“recombinant cellulase production in...
TRANSCRIPT

“Recombinant cellulase production in plants”
Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der
RWTH Aachen University zur Erlangung des akademischen Grades eines
Doktors der Naturwissenschaften genehmigte Dissertation
vorgelegt von
Holger Klose, M.Sc.
aus Lüdenscheid
Berichter: Universitätsprofessor Dr. Rainer Fischer
Universitätsprofessor Dr. Björn Usadel
Tag der mündlichen Prüfung: 12. Mai 2014
Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfügbar.

Meinen Eltern

Table of contents
i
Table of contents
Table of contents .......................................................................................................................................i
Abstract .................................................................................................................................................... iii
Kurzzusammenfassung ........................................................................................................................... iv
Chapter 1: Introduction ............................................................................................................................ 1
1. Bioenergy from biomass .................................................................................................................. 1
2. The next generation of advanced biofuels ....................................................................................... 2
3. Processing lignocellulosic biomass ................................................................................................. 4
3.1 Biomass recalcitrance – the structure and composition of plant cell walls ................................ 4
3.2 Pretreatment technologies for lignocellulosic biomass .............................................................. 7
3.3 Enzymes for cellulose conversion .............................................................................................. 9
3.4 Advanced feed stocks for fuels ................................................................................................ 11
4. Expression of recombinant glycosylhydrolases in plants .............................................................. 12
Objectives of this study ...................................................................................................................... 16
Chapter 2: Differential targeting of a mesophilic endoglucanase .......................................................... 17
Results ............................................................................................................................................... 17
Analyzing the transient expression of TrCel5A in tobacco plants .................................................. 17
Analyzing the localization of TrCel5A by transient expression in tobacco plants .......................... 19
Generation of transgenic tobacco plants expressing TrCel5A ....................................................... 21
Biochemical characteristics of TrCel5A produced in planta ........................................................... 23
Subcellular localization of TrCel5A in tobacco plants .................................................................... 24
Growth characteristics of the transgenic tobacco plants ................................................................ 25
Chemical analysis of tobacco cell walls ......................................................................................... 27
Hydrolysis of leafy biomass from transgenic tobacco .................................................................... 29
Discussion .......................................................................................................................................... 30
Chapter 3: Ethanol-inducible endoglucanase expression in tobacco.................................................... 35
Results ............................................................................................................................................... 35
Generation of transgenic tobacco plants expressing inducible TrCel5A ....................................... 35
Inducible TrCel5A expression in tobacco plants ............................................................................ 37
Growth characteristics and histology ............................................................................................. 38
Chemical analysis of tobacco cell walls in alcR::TrCel5AAP plants ................................................ 40
Discussion .......................................................................................................................................... 41

Table of contents
ii
Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco ........................................... 43
Results ............................................................................................................................................... 44
Expression of SSO1354 in tobacco plants and downstream purification ...................................... 44
Biochemical properties and catalytic activity of plant-derived SSO1354 ....................................... 47
SSO1354 activity in ionic liquids .................................................................................................... 49
Discussion .......................................................................................................................................... 50
Chapter 5: Summary, Conclusion and Outlook ..................................................................................... 53
Chapter 6: Materials and experimental procedures .............................................................................. 57
Materials ............................................................................................................................................. 57
Synthetic oligonucleotides .............................................................................................................. 57
Plasmids ......................................................................................................................................... 58
Bacterial strains .............................................................................................................................. 58
Plant material ................................................................................................................................. 59
Experimental procedures ................................................................................................................... 59
Cloning ........................................................................................................................................... 59
Protein production in tobacco plants .............................................................................................. 60
Protein chemical methods .............................................................................................................. 62
Enzymatic assays ........................................................................................................................... 62
Chemical analysis of plant cell wall polymers ................................................................................ 64
Histological methods ...................................................................................................................... 65
Conversion of transgenic tobacco leaf material ............................................................................. 65
Statistical methods ......................................................................................................................... 66
References ............................................................................................................................................ 67
Appendix ................................................................................................................................................ 80
Abbreviations ..................................................................................................................................... 80
Register of equipment ........................................................................................................................ 82
Table of figures .................................................................................................................................. 83
Scientific publications ......................................................................................................................... 84
Acknowledgement .............................................................................................................................. 86
Declaration ......................................................................................................................................... 87
Curriculum vitae ................................................................................................................................. 88

