reconstructing a puzzle: existence of cyanophages ... · pdf filesan diego state university,...
TRANSCRIPT

Reconstructing a puzzle: existence of cyanophagescontaining both photosystem-I and photosystem-IIgene suites inferred from oceanicmetagenomic datasetsemi_2304 24..32
Ariella Alperovitch-Lavy,1† Itai Sharon,1,2†
Forest Rohwer,3,4 Eva-Mari Aro,5 Fabian Glaser,6
Ron Milo,7 Nathan Nelson8 and Oded Béjà1*1Faculty of Biology, 2Faculty of Computer Science,6Bioinformatics Knowledge Unit, Lorry I. LokeyInterdisciplinary Center for Life Sciences andEngineering, Technion – Israel Institute of Technology,Haifa 32000, Israel.3Department of Biology, 4Center for Microbial SciencesSan Diego State University, San Diego, CA 92182,USA.5Department of Biochemistry and Food Chemistry, PlantPhysiology and Molecular Biology, University of Turku,FI-20014 Turku, Finland.7Department of Plant Sciences, Weizmann Institute ofScience, Rehovot 76100, Israel.8Department of Biochemistry, George S. Wise Faculty ofLife Sciences, Daniella Rich Institute for StructuralBiology, Tel Aviv University, Tel Aviv 69978, Israel.
Summary
Cyanobacteria play a key role in marine photosynthe-sis, which contributes to the global carbon cycle andto the world oxygen supply. Genes encoding thephotosystem-II (PSII) reaction centre are found inmany cyanophage genomes, and it was suggestedthat the horizontal transfer of these genes might beinvolved in increasing phage fitness. Recently,evidence for the existence of phages carryingPhotosystem-I (PSI) genes was also reported. Here,using a combination of different marine metagenomicdatasets and a unique crossing of the datasets, wenow describe the finding of phages that, as in plantsand cyanobacteria, contain both PSII and PSI genes.In addition, these phages also contain NADH dehy-drogenase genes. The presence of modified PSII andPSI genes in the same viral entities in combination
with electron transfer proteins like NAD(P)H dehydro-genase (NDH-1) strongly points to a role in perturba-tion of the cyanobacterial host photosyntheticelectron flow. We therefore suggest that, dependingon the physiological condition of the infected cyano-bacterial host, the viruses may use different optionsto maximize survival. The modified PSI may alternatebetween functioning with PSII in linear electron trans-fer and contributing to the production of both NADPHand ATP or functioning independently of PSII in cyclicmode via the NDH-1 complex and thus producingonly ATP.
Introduction
Several marine cyanophages from the myovirus andpodovirus families carry photosynthetic genes, and it wassuggested that these genes increase phage fitness (Mannet al., 2003; Lindell et al., 2004; 2005; Millard et al., 2004;Sullivan et al., 2005; Bragg and Chisholm, 2008; Dam-meyer et al., 2008). Cyanobacterial photosynthetic mem-branes contain two photosystems, of which PSII mediatesthe transfer of electrons from water, the initial electrondonor, to the plastoquinone pool whereas PSI mediateselectron transfer from plastocyanin to ferredoxin, therebygenerating reducing power needed for CO2 fixation in theform of NADPH. While PSII is known to be sensitive tophotodamage, PSI is considered to be more stable thanPSII.
The PSII gene psbA coding for the labile D1 protein isreadily detected in various cultured and environmentalmyoviruses and podoviruses infecting Prochlorococcusand marine Synechococcus (Mann et al., 2003; Lindellet al., 2004; Zeidner et al., 2005; Sullivan et al., 2006;Sharon et al., 2007; Weigele et al., 2007; Millard et al.,2009). In myoviruses, genes encoding the PSII D2 protein(Mann et al., 2003; Lindell et al., 2004; Millard et al., 2004;Sullivan et al., 2005; 2006) are also found in the vicinity ofthe psbA gene. Recently, PSI gene cassettes containingwhole gene suites (psaJF, C, A, B, K, E and D) sufficientto build a monomeric PSI were reported to exist in marinecyanophages from the Pacific and Indian Oceans (Sharonet al., 2009). This was observed using both the Global
Received 11 January, 2010; accepted 8 June, 2010. *For corre-spondence. E-mail [email protected]; Tel. (+972) 4 829 3961;Fax (+972) 4 822 5153. †These authors contributed equally to thiswork.
