regional differences in cat esophageal circular … · regional dinerences in cat esophageal...
TRANSCRIPT

REGIONAL DIFFERENCES IN CAT ESOPHAGEAL CIRCULAR SMOOTH MUSCLE: A WHITE NOISE ANALYSIS
Robert Yu
A thesis submitted in conformity with the requirements for the degree of Master of Science Graduate Department of Physiology
Institute of Biomaterials and Biomedical Engineering University of Toronto
O Copyright by Robert Yu 1999

National Library Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques 395 Wellington Street 395. rire Wellington OttawaON K l A W W O N K l A W Canada Canada
The author has graated a non- exclusive licence allowing the National Library of Canada to reproduce, loan, distribute or seil copies of this thesis in microfom, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.

Regional Dinerences in Cat Esophageal Circular Smooth Muscle: A White Noise Analysis
Robert Yu Masîer of Science
Department of Physiology and Institute of Biomateriais and Biomedical Engineering
University of Toronto 1999
By using a Gaussian white noise identijïcation technique, regional differences are
demonstrated by dxerences in electrical response characteristics.
Gaussian white noise rneasurements and analysis techniques were used to obtain
input impedance functions of myocytes fiom feline esophageal body cells (EBC) isolated
from 2 and 4 cm above the lower esophageal sphincter. A whole ce11 patch-clamp
configuration was used to inject white noise current and record a voltage response. The
magnitude of the input iïnpedance of the ce11 was calculated fiom the first order kemel.
Fourier transforms of the kemel indicate a difference in input resistance. The time
constants obtained indicated that EBC4 was quicker to respond since the higher
fkequency values correspond to a fster response characteristic of the cell.
There are regional ciifferences in the passive cable properties of smooth muscle
cells fkom the smooth muscle esophagus which likely contribute to the contraction profile
and sequential nature of peristalsis.

1 would like to offer Drs, Nicholas Diamant and Berj Bardakjian my most sincere
gratitude for their guidance and wisdom. Though 1 have only worked with them for two years,
they have given me a lifetime's worüi of practical insight.
1 would also like to tfiank Dr. Salapatek and Junzhi Ji for their patience, smooth muscles
are not always cooperative. AIong with the rest of my labmates, Shuwen Xue, Dr. H. Lau, Diana
Valdez, Ahmad Muinuddin and Jervana Valdez, thank you ail for your intelligence and
friendship, it will always be cherished.
To my family, mom, dad and Sue, and my fnends, especially Elaine, without whose
encouragement and moral support, none of this would have been possible, 1 extend my deepest
appreciation and Iove.
iii

-TABLE OF CONTENTS-
Abstract Acknowledgements Table of Contents List of Tables & Figures List of Abbreviations
Chanter One 1.1 Esop hageai Anatom y and Physiology
Physiology of the Esophagus Upper Esophageal Sphincter Esophageal Body Lower Esophageal Sphincter Histology
Esophageal Motor Physiology Primary Peristalsis Secondary Peristalsis
1.2 Overview of the Control of Esophageal Motor Function Innervation of the Esophagus
Swallowing Centre i) Afferent Input ii) Coordinating Region iii) Efferent Output
Extrinsic Innervation i) Motor Innervation
Intrinsic Innervation i)Enteric Nervous System
Control of Esophageal Perïstalsis Intrinsic Myogenic Control
1.3 Regional Differences
1.4 Modeling of Physiological Systems Interpretation of Kernels
1.5 Experimental Objective Rationale and Aim of Study
Chanter Two 2. I Materials
Animal Models P harmacological Agents
iv vi vii

2.2 Methods Specific Protocols
Ceil Isolation Biophysical Measurements White-Noise Current Injection and Cell Viability Test
2.3 Data Acquisition and Analysis Computations
I st Order Kemel Fourier Transform
Statistical Methods
Chapter Three 3.1 Equipment Validation
Model Ce11 Electrode
3.2 White Noise Injection First Order Kemels Fourier Transform
Input Resistance i) C tassical Technique
Cut-Off Frequency SIope of Fourier Transform (Roll-On)
3 -3 Statistical analysis Between EBC-2 and EBC-4
Chabter Four 4.1 Discussion
Equi pment Validation Between EBC-2 and EBC-4
Input Resistance Time Constants Order of System
4.2 Future Investigations
References

Cha~te r One
Figure 1.1 The human esophagus 1 -2 Diagrammatic fepresentation of misculmis externa of the esophagus
Figure 2.1 Schematic showing procedure for obtainïng whole ce11 patch that can be studied with patch-clamp technique
2.2 Electrophysiological experiment with Gaussian white noise (GWN) input current
2.3 Total outward current voltage-dependant activation along the esophageal body
Figure 3.1 The cut-off fiequency of mode1 ce11 3.2 The time constant of the electrode 3.3 First order kemel fiom two regions 3 -4 Input resistance derived fiom input impedance plot 3 -5 Cut-off frequency and time constant of the cells 3.6 Derivation of the order of system fiom an input irnpedance plot
Table 3.1 Input resistances
Table 3.2 Surnmary of results

acetylcholine - ACh
central nervous system - CNS circuiar muscle - CM centimeter - cm current - 1 curent-voltage - IV
enteric nervous system - ENS esophageal body cell - EBC esophageal body ce11 two centimeters fiom LES - EBC-2 esophageal body ce11 four centimeters h m LES- EBC-4
fast orthogonal algorithm - FOA gastrohtestinal tract - GI tract Gaussian white noise - GWN
hertz - Hz
interstitial cells of Cajal - [CC input impedance - Z h
longitudinal muscle - LM lower esophageal sphincter - LES
megaohrn - MST millilitre - ml millimoIar - mM millisecond - ms millivolt - mV microsecond - ps molar- M
neurotransmitter - NT nitric oxide - NO
ohm -R
plus or minus - I potassium ion - K' seconds - s standard error o f the mean - SEM
upper esophageal sphincter - LES
volt - V
vii

1.1 Esophageal Anatomy and Physiology
Ph~siologv of the E s o ~ h n u s
The esophagus is a hollow muscdar tube that connects the pharynx to the
stomach. It is guarded at the proximal end by the upper esophageal sphincter W S ) and
at the distal end by the lower esophageal sphincter (LES) (Fig. 1.1). The prirnary role of
the esophagus is to propel food or fluid bolus aborally from the pharynx to the stomach.
The driving force in the aborad direction is deglutition or the coordinated contraction
induced by swallowing aided by gravity. The UES acts to prevent air entry into the
esophagus during respiration and to prevent retrograde movement of material fiom the
esophagus into the hypopharynx. The LES acts to prevent reflux of gastric secretions
into the esophagus. The interactions between the centrai nervous system (CNS), enteric
nervous system @NS) and muscular components of the esophagus that underlie the
motor reflexes of this organ are cornplex and incompletely understood.
The purpose of this thesis is to use a new technique, white noise andysis, for
measurement of passive membrane properties in order to assess if regional ciifferences
exist in the esophageal body smooth muscle.

Figure 1.1 The human esophagus. Guarded proximally by the upper esophageal sphincter ( E S ) and distaltly by the lower esophageal sphincter (LES), it serves as a hollow muscula. tube which connects the pharynx to the stomach.

Studies were performed in isolated smooth muscle cells dissociated Erom circular
muscle layers dong the esophagus. The long-term goal of our laboratory is to define
esophageal îùnction through study of its neural and muscle control mechanisms.
Therefore, a brief description of the anatomy and control mechanisms of
esophageal motor fünction is presented in this section. From a fiuictional perspective, the
esophagus can be divided into three sections, the UES, esophageai body and the LES.
Upper Esophageal Sphincter
The esophagus begins at the UES, a purely striated muscle structure, which
consists predominantly of the cricopharyngeus and caudal fibres of the inferior
pharyngeal constrictor muscle. Circular muscle (CM) fibres fiom the proximal
esophagus also contribute to the sphincter (Netter 197 1, Asoh and Goyal 1978, Welch et
al. 1979). The UES is functionally defmed as a zone of high intraluminal pressure that
separates the pharynx fiom the upper esophagus. In humans, the axial length of the UES,
as measured by intraluminal manometry, is between 2 and 4 cm (Goyai et al. 1970, Ellis
1971).
Esophageal Body
The esophagus is collapsed in its resting state and is capable of distending to
accommodate fiuid and solid material. The esophageai body begins at the inferior margin
of the cricopharyngeus muscle and extends to the LES. In the human adult, the average
length between the UES and the LES is between 23-25 cm (Csendes et al. 1993, Denardi
and Riddell199 1, Li and Rand 1989) and 18-19 cm in the cat (Reynolds et al. 1984 and