Abstract
iii
Abstract
Enzyme-catalyzed lignocellulose conversion is a promising pathway to increase the
feasibility of second generation biofuel production. The development of viable
processes for enzymatic lignocellulose utilization must address different issues to
become economically feasible. One important issue is the production of appropriate
and cost-effective enzymes to deconstruct the complex and heterogeneous structure
of the lignocellulose. Plant based expression of cell wall degrading enzymes is a
promising technique, but it currently faces a number of challenges, such as enzyme
expression levels and auto-hydrolysis of the plant cell walls.
This thesis deals with establishing and evaluating cellulase expression strategies in
tobacco plants, including differential protein targeting, inducible protein expression
and the implementation of enzymes with different biochemical properties, such as
thermophile. Different approaches of in planta produced cellulases are evaluated by
the analysis and their properties as well as by detailed phenotypical analyses, such
as biochemical characterization of plant cell walls. Constitutive high level expression
of a mesophilic cellulase led to detrimental changes in the plant phenotype, including
growth and development. The retention of the enzyme inside the endoplasmic
reticulum could not completely avoid this impact on the plant development. The
application of an alcohol inducible promoter for cellulase production led to enzyme
expression rate without any significant phenotypical effects on the plant.
Hyperthermophilic cellulases are inactive under normal plant growth conditions,
allowing their expression in plants without negative effects on growth and
development. Subsequent heat-inducible activation enables lignocellulose
degradation. The expression rate of these hyperthermophilic enzymes is at lower
levels than that of mesophilic enzymes. However, this enzyme was shown to survive
harsh pretreatment conditions. Therefore it is a promising candidate for an advanced
enzymatic conversion of lignocellulosic biomass.
This study established advanced basic knowledge for plant based production of
cellulolytic enzymes and subsequently the developed and evaluated new production
strategies for cellulases would provide an advanced enzymatic conversion processes
of plant biomass.

Kurzzusammenfassung
iv
Kurzzusammenfassung
Die Umwandlung von Lignocellulose zu fermentierbaren Zuckern durch Enzyme ist
ein vielversprechender Ansatz zur nachhaltigen Herstellung von Biokraftstoffen. Die
Entwicklung realisierbarer Prozesse für die Enzym-katalysierte Nutzung von
Lignocellulose muss verschiedene Gesichtspunkte berücksichtigen, um ökonomisch
rentable und nachhaltige Verfahren zu erstellen. Die kostengünstige Produktion
geeigneter Enzyme zum Abbau der komplexen Struktur lignocellulosehaltiger
Biomasse ist dabei ein wesentlicher Kernpunkt. Pflanzen bieten eine kostengünstige
und vielversprechende Alternative zur Produktion solcher Enzyme. Zurzeit müssen
solche pflanzlichen Systeme jedoch noch hinsichtlich der Enzym-Ausbeute und der
potentiellen Schädigung der Pflanze durch das Enzym optimiert werden.
Die vorliegende Studie diente der Entwicklung und Evaluation von Strategien zur
Expression von Cellulasen in Tabakpflanzen. Hierbei wurden unterschiedliches
Protein-Targeting, induzierbare Expression sowie die Expression Temperatur-
aktivierbarer Enzyme untersucht. Die verschiedenen Ansätze wurden hinsichtlich der
biochemischen Eigenschaften der produzierten Cellulasen analysiert. Zudem wurden
die erstellten transgenen Tabakpflanzen auf Veränderungen des Phänotyps,
insbesondere der biochemischen Zusammensetzung der pflanzlichen Zellwände hin
untersucht. Die konstitutive Expression einer mesophilen Cellulase führte zwar zu
hohen Expressionsraten, jedoch wurden auch signifikant Änderungen im Wachstum
und der Entwicklung der transgenen Pflanzen festgestellt. Die Retention der
Cellulase im Endoplasmatischen Reticulum konnte diesen Einfluss auf die
Entwicklung der Pflanze nicht vollständig verhindern. Die Verwendung eines Alkohol-
induzierbaren Promotorsystems zur Produktion der Cellulase führte hingegen nicht
zu signifikanten Änderungen im Phänotyp der Pflanzen. Vergleichbare Ergebnisse
wurden mit der Verwendung einer hyperthermophilen Cellulase erzielt, die unter den
Wachstumsbedingungen der Pflanze inaktiv vorliegt und erst durch hohe
Temperaturen aktiviert werden kann. Zwar wurde für dieses Enzym eine geringere
Ausbeute erzielt als für das mesophile Enzym, jedoch zeigt es sehr hohe Toleranz
gegenüber den harschen Bedingungen während bei konventioneller
Vorbehandlungsmethoden zur Verzuckerung von Biomasse und ist ein
vielversprechender Kandidat für neue verbesserte Prozesse zur enzymatischen
Biomassekonversion.