Environmental Microbiology (2011) 13(1), 24–32 doi:10.1111/j.1462-2920.2010.02304.x
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd
Ocean Sampling (GOS) phase-I (GOS-I) dataset (Ruschet al., 2007) and viral marine biomes (Dinsdale et al.,2008a,b). The presence of modified PSI genes in phageswas hypothesized to be connected to alternative electrontransfer pathways and alternate energy acquisition(Sharon et al., 2009).
Abundance measures performed with Pacific Oceansamples (Sharon et al., 2009) suggested that some sitesmight contain phages that do carry both photosystems.
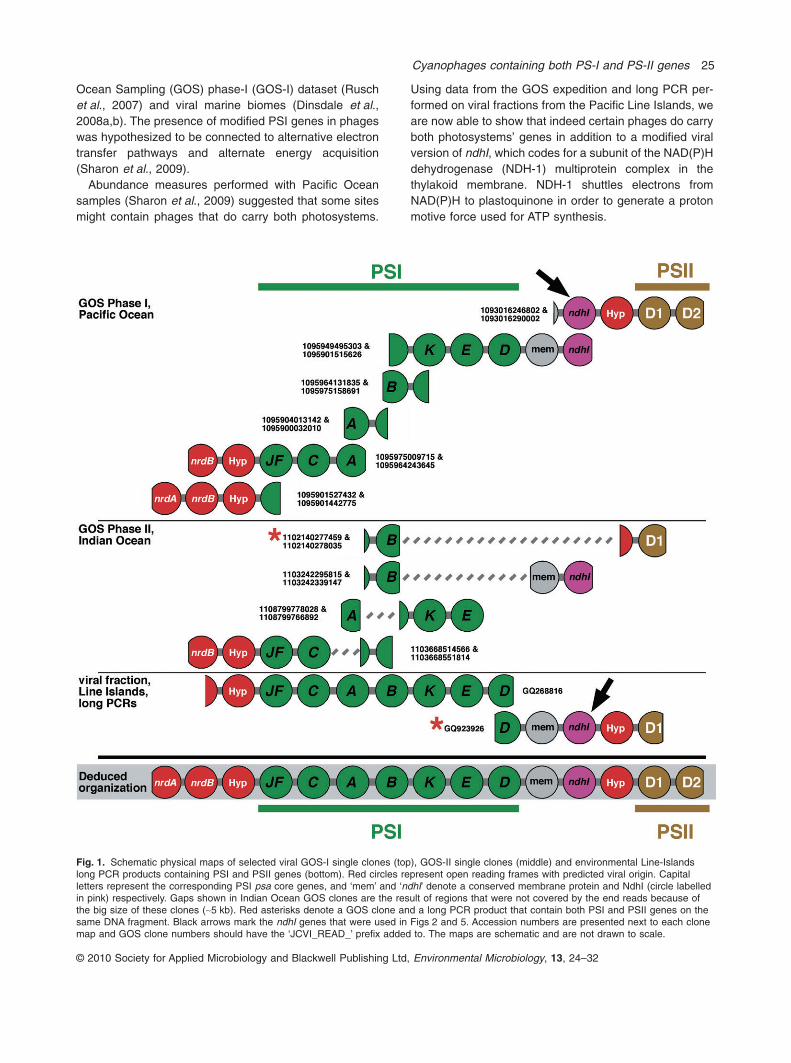
Using data from the GOS expedition and long PCR per-formed on viral fractions from the Pacific Line Islands, weare now able to show that indeed certain phages do carryboth photosystems’ genes in addition to a modified viralversion of ndhI, which codes for a subunit of the NAD(P)Hdehydrogenase (NDH-1) multiprotein complex in thethylakoid membrane. NDH-1 shuttles electrons fromNAD(P)H to plastoquinone in order to generate a protonmotive force used for ATP synthesis.