1985). Measuring fiom the incisors in the human, it is approximately 40 cm to the
gastroesophageal junction Oenardi and Riddell 199 1).
In the proximal onequarter to one-third of the esophagus, the muscle is striated in
both the inner cucular muscle (CM) layer and the outer longitudinal muscle (LM) layes
of the muscularispropria (Fig. 1.2). Striated muscle begins to be replaced by smooth
muscle about 4 cm below the UES. This marks the beginning of the transition zone
(Meyer et al. t 986). The transition zone is of variable length; striated muscle and smooth
muscle fibres continue to be intermingled up to 10-13 cm below the cricopharyngeous.
The distal one-half to one-third of the esophageal body is composed entirely of smooth
muscle in both layers (Meyer et al. 1986). Occasionally, single bundles of striated
muscle extend within the smooth muscle al1 the way to the caudal end of the esophageal
body (Arey & Tremaine 1933). The major esophageal motor disorders including that
associated with gastroesophageal reflux disease and the spastic motility disorders occur in
the smooth muscle portion.
Lower Esophageal Sphincter
The LES is located at the junction between the esophagus and the stomach
(gastroesophageal junction). It cm be identified with intraluminal manometry as a 2-4
cm zone of high pressure. The LES is composed of smooth muscle in most species,
including humans, dogs, cats and opossums. Anatomic evidence of a sphincter has k e n
difficult to obtain. Liebermann-Meffert and colleagues (1979) gave the most detailed
description of the muscular anatomy of the human sphincter. They found an asymmetric
thickening of the CM in the terminal esophagus, just above the angle of His (the junction
Vagus ncrvc Loncirudinai muscfe
Meissner's !siibmucusall
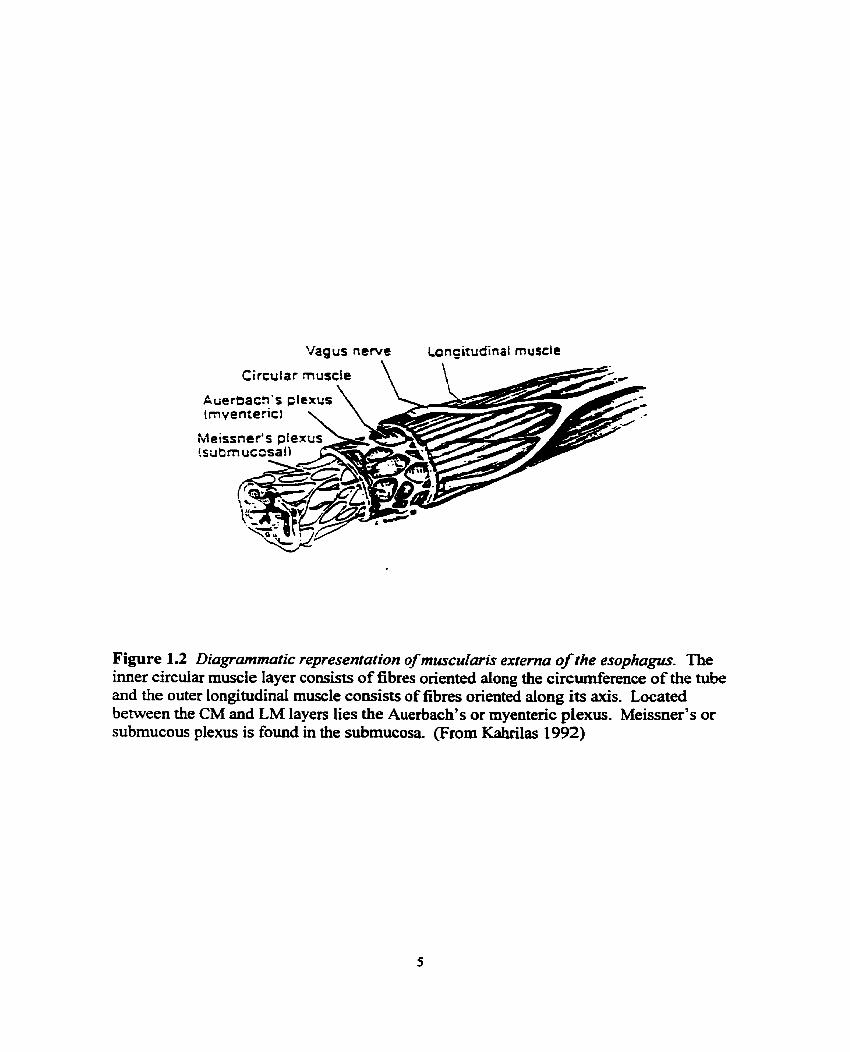
Figure 1.2 Diagrammatic representafion of muscularis externa of the esophagus. The imer circular muscle layer consists of fibres oriented dong the circderence of the tube and the outer longitudinal muscle coasists of fibres oriented dong its axis. Located between the CM and LM layers lies the Auerbach's or myentenc plexus. Meissner's or submucous plexus is found in the submucosa (From Kahrilas 1992)

of the tubular esophagus and the saccular stomach). In experimental animals, the
relationsbip behveen anatomic structure and fiuiction has been more precisely studied.
Similar thickening of circular, but not longitudinal, muscle has been observed in the
opossum and cat (Seelig and Goyal 1978, Biancani et al. 1982). In the opossum,
ultrastuctural studies have demonstrated severai differences between LES muscle cells
and those of the esophageai body. These differences may help to explain some of the
mechanisms responsible for the functional ciifferences between LES and esophageal body
contractility. Muscle cells of the LES are larger in diameter and form fewer gap junctions
(Daniel and Posey-Daniel 1984). However, neuronai ce11 bodies in the LES are smaller
than those in the esophageal body, perhaps due to the tonically contracted state of the
sphincter (Sengupta et al. 1987). Mitochondrial and smooth endoplasmic reticulum mass
is greater in the LES than in the esophageal body (Christensen and Roberts 1983).
Morphologically, the LES can also be distinguished fkom the esophageal body by the
presence of numerous intemuscular spaces containhg co~ec t ive tissue and blood
vessels (Seelig and Goyal 1978).
Hisrology
The esophageal wali, Iike other regions of the gastrointestinal (GI) tract, is
composed of three distinct layers: the mucosa, submucosa and muscularis propria (Fig.
1.2). However, what differentiates this organ nom the rest of the GI tract is the absence
of a serosal layer. The mucosal lïning is stratifi:ed squamous epithelium in al1 regions of
the esophagus except the LES, where both squamous and columnar epithelium may
coexist (Al Yassin and Toner 1977, DeNardi and Riddell 1991). The submucosa

comprises a dense network of co~ec t ive tissue within which are blood vessels,
lymphatic channels, esophageal glands and a nerve plexus called the submucous or
Meissner's plexus. The muscularis propria, the muscular wall proper, consists of muscle
which is divided into two layers: the inner circula muscle (CM) layer and the out
longitudinal muscle (LM) layer. The inner CM layer, with fibres oriented dong the
circumference of the tube, serves to constrict the esophageal lumen. The outer LM layer,
with fibres oriented dong its axis, serves to shorten the esophagus (Lerche 1950). The
combined action of these smooth muscle layers, opposed at right angles to one another, is
the basis of peristaltic contraction. Located between the CM and LM layers lies a nerve
plexus called the myenteric or Auerbach's plexus. Both the rnyenteric and submucosal
plexuses are continuos around the circumference of the esophageai wall (Sobotta and
Hammersen 1985).
Esophageal Motor Phvsiology
The major fiinction of the esophagus is to propel food and fluid into the stomach.
This is accomplished by way of sequential or peristaltic contractions of the esophageal
body in concert with an appropnately timed relaxation of the UES and LES. The
esophagus also clears any refluxed gastric contents back hto the stomach and takes part
in such reflex activities as vomiting and belching. Normal p e n d s i s is categorized into
either primary or secondary peristalsis, depending on its mode of initiation.
Primary Peristalsis
Primary peristalsis in the esophagus is a coordinated motor pattern initiated by the

act of swallowing. Continuous neuronal discharge of vagai lower motor neurons to the
UES creates a high-pressure zone through tonic contraction of the UES muscles (Asoh
and Goyal 1978, Yoshida et al. 198 1). During swallowing, this neuronal discharge
temporarily ceases allowing relaxation of the UES to occur (Van Overbeck et al. 1985).
M e r the bolus passes through the UES, the UES closes and r e m to its tonically
contracted state and a rapidly progressive CM contraction proceeds distally through the
esophagus starting in the upper esophagus and moving towards the LES. Each location
dong the esophageal axis contracts with a latency that increases gradually fkom UES to
the LES. In the upper third of the esophagus, contractions occur within 1-2 seconds afler
swallowing whereas in the lower third, this latency increases to between 5-8 seconds
(Biancani and Behar 1995).
There is a coordinated contraction of the LM and CM layers of the smooth muscle
esophagus during peristalsis, though each layer contributes to the process differently.
CM contraction produces the lumen-occluding propulsive wave front, whereas LM
contraction is associated with esophageal shortening. Sugarbaker and colleagues (1 984)
directly recorded local contractile and electrical activity of opossum CM and LM in vivo.
They found a sequential contraction of both layers in response to swallowing. With vagal
stimuiation, simultaneous LM shortening and sequential CM contractions were observed.
The LM contraction in each segment of the smooth muscle esophagus preceded and
outlasted the CM contraction, hence augmenting the amplitude of the propulsive
contraction (Sugarbaker et al. 1984). The LM contraction is associated with membrane
depolarization, whereas CM contraction is accompanied by a bimodai membrane
potential response; an initial hyperpolarization (inhibition) followed by depolarization.

The LES relaxes during swallowing and thus aiiows the aborally directed contraction
wave to pass the bolus fiom the esophagus to the stomach. AAer passage of the bolus
through the LES into the stomach, the LES closes with a prolonged contraction.
Motility studies with dry swallows reveal that contractions in the striated muscle
segment are shorter in duration (1-2 sec.) than those in the smooth muscle segment (4-7
sec). The velocity of the peristaltic wave is 3 c d s e c just above the LES (Humphries and
Castell 1977). Though the force of contractions can Vary fkom segment to segment and
even from swaliow to swallow, contractions in the distal one third of the esophagus are
also usually stronger (50- 150 mmHg) than those in the upper one-tkd (40-1 23 mmHg)
(Biancani et al. 1975). However, contractions in the proximal and distal esophagus are
stronger than those in the middle third (20-80 mmHg), perhaps because of the transition
between striated and smooth muscle esophagus in this region.
The physical characteristics of the ingested bolus also affect the speed and force
of esophageal phasic contractions. Increasing the size of the bolus tends to slow its
propagation velocity while increasing the force of contractions (Janssens et al. 1973,
Hollis 1975)- An increased viscosity dso increases the force of contractions and reduces
the speed of perktalsis (Dodds et al. 1973, Dooley et al. 1988). Bolus temperature can
also modulate esophageal peristalsis; warm boluses increase peristaltic velocity, and cold
boluses reduce it (Winship et al. 1970, Decarle et ai. 1977).
Secondury Peristalsis
Distention-induced or secondary peristalsis is the physiological mechanism
whereby reflwced gastric contents or food lefi behind d e r primary peristalsis are

propelled into the stomach. Distention-induced peristalsis is triggered by stimulation of
sensory receptors in the esophageal body.
Balloon distention (BD) is a commonly used method to induce secondary
peristalsis under experimental conditions. The esophageal response to intraluminal
distention consists of proximal excitation and distal inhibition, which serves to aid aboral
propulsion of the intraluminal bolus.
EarIy studies suggested that the esophageal response to swdowing resembled
that of bdoon distention (Craemer and Schlegel 1957, Fleshler et ai. 1959). However
more recent quantitative work by Paterson and colleagues (1 991a) has demonstrated that
contrary to earlier reports, secondary peristalsis induced by isometric distension can differ
significantly fiom primary peristalsis. This area continues to be one of considerable
debate as results fiom isobaric distension are studied.
1.2 Overview of the Control of Esophageal Motor Fuoction
Innervation of the E s o ~ h a w
The control mechanisms of esophageal peristalsis include those of the CNS, ENS,
and myogenic properties of the esophageai musculature. The esophageal innervation
provides the mechanism for the excitation and inhibition of the muscle at al1 these levels
and also provides vital sensory feedback via aflierent pathways which play important roles
in reflex modulation of the esophageal body and sphincter motor activities.