Kurzzusammenfassung
v
Die innerhalb dieser Arbeit erzielten Daten ermöglichen erweiterte Erkenntnisse zur
Expression von Cellulasen in Pflanzen. Die daraus erfolgte Entwicklung und
Evaluierung neuer Produktionsstrategien kann in Zukunft einen Beitrag zu einer
verbesserten und nachhaltigen Nutzung pflanzlicher Biomasse liefern.

Chapter 1: Introduction
1
Chapter 1: Introduction
1. Bioenergy from biomass
One of the most important challenges in 21st century is the rapid depletion of fossil
fuel resources. Energy from the sun is the most sustainable alternative to oil, gas and
coal. The surface of the earth receives more than 9000 times more energy than is
consumed by humans (Lewis and Nocera, 2006). This energy can be utilized directly
to produce electricity, e.g. wind turbines and photovoltaic cells. However in contrast
to these utilization forms which are highly fluctuating, plant biomass can also be used
to create liquid or solid fuels, and this process is currently considered the most
effective route to convert the sun’s energy into power (Carroll and Somerville, 2009).
Bioethanol has a comparably long history as a transportation fuel and fuel additive. In
Brazil it has been mandatory to supplement transportation fuels with bioethanol
derived from sugarcane since 1929 (Graham-Rowe, 2011). However, it was the
worldwide oil crisis in 1973 and the launching of the “ProAlcool” program by the
Brazilian government that transformed bioethanol production into an economically
relevant industrial sector (Amorim et al., 2011). In 2008, sugarcane-based fuel
surpassed petroleum-based gasoline in Brazil (Chaddad, 2010) and 95% of all
vehicles sold in Brazil use “flex-fuel”, i.e. any blend of ethanol and gasoline (Amorim
et al., 2011).
First-generation biofuels, e.g. ethanol from starch or sugar based, are
disadvantageous because they compete with food and feed crops. Recent concerns
about energy security and the massive increase in energy consumption has
increased the use of edible crops for the production of fuels, e.g. one third of all
harvested corn was used for bioethanol generation in 2007, resulting in a 73% price
rise by the end of 2010 (Graham-Rowe, 2011). The World Bank and the UN Food
and Agriculture Organization (FAO) reported this as the highest price in the last 20
years, contributing to food riots in Mexico and other developing countries (Fairley,
2011). This underlines the need and the great challenge to provide the growing world
population with sustainable supplies of food and energy.

Chapter 1: Introduction
2
Figure 1: Global demographic development. Between today and 2050, the world population is
predicted to rise above 9 billion people. Each person will also consume more calories per day and use
more energy to power their lives. Food and fuel supplies will need to increase massively to meet this
demand. Reproduced from (Graham-Rowe, 2011).
2. The next generation of advanced biofuels
Instead of using the edible parts of the plant, fuels can be sourced from
lignocellulosic residues to avoid competition between food and fuel. Lignocellulose
(from plant cell wall residues) is a completely different starting material for fuel
production, which contains much more energy than grain at the cost of more difficult
and intensive processing (Carroll and Somerville, 2009). The production costs (up to
US$123 per barrel or €0.6 per liter) are much higher than first-generation biofuels
and this may be the most important drawback to ethanol production from
lignocellulosic sources (Tyner, 2008). However, these production costs are estimates
based mostly on laboratory or pilot scale plants (Sainz, 2011), because there are only
a few large scale facilities worldwide.