Fig. 1. Schematic physical maps of selected viral GOS-I single clones (top), GOS-II single clones (middle) and environmental Line-Islandslong PCR products containing PSI and PSII genes (bottom). Red circles represent open reading frames with predicted viral origin. Capitalletters represent the corresponding PSI psa core genes, and ‘mem’ and ‘ndhI’ denote a conserved membrane protein and NdhI (circle labelledin pink) respectively. Gaps shown in Indian Ocean GOS clones are the result of regions that were not covered by the end reads because ofthe big size of these clones (~5 kb). Red asterisks denote a GOS clone and a long PCR product that contain both PSI and PSII genes on thesame DNA fragment. Black arrows mark the ndhI genes that were used in Figs 2 and 5. Accession numbers are presented next to each clonemap and GOS clone numbers should have the ‘JCVI_READ_’ prefix added to. The maps are schematic and are not drawn to scale.
Cyanophages containing both PS-I and PS-II genes 25
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

Results and discussion
Previous abundance measures (Sharon et al., 2009, seethe methods section for a brief description) performedwith Line Islands samples have hinted that in some LineIslands sites [Tabuarean (Fanning Atoll) and Kiritimati(Christmas Island)] all cyanophages contain PSII geneswhile 50% to 60% contain PSI genes. Potentially thismeans that 50% to 60% of cyanophages in these sitescontain both PSII and PSI genes, as do plants and cyano-bacteria (see table S3 in Sharon et al., 2009).
An in silico search for PSII- and PSI-containing phageentities was therefore conducted on the publicly available(http://camera.calit2.net/) GOS phase-II dataset from theIndian Ocean gyre (GOS-II). As opposed to the earlierGOS datasets that comprised small DNA inserts with anaverage size of < 2.0 kb (Rusch et al., 2007), the GOS-IIdataset was prepared using clones with inserts of ~5.0 to6.0 kb. The use of a bigger insert size in the GOS-IIlibraries enabled us to detect and link both photosystemgenes on one clone (i.e. PSI genes psaA and psaB andPSII gene psbA; see Indian Ocean GOS-II clone labelled
Fig. 2. FastTree approximated maximum-likelihood phylogenetic tree of NdhI proteins from the GOS project. Cyan branches denoteProchlorococcus proteins, blue denotes Synechococcus, red denotes predicted viral origin based on neighbour proteins on the scaffold, andarrows denotes the NdhI proteins shown in Fig. 1. Sequence 1096627172387 represents the sequence of JCVI_SCAF_1096627172387 builtby reads JCVI_READ_1093016246802 and 1093016290002 (top Fig. 1). The tree is based on 198 amino acid positions. Local bootstrapvalues are shown next to the connecting nodes.
26 A. Alperovitch-Lavy et al.
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

with a red asterisk in the middle panel of Fig. 1). As theGOS general scaffold assembly represents reads thatcome from different GOS sites or from different clonesand hence by definition are chimeric, all analyses wererestricted to sequences assembled from single clonereads only. Our search revealed a GOS-II clone that con-tains the PSI genes psaA and psaB and the PSII genepsbA. We also found GOS-I clones that contain eitherthe PSI genes psaBKED, a hypothetical membraneprotein and a viral NAD(P)H dehydrogenase gene (ndhI),a viral hypothetical gene, or the psbAD genes (see twoupper clones in Fig. 1). These findings strongly pointstowards the existence of phage genomes containinggenes of both photosystems separated by a gene codingfor a crucial subunit, NdhI, of the thylakoid NDH-1complex.