SwaZZowing Centre
CNS control of esophageal motor function resides in the swallowing centre - a
group of intercomected brainstem nuclei that receive cranial as weii as peripheral inputs.
The centre is composed of two intimately comected half centres and is located in the
medulla and pons (Jean 1984). Penstalsis cm either be triggered voluntarily, via higher
brain centres, or reflexly, by stimulation of peripheral afferents which go to the brain
stem swallowing centre (Clouse and Diamant 1988). Stimulation of the swallowing
centre activates vagal efferent nerves to the oropharyngeal and the esophageal muscle.
The vagal nerve mediates prirnary peristalsis in the striated muscle segment and
contributes to that in the smooth muscle segment of the esophagus. Conceptually, the
swallowing centre has t h e fimctional components: an afferent reception system, a
complex organizing system of neurons, and an efferent system of motor neurons
(Diamant 1989).
i) Afferent Input
Afferent information fiom the periphery enters into the solitary tract, the aEerent
reception portion of the swallowing centre. This sensory information serves to initiate
the swallowing sequence, as well as to alter previously initiated swallowing centre
activity and therefore modie ongoing motor activity. Sensory information fiom the
esophageal body and sphincters is carried in the vagus nerve, with the ce11 bodies located
in the nodose ganglion. This sensory pathway plays an important role in monitoring and
modulation of esophageal motor activity. Sensory idormation is also carried by the
sympathetic nerves to the spinal cord segments Cl to L3 (Sengupta et al. 1990, Collman

et al. 1991). This pathway is believed to primarily mediate nociception.
ii) Coordinating Region
The nucleus tract solitarii and the neighboring reticuiar substance make up the
portion of the swallowing centre that is considered responsible for the programming of
the entire swdowing sequence (Jean 1984). One level of integration (dorsal) within this
centre is invoIved in both the initiation and organization of the swaüowing sequence. A
second level of organjzation (ventrai) appears to act as a connection pathway to the
various motor neuron pools involved in the swallowuig act, including the integration of
the deglutition reflex with activity in other medullary centres, such as the respiratory
centre.
iii) Efferent Output
The motor neurons involved in the swallowing sequence lie mainly in the
trigeminal, facial and hypoglosd nuclei, the nucleus ambiguus (NA) of the vagus (for
esophageal saiated muscle), and the dorsal motor nucleus of the vagus (DMNV) (for
smooth muscle esophagus) (Fryscak et al. 1984, Carpenter 1989).
Ekîrinsic Innervarion
i) Motor Innervation
The primary extrinsic innervation of the esophagus is via the vagus nerve. Motor
fibres to the striated muscle esophagus &se fiom the vagus, originating in the upper
portion of the neck. The fibers are myelinated and make direct contact with the striated

muscle fibres through the motor end plate (Toyama et al. 1975). Acetylcholine (ACh) is
the neurotransrnitter (NT) involved at the motor end plate, exerting its effects through
stimulation of nicotinic cholinergie receptors on the striated muscle (Toyama et al. 1975).
The vagus nerves also carry preganglionic fibres to the srnooth muscle portion of
the esophagus (including the LES) where they branch out to form the esophageal plexus,
entering the esophagus at different levels (Penden et al. 1950). Preganglionic fibres
within the esophageal wall travel for several centimeters before reaching the
postganglionic neurons in the inttamural plexus, where they release ACh that activates
muscarinic and nicotinic receptors of myenteric nerve ceUs (Kravitz et al. 1978).
Sympathetic efferent pathways play a minor role in esophageal motor h c t i o n
compared to vagal (parasympathetic) efferents. The smooth muscle esophagus and LES
receive sympathetic nerve innemation that arises fiom the ce11 bodies in the
intermediolateral celi columns of spinal segments T4L2 (Coliman et al. 1992). In the
smooth muscle esophagus, sympathetic nerves affect muscle contractility mainiy through
modulation of other neurons (Seelig and Goyal 1978). The density of adrenergic
innervation varies in the esophagus and is lower in the LES than in the more proximal
esophagus (Baumgarten and Lange 1969).
InPinsic Innentaf ion
i)Enteric Nervous Sysfem
In humans, the ENS is estimated to contain 100 miuion neurons (Furness and
Costa 1987). This system controls motiiity, exocrine and endocrine secretions and the
microcirculation of the GI tract. It is also involved in regulating immune and

inflammatory processes (Goyal and Hirano 1996).
There are two major networks of nerve fibres that are intriasic to the gut: the
myenteric (Auerbach's) plexus and the submucous (Meissner's) plexus. The myenteric
plexus provides prirnarily motor innewation to the circula. and longitudinal muscle layers
and is involved in the control of peristaltic activity. Two major neurons are present, an
excitatory neuron with ACh as its neurotransmitter, and an inhibitory neuron with nitric
oxide (NO) as its main neurotransmitter. The submucous plexus contains secretory
newons that play a crucial role in exocrine and endocrine ce11 secretory control in the gut
(Costa and Brookes 1994). Sensory neurons that respond to stretch, tonicity and
temperature are also found in the enteric plexuses.
Another group of cells, the interstitial cells of Cajal (ICCs), are probably of major
functional importance. The interstitial celi was discovered in 1893 by Ramon Y Cajal,
who suggested that it was intercalated anatomically and functionally between autonomie
nerves and egector ceils. ICCs are mononudear poiymorphic cells that branch out
extensively, the branches intersechg to forrn a network throughout which the axons
extend. These branches fonn gap junctions with one another and with smooth muscle
cells and nerve varicosities (Christensen 1993). ICCs bear a certain ultrastructural
resemblance to both smooth muscle cells and fibroblasts, suggesting that they have a
mesenchpal origin which has recentIy k e n confirmed (Burns et al. 1997, Horowitz et
al. 1999). The distribution of ICCs varies throughout the GI tract. In humans and cats,
ICCs are found in both the circular and longitudinai layers of the esophageai smooth
musc le (Faussone-Pellegrhi 1 987, Faussone-Pellegrini and Cortiesini 1 987).

Contro 1 of E s o ~ h a p d Peristaisis
Control of esophageal peristalsis involves a combination of centrai (exainsic) and
penpheral (intrinsic) neuronal mechanisms using an array of putative transmitter
substances, as well as myogenic factors, varying regionaliy within the organ because of
variations in muscular composition. Both neurogenic and myogenic models have been
proposed to explain the control of esophageal peristalsis.
Intarinsic Myogenic Control
The basis for myogenic control of phasic contractility in the GI tract is manifested
by two fundamental charactenstics: i) the electrical control activity, an omnipresent
oscillation of membrane potential ("slow wave") that controls smooth muscle
excitability; and ii) the ability of the smooth muscle cells to communicate with each other
(coupling) in such a way that the entire tissue can operate as a fùnctional unit.
Esophageal contractions occur when the membrane potentid depolarizes above its
excitation threshold. Wnlike other GI smooth muscle, however, esophageal smooth
muscle is electrically quiescent (no slow wave) and does not contract spontaneously
(Crist et al. 1987). Recent studies in muscle strip and whole organ in vitro preparations
have demonstrated that a latent myogenic oscillatory mechanism for control of phasic
contractions exists in esophageal smooth muscle and that it may be activated by
nonspecific excitation of the smooth muscle membrane (Helm et al. 199 1, Helm et al.
1 WZ).
The most direct evidence that the mechanism for generation of phasic contractions
lying within the smooth muscle itself is the hding that in in viîro esophageal

preparations treated with the nerve toxin tetrodotoxin (TIX), the smooth muscle
esophagus is capable of generating propulsive movements in response to direct muscle
stimulation using electrical square waves of long pulse duration (Sarna et al. 1977, H e h
et al. 1992, Preiksaitis & Diamant in press). The myogenic mechanism of this
propagation is believed to operate partially through electrotonic spread of activation by
gap junctions present between smooth muscle cells.
Gap junctions are symmeaical patches of the membrane of two cells, occupied by
special intraceilular channels that facilitate the movement of iniracellular regulatory
molecules and ions form ce11 to celi (Perachia 1980). When an action potentiat occurs
across the membrane of a single-unit smooth muscle, it is rapidly propagated via these
gap junctions to the entire group of intercomected nuscie celis that h c t i o n eiectricaiiy
and mechanically as a fhctional syncytium.
The mechanisms involved in the integration of the myogenic and neural control
systems of peristalsis in the smooth muscle esophagus have yet to be determined. The
role of intramurai inhibitory nerves under this mode1 codd be to inhibit retrograde
peristalsis, mediate descending inhibition in advance of the penstaltic wave, and to
modulate the velocity of myogenic propagation of contractions (Clouse & Diamant
1998). Furthemore, if the ICCs serve as transducers for nerve to muscle signaling, and
as conduction paths for muscle to muscle communication, then they are likely integral
components of any myogenic control system.

13 Regional Differences
Regional ciifferences in muscle characteristics such as the resting membrane
potential have been reported in stomach, small bowel and colon. These regions dso
demonstrate regional differences in the fkequency of the electricai slow wave activity
which plays an important role in regulating cwrdinated motor activity and muscle
contraction responses when electricaliy excited (Daniel et al. 1994). As well, regional
electrophysiological differences are well described and are of fimctional importance
(Sanguinetti & Spector 1997) in many other muscle tissues. Regional diversity has been
documented in the heart (Barry & Nerbome 1996), vascular smooth muscle (Michelakis
et al. 1997), and in neurons (Storm 1993).
In the lower esophageal sphincter (LES) there are regional merences in response
to cholinergic excitation of the clasp and sling smooth muscle in both cat and human
(Preiksaitis et al. 1994, Preiksaitis and Diamant 1994). In response to cholinergic
stimulation, differences between the LES and the esophageal body in CaZ' sources
utilized has dso been shown (Biaacani et al. 1987). In both the cat (Zelcer et al. 1984,
Niel et al. 1990) and opossum (Daniel et al. 1976), RMP of the LES (-40 to -50 mV) is
consistently reported as less negative than that seen in the esophageal body (-50 to -60
mV) (Kaman et al. 1985, Crist et al. 1987, Decktor et al. 1982, Cnst ef al. 1991).
However, within and dong the esophageai body, there has been little investigation
of regional dinerences in smooth muscle properties and Uiformation remains inconsistent.
Decktor and Ryan reported a steady decrease of the RMP dong the opossum circular
muscle esophagus (-52.8 to -43.5 mV) (Decktor et al. 1982); however, others cornparhg
opossum circular muscle esophagus between 8 and 2 cm above the LES have found no

difference in the RMP (Cnst et ai. 1987, Crist et ai. 199 1 ) . Regional differences in
neural mediation have k e n described dong the esophageal body with the excitatory
(cholinergie) influence most prominent proximaily (Cnst et al. 1984, Gilbert et al. 1986,
Seno et al. 1988, Crist et al. 199 1, Dodds et al. 1979, Anad er al- 1994, Yamato et al.
1992, Paterson et al. L991) and the inhibitory (nitrergic-nitric oxide) influence most
active distally (Anad et al. 1994, Yarnato et al. 1992, Conklin et ai. 1995, Murray et al.
1995). However, there is no rnorphologicai nor biochemicd neural evidence of
differences between proximal and distal esophagus that c m explain the ciifTering neural
influences (Serio et al. 1988, Seeling et al. 1984, Rimele et al. 1979, Ny et ai. 1994 &
1 995, Murray et al. 1 994). Salapatek and colleagues (in press) have s h o w that isolated
smooth muscle cells fiom different esophageal regions display ciifferences in electricai
properties in RMP and voltage dependent K' channels. Therefore, regional differences in
muscle properties are present to provide an alternative for the gradients in excitatory and
inhibitory neural mechanisms.
The present experiments were therefore performed to provide another tool for
exploring the regional differences in muscle properties. In single cells, there exist both
passive properties and active properties. The modeling will begi. in this thesis with the
passive properties, which will be studied and presented using sub-threshold Gaussian
White Noise (GWN) and the Volterra-Wiener approach. The active properties can
subsequently be studied using high powered white noise which will evoke action
potentials, and the results c m be analyzed using the parallei NL model.

