reductionism & the modern synthesis “change in allele frequency over time” wright dobzhansky...
TRANSCRIPT

The macroevolutionary dynamics of
adaptive landscapesJosef C UyedaPostdoctoral fellow
iBEST & Dept of BiologyUniversity of Idaho

Reductionism & the Modern Synthesis
“Change in allele frequency over time”
Wright
Dobzhansky
Stebbins Fisher
Simpson
Haldane

Microevolution
Macroevolution

Microevolutionary patternsWe now know that we can study evolution in real time
16.06 g(1976)
17.13 g(1978)
Average weight
Response to selection
6.66% change in body size in 1 generation
(2 years)Conservatively, let’s assume that only a fraction is due to
evolution:i.e. 1% change in body size in
1 generationGrant and Grant, 2002

Let’s do the calculations
• If a population increased by 1% every generation starting from ~16 g:In 200 years
In 500 years
In 1,000 yearsIn 2,000 years
43 g193 g
2.3 kg335 kg
In 10,000 years
6.47 x 1019 kg

Thousands of years

Thousands of years
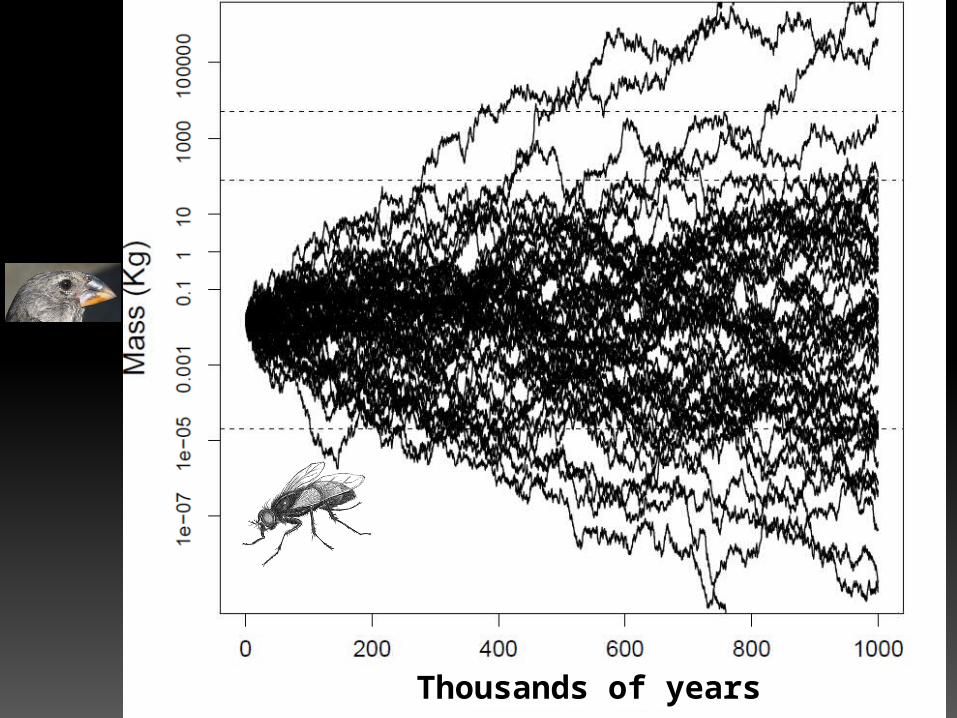
Thousands of years

The Paradox of Stasis (Hansen & Houle 2004):Organisms seem to be able to evolve far more than they do over macroevolutionary scales
Empirical studies often find:• Strong (and often persistent) directional selection
(Hereford et al. 2004, Morrissey & Hadfield 2012)
• High levels of additive genetic variance (Mousseau & Roff 1987, Houle 1992)
• Rapid evolutionary rates(Hendry & Kinnison 1999, Kinnison & Hendry 2002)
…yet stasis in the fossil record (Gingerich 1983, 2002)
…and BM in comparative data?
How do we reconcile patterns at different scales?

All studies of phenotypic evolution measure comparable
quantities
How can we see the pattern across scales of time?
We measure two quantities:
(1) “time for evolution”(2) Δ mean body size
Pop Bmean(z)
Pop Amean(z)
Time interval
Pop X
Pop Bmean(z)
Pop Amean(z)
Time interval

Phenotypic divergence database (Uyeda, Hansen, Arnold & Pienaar, 2011. PNAS.)
Only animals, mostly
vertebrates, but also some inverts
Only traits related to linear body size
change
Field, Fossil and Phylogenetic
comparative data
>8000 data points from> 150 studies
Intervals from < 1 yr to 360 my

Thousands of years

Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

2 years
Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

800,000 years
Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

332.4 myMRCA of all mammals
Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

Average size change for all monotremes vs. all other mammals
Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