To further validate the connection of PSI and PSIIgenes on the viral scaffolds, long PCR reaction was per-formed on viral Line Islands DNA, using a forward primerfrom the end of the PSI gene cassette (psaD) and a
reverse primer from the beginning of the PSII gene psbA.Sequence of positive amplicons confirmed the connectionof the PSI and PSII genes on the same viral fragment (seefragment GQ923926 in Fig. 1). Sequencing of the regionbetween the photosynthetic genes revealed an organiza-tion already observed in GOS-I viral clones where threepredicted proteins are found in this space (right side of toppanel in Fig. 1). A predicted membrane protein found inboth Synechococcus (for example, gene BL107-14195in Synechococcus sp. BL107) and Prochlorococcus(for example, protein P9515-15211 in Prochlorococcusmarinus str. MIT 9515), a modified NAD(P)H dehydroge-nase subunit I (NdhI, a subunit of the NDH-1 multiproteincomplex of cyanobacterial thylakoid membrane), and apredicted hypothetical viral protein similar to hypotheticalprotein PSSM2-282 observed in the genome of Prochlo-rococcus myophage P-SSM2 (Lindell et al., 2004; Sulli-van et al., 2005). To the best of our knowledge, anNAD(P)H dehydrogenase I gene has not previously beendetected in phage genomes. Phylogenetic trees built with
Fig. 3. FastTree approximated maximum-likelihood phylogenetic tree of PSI PsaD proteins from the GOS project. Cyan branches denoteProchlorococcus proteins, blue denotes Synechococcus, and red denotes predicted viral origin based on neighbour proteins on the scaffold.Local bootstrap values are shown.
Cyanophages containing both PS-I and PS-II genes 27
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

Fig. 4. FastTree approximated maximum-likelihood phylogenetic tree of PSI PsaB proteins from the GOS project. Cyan branches denoteProchlorococcus proteins, blue denotes Synechococcus, and red denotes predicted viral origin based on neighbour proteins on the scaffold.Local bootstrap values are shown.
28 A. Alperovitch-Lavy et al.
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

the suspected viral NdhI proteins (marked with an arrow inFig. 2) revealed numerous GOS viral-suspected (basedon neighbouring genes on the scaffolds) peptides thatcluster together (labelled red in Fig. 2) and separatelyfrom their cyanobacterial counterparts, further supportinga viral origin (a similar trend is observed with trees con-structed with the viral PSI proteins PsaD and PsaB (Figs 3and 4)). This separation into two distinguished popula-tions, one from bacteria (cyanobacteria) and one fromviruses (phages) is also observed when coverage mea-sures (Fig. 5) to all different GOS ndhI genes are per-formed against 454 pyrosequencing generated marineviromes and microbiomes (Dinsdale et al., 2008a,b). Thisalso increased the credibility of our search as the twodifferent approaches (shotgun sequencing vs. pyrose-quencing) each introduce completely different bias noisessome of which are the result of differential DNA extractiontechniques, the 454’s lack of cloning bias, the ability tosequence through regions of the genome that exhibitstrong secondary structures or problems with resolvingrepeat regions [for discussion on pros and cons ofshotgun sequencing and 454 sequencing approaches,see Goldberg and colleagues (2006) and Harismendy andcolleagues (2009)].
In an attempt to try and find the potential host for theviruses that carry the PSI and PSII genes, a tree based onthe PSII D1 protein (PsbA) was constructed (Fig. 6). The
D1 protein from the long PCR product (that contain bothPSI and PSII genes) falls, with high bootstrap values, nextto the D1 protein from cyanophage P-SSM4 [whose hostscould be low-light or high-light Prochlorococcus strains(Sullivan et al., 2003)] and with D1s from differentProchlorococcus strains. We therefore suggest that thepotential host for the cyanophages, detected in our study,that contain both PSI and PSII are Prochlorococcus cells.
The thylakoid membranes of cyanobacteria harbour atleast two types of NDH-1 complexes: NDH-1L beinginvolved in cyclic electron flow around PSI and in respi-ration and the NDH-1MS being involved in CO2 uptake(Battchikova and Aro, 2007). Genome-wide analysis pre-dicts the presence of still different NDH-1 complexes butsuch complexes have never been identified at proteinlevel in cyanobacterial membranes. The soluble NdhIprotein, which is encoded by the viral ndhI gene identifiedhere, is found in both NDH-1L and NDH-1MS complexesand it is predicted to be present in the so far ‘undiscov-ered’ NDH-1 complexes as well (Battchikova and Aro,2007).
