reconstitution of phosphorylating electron transport in mitochondria from a cytochrome c
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEWWRY Vol. 241, No. 19, Issue of October 10, PP. 4330-4338, 1966
Printed in U.S.A.
Reconstitution of Phosphorylating Electron Transport in Mitochondria from a Cytochrome c-deficient Yeast Mutant*
(Received for publication, March 2, 1966)
JAMES R. MATTOON AND FRED SHERMAN
From the Department of Physiological Chemistry, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, and the Department of Radiation Biology and Biophysics, University of Rochester School oj Medicine and Dentistry, Rochester, New York 14620
SUMMARY
Electron transport and oxidative phosphorylation in mito- chondria isolated from a cytochrome c-deficient mutant of the yeast, Saccharomyces cerevisiae, may be efficiently re- constituted by addition of purified yeast or horse heart cyto- chrome c. Added cytochrome c is tightly bound and is retained during reisolation of mitochondria.
Reconstituted mutant mitochondria carry out oxidative phosphorylation with substrates linked to nicotinamide adenine dinucleotide, flavoprotein, and cytochrome c as efficiently as mitochondria isolated from wild type yeast under the same conditions.
Titration of mutant mitochondria with yeast cytochrome c leads to progressive increases in P:O ratios as well as to increases in rates of phosphate and oxygen uptake. The end points of such titrations indicate that reconstitution is stoichi- ometric at a ratio of 0.2 mpmole of cytochrome c per mg of mitochondrial protein, approximately the amount of cyto- chrome c in normal wild type mitochondria.
Respiration in mutant mitochondria can be restored equally well by two types of yeast cytochrome c (iso-l- and iso-2-cytochrome c) and by horse heart cytochrome c. The two types of yeast cytochrome c are virtually equivalent in restoring phosphorylation coupled to succinate oxidation, whereas the horse heart enzyme is less effective.
Mutant mitochondria reconstituted with iso-1-cytochrome c catalyze phosphorylating oxidation of succinate, ar-keto- glutarate, glutathione plus tetramethyl-j-phenylenediamine, pyruvate plus malate, and D-lactate plus malate with effi- ciency comparable to that of normal yeast mitochondria. Ethanol oxidation was partially restored. Oxidative phos- phorylation with L-lactate, even in the presence of added malate to provide citric acid cycle intermediates, was poorly restored by either of the isocytochromes c.
* This investigation was supported in part by the United States Atomic Energy Commission at The University of Rochester Atomic Energy Project, Rochester, New York, and in part by Public? Health Research Grants GM 12702. HD 10677, and GM 05919 from the National Institutes of Health’and by Pudlic Health Service Special Research Fellowship l-F3 GM 16,634-0181.
In the past, the reconstitution of electron transport and oxida- tive phosphorylation has been limited to the use of mitochondrial fragments or mitochondria which were depleted of one or more components by nonspecific chemical or mechanical treatments. This investigation concerns the use of mitochondria isolated from a cytochrome c-deficient mutant of yeast. These mito- chondria were shown to be highly specific “acceptors” for various kinds of cytochrome c and had a greater degree of integrity compared to acceptor mitochondria obtained by other methods.
Normal wild type bakers’ yeast contains two distinct molecular types of cytochrome c which are referred to as iso-1-cytochrome c and iso-2cytochrome c (1, 2). In many aerobically grown strains of Saccharomyces cerevisiae, iso-1-cytochrome c con- stitutes a major fraction while iso-2-cytochrome c comprises only about 5% of the total cytochrome c. However, single gene mutants (cyl) contain exclusively iso-2-cytochrome c at concentrations roughly equal to those found in wild type strains (1). Although intact cells of cyl strains contain only 5 to 10% of the normal amounts of total cytochrome c, they still have the capacity for some respiration (I), and exhibit approximately normal amounts of various other mitochondrial enzymes (3). However, cyl mutants cannot utilize nn-lactate for growth, although they can utilize a variety of other nonfermentable substrates (1, 2).
The cyl mutant was chosen for this initial study since cyto- chrome c is soluble, stable, and readily obtained in highly purified form from different species. Also, cytochrome c can be reversibly removed from mammalian mitochondria by treatment with hypotonic KC1 without excessive destruction of phosphorylative capacity (4), and it therefore seemed probable that a cytochrome c deficiency would not cause significant secondary alterations in yeast mitochondria. In addition, the mutant character is sufficiently stable to allow production of large quantities of homogeneous cells, thereby permitting isolation of adequate quantities of mitochondria.
In this investigation, we have shown that both electron trans- port and oxidative phosphorylation could be restored in cyl mitochondria by simple additions of purified cytochromes c. Comparisons have been made of the relative effectiveness of horse heart, iso-1-, and iso-2-cytochromes c.
4330
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of October 10, 1966 J. R. Mattoon and F. Sherman 4331
MATERIALS AND METHODS
Yeast Strains
The two diploid strains of Saccharomyces cerevisiae employed in this investigation were the cytochrome c-deficient mutant, D-247, and the normal strain, D-261. The strain D-247 which is homozygous for the cylbl gene (1, 2), contains exclusively iso-2-cytochrome c at concentrations of 5 to 10% of the total cytochrome c found in normal strains. D-247 is especially stable both with respect to back-mutations and cytoplasmic mutations (p-) .
Growth Media
The media designated as “natural” were comprised of 1% Fisher yeast extract, 2% Fisher peptone, and various concentra- tions of different carbon sources. The pH of all natural media was 4.5. Natural media were used for all experiments except in testing for back-mutants. Synthetic media, used only for testing for the presence of back-mutants in cultures of D-247, contained one-tenth the concentration of Medium B of Vavra and Johnson (5) except for the carbon source and MgS04.7Hz0, which was added at 183 mg per liter. The carbon sources were either 2% glucose (pH 4.5) or 2 ml of 85% lactic acid per 100 ml of medium (pH 6.5). Two per cent agar was included for solid medium.
Culture Methods
Yeast strains were grown at 30” in 8 to 11-liter batches in 14-liter fermentor jars on a New Brunswick Microferm model MF 314 fermentor drive assembly. Effective aeration was assured by bubbling air through the medium at 14 liters per min by means of a sparger; the impeller was operated at 650 to 700 rpm. Samples were withdrawn at intervals for dry weight, Q 022 and pH determinations. Growth periods varied from 16 to 27 hours depending on the strain of yeast and the medium used. Inoculum was grown in two 2-liter Erlenmeyer flasks containing 300 ml of 2 y0 glucose medium per flask. Flasks were inoculated with about 2 to 4 mg of cells (dry weight) and incu- bated for 24 hours at 30” on a New Brunswick model G-10 Gyrorotary shaker operated at 250 rpm. The entire contents of the 2 flasks, amounting to 2.5 to 3.0 g (dry weight) of mutant cells or 3 to 4 g of normal cells, was used to inoculate a fermentor jar. As a test for back-mutants, samples of cells from the fermentor were streaked on plates of solid synthetic medium containing either lactate or glucose as the sole carbon sources. Strain D-247 gives no detectable growth on m-lactate over a 3- to 4-day incubation period. Cells were collected by centrifu- gation in a Westphalia model LWA laboratory separator and washed twice with distilled water. In most cases, mitochondria from normal yeast were prepared within a few hours after the end of the fermentation, but occasionally harvested cells were stored overnight at 4” before use. Longer periods of storage caused some decline in mitochondrial activity. Mutant yeast cells were stored from 12 to 70 hours before use. No adverse effects of this storage on mutant mitochondria could be detected.
