regulation of cellular nitrogen metabolism carbon and...
TRANSCRIPT

Regulation of Cellular Nitrogen Metabolism Carbon and Nitrogen Metabolism in Model Marine Diatoms
Andrew E. Allen (JCVI, Scripps Oceanography, San Diego)

Upwelling Boundary Currents: ~25-50% of Total Marine ProductivityDiatom dominated
J. Schmaltz, NASA (from Falkowski and Oliver (2007), Nature Reviews, Micro.)
Images, A. Allen (Puget Sound Phytoplankton Metatranscriptomics Project) (collaboration with Ginger Armbrust, Micaela Parker, Ahmed Moustafa)
Diatoms are thought to be the source of many modern day oil deposits
glycolate 2-P
glycolate
1,3 bisphosphate
glycerate
glyceraldehy
de
3-Pdihydroacetone
phosphateseduloheptulose
7-P
ribose 5-P
ribulose 5-P
ribulose
1,5 bisphosphate
Xylulose 5-P
Plastid
HCO3 + glutamine
carbamoyl-P
carbamolyl aspartate
dihydrorotate
orotate
UMP
UDP
UTP
CTP
UMP
uridine
uridineuracil
Triose/hexose phosphate/
Phosphate
Glucose 6-P/
Phosphate
acyl carnitinecarnitine
acyl
carnitinecarnitine
L-3-hydroxyacyl CoA
Mitochondrion
glycineserine
glyoxylate
hydroxypyruvate
glycerate
cysteine
alanineS
thiamine
2-ketobutyrate
2-
acetohydroxy
butyrate2,3 hydroxy
3-
methylvalerate2-keto 3-
methyl
valerate
fructose
1, 6 bisphosphatefructose
6-P
erythrose 4-P
hypoxanthine guanine
inosine quanosine
IMP
GMP
adenine
adenosine
AMP
PUFAfatty acid
-acyl CoA
trans enoyl CoA
beta hydroxyacyl CoA
fatty acid acyl CoA
acetyl CoAbeta ketoacyl CoA
OAA
citrate
succinate
glyoxylateacetyl CoA
H2O2
Peroxisome
pyruvateOAA
malate
PEP
2-phosphoglycerate