Chapter 1: Introduction
3
Nevertheless world leading energy suppliers have started to focus on production
facilities for lignocellulosic biofuels. The joint venture “project liberty” between POET
and Royal DSM will be one of the first commercial-scale facilities for cellulosic
ethanol. The plant is under construction in Emmetsburg, Iowa, and will produce 20-
25 million gallons of ethanol per year from corn cobs, leaf husks and stalks starting in
late 2013 (http://www.poetdsm.com/). Likewise, DuPont is constructing a commercial-
scale cellulosic ethanol facility in Nevada, Iowa, which will become operational in
mid-2014 and will produce 30 million gallons per year
(http://biofuels.dupont.com/cellulosic-ethanol/nevada-site-ce-facility/). Both projects
benefit from the US Department of Energy thanks to the Energy Independence and
Security Act 2007.
Oil and automobile companies have also funded cellulosic ethanol research. Dutch
Royal Shell and Volkswagen have invested in, among others, the Canadian company
Iogen to further develop cellulosic ethanol at a large pilot scale facility in Ottawa,
Canada (http://www.iogen.ca).
European examples include the Spanish company Abengoa Bioenergy and the
Danish company Inbicon. Abengoa runs 14 bioethanol facilities worldwide and a
demonstration plant in Salamanca, Spain, which uses wheat and barley straw for the
production of cellulosic ethanol. Inbicon is a subsidiary of Denmark´s largest energy
supplier Dong Energy. Its demonstration plant in Kalundborg, Denmark, produces 1.5
million gallons of cellulosic ethanol per year (www.inbicon.com). In Germany, the
Swiss company Clariant launched a pilot scale plant for cellulosic ethanol in
Straubing in 2012, supported by German governmental research initiative
BioEconomy 2030 (www.clariant.com). This increased investment is encouraged by
the potential of energy and transportation fuels derived from lignocellulosic biomass.
Up to 30% of global gasoline demand could be met from agricultural crop residues
(Michael et al., 2007). Funding and investment has recently accelerated, focusing on
all steps of the process chain (Sainz, 2011).

Chapter 1: Introduction
4
3. Processing lignocellulosic biomass
Plants have evolved to resist microbial attack (Scheller and Ulvskov, 2010). Plant cell
walls play an important role in this process, by providing a barrier against potentially
pathogenic organisms. A greater understanding of plant cell wall architecture and
adaptive strategies for the deconstruction of plant biomass are therefore key
requirements in the development of novel and sustainable processes for
lignocellulose utilization.
3.1 Biomass recalcitrance – the structure and composition of plant cell walls
Many plant cell walls are basically composed of lignocellulose, which comprises three
carbon-based polymers: lignin, hemicellulose and cellulose.
Figure 2: The primary cell wall of a growing cell as found in most flowering plants. The cellulose
chains were organized in microfibrils which are embedded in a matrix of heterogenous
polysaccharides. This matrix-phase consists mainly of pectic polysaccharides like polygalacturonic
acid and rhamnogalacturonan, the latter is often substituted with small polymeric side groups of
arabinan, galactan and arabinogalactan. Xyloglucans have only a single face and therefore can attach
other glucan chains via hydrogen bonds. Highly-esterified PGA is an important component of the
young cell wall and the junction zones are enriched in the middle lamella. Abbreviations: PGA,
polygalacturonic acid; RG I, rhamnogalacturonan I. Figure reproduced from (Carpita and Gibeaut,
1993).