Determination of Yeast Respiratory Capacity
Oxygen uptake of intact yeast cells was determined with a Clark polarographic electrode connected to a Varian G-10 re-
corder which was equipped with a 0.6-volt power supply, as outlined by Kielley (6). The electrode was inserted into a cuvette (15 X 45 mm) mounted in a 25” water bath. The medium (1.8 ml), containing 200 pmoles of glucose and 30 Mmoles of inorganic phosphate and having a pH of 4.5, was stirred with a Teflon-coated magnetic stirring bar. A small volume (0.2 or 0.1 ml) of an aqueous suspension of washed cells was injected into the medium, and the rate of 02 uptake was determined from the slope of the recorder trace. Dry weight was estimated turbidimetrically at 660 rnp after appropriate dilution of an aliquot of the suspension. It was found’ that the most reproduci- ble QoZ values could be obtained when the rate of oxygen uptake was between 150 and 300 mpatoms per min.
Preparation of Cytochrome c
Yeast iso-1-cytochrome c was prepared from commercial Fleischmann’s bakers’ yeast (Standard Brands). Iso-2-cyto- chrome c was prepared either from the mutant strain D-247 or from Fleischmann’s yeast. The iso-1-cytochromes c obtained from Fleischmann’s yeast and various laboratory strains were shown to be identical by amino acid analyses and peptide maps.z Also the iso-2-cytochromes c from strain D-247 and Fleisch- mann’s yeast were indistinguishable. The procedures for isola- tion and purification of the yeast isocytochromes c have been presented in detail elsewhere3 and are summarized as follows. The cytochrome c was extracted from yeast with ethyl acetate and NaCl and absorbed on coarse cation exchange resin accord- ing to procedures previously described (1). The cytochrome c was eluted with 0.8 M NaCl in 0.1 M sodium phosphate buffer (pH 7.2) and some impurities were precipitated by the addition of (NH,)&04 to 80% saturation. Final purifications and separations of the two isocytochromes c were performed on cation exchange columns by using a linear gradient of NaCl (1).
Horse heart cytochrome e (Type III) was obtained from Sigma and further purified by ion exchange chromatography.
Preparation of Yeast Mitochondria
Method A-A packed pellet of about 40 g (wet weight) of washed yeast cells was suspended in about 60 ml of a medium, designated MTEB, of the composition 0.25 M mannitol, 0.02 M
Tris, 10-s M EDTA, and 0.2% crystalline bovine serum albumin (Armour), at pH 7.4. The cells were then collected by centrifu- gation, and the packed pellet was suspended in fresh Medium MTEB to give a final volume of 150 to 200 ml. Fifty milliliters (70 g) of 0.2-mm diy glass beads (No. 4285-M20, Arthur H. Thomas) were added to the suspension, and the pH was adjusted to 7.4. (The glass beads were previously cleaned by soaking in aqua regia for about 24 hours followed by very thorough wash- ing with distilled water of high purity.) The suspension was chilled to 0” and homogenized, with recycling, in an Eppenbach MV-6-3 Micro-mill (Gifford-Wood) cooled by circulation of ethanol at -5” through the jacket and coils of the hopper.
The mill was precooled for 3 to 5 min by operating at low speed at a gap setting of 60 (gap settings represent thousandths of an inch clearance between mill rotor and stator) with about 200
1 C. R. Alving and J. R. Mattoon, unpublished experiments. 2 J. Stewart, H. Taber. F. Sherman, E. Margoliash. and W.
Campbell, unpublished experiments. - 3 F. Sherman, J. Parker, J. Stewart, E. Margoliash, and W.
Campbell, unpublished experiments.
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

4332 Reconstitution of Cytochrome c-deficient Yeast Mitochondria Vol. 241, No. 19
ml of distilled water in the hopper. Care was taken to avoid freezing. With the mill still running at low speed the water was quickly drained, and the cold yeast-bead suspension immediately added. The powerstat of the mill was then set at 85 (120 volts), the gap was closed to a setting of 35, and the yeast was homoge- nized for 25 set with the mutant strain, and 30 set with the normal strain. The optimal gap setting and homogenization time were chosen to maximize t,he yield of mitochondria and capacity for phosphorylation, since both mitochondrial damage and cell breakage are increased by lowering the gap setting and by lengthening the homogenization time. At the end of the homogenization period, the gap was opened to 60 and the mill speed was reduced. The homogenate was drained, and a second suspension of 40 g of cells and 70 g of beads was homogenized. Additional batches were homogenized as desired. When the last homogenization was complete, the mill was quickly rinsed by adding 50 ml of cold Medium MTEB and operating the mill at top speed (gap 60) for about 30 set to force the medium through the recycling tube, and the rinse was pooled with the homogenate. The final temperature of this mixture was about 4”. The pH, which was usually slightly higher than 7.0, was immediately adjusted to 7.1, and the beads were separated by decantation. The beads were then washed with 100 ml of Medium MTEB, and the wash w-as added to the homogenate.
The homogenate was centrifuged for 10 min at 780 X g to remove debris and nuclei. Care was taken in decanting the supernatant fraction to avoid its contamination from any loosely packed layer on the pellet. This layer was usually absent in mutant yeast preparations. Mitochondria and mitochondrial membrane fragments were collected by centrifugation of the decanted supernatant fraction for 20 min at 8800 X g. The resulting supernatant layer had a pH of about 6.7. The pellet was suspended in Medium MTEB to give a final volume of 20 ml and fractionated again by centrifuging at 1400 x g for either 20 min (strain D-247) or 15 min (strain D-261). The resulting pellet contained a few whole cells, nuclear material, possibly some cell wall fragments and glycogen, as well as a considerable amount of mitochondrial material. This fraction of the mito- chondria was discarded since it exhibited a high level of endoge- nous respiration which obscured oxidative phosphorylation measurements. Washing and final separation of reasonably intact mitochondria from the bulk of the phosphorylating mito- chondrial membrane fragments was accomplished by centrifuging the supernatant fraction from the preceding step for 20 min at 6800 x g. The resulting mitochondrial pellet displayed a high level of oxidative phosphorylation with either pyruvate-malate or succinate as substrates. The yield of mitochondrial protein in the final fractionated pellet varied from 15 to 25 mg of mito- chondrial protein per 40-g sample of cells (corrected for nitrogen contribution of the suspending medium). The supernatant fraction, which was ordinarily discarded, contained particles displaying appreciable oxidative phosphorylation only with substrates linked to membrane-bound dehydrogenases, e.g. succinate and NADH.