Microevolutionary dataFossil dataPhylogenetic comparative data
Uyeda et al., PNAS, 2011

Simpson’s Adaptive Zones

Model fits:Brownian Motion (BM)Multiple-burst (MB) – Poisson Point Process

MB model fits:
Dataset Parameter Estimates
Stasis SD Burst SD Ave. burst time
Whole dataset 𝜎ො��𝑝 = 0.096 𝜎ො��𝑘 = 0.27 25.0 my
Microevolutionary & Fossil 𝜎ො��𝑝 = 0.087 𝜎ො��𝑘 = 0.25 1.5 my
Phylogenetic 𝜎ො��𝑝 = 0.086 𝜎ො��𝑘 = 0.22 21.8 my
AIC = -9142.5 AIC = -9018.0 AIC = -7878.0

Time
Ph
en
oty
pe
Million-year waiting times
“Ephemeral” divergence over short timescales(Futuyma 1987)

Time
Ph
en
oty
pe
Adaptive peak shifts, stabilizing selection & genetic drift
Adaptive Zones/Niches


θ1 θ2 θ3
α
σ2
ln2/α = Phylogenetic Half-Life

Compare models(e.g. by AIC)

The “best” model may still be bad
OU models are not always well-behaved statistically
Problems:
OU models are valuable because of their biological
realism

…but where did the biology go?

Bayesian Reversible-Jump OU Modeling
FlexibleCan infer adaptive shifts and compare to a
priori hypotheses
Can incorporate additional data/realism through informative priors
Customize to test specific hypotheses



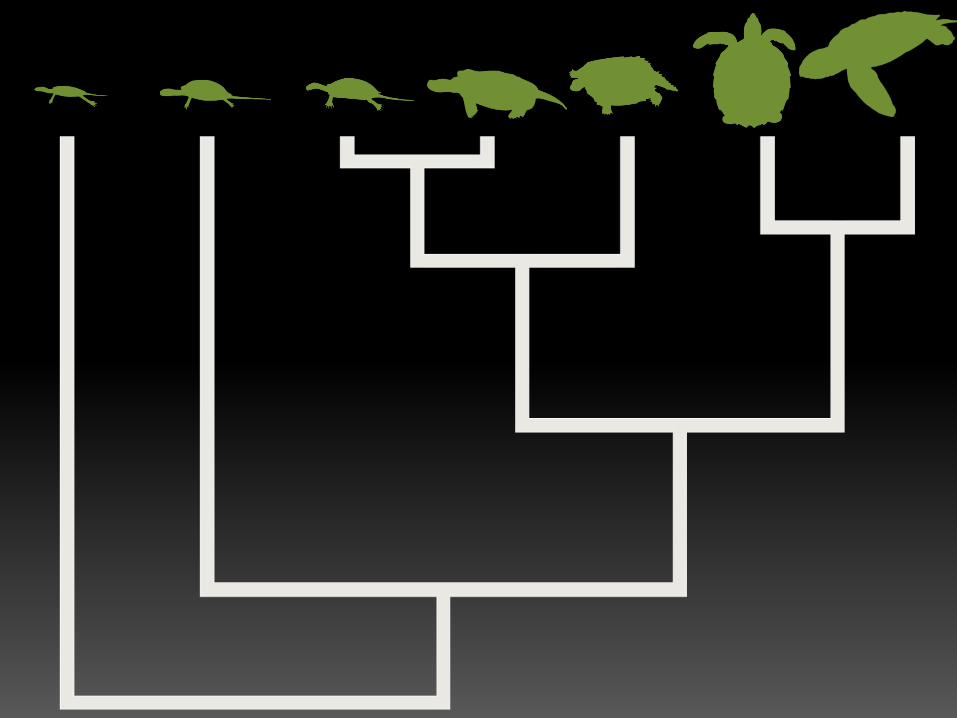
Turtles and Tortoises (Jaffe et al. 2011)

(Uyeda and
Harmon, 2014)
Habitat Modelbayou Model

2 ln BF = 15.24
surface
(Ingram &
Mahler, 2012)
bayou : 8 – 167 cm surface : 2.9x10-170fm– 1.96x1044 km surface:
92.6my
(Uyeda and
Harmon, submitt
ed)
habitatbayou
Habitat Modelbayou Modelsurface Modelbayou Model
𝝈 𝟐 Optima Phylogenetic half-life (my)
prior

We can generate better hypotheses
Jaffe et al. (2011) Marine, Freshwater, Terrestrial and
Island
Only Marine was found by bayou.
Better hypothesis? Aquatic life history + high
environmental temperature

But is it stabilizing selection or adaptive zone shifts or….?
• 85 taxa• Body size (SVL)• Use 3 different
priors:• Weakly
informative (Free)• Blunderbuss
model (Stasis shifts)
• Quantitative genetic model (Peak shifts)
Anolis

