressor gene adenomatous polyposis coli (APC)are common in individuals with colorectal cancer
• Mutations in the BRCA1 or BRCA2 gene are found in at least half of inherited breast cancers
• A woman who inherits one mutant BRCA1 allele has a 60% chance of breast cancer vesus a 2% chance if you don’t have the mutant allele