Method B-This procedure was employed only in the experi- ments reported in Table I. Larger quantities of cells, 100 or 150 g, were suspended in a final volume of 200 or 300 ml with Medium MTEB (without pH adjustment), chilled, and placed in the hopper while the mill was running at low speed. Chilled beads were added directly to the suspension in the mill. The power- stat was set at 80 (112 volts), the gap was set at 30 and homogeni-
zation was carried out for 45 sec. The rest of the procedure was the same as that given for Method A. This method, which was used in the earlier phases of the study, gave more variable results than Method A, probably because of greater variations in pH and a greater degree of mechanical damage to mitochondria.
Protein Determination
Protein content of samples was calculated from nitrogen content determined by the micro-Kjeldahl method of Johnson (7).
Measurement of Oxidative Phosphorylation
Oxygen uptake was measured manometrically in Warburg flasks of approximately 20-ml capacity. A total of 2 ml of reaction mixture was placed in the main compartment; the center well contained 0.2 ml of 2 M KOH and fluted filter paper which extended about 0.5 cm above the top of the well.
The side arm contained 0.2 ml of 3.3 M trichloracetic acid. The reaction mixture contained the following components in a total volume of 2.0 ml: 20 pmoles of substrate, 20 pmoles of KH~POI, 2 pmoles of ADP, 20 pmoles of glucose, 150 Kunitz-MacDonald units of hexokinase, 10 pmoles of MgC12, 10 mg of crystalline bovine serum albumin, 60 pmoles of Tris, 2 pmoles of EDTA, and 0.5 to 3.0 mg of mitochondrial protein. The final pH of the reaction mixture was 6.7. The hexokinase, dissolved in a glucose solution, was added just before addition of mitochondria and starting the manometric measurements. All additions were made to flasks at room temperature (23-25”); the incuba- tions were at 25”. Taps were closed after 32 min had elapsed, a reading was taken at 4.0 min, and interval readings were taken every 3 min thereafter. After 30 min of incubation, the tri- chloracetic acid was tipped in from the side arm to stop the reaction. A 0.5-ml aliquot of the reaction mixture was then withdrawn, 5.0 ml of 0.3 M trichloracetic acid were added, protein was removed by centrifugation, and 1.0 ml of the resulting supernatant was used for inorganic phosphate determination by the method of Gomori (8). Completeness of temperature equilibration was checked by including a control flask to which cold Medium MTEB was added instead of the mitochondrial suspension. In only a single experiment (Table I), in which a different Warburg apparatus was employed, was it necessary to correct for incomplete temperature equilibration by the method of Slater and Holton (9).
RESULTS
Reconstitution of Oxidative Phosphorylation with Xuccinate- Table I gives data on the reconstitution of succinate oxidation in mutant mitochondria by addition of horse heart cytochrome c. The mutant mitochondria show very low rates of succinate oxidation compared to normal mitochondria. On addition of horse heart cytochrome c, oxidation is restored to about the same rate that occurs in normal wild type mitochondria. It is most important that addition of the cytochrome c also restores oxida- tive phosphorylation. Whereas the mutant mitochondria give a P:O ratio of only 0.3 in the absence of added cytochrome c, P :0 ratios as high as 1.2 were observed after addition of horse heart cytochrome c and as high as 1.6 when yeast cytochrome c was added. This increase in P:O ratio was invariably observed in many experiments.
The presence of limited oxidative phosphorylation in mutant mitochondria to which no cytochrome c has been added is not
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of Oct.ober 10, 1966 J. R. Mattoon and F. Sherman 4333
unexpected because they already contain the normal complement of iso-2-cytochrome c. Furthermore, the cells from which the mitochondria were isolated were capable of some respiration and growth on ethanol, a nonfermentable carbon source. Since the amount of mitochondrial protein isolated from mutant yeast is as great as, or greater than, that isolated from normal cells, and since added cytochrome c completely restores oxidative phos- phorylation with succinate to these isolated mitochondria, it appears that the relatively low basal respiration observed in mitochondria in the absence of added cytochrome c is due primarily to lack of iso-1-cytochrome c and not to other changes in mitochondrial structure or function. Presumably, the basal rate of respiration in the mutant mitochondria is supported by the small amount of iso-2-cytochrome c known to be present in the cells (1).
Reconstitution of Oxidative Phosphorylation with Other Sub- stratesData in Table II show that oxidative phosphorylation in the mutant mitochondria may also be restored at all three sites with yeast iso-1-cytochrome c. Site III phosphorylation, which is associated with the cytochrome c-cytochrome oxidase segment of the respiratory chain, was measured with GSH rather than ascorbate as substrate because GSH gives far less inter- ference in the phosphate determination. It is apparent from the data that, although the P:O ratios are considerably below theoretical maximum values, all three phosphorylation sites are functional in the reconstituted mutant mitochondria. The third site appears to have been damaged most extensively during mitochondrial preparation. The reconstituted mutant mito- chondria catalyze uptake of oxygen and phosphate with all subst,rates except ethanol at specific rates and P:O ratios nearly equal to, or exceeding, those obtained with mitochondria isolated from normal yeast cells. Two differences between normal and reconstituted mutant mitochondria are illustrated in this table: (a) ethanol oxidation is poorly reconstituted in the mutant mitochondria and (b) the specific activities of GSH + tetra- methyl-p-phenylenediamine oxidation and phosphorylation in reconstituted mutant mitochondria greatly exceed those in the normal mitochondria, while the P:O ratios are nearly the same.
Titration of Mutant Acceptor Mitochondria with Iso-l -cyto-
TABLE I Effects of horse heart cytochrome c on oxidative phosphorylation in
normal and muta.nt mitochondria oxidizing succinate
Mitochondria were prepared by Method B from the normal strain (D-261) grown in 3% ethanol medium for 22.3 hours, and from the mutant strain (D-247) grown in 3y0 ethanol-0.1% glu- cose medium for 25 hours. The 2 ml of the appropriate reaction mixtures contained 10 mM potassium succinate, 14.8 mpmoles of Sigma Type III horse heart cytochrome c, and 0.806 or 1.95 mg of mitochondrial protein, respectively, from normal or mutant strains.
P:O
Substrate
Mutant mitochondria. Normal mitochondria...