Chapter 1: Introduction
5
There are primary and secondary cell walls. The primary cell wall is formed when
cells develop and typically does not contain any lignin (Carpita and Gibeaut, 1993;
Cosgrove, 1997).
Secondary cell walls were formed after differentiation as part of structural and
functional specialization (Boerjan et al., 2003; Bonawitz and Chapple, 2010). Lignin
contributes up to 30% of the total weight of some secondary cell walls (Scheller and
Ulvskov, 2010). It is a heterogeneous complex polymer comprising diverse
phenylpropanoid subunits (Boerjan et al., 2003). It provides strength, rigidity and
hydrophobicity, although the amount varies between different species and tissues
(Bonawitz and Chapple, 2010).
Cell wall polysaccharides form two different subgroups; the stereoregular homo-
polymer cellulose forms the microfibrilar phase in all cell walls, whereas the stereo-
irregular heteropolysaccharides are associated with these microfibrils. The latter
consist of structurally and compositionally distinct polysaccharides, often described
as hemicellulose and pectin (Pauly et al., 2013). These polymers are located in the
matrix phase between the fibers and are responsible for covering the fibrils and
connecting them to the lignin polymer in the secondary cell wall (Popper, 2008).
Hemicelluloses are categorized by the components of their backbone, mainly
mannans, glucans and xylans (Pauly et al., 2013; Scheller and Ulvskov, 2010). Pectic
polysaccharides contain large amounts of galacturonic acid and their structure and
composition is even more complex and diverse (Atmodjo et al., 2013). The
biosynthesis of these heterogenous polysaccharides takes place in the cisterna of
the Golgi apparatus by plant glycosyltransferases (GTs) that generate the sugar
backbone and subsequently decorate it with side chain sugars (Scheller and Ulvskov,
2010). These precursors can be trimmed by glycosylhydrolases (GHs) before transfer
to the apoplast and incorporation into the cell wall (Günl and Pauly, 2011; Minic,
2008).
Cellulose is usually the most abundant component of plant cell walls (Endler and
Persson, 2011). It consists of long unbranched chains of β-glucopyranose which are
linked via β 1,4 glycosidic bonds. The sugar chains have a ribbon-like conformation
which allows them to be packed into microfibrils stabilized by intermolecular
hydrogen bonds and van der Waals interactions (Harris and Stone, 2009). Although
cellulose has been described as crystalline this is not true in the classical sense.

Chapter 1: Introduction
6
Native cellulose in living plants varies in structure from amorphous to crystalline
depending on species and tissue (Atalla et al., 2009).
Figure 3: Schematic representation of hemicellulose polysaccharides. Symbols representing
different monosaccharides were adopted from the Nomenclature Committee Consortium for Functional
Glycomics (Varki et al. 2009). Glycosidic linkages are presented in their anomeric configurations (α
and β) and their position, e.g. β4 indicates a β 1,4 linkage. Mannans are abundant and are present in
variable amounts in the cell wall. Glucomannes are connected to the secondary cell wall. Xyloglucans
(XyGs) vary in structure and composition between dicots (where predominantly fucogalactoXyG
appears) and monocots and solanales (where arabinogalactoXyG is present). Xylan is found as
Glucoronoxylan (GX) in dicots but more often as glucoronoarabinoxylan (GAX) in grasses.
Reproduced from (Pauly et al., 2013).

Chapter 1: Introduction
7
Cellulose is synthesized by globular protein complexes located in the plasma
membrane. These so-called cellulose synthase complexes probably use a steryl
glycoside primer and therefore first accept UDP-glucose before forming the long
β 1,4-glucan chains. GHs are also required for cellulose biosynthesis (Minic, 2008).
For example, the endoglucanase KORRIGAN may contribute to the assembly of the
microfibrils and/or chain termination (Molhoj et al., 2001).
The heterologous composition of lignocellulose and the variety of linkages within and
between the different polymers explain the recalcitrance of biomass, and this must be
overcome to develop an economically feasible process for lignocellulose conversion
(Chundawat et al., 2011).
3.2 Pretreatment technologies for lignocellulosic biomass
The first step in the conversion of lignocellulose to single sugars is the pretreatment
of the biomass, which had been practiced for more than 100 years but has seen a
resurgence of interest in the last decade (Saville, 2011). The aim of pretreatment is to
improve the subsequent enzymatic hydrolysis and fermentation. The recovery of
soluble and therefore fermentable sugars should be maximized, whereas energy
costs, chemical inputs and waste stream disposal should be minimized (Galbe and
Zacchi, 2007). Currently, pretreatment represents 18% of overall production costs,
which is the second most expensive step (Mosier et al., 2005; Yang and Wyman,
2008) . The greatest hurdle in the pretreatment step is the resistance of plant cell
walls to deconstruction. This recalcitrance of lignocellulosic biomass reflects the
highly crystalline structure of cellulose, which is embedded into a complex matrix of
hemicellulose and lignin (Himmel et al., 2007). Pretreatment can involve physical,
chemical or biological processes, and based on the heterogeneous nature of
lignocellulosic substrates it is difficult to develop a universal process (Alvira et al.,
2010).
Physical pretreatment technologies include mechanical comminution or extrusion in
order to reduce the size of particles and increase the surface area. Milling, chipping
and grinding produce particles with a size between 30 and 0.2 mm (Sun and Chen,
2008) and are often combined with other pretreatment technologies.
Chemical pretreatment methods can be classified based on the chemical agent.
Alkaline pretreatment uses sodium or ammonium hydroxide to de-esterify the lignin
polymer (Cheng et al., 2010) and to cause swelling and the partial decrystallization of