3-phosphoglycerate
1,3 DPG
glyceraldehyde 3-P
fructose 1,6 diphosphate
DHAPglycerol 3-P
glycerol
fructose 6-P
glucosamine 6-P
glucose 6-P
ribose 5-P + xylulose 5-P
ribulose 5-P
6-phosphogluconate
triose
phosphates
NO3Silicate
lactate
cysteine
cystathione
L-homocysteine
methionine
IMP
XMPInome
xanthosine
AMP
xanthine
uric acid
hypoxathine
urea
urea CO2 + NH4
S-adenosyl-L-
methionine
putrescine
spermidine
S-adenosyl
methioninamine
spermidine
S-adenosyl
methioninamine
glucose
glutamate
acetate
N-acetylglucosamine
1-P
N-acetylglucosamine
6-P
UDP-N-acetylglucosamine
glutamine
NH4
4-α-methylcholestra-
8-en-3-β-ol
anthranilate
erythrose 4-P
DAHP
3-dehydro
shikimate
shikimate
shikimate 3-P
EPSP
chorismate
prephenate
phenyl pyruvate
arogenate
malonyl CoA
acetyl CoA
malonyl ACP
3-
ketobutyryl
ACPbutyryl ACP
3-ketoacyl
ACP
continued cycles
16:0 ACP
18:0 ACP
O-acetyl serine
sulfate
APS
PAPS
sulfite
H2S
Serine
NO2
NO2 NH3
glutamate
glutamate 1-semialdehyde
glutamyl tRNA
5-aminolevulinate
porphobillinogen
preuroporphyrinogen
uroporphyrinogen III
coproporphyrinogen III
protoporphyrinogen IX
protoporphyrin IX
Mg protophorphyrin IX
Mg protoporphyrin
monomethyl ester
monovinyl
protochlorophyllide
chlorophyllide a
pyruvate
2-
acetolactate2,3
dihydroxy
isovalerate2-
ketoisovalerate
3-
isopropylmalate
glutathione ox glutathione red pyruvate
+
glyceraldehyde 3-P
deoxyxylulose 5-P
MEP
4-cytodine-5-diphospho
2-C-methyl-D-erythritol
IPP DMAPP
GGPP
phytoene
lycopene
beta carotene
zeaxanthin
antheraxanthine
violaxanthin