NO Plus cyto- cyto-
chrome chrome G ‘
TABLE II Iso-l -cytochrome c stinzulation of oxidative phosphorylation in
normal and mutant mitochondria oxidizing various substrates
Mitochondria were prepared by Method A from mutant (D-247) and normal (D-261) yeast grown 26 and 21 hours, respectively, in 3% ethanol-O.lyO glucose medium. The appropriate reaction mixtures contained: 25 mM GSH; 10 mM for other substrates; 0.3 mM TMPD;e 1.0 mM malate; 1 pg of antimycin B in ethanolic solution; 1.28 mg of mitochondrial protein from the normal strain with GSH and succinate experiments, and 2.56 mg with ot,her experiments; 1.16 mg of mitochondrial protein from the mutant strain with GSH and succinate experiments, and 2.32 mg with other experiments; and 0.6 and 1.2 mpmoles of iso-1-cytochrome c, respectively, with the lower and higher amounts of mitochon- drial protein.
Substrate
Mutant. mitochondria Endogenous GSH + TMPD + an-
timycin A. GSH + TMPD...... Ethanol. Pyruvate + malate.. Succinate SuccinaterJ. . . cu-Ketoglutarate
Normal mitochondria Endogenous GSH + TMPD + an-
timycin A. _. GSH + TMPD...... Ethanol Pyruvate + malat,e. Succinate . a-Ketoglutarate
.I
-
Specific Specific 0% uptake Pi uptake
NO Plus No cyto-
I I
PIUS CytO- cyto- cyto-
chrome chrome chrome chrome ‘ c c c
petoms/hr/mg pmoles/hr/ng mitochondrial mitochondrial
protein protein
<l.O 1.02 0.45 0.69 O.GO
<l.O 4.6C
<l.O <l.O <l.O
<l.O
25.2 0.43 6.32 15.5 1.48 8.58
1.72 0.66 2.97 5.G7 1.74 7.21
11.4 0.55 13.5 11.1 10.9
6.81 0.97 1.1
0.32 0.25 0.55 1.73 1.27 1.18 0.98 1.63
0.00 0.18
8.31 6.16 1.76 0.98 0.21 0.16 5.05 5.41 2.86 3.G3 0.56 O.G7 2.41 4.22 4.40 6.58 1.83 1.56 4.1: 5.58 6.58 6.G9 1.59 1.20
10.7 11.0 10.8 9.74 1.01 0.89 7.21 7.18 10.7 1.5 1.48 1.60
PI0
- No
lx%e c
Plus cyto- hrome
‘
a The abbreviation in the table is. TMPD, tetramethyl-p- phenylenediamine.
b Only 0.09 mpmole of iso-1-cytochrome c was added in this determination, somewhat less than the amount required to ti- trate the mitochondria.
chrome c-The results in Tables I and II show that an increase in P : 0 ratio occurs when deficient mitochondria are reconstituted with an excess of horse heart or yeast iso-1-cytochrome c, respec- tively. Data presented in Fig. 1 show the effect of “titrating” the mutant mitochondria with increasing concentration of puri- fied yeast iso-1-cytochrome c. Increasing rates of oxygen uptake and phosphate uptake were observed. Furthermore, there was an increase in the P : 0 ratio with increasing degree of reconstitu- tion. Low levels of cytochrome c are less effective in recon- stituting the phosphorylation mechanisms than in restoring electron transport. Small errors in cytochrome c or mito- chondrial protein concentrations give rise to relatively large errors in phosphate uptake rate in the accelerated region of the curve. Nevertheless, the somewhat sigmoid curve for restora-
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

4334 Reconstitution of Cytochrome c-dejicient Yeast Mitochondria Vol. 241, No. 19
2 I I I I I I I I I I 0 IO 20 30 40 50
IO’xmpMOLES ISO-I-CYTOCHROME C/mg. PROTEIN
FIG. 1. Titration of mutant mitochondria with iso-l-cyto- chrome c. Increasing amounts of cytochrome c were added directly to Warburg vessels containing 1.17 mg of mitochondrial protein and 20rmoles of potassium succinate per 2.0 ml of medium. Mitochondria were prepared by Method A from mutant yeast (D-247) grown 26 hours in 3y0 ethanol-O.lyO glucose medium.
tion of phosphorylation has been obtained in many similar experiments. Reconstitution of the phosphorylation mechanism approaches, then exceeds, that of the oxidation mechanism with increasing amounts of cytochrome c.
Approximately half-maximal stimulation of respiration was given by about 0.10 mpmole of cytochrome c per mg of protein, and the maximal or saturating level was about 0.18 mpmole per mg of protein; this is considered to be the end point of the titra- tion. If it is assumed that the mutant mitochondria contained iso-2-cytochrome c at 50/, of t.he maximum concentration, then the final content of cytochrome c in reconstituted mitochondria would be about 0.19 mpmole per mg of protein. This saturation value may be compared with the value of 0.15 mpmole of cyto- chrome a per mg of protein obtained spectrophotometrically by Chance and Hess (10) with yeast mitochondria prepared with a Nossal shaker, and 0.17 mpmole of cytochrome c per mg of
0 iPi (ISO-I)
protein (one-half the cytochromes c + c1 value) with rat liver mitochondria.
Titration of Mutant Mitochondria with Iso-d-cytochrome c- Iso-2-cytochrome c was compared with iso-1-cytochrome c in titration experiments shown in Fig. 2A and Fig. 2B. The over-all effectiveness of either type of cytochrome c in recon- stitution of succinoxidase and the accompanying phosphorylation was quite similar. For both types of yeast cytochrome c, approximately half-maximal stimulation of respiration was given by about 0.10 mpmole per mg of protein and the saturation level at about 0.2 mpmole per mg of protein.
Properties of Mitochondria Reisolated after Reconstitution-In a third type of reconstitution experiment, acceptor mitochondria prepared from the mutant were treated with an excess of cyto- chrome c, followed by reisolation and washing to remove any nnbound cytochrome c. The mitochondria were then tested for oxidation and phosphorylation. The data in Table III illus- trate this type of experiment. The mutant acceptor mito- chondria firmly bound not only both types of yeast cytochrome c, but also bound horse heart cytochrome c in such a way that it was functional in respiration. The differences between mito- chondria reconstituted with the two yeast isocytochromes c were usually small, and showed little or no increase in P : 0 ratio when additional exogenous cytochrome c of the same type was added. However, mitochondria reconstituted with highly purified horse heart cytochrome c always gave somewhat lower P:O ratios which could be increased slightly by adding exogenous horse heart cytochrome c. Control experiments in which excess cytochrome c was added directly to mutant mitochondria which had not been reconstituted gave essentially the same behavior as reconstituted mitochondria.