1.4 Modeling of Physiological Systems
For a linear, causal and time-invariant systems, the input-output relation is
represented by:
where x is the input, y is the output, t is tirne. The variable h(t), the impulse response
function of the system, characterizes the input-output mapping. The duration of the
impulse response function represents the linear system memory which is quantified by
the largest value of t for which the integrand of the equation above is non-zero. The input
impedance can be defmed as
where s is the complex fiequency variable of the input impedance Zjn which is the
Fourier transform (3) of h(t) when xft) is current and y@) is voltage.
The system dynamics white noise is the element of the model that represents
measurement errors, noise and possible inputs. Here we use a filtered version of an
inaccessible white noise process to identiS. the deterministic model, or kernel functional
expressions. Experimentally, th is technique relates the inputs to the outputs using
arbitrary basis functions rather than anything related to underlying physical anatomy and
physiology of the system.

Intemretation of Kemels
The system dynamics are M y described by kemel fwictions. Marmarelis and
Marmarelis (1978) described the set of GWN derived kernels as generalized combined
impulse responses of the system. This is perhaps seen as a descriptor of a system.
Marmarelis and Marmarelis fiirtber described the kemel as a provider of a quantitative
measure of the association or "cross taik" between varying stimuli.
1.5 EXPERIMENTAL OBJECTIVE
Rationale and Aim of Study
Regional differences are important in the regdation of peristaltic waves in normal
gastrointestinal fiuiction of transporthg the bolus (Salapatek et al. in press), and we
speculate that they serve a similar h c t i o n in the esophagus. The inconsistencies dong
the gut are well documented and regional differences may also have a role in myogenic
control of esophageal peristalsis. To develop tbis thesis, the patch-electrode current
clamp technique was used to measure the voltage response in isolated circular smooth
muscle cells dissociated fiom the esophagus of adult felines and to show regional
differences. The present experiment was perfonned to determine if regional differences
in the esophageal body smooth muscle could be detemillied by this biophysical
technique. White noise studies were performed in isolated smooth muscle cells
dissociated from the circular muscle layer dong the esophagus.

2.1 Materials
AnUnal Models
Current understanding of esophageal function is denved primarily fiom animal
studies. There is considerable variation in the anatomy and physiology of the esophagus
among different species. Do~s, rabbits, sheep, cows, guhea pigs, rats, lions and giraffes
have an esophageal body cornposed completely of striated muscle; whereas, primates,
hones, pigs, opossums and cats have a considerable portion of the distal esophagus
composed of smooth muscle (Diamant 1982). Hence in cats, there exists a complex
interplay of both central and intrinsic neural mechanisms, simitar to what is found in
humans. The placement of the gastroesophageal junction relative to the diaphragrn, a
significant percentage of the esophagus that is composed of striated or smooth muscle,
and the cholinergic sensitivity of the smooth muscle esophagus and LES are very similar
in hurnans and cats. For these reasons, the cat was chosen for these investigations.
P harmacoloaical Agents
The enzymes used were tiom Sigma Chernical Co. (St, Louis, MO) and al1 other

chemicals originated from Fisher Scientific Company (Fair Lawn, NI), Mallmckrodt
Specialty Chernicals Company (Paris, Kentucky), or Sigma Chernical Co. (St. Louis,
MO).
2.2 Methods
S~ecific Protocols
Cell Isolation
Adult cats of either sex were killed by intravenously injecting euthanyl(0.5
mgkg), following a protocol approved by the University of Toronto. The entire
esophagus was quickly excised and placed in oxygenated (with 95% 0, and 5% CO3
Krebs solution containing the following composition (in mM): 1 15.0 NaCl, 4.6 KCi, 1.2
NaH,PO,, 1.2 MgSO,, 22.0 NaHCO,, 2.5 CaCl,, and 1 1.0 glucose. M e r removing
connective tissues surrounding the esophagus, it was opened dong the greater cwature
of stomach. The mucosa and the circular muscle were stripped off, leaving the exposed
longitudinal muscle whose strips were dissected out and cut into squares of -2 mm2.
Three to five smooth muscle squares were then placed in 1 mi dissociation solution with a
composition of (in mM) 125.0 NaCl, 5.0 KCI, 1.0 CaCI,, 1.0 MgCl,, 10.0 HEPES, 2.5
EDTA and 10.0 glucose (pH 7.2). The dissociation solution was added 10 pl collagenase
blends F (1 30 mg/ml), 20 pl papain (500 mghl), 10 pl 1,4-dithio-L-threitol(15.4 mg/ml)
and 10 pl BSA (100 mg/ml) and then incubated at 37OC for about 45 minutes. M e r
incubation, tissues were washed wiîh enzyme-free dissociation solution 3 times and

gently agitated with a plastic transfer pipette. Spincile-shaped single SMCs were then
dispersed and used for patch clamp study within the following five hours.
Biophysical Measurements
The patch clamp technique was used to observe cellular electrical activity through
current flow and voltage response. The actual patch clamping of the cells was performed
by Dr. AM Salapatek and J Ji.
Patch pipettes were made fiom thin-walled borosilicate glass capillary tubes (OD
1.5 mm, ID 1.10 mm, Sutter Instrument Co. Novato, CA) and were heat polished prior to
use. They were pulled with a Narishige PP-83 (Tobo, Japan) two stage micropipette
puller and the raw tips were polished with a Narishige MF-83 (Tokyo, Japan) microforge.
The polishing ensures a microscopie smooth opening at the pipette tip. The reference
electrode made fiom Ag-AgC1 wire was directly connected to the bath. Pipettes were
filled with a pipette solution composed of (in mM) 140.0 KCl, 0.5 CaCl,, 1 .O MgCl,, 10.0
HEPES, 5.0 N a m (pH 7.2) pnor to attaching it to the microscope.
Pipettes were fastened ont0 an Axopatch 200B amplifier (Axon Instruments Inc.,
Foster City, Ca.) headstage electrode, which in tum was attached to an inverted
microscope (Olympus CK20, Olympus America, NY, USA). This microscope rested on
a air-cushioned table which was enclosed by a Faraday cage, minuni;ring both extemai
vibrations and electrical interference. Isolated cells in dissociation solution were placed
in a 1-ml glas-bottom dish mounted on the stage of an uiverted microscope and allowed
for 30 min. to adhere to the bottom. The ceils were then washed with extemal solution
containing (in mM) 140.0 NaCl, 5.0 KCl, 2.5 CaCl,, 1.0 MgCl,, 10.0 HEPES, and 5.5

glucose (pH 7.4). The pipette was lowered into the bath using a coarse manipdator and
placed near a SMC using a remote fine-manipulating controller.
Once in the bath, the resistance for each tip opening was calculated using Ohm's
law. Ohm's law States,
where V = potentid difference in volts, V
1 = current in amperes, A
R = resistance in ohms, S2.
The pipette resistance varied between 3 to 6 MR.
Healthy SMCs were chosen on the basis of location and appearance. The most
desired cells were individual cells that consisted of a clean membrane. Prior to patching
onto a cell, the pipette was calibrated (or zeroed) using the pipette offset knob and a "seal
test" 5 mV step was applied in the bath. This was seen as an actual step on the
oscilloscope. Using the fine-manipulating controller, the pipette was 10 wered onto a cell.
As the tip made contact and became obstructed with the ce11 membrane, a slight electrical
resistance occurred appearing as a deflection in the "seal test" current. Gentle suction
was applied on the pipette until a gigaseal(>l GS2) forms with the ce11 membrane. The
disappearance of the "seal test" step indicated that no m e r current flow existed
between the pipette and the ground, and thus a gigaseal was ensured. This is also known
as the cell-attached patch (Fig. 2. l a). Membrane potential (or chamel) characteristics

GEGAOHM
PULSE OF SUCTION OR VOLTAGE
- Whole
ce11
Fi p r e 2.1 Schematic showing procedure for obtaining whole cell patch that can be studied with patch-clamp technique (A) Configuration of ceil-attached patch. Blunt pipette tip pressed agaiast ceii membrane. With suction a gigaseal can be formed that isolates interior of pipette and surface of membrane patch fiom bath solution. (B) Configuration of a whole cell patch. By passing a small pulse of current or suction, it is possible to break the isolated patch while maintaining a gigaohm seal. This provides low-resistance access to ce11 interior. Contents of pipette rapidly exchange with cellular contents (dialyze). One now has control over ionic gradients and trammembrane potential and whole-celi currents can be quantitated.