diatoxanthin
diadaxanthin
Acyl CoA
trans enoyl CoA
3-keto acyl CoA
acyl CoA
acetyl COA
acetoacetyl COA
HMG COA
mevalonate
MVPP
IPP
DMAPP
farnesyl
diphosphateprotein
prenylation
presqualene
diphosphate
(S)-squalene-
2,3-epoxide
lanosterol
12-demethyl-
lanosterol
zymosterol
cholesta-7,24-dien-3-β-ol
cycloartenollathosterol
7-dehydro-cholesterol
cholesterol
Cholesterol
-5α6β-epoxide
ergosterol phytosterol
PRPP
PRA
GAR
FGAR
FGAM
AIR
CAIR
SAICAR
AICAR
CAICAR
FAICAR
IMP
SAMP XMP
GMPAMP
Cytosol
3-carboxy-3-
hydroxy-
isocaproate
2-oxo-
isocaproate
pyruvate
OAAcitrate
isocitrate
succinyl CoA
succinate
fumarate
malate
acetyl CoACO2
TCA cycle
apha keto
glutarate
H
H
H
H
H
H
Calvin cycle
Glycolat
e
cyclemalate
H
H
R
Polyamines
Fatty acid
oxidation
Phosphat
e
sugars
Photo
respiratio
n
H
Chitin
NO3
H
H
Triglycerides
H
Creatine-P
H
H
H
Purine
biosynthesi
s
Pyrimidine
biosynthesi
s
Fatty acid
oxidation
H
Isoprenoid
biosynthesi
s
Glycolysis &
Gluconeogene
sis
H
3-phosphoglycerate
Chlorophyll a
Heme
Isoprenoid
biosynthesis
Fatty acid biosynthesis
H
Tyrosine
Phenylalanine Cysteine
Isoleucine
Valine
Leucine
Fucoxanthin
glutamate glutamine
H
H
H
Purine SalvagePyrimidine Salvage
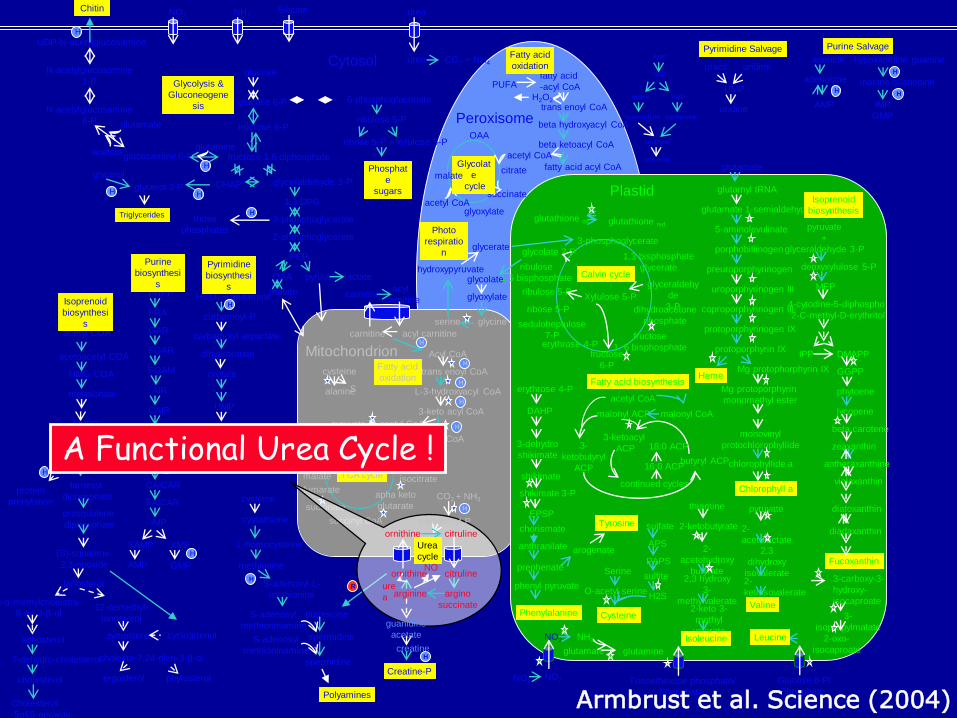
A Functional Urea Cycle !
creatine
guanidino
acetate
citruline
carbamoyl P
CO2 + NH3
NO
ornithine
Urea
cycle
ornithine citruline
ure
a arginine argino
succinate
Armbrust et al. Science (2004)
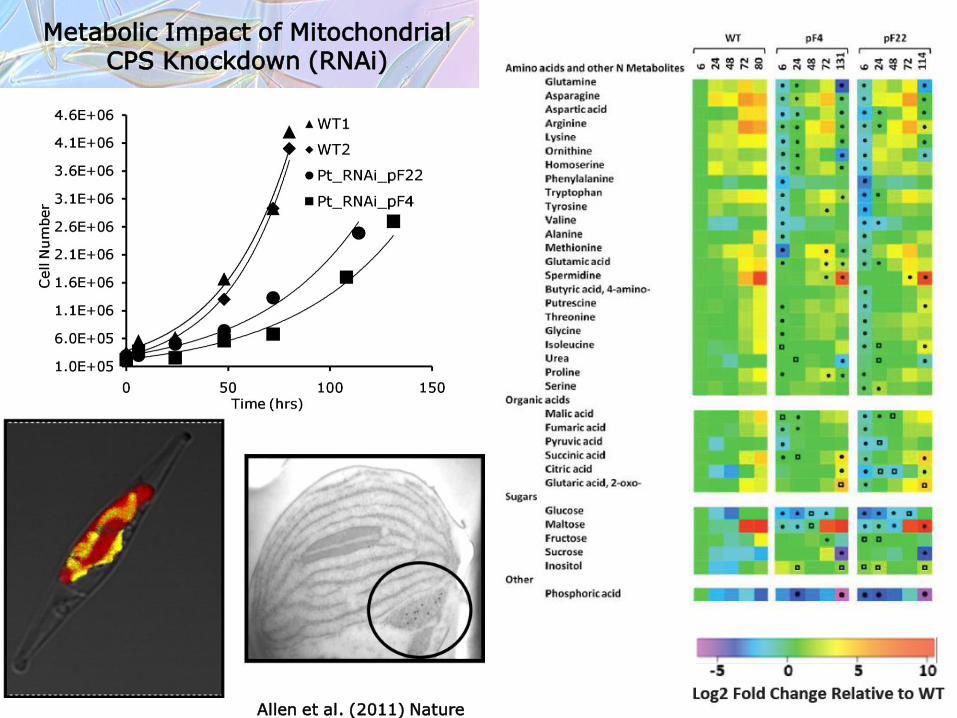
Metabolic Impact of Mitochondrial CPS Knockdown (RNAi)
Allen et al. (2011) Nature