Spectroplwtometric Determination of Cytochrome c Binding- In order to determine whether the end point in a titration experi- ment such as that described above actually represents the point at which mitochondria become saturated with cytochrome c, the binding capacity of normal and mutant mitochondria for iso-1-cytochrome c was determined spectrophotometrically
1.4 j- ,> ISO-
0.6 - I I I I I I I I I I I I I I
10 20 30 40 50 60 70 80 90 20 40 60 80 100 120
IO2 X mpMOLES CYTOCHROME C/mg. PROTEIN 102xmpMOLES YEAST CYTOCHROME C/mg. PROTEIN
FIG. 2. A, comparative titrations of mutant mitochondria with 0.691 mg of mitochondrial protein and 20 pmoles of potassium SUC- iso-1-cytochrome c and iso-2.cytochrome c. This experiment is cinate per 2.0 ml of medium. Mitochondria were prepared by similar to that shown in Fig. 1. The same mitochondrial prepara- Method A from yeast (D-247) grown 26 hours in 3% ethanol-0.1% tion was used for the entire experiment. Flasks containing simi- glucose medium. B, changes in P:O during titration of mutant lar amount,s of iso-l- and iso-2-cytochrome c were arranged in al- mitochondria with iso-l- and iso-2-cytochrome c. These data ternating pairs on the Warburg apparatus. Each flask contained are from the same experiment described in the legend of Fig. 2A.
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from
Issue of October 10, 1966 J. R. Mattoon and F. Sherman 4335
The data in Table IV show that under the conditions of ionic strength and pH used in the reisolation studies, the capacity of yeast mitochondria to remove iso-1-cytochrome c from solution far exceeds the saturation value of about 0.2 mpmole per mg of protein for electron transport as obtained by the titration experi- ments. Thus, there appear to be two species of bound cyto- chrome c, differing according to the class of binding site each oc- cupies: one species is functional, apparently because it occupies a class of site in the mitochondrial membrane which permits re- covery of full oxidative and phosphorylative capacity; the other species is essentially nonfunctional, since it appears to bind at a class of site(s) where it exerts only a minor effect, if any, on succinate oxidation and phosphorylation. Most of the cyto- chrome c bound by normal yeast mitochondria is undoubtedly of this second, nonparticipating type (cf. Tables I and II).
Two factors which affect the degree of cytochrome c binding are the concentration of the mitochondria and the concentration of cytochrome c. Table IV shows that the specific binding (mil- limicromoles of cytochrome c bound per mg of mitochondrial pro- tein) increases markedly with the degree of mitochondrial dilution. This phenomenon suggests that dilution exposes more nonfunc- tional binding surface per mitochondrion, perhaps as a result of mitochondrial swelling and disruption of the outer membrane. This may be related to the relatively high capacity of water- washed or fragmented animal mitochondria to bind relatively large amounts of cytochrome c (11-15). The effect of the con- centration of cytochrome c in the medium upon the level of cyto- chrome c binding is also shown in Table IV (Lines 5 and 6). It
TABLE III
Oxidative phosphorylation catalyzed by mitochondria reisolated after reconstitution with difierent types of cytochrome c
Mitochondria were prepared by Method A from mutant yeast (D-247) grown 27 hours in 3% ethanol-0.1% glucose medium. Reconstitution was carried out by incubating 30 mpmoles of cytochrome c (in Medium MTEB) and 11.3 mg of mitochondrial protein in a final volume of 3 ml for 5 min with frequent mixing in an ice bath. Control mitochondria were treated in the same way except cytochrome c was omitted from the reaction mixture. Reconstituted mitochondria were recovered by a IO-min centrif- ugation at 15,000 X g, washed with about 3 ml of Medium MTEB, and recovered by a similar centrifugation. Warburg vessels con- tained 0.55 to 0.72 mg of mitochondrial protein (reconstituted), 3.0 mpmoles of cytochrome c (when present), and 20 pmoles of potassium succinate in a 2.0-ml volume.
Type of Type of cytochrome c cytochrome c added before added to
reisolation reaction mixture
Iso-l None Iso-l Iso-l Iso- None Iso- Iso- Horse heart None Horse heart Horse heart None None None Iso-l None Iso- None Horse-heart
-
.-
I
-
Specific Spt?CiiiC 0% uptake Pi Uptake
uztoms hrlmg protein
14.9 16.1 16.0 16.7 20.1 19.4
<2.0 21.6 16.5 19.2
I MWltX/h~/~g protein
22.8 21.1 19.7 21.3 21.2 22.9
1.93 27.6 26.4 20.6
-
-
P:O
1.53 1.31 1.23 1.28 1.05 1.18
1.28 1.59 1.07
TABLE IV
Spectrophotometric determination of Go-1 -cytochrome c binding to mutant and normal mitochondria
Cells were grown in 3y0 ethanol-0.1% glucose medium for 26 and 21 hours, respectively, for mutant and normal strains. Reac- tion mixtures contained the amounts of mitochondrial protein indicated, and 0.05 to 0.6 ml of 100~~ iso-1-cytochrome c dissolved in Medium MTEB. Final volume was adjusted to 2.5 ml with Medium MTEB. Binding was allowed to proceed 5 min at 0” with frequent gentle mixing. Mitochondria were removed by centrifugation at 50,000 X g for 10 min. Free cytochrome c in the supernatant was determined after reduction with dithionite by measuring absorption at 550 rnp. Appropriate blanks without additional cytochrome c were included.
Yeast strain ’
Mutant Normal Normal Normal Mutant Mutant Mutant Mutant Normal
Mitochon- lrial protein
added
mg 4.74 4.24 5.52 1.38 0.335 0.335 0.201 0.067 4.24
Initial Final ytochrome c cytochrome c mcentration concentration
PM 14.1 11.42 22.2 3.04 22.6 2.75 11.3 2.36
3.75 1.98 11.3 8.46
7.50 5.56 7.50 6.44 8.84 0
:ytochrome c bound to protein
1.39 11.3
8.98 16.2 13.2 20.7 24.2 39.1
5.21
0
s I I I I -. I 5 IO 15 20 25 30
BOUND CYTOCHROME C (mpMOLES/mg. PROTEIN)
FIG. 3. Scatchard plot of the binding of iso-1-cytochrome c to normal yeast mitochondria. Experimental conditions are given in Table IV. Free cytochrome e &,x) refers to cytochrome c remaining in solution after removal of the mitochondria by cen- trifugation. Mitochondrial protein concentration was 0.55 mg per ml, and initial iso-1-cytochrome c concentrations were from 7.53 to 22.6 PM. A linear relationship would be expected if all sites had equal affinities. The theoretical maximum binding, where all of the binding sites are occupied, is determined by extrapolating to the abscissa intercept.
may also be seen that at approximately equal degrees of dilution, mutant mitochondria display a far lower affiity for cytochrome c than do normal mitochondria (Lines 1 to 3), even though final concentration of free cytochrome c is higher with the mutant
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

4336 Reconstitution of Cytochrome c-dejkient Yeast Mitochondria Vol. 241, No. 19
TABLE V
Phosphorylation coupled to oxidation of lactate, succinate, a.nd pyruvate in normal and mutant mitochondria reconstituted
with iso-I-cytochrome c
Mitochondria were prepared by Method A from yeast grown in 3% ethanol-O.lyO glucose medium for 26 hours. Reconstitution was as described in Table IT’, except for use of 115 mpmoles of iso-l-cytochrome c, 81 mg of mitochondrial protein, and a final volume of 5.2 ml. The wash volume was approximately 5 ml. The appropriate Warburg vessels contained: 1.04 or 1.40 mg of protein, respectively, of normal or reconstituted mitochondria; 1.0 mM malate; and 10.0 rnM for other substrates.