within the patch can be observed in cell-attached patch con£iguration. Access to the
intracellular space was obtained by additional suction, thus resulting in a whole-ceil
configuration (Fig. 2.1 b). This is the configuration used in the present experiments. Due
to the nature of the technique used, the flow of ions (or current) through the ion channels
of the whole ceii could be measured with stringent accuracy and monitored with a tirne
resolution of milliseconds.
White-Noise Current Injection and Cell Viabiliîy Test
An adjustable high power level current stimulus of 0.4 second duration (400 ms =
7932 points) was injected using pClamp6 software (Fig. 2.2a). The voltage response is
captured and saved in a computer file for analysis later on (Fig. 2.2b).
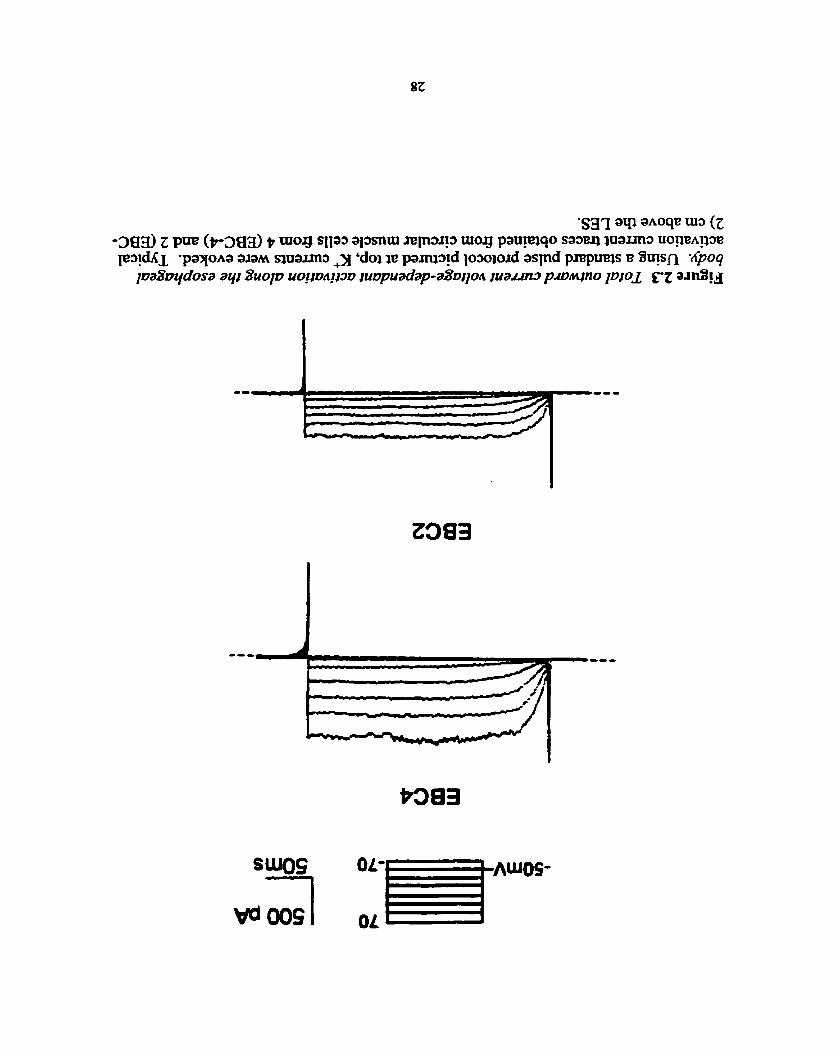
Prier to and following the injection of experimental current, a standard current-
voltage (IV) protocol was applied to the ce11 to detennine the viability of both the ce11 and
the seal. The results will also be used to calculate cellular input resistance for later
cornparison. In essence, this protocol hoIds the ce11 at -50 mV for 235 ms and increases
by 20 mV every 255 ms. Throughout this process, voltage-activated chamels in properly
configured cells wili open and produce an outward current, normally at -40 mV, which
will result in a series of outward currents (Fig. 2.3).
2.3 Data Acquisition and Analysis
Whole ce11 current-clamp protocols were generated by pClamp6 software (Axon
instruments, Foster City, USA). Electmphysiological signals were relayed back to an

O 0.002 0.004 0.006 0.008 0.01
nfrœ (sec)
Figure 2.2 EZectrophysioZogicuI experimenr with Guussian white noise (GWN) input currenr. (A) The GWN input current. (B) The output voltage. (C) The nrst order kemel and close up of kemel (inset). @) The input impedance, a Fourier transform fiom time domain to frequency domain and close up of area of interest (inset).

oscilloscope where ceil characteristics were observed while data were low-pass filtered at
10 kHz by an on-board eight-pole Bessel filter before digitization with a DigiData 1200
analog-to-digital converter (Axon Instruments). Recordings were commenced 2-5 min.
after the formation of whole cell configuration in order to allow for ce11 dialysis with the
pipette solution. Al1 experiments were performed at r w m temperature of 20-22OC.
The technique chosen for kemel computation was Fast Orthogonal Algorithm
(FOA). The FOA was implemented using an algorithm developed by Korenberg (1988)
and adapted to our cornputers by William Neil Wright (1993). Utilization of the FOA is
advantageous because it requises no apriori estimates of kemel memory length. This
technique involves the creation of an orthogonal set of fùnctions fiom the actuaî input
sequence used (GWN) and these fùnctions are used to synthesize the kemel. Kernels
were calculated with 1024 lags which adequately allows the memory of the kemel to
decay to zero (Fig. 2.2~).
Fourier Transform
The kemel data was transformed fiom time domain to fkquency domain (Fig.
2.2d) in order to determine the input impedance fiom which the input resistance, the cut-
off fiequency and the order of the system c m be estimated. The input resistance is a
measure of the ce11 structure and is quantified by the magnitude of the input impedance at
zero fiequency. The cut-off tiequency is a measure of the responsiveness of a ce11 which

has low pass fiequency characteristics. The kquency that matches the 3 dB level of
impedance, a point when impedance reaches 0.707 of its maximum, is the cut-off
fiequency. The order of the system is indicative of the amount of exhibited RC circuit-
Iike characteristics which tells us the rate of attenuation of the cell. This can be shown as
sirnpiy the dope at which the impedance decreases as fiequency increases.
Statistical Methods
The data will be analyzed to determine if there is a significaat difference in input
resistance, cut-off fkequency and the order of the system. Since a small sample size was
obtained, the t-distribution should be used. Using at les t a 5% level of significance, the
nul1 hypothesis will be that the two regions, EBC-2 and EBC4, are the same.

3.1 Equipment Validation
Mode1 Ce11
First of dl, the system undenvent testing to determine its fiinctionality and its
accuracy. A model ce11 (PATCH-1 U, Axon Instruments) was provided by the
manufacturer of the amplifier dong with general information regarding its senings and
parameters. The purpose of this exercise was to see if the parameters of the model ce11
could be captured through white noise current injection and analysis. The model ce11 was
reported to have a time constant of 16.5 ms or a cut-off fiequency of 60.6 Hz. The time
constant is the inverse of the cut-off fiequency, the fiequency that matches approximately
0.707 of the maximum magnitude of impedance fiom a fiequency domain plot. The cut-
off fiequency of the model ce11 was about 65 Hz, a tirne constant of 15.4 ms (Fig. 3.1).
EIectrode
The electrode was tested to see if its response characteristics were adequate to
capture the output noise that it will be provided throughout the experiment. The effect of
the electrode shouid not dominate; in other words, the glass tip should not be too small so

Magnitude of Input lmpedance (Model Cell)
Fmquency (Hz)
Figure 3.1 The cut-offfiequency of mode2 cell. The cut-off fiequency of the mode1 celi was 65 Hz ( t h e constant 15.4 ms) which closely matches the parameters &en by the manufacturer with a t h e constant of 16.5 ms. (note: cut-off fiequency is the inverse of t h e constant)

that it causes resistance in measuring the injected current An electrode was lowered into
the bath medium and white noise current was applied in order to record the characteristics
of the electrode. Under the kquency domain, the results indicated that the electrode far
exceeded EBC nom both levels in cut-off fkquency and consequently possessed a faster
tirne constant in the ps range (Fig. 3.2). ï hus the electrode tip was small enough to patch
and yet not hinder (or swamp the noise artifact) the white noise current.
3.2 White Noise Injection
After validating the system, cells fiom both regions were patched, injected with
white noise current and its voltage response were recorded and analyzed. In total, five
cells fiom each region (ten experirnents in total) were successfully patched, studied and
reported in this section.
First Order Kemels
First order kernels were calcdated and plotted against t h e (Fig. 3.3). It is
already apparent that the two kemels show two distinct patterns generated fkom the two
regions. Generally, cells h m two centimeters above LES (EBC-2) displayed a much
slower decay curve than those isolated fiom EBC-4.
Fourier Transform
Fourier transforms of the kemels fiom time domain to kquency domain contain a

Magnitude of Input lmpedance (Elecaode vs EBC4)
Fmquency (Hz)
Figure 3.2 The time constant of the eleclrode. Under the fiequency domain, the results indicated that the electrode far exceeded EBC-4 in cut-off fkequency with a time constant of < 1 .O ms. Thus the electrode tip was srnail enough to patch and yet not hinder the white noise current.

l
O 0.001 0.002 0.003 0.004 0.005 0.006 0.007 0.008 0.009 4 . 1 -
Tirne (sec)
Figure 3 3 First order kernelfiom two regionS. Fùsî order kemels were calculated and plotted against t h e . Cells fkom four centimeters above LES (EBC-4) have a steeper decay cuve than those h m EBC-2.

wealth of information about the cells that can be elucidated by foilowing these methods.
Input Resistance
Input resistance of a ce11 is the impedance value at zero fiequency (Fig. 3.4). The
resistance values obtained were 3 10.8 i 42.6 MS2 and 39.1 f 36.9 MS2 for EBC-2 and
EBC-4 respectively.
i ) Classical Technique
In order to confirm these input resistance values, the numbers were compared
with input resistances derived from standard means which were accomplished by voltage
clamping the cells and recording K' outward current (Fig. 2.3). The data was taken,
plotted by linear regession and a line was fitted dong a linear area of Iow v~ltages since
nonlinear data should not be included. Small hypepolarizing pulses (-30 mV to 10 mV)
were chosen as to not take in effect of inward K+ rectifiers. The foilowing table displays
both results (Table 1).
The standard technique showed varying values of resistances with a mean value + SE of 290.3 k 147.9 MS2 and 20.7 + 1.1 Ml2 for EBC-2 and EBC-4 respectively. The
maximum percentage difference was 40 % with t-tests indicating no significant
differences between the two measurements.
Cut-Off Frequency
The cut-off fiequency is the fiequency that matches the 3-decibel level of
impedance (Fig. 3.5). The 3 dB is approxirnately 0.707 of the maximum magnitude of

Magnitude of lnput Impedance
lnput resistance
O 500 lm 1500 2000 2500 3000 3500
Frequency (Hz)
Figure 3.4 Input resistance derivedfi.om input impedance plot. input resistance of a ce11 is the impedance value at zero fiequency. The values obtained were 3 10.8 f 42.6 MR and 39.1 + 36.9 MR for EBC-2 and EBC-4 respectively.