% 15N Enrichment During Growth on Nitrate or Urea in Wild Type Cells
Dupont et al. in prep
Synthesis pathways in chloroplast

Metabolic Consequences of TALEN-Based Urea Cycle Manipulation in Diatoms
Graham Peers, Colorado State University
Weyman et al. Plant Biotechnology Journal (In Press)

Nitrate Reductase Homologous Recombination Vectors for Gene Inactivation
NR Talen vector recognizes NR target sequence; endonuclease fused to recognition domain make double-stranded cut at target site.
NR homologous recombination vector provides homologous NR sequences for Pt’s DNA repair mechanism. The insertion of the Sh blesequence to the NR gene during recombination inactivates NR gene.

P. tricornutum NR Knock Out Results after Biolistic Transformation
WT cultures were grown simultaneously as a
control. Plates were photographed 10 day
after inoculation: 5 ul NR-KO cells into 2 ml
F/2 media; auto-fluorescence was recorded
daily as a proxy for growth for two weeks.
Experiments were successfully repeated over several months.
NR-KO #9 NR-KO #14NH4 vs NO3 growth assay
NH4 NO3 NH4 NO3
blank #9 WT blank #9 WT blank #14 WT blank #14 WT
1 2 3 4 5 6 7 8
1. Ladder
2. WT-1 NH4 day zero
3. WT-2 NH4 day zero
4. KO-9 NH4 day zero
5. KO-14 NH4 day zero
6. WT-1 NO3 day 3
7. KO-9 NO3 day 3
8. KO-14 NO3 day 3NR
Western blot of WT vs KO-9, KO14

Confocal Images of NR KO-1 Cells Grown on NH4 vs NO3
Amended F/2 Media
[880 uM] ammonium [880 uM] nitrate
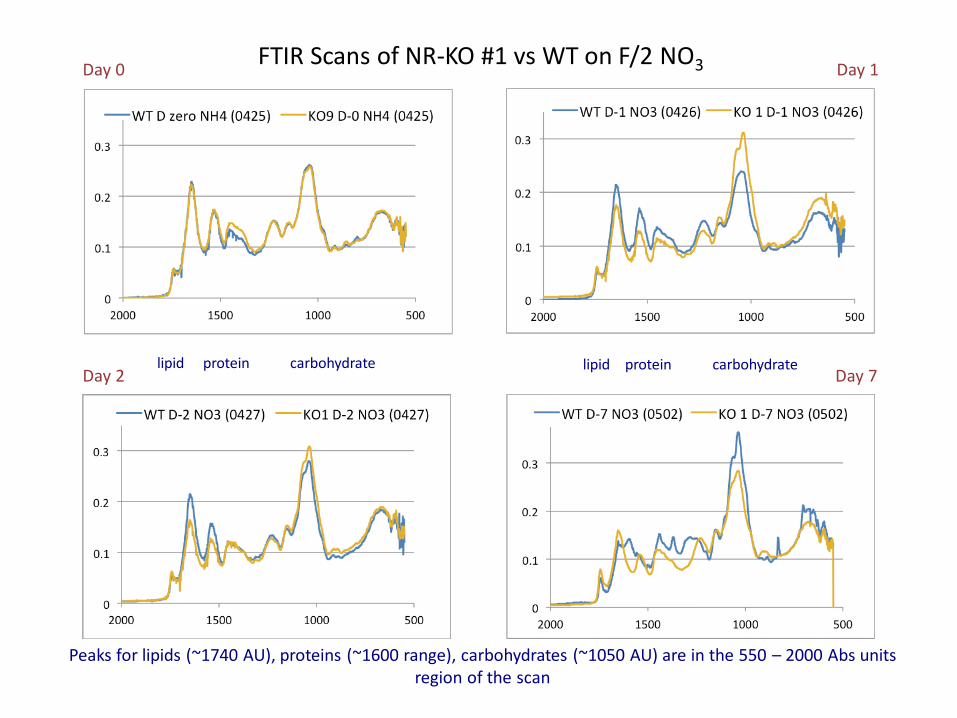
Day 0 Day 1
Day 2 Day 7
FTIR Scans of NR-KO #1 vs WT on F/2 NO3
lipid protein carbohydrate lipid protein carbohydrate
Peaks for lipids (~1740 AU), proteins (~1600 range), carbohydrates (~1050 AU) are in the 550 – 2000 Abs units region of the scan

Scaled-Up View of Lipid Peaks –
FTIR scans of NR-KO #1 & #9 vs WT on F/2 (NO3)