Parameterize with the “Blunderbuss”
σ2/(2α)
ln2/α

Quantitative Genetics Model(Lande 1976)
𝑑𝑍 𝑡=−α (𝑍 𝑡−1−θ )𝑑𝑡+σ 𝑑𝑊𝑡
Stabilizing Selection
Genetic Driftα = h2VP/(VP+ω2) σ2 = h2VP/Ne

QG model priors
h2=estimated in anoles to be ~0.55 for body size
VP=estimated from the data
Ne = 99% CI between 1000 and 400,000 (mean 22,000)
ω2= Stabilizing & directional selection gradients estimated from wild populations

QG Model Posteriors
Posterior
Prior

Parameterization Marginal lnL 2 ln BF: Free vs. _____
Free -16.4 0
Blunderbuss -24.9 17.0Lande (QG) -37.7 21.3
Both Blunderbuss and Lande models perform much more poorly than free parameterization
Can also combine these interpretations with ecotype-based hypotheses…..

Regimes Parameterization Convergence Marginal lnLFixed- Ecotype Free Yes 9.83Fixed- Ecotype Blunderbuss Yes 2.73Fixed- Ecotype Free No -3.62Fixed- Ecotype Lande (QG) Yes -6.70Fixed- Ecotype Blunderbuss No -14.16Free Free No -16.41Fixed- Ecotype Lande (QG) No -18.66Free Blunderbuss No -24.93Free Lande (QG) No -37.73
Convergent, Ecotype-based regimes fit tree best, but should not be interpreted as either Blunderbuss or Lande model
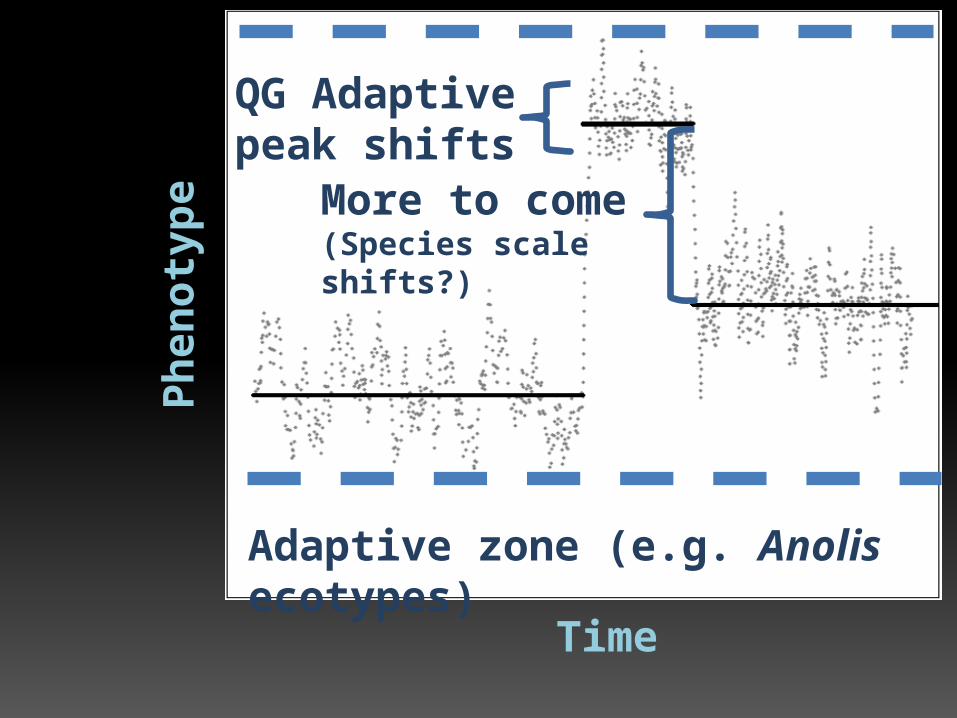
Time
Ph
en
oty
pe
Adaptive zone (e.g. Anolis ecotypes)
QG Adaptive peak shifts
More to come(Species scale shifts?)

A broader framework: Combining fossils, microevolution and
phylogenetic comparative data
Fossil timeseries
Microevolutionary timeseries
Quantitative genetic & selection parameters

Goal: Powerful, customizable phylogenetic comparative methods for testing user-specific biological hypotheses
http://www.arborworkflows.com/

AcknowledgementsFunding and Support
National Science FoundationResearch Council of NorwayBEACON CenteriBEST, U. of IdahoOSU Zoology departmentCEES, U. of Oslo, Norway
Other assistance Phil Gingerich
Andrew HendryLynne HouckPaul JoyceJoe Felsenstein
Coauthors and collaborators
Thomas HansenJon EastmanLuke HarmonStevan J ArnoldJason PienaarMatt Pennell
Aaron ListonMike BlouinDavid LytleHarmon LabHansen LabArnold Lab