Table 3.1 Input resistances Smooth Muscle
Type
EBC-2
E B C 4
White Noise Technique (Msr) 365 294
In calcuiation of percentage difference, standard technique was taken as tme value.
19.4 21.2 21.5
18.3 23.4 24.7
5.7 10.4 14.9
Standard Technique @lm 461 210
Percentage Dwerence
20.8 40.0

rsspsdive tim constants
3dB cut-Off
EBG2
O 0.0001 O.Oûû2 0.0003 0.OOM 0.0005 0.- 0.0007 0.0008 0.0009 0.001
Time (sec)
Figure 3.5 Cut-oflfiequency and time constant of the cells. (A) The cut-off frequency is the fiequency that matches the 34ecibel level or 0.707 of the maximum magnitude of impedance. EBC-2 and EBC-4 redteà in highly variable cut-off fkquencies of 200.6 k 55.5 Hz and 1287.0 f 1 65.2 Hz respectively. (B) The tirne constant is the time that matches the 3-decibel level of the nrst order kemel. EBC-î and EBC-4 resulted in time constants of 2.9 +, 2.2 ms and 0.9 f 0.4 ms respectively .

impedance fiom a fkequency domain plot. The t h e constant of the membrane is
inversely proportional to the cut-off fiequency. EBC-2 and EBC-4 resulted in average
cut-off fiequencies of 200.6 Hz and 1287.0 Hz respectively. These averages did not
follow the normal distribution. Therefore, more accurate tirne constants (2.9 + 2.2 ms
and 0.9 + 0.4 ms respectively) were obtained by taking the 3 dB fiom the kemei (Fig.
3.5b).
Slope of Fourier Transform (Roll Ofl
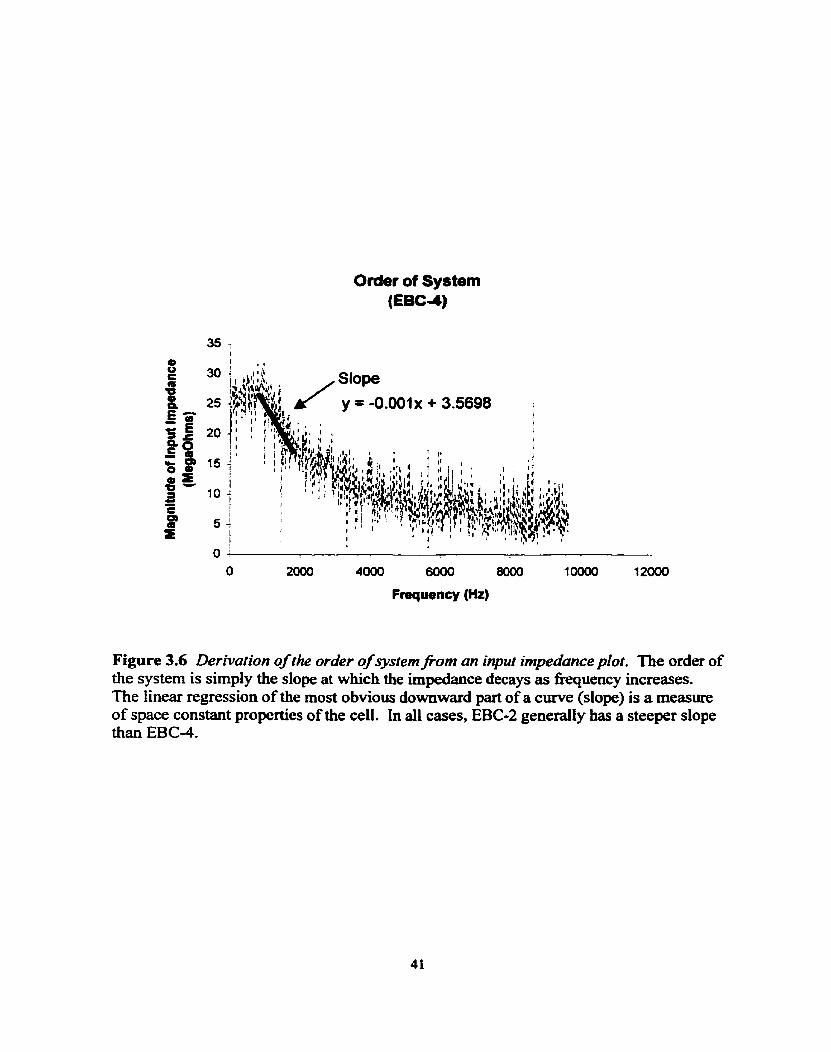
The order of the system is simply the slope at which the impedance decays as
fiequency increases. A simple technique exists for obtaining an approximate plot of the
magnitude and phase of a transfer fhction by using diagrams called Bode plots. On a
plot of decibels versus log fiequency, one can get straight-line plots or one can get the
linear regression of the most obvious downward part of a simple curve (Fig. 3.6). This
slope is a measure of the average order of a system consisting of RC networks. As the
slope gets steeper, there is an indication of a higher space constant and the presence of a
higher order in the cable properties of the cell. In al1 these cases, EBC-2 generally has a
steeper slope than EBC-4.
3.3 Statistical Analysis
Between EBC-2 and EBC-4
Using a two-tail t-test ofsignificance, al1 pairs of data (Table 2) were examined to
Order of System (EBC4)
Slope
y = -0.001~ + 3.5698 i 1 I l 1 . i
r
I
Frequency (Hz)
Figure 3.6 Derivation of the order of system f'om an input imped4nce plot. The order of the system is simply the dope at which the impedance decays as fiequency increases. The Iinear regessian of the rnost obvious dowuward part of a curve (slope) is a mesure of space constant properties of the cell. In al1 cases, EBC-2 generally has a steeper dope than EBC-4.

Table 3.2 Summary of remfis Smooth Muscle
Type
EBC-2
Average (EBC-2) ,
Average (EBC-4)
Input Resistance (MQ) 270
1 1 I - - -
Each row represents values of an individual cell,
T h e Constant (ma 1 -3
347 365 294 278
310.8 + 42.6
-0.00 1 5 -0.0009 -0.00 1 O
-0.0029 f 0.004
1
Slope of Input Impedance ( M W )
-0.0823
18.3 23.4 24.7
39.1 t 36.9
6.8 1 -0.0228 2.5 -0.0135
1.4 0.6 0.8
0.9 + 0.4
1 -9 2.0
2.9 + 2.2
-0.082 1 -0.0805
4.056 f 0.04

see if there was a significant difference betweea them. The test statistic, t, was
detennined by
where
and F = tested mean a= standard error n = sarnple sîze
In the present case, the t value for input resistance, tirne constant and slope were 10.780,
2.002 and 2.953 respectively. By using a 95 % codidence level, the hrpothesis would be
rejected if t > 1.86; therefore, a significant difference between EBC-2 and EBC-4 is
present in al1 three parameters: input resistance, cut-off fiequency and slope.

4.1 Discussion
These studies demonstrate for the fhst t h e that white noise c m be used on single
smooth muscle cells, in this case from the smooth muscle esophagus. The white noise
technique proved to be a valuable tool in detemiining two aspects of the celi not usually
characterized by other means, the cut-off fiequency (tirne constant) and the differences in
the average order of a system (space constant) between the two regions of the circular
muscle of the esophageal body.
Eaui~ment Validation
First of dl , the system undenvent tests to determine its hctionality and its
accuracy. The mode1 ce11 was docurnented to have a time constant of 16.5 ms in which
the analysis confirmed it at 15.4 ms. The 1. t rns error could have risen through
extrapolating the information from the plot where a polynornial curve was fitted, and
perhaps the curve fitting software was inadequate for our needs. Until a better curve fit
tool is acquired, a 5 % error demonstrates that the system is most reliable. Therefore the
parameters of the mode1 ce11 can reasonably be captured through white noise current

injection and analysis.
The electrode was also tested to see if the speed of electrode was adequate to
capture al1 the noise that will be provided. The results showed that the time constant of
the electrode was safely < 100 ps, shorter than the quickest celi recorded at around 0.5
ms. If the results were reverse& the electrode would not have been able to record the
cell, instead it would in essence have recorded itseff. By measuring the characteristics of
the electrode, it was determiaed that the electrode tip was suitable for experimentation.
Differences Between EBC-2 and EBC-4
This report shows evidence of regional differences in circuiar smooth muscle
properties dong the esophageal body. It was apparent by the two kemels that the cells
fiom the two regions generated two distinct patterns. Kemels of ceils fiom EBC-2 had a
much slower decay curve than those fiom EBC-4. The regionai diversity includes
differences in input resistance, ciifferences in cut-off fiequency and differences in order.
Proximally, the cells are more active, posses higher input resistaaces, demonstrate
quicker time constants (more responsive) and have smaller space constants.
Input Resistance
The input resistance is a measure of the resistive components of the cell. Input
resistances obtained were 3 10.8 f 42.6 MC2 and 39.1 + 36.9 MC2 for EBC-2 and EBC-4
respectively. Using a two-tail t-test of significance, the pair of data resulted in a
significant difference, as much as 99.9%, between them. The differences perhaps are due

in part to some ion channel differences within the ceils of both regions. Evidence of
differences dong the cat esophagus have been found and implicate diverse voltage
dependent K+ channel densities dong the esophagus (Salapatek in press). The density is
greater at EBC-4.
In order to ver* the white noise technique on isolated smooth muscle celis, input
resistances were caiculated the standard way by voltage clamping the cells. Upon
comparing the two sets, there were varying differences in magnitude among them that
range fiom 5% to 40 %. Calculating voltage-clamped input resistances were quite
arbitrary because a linear regression is fitted dong data points derived fiom a series of
hyperpolarizing pulses (-50 mV to 70 mV). The 10 to 70 voltage range, wbich represents
K+ rectiQing channel activity, was cut off leaving a few points to calculate conductance.
This method accounts for the high variability of the input resistances taken using standard
techniques. The white noise method is advantageous because from five samples, a
maximum standard error of 40 MC2 was recorded compared to a standard error 150 MR
using standard means. Upon using both white noise curent inputs and voltage outputs,
the input impedance is estimated using theoretical and mathematicd grounds, instead of
relying on numerous experimental trials and data.
Time Constants
The tirne constant is a measure of the responsiveness of a ce11 which has low pass
fiequency characteristics. EBC-2 and EBC-4 resulted in time constants of 2.9 ms and 0.9
ms respectively. Systems with lower memory normally have a higher cut-off fiequency

which also relates directly to how the ce11 responds to a given input. The striated region
is neurally mediated, which requires muscle cells to quickly respond to neural messages
and sometimes quick changes of action (Le. vomiting). Since EBC4 is doser to the
striated, these findings support the notion of a muscle gradient dong the esophagus where
reaction tirne dimiaishes as one moves distally nom striated muscle to smooth muscle.
Other regions in the gut have been found to have varying time constants. In the
canine gastric antrai circular muscles, the time constant of the circular muscle near the
myenteric plexus tended to be greater than that of the circular muscle near the submucosa,
however this clifference was not statistically different (Bauer & Sanders 1986). Their
experiment was performed on tissue where ceils are coupled.
Order of System
The average order of the system is indicative of the amount of RC circuit-like
characteristics and tells us the rate of responsiveness at which a ce11 is capable of
handling. Generally, the space constant is a fùuction of the dope of the input impedance
which is a fünction of the axial and membrane resistances of the cell. EBC-2 generally
has a steeper slope than EBC-4 meaning that EBC-2 has an overall higher space constant
and the presence of a higher order in the cable properties of the cell. Since the space
constant is the distance at which the signal travels as it attenuates, a quicker responding
ce11 shouid ideally have lower memory, shorter space constant. This lack of memory
allows for the celi to quickly respond to new stimuli. From the aspect of space constants,
regional differences exist in antral circula. muscles (Bauer & Sanders 1986). The length
constant of the circular muscle near the myenteric plexus (2.4 mm) was significantly

greater that the space constant of the circular muscle near the submucosa (1 -7 mm).
The diversity is seen as important in other muscle tissues such as those in the
heart, vascular smooth muscle and in neurons. In the esophagus, excitatory (ACh) and
inhibitory neurotransmitters (NO) have been implicated as the main mechanism for
esophageal controt by depolarking or hyperpolaripng cells, and show regional
ciifferences in effects not accounted for by the neural elernents themselves. The evidence
of regional muscle differences therefore provides a rational basis for the fiinctional
differences in neural effects, and is key in understanding how myogenic control
contributes to esophageal peristalsis. Perhaps anned with the understanding of myogenic
properties, new therapies can be developed at targeting the esophageal muscle itself in
helping people with such disorders as gastroesophageal reflux disease and spastic motor
disorders.
4.2 Future Investigations
This study should be taken on to M e r levels. The white noise technique
illustrates the dynamics of a ce11 by capturing not only the passive properties, but the
active ones. The classic way of plotting N cuves is by changing the voltage and
recording current. However, in this method, we can change the current and record the
voltage and therefore capture a nodinear c w e . This technique has been utilized in
hippocampal neurons (Bardakjian et al. 1994). Future studies in smooth muscle provide
the potential for similar assessments.