Bacterial Conjugation
P. tricornutum
E. coli
Conjugative Plasmid
Bacteria
Yeast Mammalian Cell
?
F pilus
Conjugative Plasmid
Conjugative Plasmid
Conjugative Plasmid
Conjugative Plasmid
Karas et al. in prep
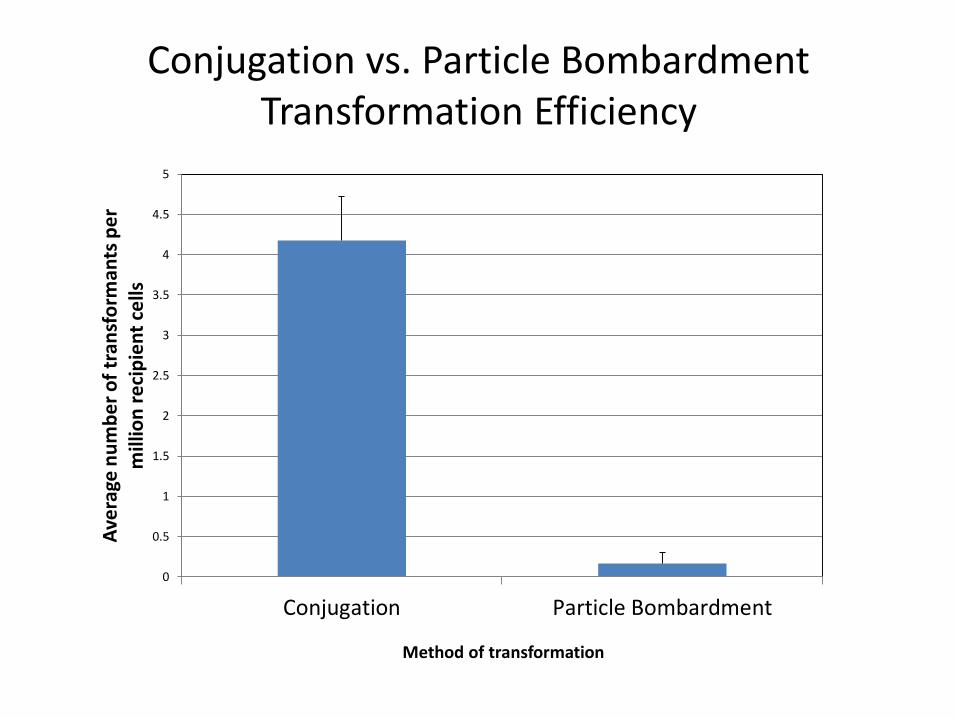
Conjugation vs. Particle Bombardment Transformation Efficiency
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
Conjugation Particle Bombardment
Ave
rage
nu
mb
er
of
tran
sfo
rman
tsp
er
mill
ion
re
cip
ien
t ce
lls
Method of transformation

Rescued cells growing in F/2 media show that
both the N’YFP and NR proteins are active
A conjugation plasmid, containing the above N’YFP-NR cassette, driven by the
NR promoter, was transformed into a conjugating strain of E. coli. Pt WT and KO
lines were incubated with the E.coli and plated on selective media.
NH
4
NO3
Potential KO-complemented conjugates
formed colonies large enough to transfer
to liquid media after 10 days. C1 -6 were
transferred to 24 well plates with NH4 and
NO3 amended F/2 media. Strains growing
in both N-media (C2-5) indicate that the
the nitrate reductase gene on the plasmid
is being expressed.
NR knockout complementation by conjugation of P. tricornutum
with E. coli

251 gene cluster of predominantly chloroplast localized genes
Short Time Sacle Addition or Removal of Nitrogen is Major Transcriptional Trigger
Cluster of 428 genes, in addition to fatty acid functional genes, includes transcription factors, vacuolar, peroxisomal, ubiquitin-conjugating enzymes, kinases, and N-response regulators
p-value 1.0E-07p-value 5.0E-04
N-depleted time points15 min after N-addition
McCarthy et al. in prep

Regulation of Nitrogen Metabolism in the
Marine Diatom Phaeodactylum tricornutum
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
N-freewash
NO3spike Wash*
T15 T45 T-end T15 T45 T-end T15 T45 T-end T15 T45 T-end
RP
KM
s (
no
rma
lize
d m
RN
A r
ea
ds)
nitrate reductase
nitrite transporter
glutamine synthetase
N-
free
NH4+ NO3
- NO2-
Experimental design & key enzymes
McCarthy et al. in prep
Nitrate Reductase R and 34 genes that most closely co-vary in
expression; p=6.0E-04.
Transcripts (annotated) include: nitrate & nitrite transporter, nitrite reductase(s), glutamine synthetase, HIS acid phosphastase, protein kinases, voltage-gated chloride channel, vaculor amino acid transporter, silicon transporter, NO dioxygenase (globin domain protein), mitochondrial carrier protein, among others