Specific Pi uptake P:O
Substrate Sormal mito-
chondria
Ryo$- S0rltlal mito- mito-
chondria chondria
Malate. n-Lactate. n-Lactate + malate. Pyruvate + malate. Succinate
pntoles/hr/ng protein
0.57 0.57 0.78 0.86 G.86 5.36
11.2 8.71 12.G 14.8
1.63 1.44 1.48 2.36 1.01 1.15
TABLE TI Phosphoyylation in iso-l- and iso-Z-reconstituted mutant
mitochondria oxidizing lactate, ethanol, or succinate
Mitochondria were prepared by Method A from mutant yeast grown in 3% ethanol-O.lyO glucose medium for 243 hours. Re- constitution was carried out as described in Table 11. except for use of 50 mpmoles of iso-l- or isoS-cytochromes c, 25 mg of mito- chondrial protein, and a final volume of 5.4 ml. Wash volume was about 5 ml. Warburg vessels contained 1.54 mg and 1.50 mg of protein for iso-l- and iso-2-mitochondria, respectively. Iso-2-cytochrome c was a freshly dialyzed preparation extracted from wild type yeast, whereas iso-1-cytochrome c had been frozen and thawed after dialysis. Malate was added to a final concen- tration of 1.0 mM; other substrates were 10 mM.
Substrate
Endogenous n-Lactate. n-Lactate + malate. L-Lactate. L-Lactate + malate. Ethanol Ethanol + malate. Succinate
-
Specific Specific 0% uptake Pi uptake P:O
ISO-l- Iso-2. Iso-l- Iso-2- [SO-l- mito- mito- mito- mito- mito- chon- chon- chon- chon- chon- dria dria dria dria dria
- ,,atoms/hr/m~
protein pmoles/hr/mg
prolein
1.47 0.89 1.85 0.71 7.60 6.61 1.09 0.71 1.67 0.89 5.11 6.41 5.89 5.61
.1.6 12.7
4.40
3.28 3.69
I -
ISO-Z- mito- chon- dria
1.72 1.31
1.56 1.60
2.05 1.69 1.48
mitochondria. However, if mutant mitochondria are diluted sufficiently, a high binding capacity can be observed (compare Lines 1 and 5). Thus, the capacity of the mitoehondria for nonfunctional binding is several times the capacity for func- tional binding (Fig. 1).
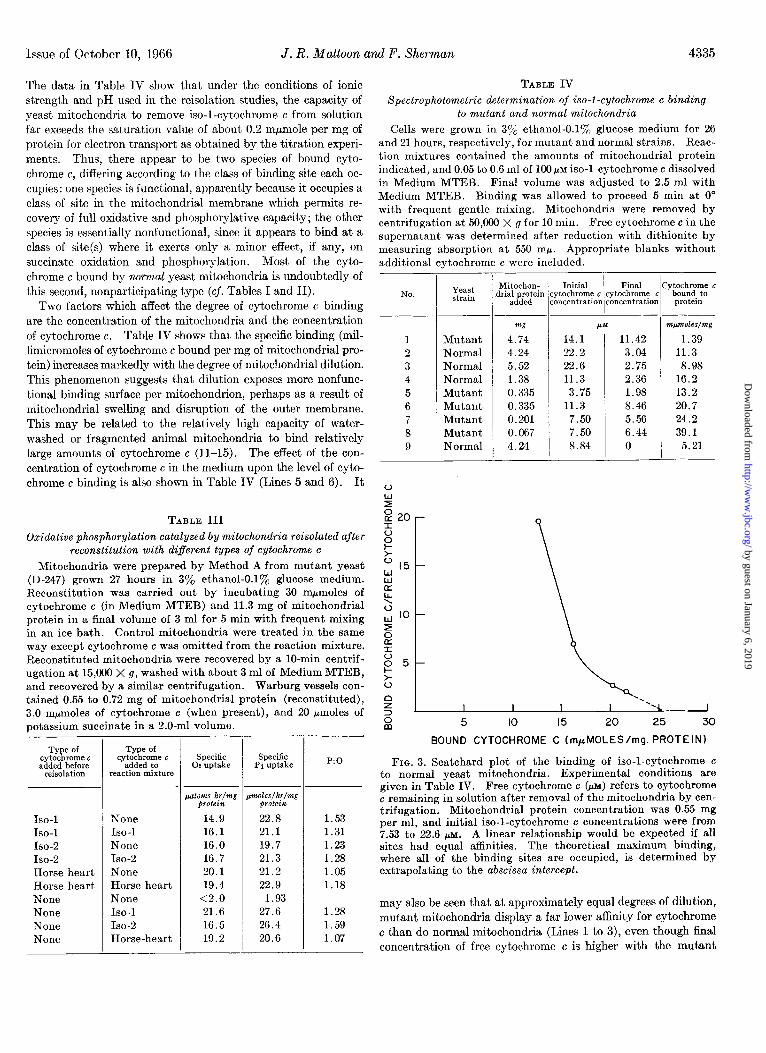
Fig. 3 is a Scatchard plot (16) relating the bound cytochrome c
to the ratio of bound to free cytochrome c in normal wild type mitochondria which already contain a full complement of func- tional cytochrome c. Although the data are not extensive, the results are consistent with the view that in addition to the func- tional binding sites there are at least two classes of nonfunctional binding sites having different binding constants. Alternatively, the marked curvature may indicate an interaction of binding sites which is dependent upon the degree of site saturation. This figure also clearly indicates that the total number of such sites, determined by extrapolation, far exceeds the number necessary completely to reconstitute oxidative phosphorylation in mutant mitochondria. Thus, in normal mitochondria, the total number of nonfunctional sites is sufficient to bind approximately 25 mpmoles per mg of protein, 125 times the number of functional sites indicated by titration experiments.
At appropriate levels of mitochondrial protein andcytochrome c, all detectable cytochrome c can be removed from solution by either normal or mutant mitochondria. This affinity of yeast mitochondria for cytochrome c makes it unlikely, therefore, that the reisolation of mutant mitochondria after reconstitution with cytochrome c will result in complete removal of the excess of nonfunctional cytochrome c which may be bound, even when the reisolated mitochondria are washed with fresh medium.