This technique may also be used to identifjr the system mode1 and to provide
passive electrïcal properties such as ce11 capacitance and coupihg resistance in a tissue of
coupled cells. This involves an optimization of the system (Fu et al. 1991), and holds the
potential to provide new insight into the coupling mechanisms of smooth muscle celis.

Al Yassin, T.M. and P.G. Toner- Fine structure of quamous epithelium and submucosal glands of human esophagus. J. Anal. 123:705,1977.
Anand, N. and W.G. Paterson. Role of nitric oxide in esophageal peristalsis. Am. J. Physiol. 266:G123-G13 1, 1994.
Arey, L.B. and M.J. Tremaine. The muscle content of the lower esophagus in man. Anat Rec. S6:3 15, 1933.
Asoh, R. and R.K. Goyal. Manometry and electromyography of the upper esophageal sphincter in the opossum. Gastroenterology 745 14-520, 1978.
Bardakj ian, B .L., Wright, W.N., Valiante, T.A. and P.L. Carlen. Nonlinear system identification of hippocampal neurons. In: Advanced Methods of Physiological System Modeling, Vol. 3. V.Z. Marmarelis (ed.). New York, Plenum Press, pp. 179- 194, 1994.
Barry, D .M. and J.M. Nerbonne. Myocardial potassium channel: elecrophy siological and molecular diversity . Annu. Rev. Physiol. 58:363-394, 1 996.
Baumgarten, M.G. and W. Lange. Adrenergic innervation of the oesophagus in the cat (Felis dornestica) and rhesus monkey (Macacus rhesirs). 2. Zellorsch. Mikrosk Anat. 95529-545, 1969.
Bauer, A. J. and K.M. Sanders. Passive and active membrane properties of canine gastric antral circular muscles. Am. J. Physiol. 25 1 :C268-C273, 1 986.
Biancani, P. and J. Behar. Esophageal Motor Function. In: Textbook of Gastroenterology, Section B. Motility, Second edition, Ed. Tadataka Yamah. J.B. Lippincott Company, PPhiladelphia, 1 995.
Biancani, P., Hillemeier, C., Bitar, K.N. and G.M. Makhiouf. Contraction mediated by Ca2+ release in tbe LES. Am. J. Physiol. 253:G760-G766, 1987.

Biancani, P., Zabinski, M.P., Kernstein, M. and J. Behar. Lower esophageal sphincter mechanics: anatomic and physiologie relaîionships of the esophagogastric junction of cat. Gastroenterology 82:468475, 1982.
Biancani, P., Zabinski, M. and J. Behar. Pressure, tension, and force of closure of the human lower esophageal sphincter and esophagus. J. Clin. Invest. 56:476, 1975.
Burns, A.J., Herbert, T.M., Ward, S.M. and K.M. Sanders. Interstitial ceils of Cajal in the guinea-pig gastrointestinal tract as revealed by c-Kit immunohistochemistry. Cell and Tissue Res. 290: 1 1 -20, 1997.
Carpenter, D.O. Central nervous system mechanisms in deglutition and emesis. In: Handbook of Physiology. Section 6: The Gastrointestinal System, edited by S.G. Physiological Society, pp. 685-7 14, 1989.
Christensen, J. The enteric nervous system. In: An illustrated guide to gastrointestinal motility. D. Kumar and D. Wigate (eds). New York: Churchill Livingstone, pp. 10-31, 1993.
Christensen, J. and R.L. Roberts. Differences between esophageal body and Iower esophageal sphincter in mitochondria of smooth muscle in opossum. Gastroenrerology 85:650-656,1983.
Cloue, R.E. and N.E. Diamant. Motor physiology and motor disordea of the esophagus. in: Feldman, M., Scharachmidt, B.F. and Sleisenger, M.H. Sleisenger & Fortran's Gastrointestinal and Liver Disease, 6: 467-497, Philadelphia, W.B. Saunders, 1998.
CoIIman, P.I., Tremblay, L. and N.E. Diamant. The distribution of spinal and vagal sensory neurons that innervate the esophagus of the cat. Gasfroenterology 1 O3 :8 17-822, 1992.
Collman, P.I., Tremblay, L. and N.E. Diamant. Distribution of vagal and spinal sensory neurons to the esophagus of the cat [Abstract]. Gastroenterology 100:A432, 1991.
Conklin, J.L., Murray, J., Ledlow, A., Clark, E., Hayek, B., Picken, H. and G. Rosenthal. Effects of recombinant human hemoglobin on motor fhctions of the opossum esophagus. Journal of Phurmacology and Ekperimentul Theropeutics 273:702- 767, 1995.
Costa, M. and S.J.H. Brookes. The Enteric Nervous System. Am. J Gastroenter. W(8):S 129-S 137, 1994.

Craemer, B. and J. Schlegel. Motor responses of the esophagus to distention. J. Appl. Physiol. 10:498-504, 1957.
Crist, J., Kauvar, D. and R.K. Goyal. Gradient of cholhergic innervation in opossum esophageal circular smooth muscle. Gullet 1 :92-98, 199 1.
Crist, J., Surprenant, A. and R.K. Goyal. Intracellular studies of electrical membrane properties of opossum esophageal circular smooth muscle. Gastroenterology 92:987-992, 1987.
Crist, J., Gidâa, J.S. and R.K. Goyal. Intramural mechanism of esophageal peristalsis: rules of chohergic and noncholinergic nerves. Proceedings of ihe National Academy of Sciences USA 8 1 :3 595-3 599, 1 984.
Csendes, A., Hemiquez, A. and P. Csendes. Length of the human esophagus in vivo during manometric studies. Diseuses of the Esophagus 6: 1 7- 19, 1993.
Daniel, E.E., Bardakj ian, B.L., Huizinga, J.D. and N.E. Diamant. Relaxation oscillators and core conductor models are needed for understanding of GI electrical activities. Am. J. Physiol. 266:G3 39-G349, 1994.
Daniel, E.E. and V. Posey-Daniel. Neurornuscular structures in opossum esophagus: role of interstitial cells of Cajal. Am. J. Physiol. 246 (Gastrointest. Liver Physiol. 9): G305-G3 15, 1984.
Daniel, E.E., Taylor, G.S. and M.E. Holman. The rnyogenic basis of active tension in the lower esophageal sphincter (abstr). Gastroenterology 70:874, 1 976.
Deca.de, D.J., Szabo, A.C. and J. Christensen. Temperature dependance of responses of esophageal smooth muscle to electrical field stimulation. Am. J. Physiol. 232:E432-E436,1977.
Decktor, D.L. and J.P. Ryan. Trammembrane voltage of opossum esophageal smooth muscle and its response to electrical stimulation of intrinsic nerves. Gastroenterology 82:30 1 -3 08, 1 982.
DeNardi, F.G. and R.H. Riddell. The Normal Esophagus. Am. J Swg. Pathol. 1 S(3):296-309, 1991.
Diamant, N.E. Physiology of Esophageal Motor Function. Gasiroenterol. Clin. N. Am. 18(2):179-194, 1989.
Dodds, W.H., Christensen, J., Dent, J., Arndorfer, RC. and J.D. Wood. Phannacologic investigation of prhary peristalsis in smooth muscle portion of opossum esophagus. Am. J. Physiol. 237:ES6 1 -E566,1979.

Dodds, W.J., Hogan, W.J., Reid, D.P., Stewart, E.T. and R.C. Amdorfer. A cornparison between primary esophageal peristaisis following wet and dry swallows. J. Appl. Physiol. 35:85 1-857, 1973.
Dooley, C.P., Schlossmacher, B. and J.E. Valenmela. EEects of alterations in bolus viscosity on esophageal peristalsis in humans. Am. J. Physiof. 254:G8, 1988.
Ellis, F.H. Upper esophageal sphincter in health and disease. Surgi Clin. N. Am. 5 1 553- 565, 1971.
Faussone-Pellegrini, M.-S. Comparative study of Interstitial Cells of Cajal. Acta Ana. 130:109-126, 1987.
Faussone-Pellegrini, M.-S . and C. Cortiesini. Ultt.astnictural feanires and localization of interstitiai cells of Cajal in the smooth muscle coat of the human esophagus. J Subrnicrusc. Cytol. 1 7 : 187- 197, 1987.
Fleshler, B., Hendrix, T.R., Kramer, P. and F.J. Ingelfinger. The characteristics and sirnilarity of primary and secondary peristalsis in the esophagus. J: Clin. Imest. 38:llO-116, 1959.
Fryscak, T., Zenker, W. and D. Kantner. Merent and efferent innervation of the rat esophagus. A tracing study with horseradish peroxidase and nuclear yellow. Anat. Embryofogy 170:63-70, 1984.
Fu, P. and B.L. Bardakjian. System identification of electrically coupled smooth muscle cells: the passive electrical properties. IEEE Tram- Biomeci Eng. 3 8 : 1 1 30- 1 140, 1991.
Furness, J.B. and M. Costa. The enteric nervous system. Edinburgh: Churchiü Livingstone, 1987.
Gilbert, R.J. and W.J. Dodds. Effect of selective muscarhic antagonists on peristaltic contractions in opossum smooth muscle. Am. J. Physiol. 250:GSO-G59, 1986.
Goyal, R.K. and 1. Hirano. The Enteric Nervous Systern. N. Engl. J . Med 3 M(l7): 1 106- 1115,1996.
Goyal, R.K., Sangree, M.H. and T. Hersh. Pressure inversion point at the upper high pressure zone and its genesis. Gmtroenterolop 59:754-759, 1970.
Helm, J.F., Bro, S.L., Dodds, W.J., Sarna, S.K. and R.G. Hofhann. Myogenic mechanism for peristalsis in opossum smooth muscle esophagus. Am. J. Physiol. 263 (Gastroktest. Liver Physiol. 26): G953oG959, 1992.