Chapter 1: Introduction
8
cellulose and hemicellulose (McIntosh and Vancov, 2010). Acid pretreatment uses
diluted or concentrated acids for the solubilization of hemicellulose and lignin with
minimal degradation (Carrasco et al., 1994). However extensive washing and
detoxification is required in both cases before subsequent hydrolysis and
fermentation (Saha et al., 2005), particularly in the case of acid pretreatment which
can generate potential fermentation inhibitors such as furfurals (Marzialetti et al.,
2008).
Over the last decade, so-called green solvents have emerged as alternatives for the
processing of lignocellulosic biomass, including the extensively studied ionic liquids
(ILs). These are salts comprising a small anion and a large organic cation, with a low
vapor pressure that typically means they remain liquid at room temperature (Vancov
et al., 2012). Ionic liquids can dissolve diverse biomaterials including crystalline
cellulose, which is most efficiently dissolved by ILs based on imidazolium (Vancov et
al., 2012; Zavrel et al., 2009). Cellulose regenerated after treatment with ILs was
more amorphous and porous than before (Kilpeläinen et al., 2007), and underwent
more efficient saccharification (Bodirlau et al., 2010). However, ILs are toxic and tend
to inhibit or inactivate enzymes even at low concentrations, thus increasing the costs
of regeneration (Turner et al., 2003). Therefore needed regeneration or separation
steps included in the process increased the overall cost. The development of
improved biorefinery concepts such as consolidated bioprocessing hence requires
the evaluation of solvents that support the activity of enzymes and microorganisms
(Brodeur et al., 2011).
Biological pretreatment involves the use of microorganisms such as brown, white or
soft-rot fungi to break down lignin and hemicellulose (Sánchez, 2009). The fungi
secrete peroxidases and laccases that oxidize lignin (Kumar et al., 2009) and may
also produce additional enzymes and proteins to enhance subsequent
saccharification, such as swollenins (Jäger et al., 2011). The low capital cost of these
systems and their minimal energy and chemical inputs offer important advantages,
but they still offer low conversion rates compared to other systems (Sun and Cheng,
2002). To establish an economically-feasible approach including biological
pretreatment the conversion rates must be improved substantially.
Other technologies for biomass conversion have been developed that combine those
described above, including ammonia fiber expulsion (AFEX) and organosolv, which

Chapter 1: Introduction
9
combine high temperature and pressure with alkaline treatment and organic solvents,
respectively.
3.3 Enzymes for cellulose conversion
The glycosylhydrolase (GH) superfamily includes many diverse polysaccharide-
degrading enzymes (http://www.cazy.org/). Cellulases are less efficient than other
sugar-hydrolyzing enzymes mainly due to the crystalline nature of the substrate.
Figure 4: Current overview on fungal enzymatic degradation of cellulose. EGs cleave
β-glycosidic bonds at internal amorphous sites, generating oligosaccharides of variable length down to
cellobiose and creating new chain ends (Lynd et al., 2002; Zhang and Lynd, 2004). CBHs release
cellobiose from the ends of cellulose chains acting in a processive manner (Lynd et al., 2002). CBH1
and CBH2 hydrolyze from the reducing and non-reducing ends, respectively. BGLs finally convert
cellobiose to glucose. Non-hydrolytic or expansin-related proteins like swollelin (Swo) can transform
crystalline areas into amorphous areas by loosening the hydrogen bonds between the chains (Jäger et
al., 2011). Recently discovered PMOs such as the CBM33 and GH61 families create new ends by
oxidizing glucopyranose units within the cellulose chains (Horn et al., 2012). Most cellulolytic systems
comprise multiple EGs and CBHs, which vary in their specificity for the substrate, acting on different
crystal faces or parts differing in crystallinity and accessibility. The PMO depicted here represents a
GH61 class enzyme, which acts on C4 atoms to generate non-oxidized ends for CBH1 (oxidized
sugars represented by white circles). CDHs could provide PMOs with electrons by oxidizing
cellobiose. Also other non-enzymatic reductants (electron donors) can induce oxidative activity, e.g.
reduced glutathione, ascorbic acid and gallic acid (Žifčáková and Baldrian, 2012). Abbreviations: BGL,
beta-glucosidase; CBH, cellobiohydrolase; CDH, cellobiose-dehydrogenase; EG, endoglucanase;
PMO, polysaccharide mono-oxigenase; Swo, Swollenin. Adapted from Horn et al. (2012) and Jäger et
al. (2012).