We recently proposed that the elimination of the specificplastocyanin binding site by the presence of PsaJF fusionprotein in the virus encoded PSI might lead to electronflow from the respiratory chain to PSI instead of cyto-chrome oxidase (Sharon et al., 2009). The presence ofmodified PSII and PSI genes in the viral entities in com-bination with electron transfer proteins like NdhI stronglypoints to a role in perturbation of the cyanobacterial hostphotosynthetic and respiratory electron flow. The conse-quence of this ensemble in the infected cyanobacteriawould be a flexibility to produce NAD(P)H and ATP ingreatly non-stoichiometric amounts, which might bettercorrespond to the needs of virus replication.
Does a virus gain a selective advantage from augment-ing the activity of PSI in parallel to PSII? A schematicenergy balance of the infection process lends someinsights. Myoviruses are about 100 nm (Weigele et al.,2007) in diameter and their burst size, although variable,is in the range of 20–80 (Wilson et al., 1996). For a char-acteristic protein radius of 4 nm (Phillips et al., 2009), andassuming a crystal like viral protein content, a productionof about 50*(100/4)3 (i.e. ~106) proteins is required in theshort time span of the viral lytic process. This is close tothe overall protein content of a 700 nm diameter Prochlo-rococcus cell (Partensky et al., 1999). Given that aminoacid polymerization into proteins is estimated to be adominant factor in the energetic load of building a new cell(Stouthamer, 1973), and that the process of infection wasobserved to be shorter than a cell cycle, we can predictthat the viral protein production energetic cost is a loadthat could limit the burst size achieved or equivalently thetime for completion of a lytic cycle once favourable con-ditions are encountered. This suggests a strong selective
Fig. 5. Recruitment coverage of GOS single clones carrying ndhIgenes with Northern Line Islands biomes [viromes (x-axis) andmicrobiomes (y-axis)]. Coverage is defined as % of GOS clonelength covered by at least one recruited read from the Line Islandsbiomes. Colours used are same as in Fig. 2. Viral/non-viralclassification assignments of the different GOS clones was basedon neighbouring genes to the ndhI gene.
Cyanophages containing both PS-I and PS-II genes 29
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

pressure for augmenting the energetics towards PSI-dependent ATP production.
We therefore speculate that depending on the physi-ological condition of the infected cyanobacterial host, theviruses may use different options to maximize survival.The modified PSI may alternate between working withPSII, working independently using other electron sources(e.g. organic material) or alternatively working in a cyclicphotophosphorylation mode.
Experimental procedures
PCR amplification of fragments connecting PSIand PSII genes
For amplifying the region linking the PSI cassette and thePSII genes, genomic DNA from viral samples from the North-ern Line Islands [‘Line Islands Christmas Reef B3 phage’(Dinsdale et al., 2008a)] was used. The PCR reaction
was preformed using TaKaRa LA Taq™ DNA polymerase(TaKaRa Bio INC, Japan). Polymerase chain reaction ampli-fication was carried out in a total volume of 50 ml containing50 ng of Northern Line Islands phage DNA, 400 mM dNTPs,2.5 mM MgCl2, 1 mM primers [psaDfwd (5′-TAYCCNAARGAYGGNGTNG-3′) and psbArev (5′-GCYTCCCADATNGGRTA-3′)] and 2.5 U TaKaRa LA Taq™ DNA polymerase. Theamplification conditions comprised steps at 95°C for2 min, 40 cycles of 95°C for 30 s, 50°C for 30 s and 68°Cfor 3 min.
Determining PSI and PSII gene abundancein cyanophages
The complete procedure is described in the supporting infor-mation for Sharon and colleagues (2009). In short, theobserved number of reads containing selected genes fromPSI (psaA and psaB) and PSII (psbA and psbD) was normal-ized by the number of reads containing selected genes foundacross all known myoviruses (gp20 and gp23) and podovi-
Fig. 6. FastTree approximated maximum-likelihood phylogenetic tree of PSII PsbA proteins from the GOS project. Cyan branches denoteProchlorococcus proteins, blue denotes Synechococcus, and red denotes predicted viral origin based on neighbour proteins on the scaffold.Local bootstrap values are shown next to the connecting nodes.