Helrn, J.F., Bro, S.L., Dodds, W.J., Sarna, S.K., HoiTinaun, R.G. and RE. Amdorfer. Myogenic osciliatory mechanism for opossum esophageal smooth muscle contractions. Am. J. Physiol. 26 1 (Gastrointest. Liver Physiol. 24): G377-G3 83, 1991.
Hollis, J.B. and D.O. Castell. Effect of dry and wet swailows of different volumes on esophageal peristalsis. J. AppI. Physiol. 28: 1 16 1 - 1 164, 1975.
Horowitz, B., Ward, S.M. and K.M. Sanders. Cellular and molecular basis for electrical rhythmicity in gastrointestinal muscles. meview] A m Rev. of Phys. 6 1 : 19-43, 1999.
Humphries, T. J. and D.O. Castell. Pressure profile of esophageal peristalsis in normal humans as rneasured by direct intraesophageal transducers. Am. J. Dig. Dis. 22: 641 -645,1977
Janssens, J., Valembois, P., Vantrappen, G., Hellemans, J. and W. Pelemans. 1s the primary peristaltic contraction of the canine esophagus bolus-dependent? Gastroenterology 65:750-756, 1 973.
Jean, A. Brainstem organization of the swailowing network. Brain Behav. Evol. 25: 109, 1984.
Kahrilas, P.J. Functional anatomy and physiology of the esophagus. In: Castel, D.O., ed. The Esophagus. Boston: Little, Brown and Company, pp. 1-27? 1992.
Kannan, M.S., Jager, L.P. and E.E.Danie1. Electrical properties of smooth muscle ce11 membrane of opossum esophagus. Am- J. Physiol. 248:G342-G346, 1985.
Korenberg, M.J. Ident-g nonlinear dserence equation and fbctional expansion representations: the fast orthogonal algorithm. Annals of Biomed Eng. 16: 123- 142,1988.
Kravitz, J.J., Snape, W.J. and S. Cohen. Effect of thoracic vagotomy and vagal stimulation on esophageal function. Am. J. Physiol. 234:E359-E364,1978.
Lerche, W. The esophagus and pharynx in action. Springfield, IL: Charles C. Thomas, 1950.
Li, C.G. and M.J. Rand. Evidence for a role of nitric oxide in the neurotransmitter system mediating relaxation of the rat annococcygeus muscle. Clin. Exp. Pharmacol. Physiol. 16:933-93 8, 1989.
Liebemann-Meffert, D., Allgower, M., Schrnid, P. and A.L. Blum. Muscular equivalent

of the lower esophageal sphincter. Gasrroenterology 76:3 1-3 8, 1979.
Marmarelis, PZ. and V.Z. Mannarelis. Analysis of Physiologicai Systems. New York, Plenum Press, 1978.
Matmarelis, V.Z. (ed.) Advanced Methods of Physiologicai System Modeling, Vol. 3. New York, Plenum Press, 1994.
Meyer, G.W., Austin, R.N., Brady, C.E. and D.O. Castell. Muscle anatomy of the human esophagus. J. Clin. Gastroenferoi. 8(2): 13 1-4, 1986.
Michelakis, ED-, Reeve H-L., Huang, J M , Tolarova, Nelson, D-P-, Weir, E.K. and S.L. Archer. Potassium channels diversity in vascular srnooth muscle cells. Can. c/: Physiol. Pharmacol. 75:889-897, 1997.
Murray, J.A., Ledlow, A., Launspach, J-, Evans, D., Loveday, M. and J.L. Conklin. The effects of recombinant human hemoglobin on esophageal motor function in humans. Gastroenterology 109: 124 1-1 248, 1995.
M ~ y y J.A. and E.D. Clark. Characterization of nitric oxide synthase in the opossum esophagus. Gastroenterology 106: 1444-1 450,1994.
Netter, F.H. Digestive system: upper digestive tract. In: The CIBA Collection of Medical Illustrations, Ed. E. Oppenheimer. New York: CIBA hblishing, 1971.
Niel, J.P. and J.P. Miolan. Involvement of a cholinergie mechanism in the sustained depolarization and contraction of the lower oesophageal sphincter muscle cells in the cat. Neuroscience 36:803-809, 1990.
Ny, L., Alrn, P., Larsson, B., Ekstrom, P. and K.E. Andersson. Nitric oxide pathway in the cat esophagus: localization of nitric oxide synthase and fimctional effects. Am. J. Physioi. 268:G59-G70, 1 995.
Nyy L., Alm, P., Ekstrom, P., Hannibal, J., Larsson, B. and K.E. Andersson. Nitric oxide synthasecontaining, peptide-containing, and acetylcholinesterase-positive nerves in the cat lower oesophagus. Hostochernical Journal 26:72 1-733, 1994.
Paterson, W.G., Hynna-Liepert, T.T., and M. Selucky. Cornparison of primary and secondary esophageal peristalsis in humans: effects of atropine. Am. J. Plîysiol. 260:G52-G57,199 1
Peden, J.K., Schneider, M.D. and R.D. Bickel. Anatomic relations of the vagus nerves to the esophagus. Am. J Surg. 80:32-34, 1950.
Perachia, C. Structural correlates of gap junction permeation. ?nt. Rev. Cyfol. 66:8 1,

Preiksaitis, H.G. and N.E. Diamant. A myogenic mechanism for peristalsis in the cat. Subrnitted to Am. J. Physiol. In press.
Preiksaitis, H.G. and N.E. Diamant. Regional ciifferences in cholinergic activity of muscle fibers fiom the human gastroesophageal junction. Am. J . Physiol. 272:G1321-G1327,1997.
Preiksaitis, H.G. and N.E. Diamant. Phasic contractions of the muscular components of human esophagus and gastroesophageal junction in vitro. C m J: Physiol. Phmucol . 73 :3 56-363, 1995.
Preiksaitis, H.G., Tremblay, L. and N.E. Diamant. Cholinergic responses in the cat lower esophageal sphincter show regional variation. Gastroenterology 1 O6:3 8 1-3 88, 1994.
Reynolds, R.P .E., El-Sharkawy, T.Y. and N.E. Diamant. Oesophageal peristalsis in the cat: the role of central innervation assessed by transient vagal blockade. Can. J Physiol. Pharmacol. 63 : 122- 130, 1985.
Reynolds, R.P.E., El-Sharkawy, T.Y. and N.E. Diamant. Lower esophageal sphincter function in the cat: role of central innervation assessed by transient vagal blockade. Am. J: Physiol. 246:G666-G674,1984.
Rimele, T.J., Rogers, W.A. and T.S. Gaginelia Characterization of muscarhic cholinergic receptors in the lower esophageal sphincter of the cat: binding of [3 HJquinuclidinyl benzilate. GastroentemZogv 77: 1225- 1234, 1979.
Salapatek, A.M.F., Ji, J. and N.E. Diamant. Ion chamel diversity dong the feline smooth muscle esophagus. In press.
Salapatek, A.M.F., Lam, A. and E.E. Daniel. Calcium source diversity in canine lower esophageal sphincter muscle. Journal of Pharmacology and Erperimental Therapeutics 287:98- 1 06, 1998.
Sanguinetti, M.C. and P.S. Spector. Potassium chanwlopathies. NeurophmmacoZo~ 35:755-762,1997.
Sama, S.K., Daniel, E.E. and W.E. Waterfall. Myogenic and neural control systems for esophageal motility. GastroenteroZogy 73: 1345-1 352, 1977.
Seelig, L.L. Jr., Doody, P., Brainard, L., Gidda, J.S. and R.K. Goyal. Acetylcholinesterase and choline actyltransferase staining of neurons in the opossum esophagus. Anat Rec. 209: 125-1 30,1984.

Seelig, L.L., Jr. and R.K. Goy al. Morphological evaluation of opossum lower esophageal sphincter. Gastroenterology 755 1-58, 1978.
Sengupta, J.N., Saha, J.K. and RK. Goyal. Stimulus-response function studies of esophageal mechanosensitive nociceptors in sympathetic affierents of opossum. J Neurophys. 63 (3):796-8 1 2, 1 990.
Sengupta, A., Paterson, W.G. and R.K. Goyal. Atypical localization of myenteric neurons in the opossum lower esophageal sphincter. American Journal o f Anatomy 180:342-348, 1987.
Serio, R. and E.E. Daniel. Electrophysiological analysis of responses to intrhic nerves in circula. muscle of opossum esophageal muscle. Am. J. Physiol. 254:G107- G116,1988.
Sobotta, J. and F. Hammersen. Histology, Color Atlas of Microscopie Anatomy. 3" ed. Munich: Kastner & Callwey, 1985.
Sohn, U.D., Han, B., Tashjian, A.H., Behar, J. and P. Bimcani. Agonist-independent, muscle-type specific signal transduction pathways in cat esophageal and lower esophageal sphincter circular smooth muscle. J. Pharmacol. &p. Ther. 273:482- 491, 1995.
Storm J.F. Functional diversity of R currents in hippocampal pyramidal neurons. Seminars in the Neurosciences. 579-92, 1993.
Sugarbaker, D.J., Rattan, S. and R.K. Goyal. Mechanical and electrical activity of esophageal smooth muscle during peristalsis. Am. J. Physid 246 (Gastrointest. Liver Physiol. 9):G l45-Gl50, 1984.
Toyama, T., Yokoyama, I. and K. Nishi. Effects of hexamethoniurn and other ganglionic blocking agents on electrical activity of the esophagus induced by vagal stimulation in the dog. Eur. J . Pharmacol. 3 1 :6W 1, 1975.
Van Overbeck, J. J.M., Witt, H.P., Paping, R.H.L. and H.M. Segenhout. Simultaneous manometry and electromyography in the pharyngoesophageal segment. Laryngoscope 95582, 1985.
Welch, R.W., Lukemann, K., Ricks, P.M., Drake, S.T. and G.A. Gates. Manometry of the normal upper esophageal sphincter and its alterations in laryngectomy. J. Clin. Invest. 63(5): 1 036-4 1, 1 979.
Winship, D.H., Deandrade, S.R. and F.F. Zboralske. Infiuence of bolus temperature on huaian esophageal motor f'wiction. J . Clin. ïnvest. 49:243-250, 1970.

Wright, W.N. Nonlïnear models of the electrical properties of neuronal membranes. MSc. Thesis, University of Toronto, 1993.
Yamato, S., Spechier, S.J. and R.K. Goyal. Role of niûic o d e in esophageal peristalsis in the opossum. Gash=oenterology 1 O3 : 1 97-204, 1 992.
Yoshida, Y., Miyazaki, T., Hirano, M., Shin, T., Totoki, T. and T. Kanaselci. Locaüzation of efferent neurons innervating the pharyngeal constrictor muscles and the cervical esophagus muscle in the cat by means of the horseradish peroxidase method. Neurosci Left. 22(2):9 1-5, 198 1.
Zelcer, E. and N.W. Weisbrodt. Electncd and mechanicd activity in the lower esophageal sphincter of the cat. Am. X Physiol. 246:G243-G247, 1984.