Reconstitution of Phosphorylation Coupled to Lactate Oxidation- It was of particular interest to determine whether oxidative phosphorylation could be reconstituted with lactate as substrate since the mutant strain gives no growth in synthetic medium when m-lactate is the sole carbon source, although it does grow at reduced rates on other nonfermentable carbon sources such as ethanol and glycerol. It has previously been shown that this deficiency does not stem from a low content of the primary dehydrogenases (3). Mutant mitochondria were reisolated and washed after reconstitution with iso-1-cytochrome c in an effort to minimize the contribution of “external” cytochrome c which might contribute to a possible by-pass of the “normal” site of entry of electrons from lactate (17). Oxidative phosphorylation with n-lactate was found to proceed at a low rate in either normal or reconstituted mutant mitochondria (Table V), but the pres- ence of malate “sparker” increased the phosphorylation rate in both types of mitochondria to 35 to 50% of the rate with suc- cinate. Inasmuch as the P:O ratios obtained with n-lactate + malate are similar for the two types of mitochondria, it seems likely that reconstitution was complete. It must be kept in mind, however, that relatively large amounts of “extra” (non- specific) cytochrome c might remain bound to reconstituted mitochondria and cause an increase in the oxidation rate of D-
lactate. Table VI illustrates an experiment designed to test the relative
effectiveness of D- and L-lactate utilization by mutant mitochon- dria reconstituted with iso-l- and iso-2-cytochrome c, respectively. Data for succinate and ethanol are included for comparison. Both types of yeast cytochrome c are effective in reconstituting the capacity for utilization of n-lactate when malate sparker is present although phosphorylation is somewhat lower with iso-2- cytochrome c. L-Lactate was utilized very inefficiently, even in the presence of malate, particularly when iso-2-cytochrome c was used for reconstitution. In other experiments not shown here, normal mitochondria oxidized L-lactate plus malate sparker at about the same rate as n-lactate plus malate sparker. The data in Table VI also show that utilization of ethanol, unlike lactate, is not affected greatly by addition of malate sparker.
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of October 10, 1966 J. R. Mattoon and F. Sherman 4337
DISCUSSION
In this paper, we have demonstrated the successful use of mitochondria from a cytochrome c-deficient mutant for reconsti- tution in vitro of phosphorylating electron transport. The mu- tant mitochondria were reconstituted to essentially 100% of the corresponding normal values after the addition of cytochrome c; in fact, reconstituted activities frequently exceeded wild type ac- tivities. Even though the normal mitochondria employed in this study had undergone considerable mechanical damage, as indica- ted by somewhat low P:O ratios, it appears that the mutant and wild type mitochondria had been affected equally duringisolation. Consequently, the genetically altered mitochondria are not sub- ject to nonspecific loss of proteins and cofactors which undoubt- edly occurs when normal mitochondria are depleted of cyto- chrome c by chemical methods such as hypotonic treatment and extraction of cytochrome c with saline solution. Respiration in liver mitochondria from which cytochrome c had been removed by the latter method could be reconstituted with cytochrome c, but the P:O ratios obtained with succinate as substrate were considerably below those of normal liver mitochondria (4).
The increase in P : 0 with increasing degree of reconstitution in titration experiments may be interpreted in different ways. Perhaps at low levels of reconstitution, where ATP formation is severely limited by respiration, a significant fraction of the glu- cose &phosphate formed by the hexokinase-glucose trapping system is hydrolyzed as a result of phosphatase action. The relative amount of “drain” caused by phosphatase action would be less at the higher rates of ATP formation which occur with in- creasing degrees of reconstitution of electron transport. Alter- natively, this phenomenon may be a reflection of a “coupling factor” role for cytochrome c. It may be added that substitution of 2-deoxyglucose for glucose gave no increase in P:O with wild type mitochondria prepared with the colloid mill, although such an effect has been observed with particles prepared with a Nossal shaker (18).
No major differences in function of the two types of yeast iso- cytochrome c were revealed by comparative titration experiments in which succinate was used as substrate. The affinity of cyto- chrome c-deficient respiratory assemblies for either type appears to be almost the same. However, horse heart cytochrome c was functionally bound somewhat less effectively than yeast cyto- chrome c, since the concentration required to reach half-maximal stimulation of respiration is considerably greater than the corre- sponding concentration of iso-l-cytochrome c.~ Nevertheless, all three types of cytochrome c were readily removed from solution by the deficient mitochondria and bound with sufficient tenacity to prevent removal by washing once with fresh medium. The orientation of the bound cytochrome c, regardless of type, is such that electron transport from succinate proceeds at normal or nearly normal rates. However, phosphorylation efficiency with horse heart cytochrome c appears to be slightly lower than that obtained with iso-1-cytochrome c. The spectrophotometric studies of iso-1-cytochrome c binding show that the assessment of the efficiency of a given type of cytochrome c in reconstitution experiments should take into account the contribution of any nonspecifically bound cytochrome c, which under certain condi- tions may exceed the normal level by several fold.
Preliminary experiments4 with the disulfide dimer of iso-l- cytochrome c show that a large excess of this form relative to the
4 J. R. Mattoon and F. Sherman, unpublished experiments.
normal thiol form is required to reconst,itute phosphorylating elect,ron transport. In manometric experiments with this ma- terial, oxygen uptake shows a continuous acceleration, indicating that a possible disulfide-thiol conversion may be occurring during incubation. In this connection, it is of interest that Nickerson and Falcone (19) have presented preliminary evidence for the existence in yeast mitochondria of a protein disulfide reductase which acts on cell wall proteins.
A significant feature of the experiments with lactate is the high degree of dependence of oxidation of either isomer on sparker addition. The marked latency of the mitochondrial lactate oxidases may also be overcome in other ways. For example, added cytochrome c stimulated n-lactate oxidation with yeast particles prepared in NaCl medium (20) or with particles pre- pared in 0.25 M sucrose and assayed in a medium laclcing added Mg++ (21). With the latter particles, low pH (6.0) and high Mg+f concentration (35 mM) induced a high rate of oxidation of n-lactate, which was insensitive to added cytochrome c. It is probable, however, that stimulation of D- or L-lactate oxidation by either external cytochrome c or high Mg++ concentration (35 mM) is accomplished by introducing abnormal, “nonphysiologi- cal” conditions. In this connection, it was noted by Gregolin and Scalella (17) that increasing Mg++ concentration from 8 to 35 mM markedly lowered P : 0 ratios for both lactate isomers.
Another factor which influences the sensitivity of n-lactate oxidase to added cytochrome c is the degree to which mitochon- dria have been damaged during preparation. n-Lactate oxidase activity diminishes with increasingly severe cell disintegration procedures, and there is a decline in the ratio of oxidase activity to n-lactate-cytochrome c reductase activity (measured with added cytochrome c in the presence of azide) (21). It is possible that the low levels of D- and L-lactate oxidation (high degree of latency) observed in the present study actually reflect a degree of mitochondrial intactness and that the strong stimula- tion obtained by sparker addition is a useful “physiological” means for studying lactate oxidase. There are many well known examples of correlation between latency of various mit,ochondrial enzymes and intactness of mitochondrial structure. However, .a rigorous assessment of phosphorylation coupled to lactate oxida- tion must take into account both the effects of Mg++ and the contribution of pyruvate oxidation.