Putative Nitrate Sensing Gene ptNIT(NO3 sensitive kinase, gene model manually extended in 3’ direction to find novel
Eukaryotic NIT domain)
NasR and NarX(Boudes et al., 2012; Chueng and Hendrickson, 2008)
Nitrate/Nitrite Sensing Kinase

In K-means cluster 1, NR, Pt54983, clusters with phosphorylated
peptides involved in nitrate assimilation:
CPS I, POT and voltage-gated Cl
channels, kinases, phosphatases,
transcription factors, etc.
Note: NR’s proteomic profile added to phosphorylated-peptide data for tracking purposes
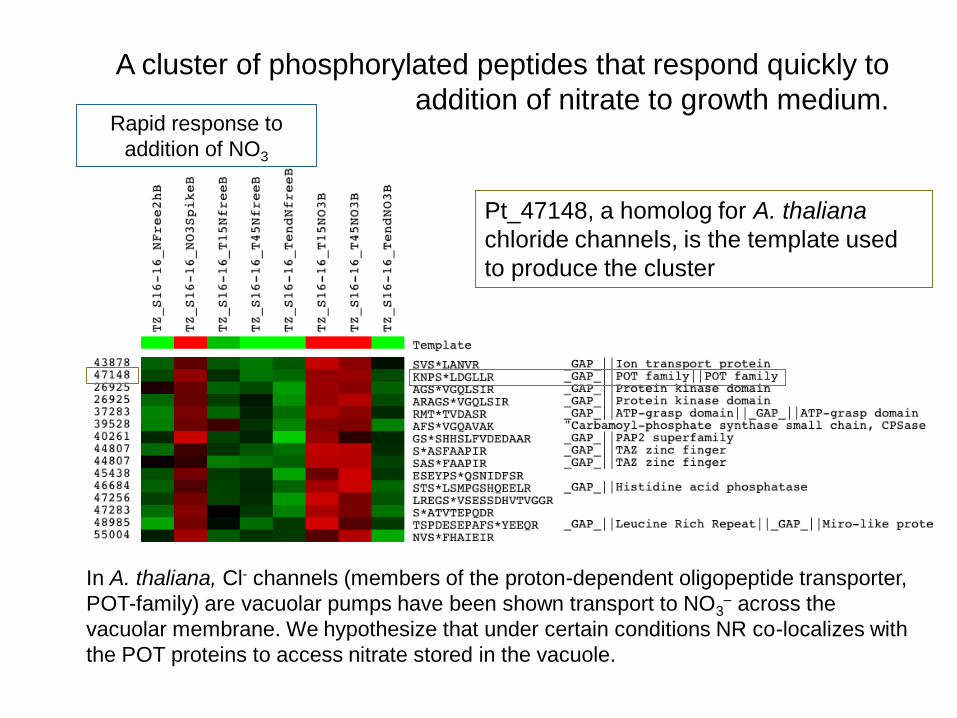
In A. thaliana, Cl- channels (members of the proton-dependent oligopeptide transporter,
POT-family) are vacuolar pumps have been shown transport to NO3– across the
vacuolar membrane. We hypothesize that under certain conditions NR co-localizes with
the POT proteins to access nitrate stored in the vacuole.
Rapid response to
addition of NO3
A cluster of phosphorylated peptides that respond quickly to
addition of nitrate to growth medium.
Pt_47148, a homolog for A. thaliana
chloride channels, is the template used
to produce the cluster