30 A. Alperovitch-Lavy et al.
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

ruses (DNAPol), taking gene length into consideration. Theresulting figures may serve as estimations for the abundanceof each gene across the cyanophages in the examinedsamples.
Recruitment coverage analysis
Procedure and parameters used for this analysis (presentedin Fig. 5) were the same as in Sharon and colleagues (2009).For each recruiting clone, reads from the four viral and fourmicrobial Northern Line Islands samples were recruited usingBLAST with parameters (-p blastn -e 1e-3 -F F -r 2). Only readsaligning with at least 85% identity for 80 bp or more of theirlength were considered (average read length in the LineIslands samples is ~100 bp). Coverage is defined as thefraction of the recruiting sequence covered by at least onerecruited read.
Phylogenetic tree reconstruction
Phylogenetic trees were constructed by the FastTree pro-gramme (Price et al., 2010), using multiple alignments pro-duced by MUSCLE (Edgar, 2004). While we used MUSCLEdefault parameters, FastTree was run using parameters forhigh accuracy: -spr 4 (to increase the number of rounds ofminimum-evolution SPR moves) and -mlacc 2 -slownni (tomake the maximum-likelihood NNIs search more exhaus-tive). These parameters can produce slight increases in accu-racy. To estimate the reliability of each split in the tree,FastTree uses the Shimodaira-Hasegawa test on the threealternate topologies (NNIs) around that split (Guindon et al.,2009).
Acknowledgements
We would like to thank D. Lindell for encouragement andideas. This work was supported in part by grants 1203/06(O.B.) and 356/06 (N.N.) from the Israel Science Foundation.The long PCR product sequence reported in this study thatcontain both PSI and PSII genes and the region betweenis deposited with GenBank under Accession numberGQ923926.
References
Battchikova, N., and Aro, E.M. (2007) Cyanobacterial NDH-1complexes: multiplicity in function and subunit composition.Physiol Plant 131: 22–32.
Bragg, J.G., and Chisholm, S.W. (2008) Modeling the fitnessconsequences of a cyanophage-encoded photosynthesisgene. PLoS ONE 3: e3550.
Dammeyer, T., Bagby, S.C., Sullivan, M.B., Chisholm, S.W.,and Frankenberg-Dinkel, N. (2008) Efficient phage-mediated pigment biosynthesis in oceanic cyanobacteria.Curr Biol 18: 442–448.
Dinsdale, E.A., Pantos, O., Smriga, S., Edwards, R.A., Angly,F., Wegley, L., et al. (2008a) Microbial ecology of fourcoral atolls in the northern Line Islands. PLoS ONE 3:e1584.
Dinsdale, E.A., Edwards, R.A., Hall, D., Angly, F., Breitbart,M., Brulc, J.M., et al. (2008b) Functional metagenomic pro-filing of nine biomes. Nature 452: 629–632.
Edgar, R.C. (2004) MUSCLE: multiple sequence alignmentwith high accuracy and high throughput. Nucleic Acids Res32: 1792–1797.
Goldberg, S.M., Johnson, J., Busam, D., Feldblyum, T.,Ferriera, S., Friedman, R., et al. (2006) A Sanger/pyrosequencing hybrid approach for the generation ofhigh-quality draft assemblies of marine microbial genomes.Proc Natl Acad Sci USA 103: 11240–11245.
Guindon, S., Delsuc, F., Dufayard, J.F., and Gascuel, O.(2009) Estimating maximum likelihood phylogenies withPhyML. Methods Mol Biol 537: 113–137.
Harismendy, O., Ng, P.C., Strausberg, R.L., Wang, X., Stock-well, T.B., Beeson, K.Y., et al. (2009) Evaluation of nextgeneration sequencing platforms for population targetedsequencing studies. Genome Biol 10: R32.
Lindell, D., Sullivan, M.B., Johnson, Z.I., Tolonen, A.C.,Rohwer, F., and Chisholm, S.W. (2004) Transferof photosynthesis genes to and from Prochloro-coccus viruses. Proc Natl Acad Sci USA 101: 11013–11018.