Iso-1-cytochrome c poorly restored oxidative phosphorylation with L-lactate even in the presence of added malate sparker. Perhaps the absence of iso-1-cyt.ochrome c during the formation of the mutant mitochondrion results in a nonspecific change or loss from the respiratory assembly of some labile component or chemical grouping essential for L-lactate oxidation which cannot be reconstituted completely simply by rebinding the iso-l-cyto- chrome c in vitro. Although cytochrome bz (L-lactate dehydro- genase) itself is rather easily detached from mitochondria, added cytochrome bz (Worthington) gave no stimulation of L-lactate oxidation or phosphorylation with mutant mitochondria.4
One important question to be asked concerns the roles of the two isocytochromes c with respect to lactate utilization. Since cyl mutants are unable to utilize nn-lactate for growth, it is of considerable interest to determine whether this inability is caused simply by a general cytochrome c deficiency, or whether it is caused specifically by loss of iso-1-cytochrome c. It has been reported that, in nonphosphorylating yeast respiratory particles, iso-2-cytochrome c was only about 25% as efficient as iso-l- cytochrome c in stimulating n-lactate oxidase, whereas little or no
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

4338 Reconstitution of Cytochrome c-de$cient Yeast MitochondGa Vol. 241, No. 19
difference was observed in L-lactate oxidation (2). The data in Table VI indicate that under more favorable test conditions (phosphorylating mitochondria supplemented with malate to avoid limitations of supply of citric acid cycle intermediates) iso-2-cytochrome c is as effective as iso-1-cytochrome c in restor- ing n-lactate oxidation. However, phosphorylation efficiency is considerably less with iso-2-cytoehrome c. When malate sparker is omitted, differences appear to be greater. Iso-2-cytochrome c is even less effective than iso-1-cytochrome c in restoring phos- phorylation coupled to L-lactate oxidation, even when sparker is added. In summary, the reconstitution experiments suggest that both the low cytochrome c concentration and the exclusive presence of iso-2-cytochrome c in cgl mutants contribute to the inability of the mutant to utilize nL-lactate for growth.
2. SLONIMSKI, P.P., ACHER, R.,P6ti, G., SELS, A., AND SOMLO, M., M&hanismes de rkgulation des activitSs cellulaires chez les microorganismes, Marseille, Centre National de la Re- cherche Scientifique, Paris, 1965, p. 435.
SHERMAN, F.. AND SLONIMSKI, P. P., Biochim. Biophus. Acta. 90, 1 (1964j.
_ I
JACOBS, E. E., AND SANDI, D. R., J. Biol. Chem., 235, 531 (1960).
V~~RA,’ J. J., AND JOHNSON, M. J., J. Biol. Chem., 220, 33 (1956).
It has been demonstrated3 that the structure of iso-l-cyto- chrome c is controlled by the chromosomal gene CYI. This implies that iso-1-cytochrome e or at least messenger RNA de- termining iso-1-cytochrome c, is synthesized outside of mito- chondria. From the results of this investigation, one can easily visualize the assembly of cytochrome c in viva into the mito- chondrial apparatus. In other words, one may suggest that cyto- chrome c is synthesized in the cytoplasm and is spontaneously incorporated into the mitochondria at a specific functional site.
We are extending the investigation of oxidative phosphoryla- tion with yeast mutants that are deficient in other cytochromes such as cytochrome a. It is anticipated that recent improve- ments in methods for preparing yeast mitochondria (22-25) will eliminate some of the problems imposed by mechanical damage.
7. 8. 9.
10. 11. 12.
KIELLEY,~.~., in S. P. COLOWICK AND N. 0. KAPLAN (Edi- tors), Methods in enzymology, vol. VI, Academic Press, Inc., New York, 1963, p. 272.
JOHNSON. M. J., J. Biol. Chem.. 137, 575 (1941). GOMORI, G., J.‘Lab. Clin. Med:, 27, 955 (1942). SLATER. E. C.. AND HOLTON. F. A.. Biochem. J.. 56. 28 (1954). CHANCE, B.,-A&D HESS, B., i. Bioi. Chem., 234,‘2464 (19‘59). ’ BEINERT, H., J. Biol. Chem., 190, 287 (1951). GREEN, D. E., MII, S., AND KOHOUT, P. M., J. Biol. Chem.,
217, 551 (1955).
Acknowledgments-We wish to acknowledge the excellent technical assistance of Mr. Richard Gottal, Mr. Wayne Campbell Mr. Andrew Kruzel, and Mr. Alan Howard. We also wish to acknowledge the expert advice of Dr. A. L. Lehninger.
13. 14.
15.
16. 17.
18.
19. 20.
21.
22.
23.
24.
GAMBLE, J. L., Biochim. Biophys. Acta, 23,306 (1957). RICHARDSON, S. H., AND FOWLER, L. R., Arch. Biochem.
Biophys., 100, 547 (1963). SMITH, L., AND MINNAERT, K., Biochim. Biophys. Acta, 105,
1 (1965). SCATCHARD, C., Ann. N. Y. Acad. Sci., 51, 660 (1949). GREGOLIN, C., AND SCALELLA, P., Biochim. Biophys. Acta,
92, 163 (1964). UTTER, M. F., KEECH, D. B., AND NOSSAL, P. M., Biochem.
J., 68, 431 (1958). NICKERSON, W.J., AND FALCONE, G.,Science, 124,318 (1956). GREGOLIN, C., AND SINGER, T. P., Biochem. Biophys. Res.
Commun., 4, 189 (1961). GREGOLIN, C., SCALELLA, P., AND D'ALBERTON, A., LVature,
204, 1302 (1964). OHNISHI, T., AND HAGIHARA, B., J. Biochem. (Tokyo), 66,
584 (1964). DUELL, E. A., INOUE, S., AND UTTER, M. F., J. Bacterial., 88,
1762 (1964).
REFERENCES
MATTOON, J. R., AND BALCAVAGE, W. X.,in S.P. COLOWICK AND N. 0. KAPLAN (Editors), Methods in enzymology, Aca- demic Press, Inc., New York, in press.
1. SHERMAN, F., TABER, H., AND CAMPBELL, W., J. Mol. Biol., 25. OHNISHI, T., KAWAGUCHI, K., AND HAGIHARA, B., J. Biol. 13, 21 (1965). Chem., 241, 1797 (1966).
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from

James R. Mattoon and Fred Sherman-deficient Yeast MutantcCytochrome
Reconstitution of Phosphorylating Electron Transport in Mitochondria from a
1966, 241:4330-4338.J. Biol. Chem.
http://www.jbc.org/content/241/19/4330Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/241/19/4330.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 6, 2019http://w
ww
.jbc.org/D
ownloaded from