N-responsive, phosphorylated transcription factors clustered by abundance per time point
Cohorts of transcription factor Pt44807 clustered by phosphoryl abundance
under replete NO3 conditions
Cluster includes a 2nd TF:
Pt47256, an ammonium
transporter and the putative
NO3, POT transporter,
Pt47148
NO3 T
end
NO3
T15
NO3
T45
N
free
NO3
spike
N-
free
T15
N-free
T45
N-free
T end
LREGS*VSESSDHVTVGGR
S*ASFAAPIR
SAS*FAAPIR
TTTPATVDPQGHQFS*PTTR
TTTPATVDPQGHQFS*PTTR
SSGVGGAAS*AAPEAIIMGRPK
AGS*VGQLSIR
ARAGS*VGQLSIR
DHVDELMEIRAS*K
RMT*TVDASR
SVS*LANVR
ESEYPS*QSNIDFSR
STS*LSMPGSHQEELR
KNPS*LDGLLR
S*ATVTEPQDR
RPS*EDDVASVLQK
TSPDESEPAFS*YEEQR
47256
44807
44807
292
292
1686
26925
26925
27877
37283
43878
45438
46684
47148
47283
48350
48985
Myb1R
_GAP_||TAZ zinc finger
_GAP_||TAZ zinc finger
Myosin head (motor domain)
Bromodomain
_GAP_||Protein kinase domain
_GAP_||Protein kinase domain
_GAP_||Ammonium Transporter Fam
_GAP_||ATP-grasp domain||_GAP_
_GAP_||Ion transport protein
_GAP_||Histidine acid phosphat
_GAP_||POT family||POT family
_GAP_||Adenylate cyclase
_GAP_||Miro-like protein||_GAP_|
NO3 T
end
NO3
T45
NO3
T15
N
free
NO3
spike
N-
free
T15
N-free
T45
N-free
T end

Changes in phosphorylated protein abundance in response to
N-sources: NH4 and NO2NH4 NO2
S1
7: clu
ste
r 1
S1
7: clu
ste
r 1
4
GREEN = Low Phosphorylation
RED = High Phosphorylation
Phosphorylated
in response to
lack of nitrogen
Phosphorylated in
response to
nitrogen
The appearance of
the ammonium
transporter in these
4 clusters suggests
it plays a complex
role in N-source
transport

Peptide profiles of the ammonium transporter Pt_27877
N free NO3

Diel clustering of functional categories within diel expression clusters: Consistent periodic modulation (P < 0.05, over all replicates)
A. Transcription Factors
4 σ-TF’s (total of 8) induced early morning Discrete peak at 1h light
B. Light Harvesting Complex proteins
Induced early morning Discrete peak at 1h light
Repressed by light (at 1h L), back upregulated in the afternoon
Repressed at dawninduced at dusk or at midnight

Kleeson et al, in prep
Testing compartment and non compartment models
Compartmentalized Metabolic Flux Reconstructions
Sabrina KleesonMax Plank, Potsdam
Alisdair Fernie and Zoran Nikoloski, Max Plank, PotsdamBernhard Palsson, UCSD

Ongoing and Future Experiments & Conclusions
In Vitro Transcription Factor Expression and DNA Porbing Targets for all TFs (DNA Aptamer Protein sequencing (DAP-seq)
Generate Biomass Objective Functions Required for Genome Scale Reconstruction (biochemical and energetic requirements for biomass)
Metabolic and Transcriptional Regulatory Network Reconstruction

Conclusions
Novel Eukaryotic recombination and repartitioning of bacterial metabolism; for example diatom mitochondria biochemistry
The diatom urea cycle represents a critical for anaplerotic carbon (CO2) fixation pathway into key nitrogenous compounds
Cellular compartmentalization is critical for cellular ecology and modeling of integrated carbon and nitrogen metabolism (we lack critical information related to intracellular transporter reactions)
Transcriptome rapidly responds to silicon and nitrogen starvation conditions in a way that “sets the stage” for lipid accumulation. Regulation of intracellular carbon flux appears to be under some degree of transcriptional control

Jonathan Badger (JCVI)John McCrow (JCVI)Flip McCarthy (JCVI)Jeroen Gillard (JCVI)Stephane Lefebvre (JCVI)Chris Dupont (JCVI)Philip Weyman (JCVI)Sabrina Kleeson (MPI, Golm)Bernhard Palsson (UCSD)Jennifer Levering (UCSD)Alessandra Gallina (UCSD)Karsten Zengler (UCSD)Graham Peers (Col. State University)Adam Kustka (Rutgers University)Chris Bowler (ENS)Uma Maheswari (EBI)
Marine Microbiology Initiative
Environmental Genomics, Biological Oceanography, Dimensions of Diversity