Lindell, D., Jaffe, J.D., Johnson, Z.I., Church, G.M., andChisholm, S.W. (2005) Photosynthesis genes in marineviruses yield proteins during host infection. Nature 438:86–89.
Mann, N.H., Cook, A., Millard, A., Bailey, S., and Clokie, M.(2003) Bacterial photosynthesis genes in a virus. Nature424: 741.
Millard, A., Clokie, M.R.J., Shub, D.A., and Mann, N.H.(2004) Genetic organization of the psbAD region in phagesinfecting marine Synechococcus strains. Proc Natl AcadSci USA 101: 11007–11012.
Millard, A.D., Zwirglmaier, K., Downey, M.J., Mann, N.H., andScanlan, D.J. (2009) Comparative genomics of marinecyanomyoviruses reveals the widespread occurrence ofSynechococcus host genes localised to a hyperplasticregion: implications for mechanisms of cyanophage evolu-tion. Environ Microbiol 11: 2370–2387.
Partensky, F., Hess, W.R., and Vaulot, D. (1999) Prochloro-coccus, a marine photosynthetic prokaryote of global sig-nificance. Microbiol Mol Biol Rev 63: 106–127.
Phillips, R., Kondev, J., and Theriot, J. (2009) PhysicalBiology of the Cell. London, UK: Taylor & Francis.
Price, M.N., Dehal, P.S., and Arkin, A.P. (2010) FastTree 2 –approximately maximum-likelihood trees for large align-ments. PLoS ONE 5: e9490.
Rusch, D.B., Halpern, A.L., Heidelberg, K.B., Sutton, G.,Williamson, S.J., Yooseph, S., et al. (2007) The sorcererII global ocean sampling expedition: I, the northwestAtlantic through the eastern tropical Pacific. PLoS Biol 5:e77.
Sharon, I., Tzahor, S., Williamson, S., Shmoish, M., Man-Aharonovich, D., Rusch, D.B., et al. (2007) Viral photosyn-thetic reaction center genes and transcripts in the marineenvironment. ISME J 1: 492–501.
Sharon, I., Alperovitch, A., Rohwer, F., Haynes, M., Glaser,F., Atamna-Ismaeel, N., et al. (2009) Photosystem-I genecassettes are present in marine virus genomes. Nature461: 258–262.
Cyanophages containing both PS-I and PS-II genes 31
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32

Stouthamer, A.H. (1973) A theoretical study on the amount ofATP required for synthesis of microbial cell material.Antonie Van Leeuwenhoek 39: 545–565.
Sullivan, M.B., Waterbury, J.B., and Chisholm, S.W. (2003)Cyanophages infecting the oceanic cyanobacteriumProchlorococcus. Nature 424: 1047–1051.
Sullivan, M.B., Coleman, M.L., Weigele, P., Rohwer, F., andChisholm, S.W. (2005) Three Prochlorococcus cyanoph-age genomes: signature features and ecological interpre-tations. PLoS Biol 3: e144.
Sullivan, M.B., Lindell, D., Lee, J.A., Thompson, L.R., Bielaw-ski, J.P., and Chisholm, S.W. (2006) Prevalence andevolution of core photosystem II genes in marine cyano-bacterial viruses and their hosts. PLoS Biol 4: e234.
Weigele, P.R., Pope, W.H., Pedulla, M.L., Houtz, J.M., Smith,A.L., Conway, J.F., et al. (2007) Genomic and structuralanalysis of Syn9, a cyanophage infecting marine Prochlo-rococcus and Synechococcus. Environ Microbiol 9: 1675–1695.
Wilson, W.H., Carr, N.G., and Mann, N.H. (1996) The effectof phosphate status on the kinetics of cyanophage infectionof the oceanic cyanobacterium Synechococcus sp.WH7803. J Phycol 32: 506–516.
Zeidner, G., Bielawski, J.P., Shmoish, M., Scanlan, D.J.,Sabehi, G., and Béjà, O. (2005) Potential photosynthesisgene recombination between Prochlorococcus & Synecho-coccus via viral intermediates. Environ Microbiol 7: 1505–1513.
32 A. Alperovitch-Lavy et al.
© 2010 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 13, 24–32