Chapter 1: Introduction
10
The complete conversion of cellulose to glucose requires the activity of three different
classes of enzymes: endoglucanases, exoglucanases and β-glucosidases (Jäger and
Büchs, 2012; Sainz, 2011).
Exo-β 1,4-D-glucanases or cellobiohydrolases (CBH; EC3.2.1.91) act on the ends of
the cellulose chains and processively cleave off cellobiose units. There are two
subgroups, depending on whether they act from the reducing or non-reducing end of
the sugar chain. Endo-β 1,4-D-glucanases (EG; EC3.2.1.4) hydrolyse β–glycosidic
bonds within sugar chains and are more active on amorphous rather than crystalline
cellulose.Finally, β-1,4-D-glucosidases (BGL; EC3.2.1.21) cleave cellobiose units into
glucose. The whole enzyme set acts synergistically to convert cellulose into its
subunits. For example, EGs generate new ends for CBHs, and BGLs remove the
cellobiose which inhibits both EGs and CBHs (Kumar et al., 2008). Many GHs feature
additional domains responsible for adsorption onto the substrate. Such carbohydrate-
binding modules (CBMs) were initially found to bind the surface of crystalline
cellulose (Gilkes et al., 1988; Tomme et al., 1988), but more recent studies have
shown that some CBMs also have disruptive functions (Boraston et al., 2004;
Forsberg et al., 2011).
The mesophilic fungus Trichoderma reesei is one of the best-known sources of
cellulolytic enzymes and is the most widely studied cellulolytic system, secreting a set
of five endoglucanases and two exoglucanases in different ratios (Peterson and
Nevalainen, 2012; Rosgaard et al., 2007). It is a key source of commercial cellulase
preparations, but bacterial and archaeal systems have also been investigated
because these microorganisms and their enzymes demonstrate thermotolerance and
survive under other harsh conditions. The incorporation such enzymes into industrial
processes therefore seems a promising approach (Barnard et al., 2010; Blumer-
Schuette et al., 2008). One example is the hyperthermophilic archaean Sulfolobus
solfataricus, which produces several enzymes involved in carbohydrate degradation
(She et al., 2001) that have been evaluated for biomass degradation activity (Cannio
et al., 2004; Cobucci-Ponzano et al., 2010; Girfoglio et al., 2012; Huang et al., 2005;
Limauro et al., 2001).
Metagenomic libraries can also be screened for novel carbohydrate-degrading
enzymes with advanced features for biomass conversion (Ilmberger et al., 2011;
Pottkämper et al., 2009; Voget et al., 2006). Known enzymes can also be re-

Chapter 1: Introduction
11
engineered to enhance their stability and activity (Lehmann et al., 2012; Liang et al.,
2011; Percival Zhang et al., 2006).
Nevertheless, enzymes for cellulose hydrolysis still represent a major component of
overall process costs (Carroll and Somerville, 2009). Up to 25 kg of enzyme is
required per ton of biomass (Taylor et al., 2008). Despite progress towards the
economical production of enzymes by commercial suppliers such as Novozyme and
Danisco-Genencor this remains a major bottleneck in the production of biofuels from
biomass.
3.4 Advanced feed stocks for fuels
Ethanol is produced from starch and sugarcane by established mature industries
(Geddes et al., 2011) but they provide only marginal societal benefits over fossil fuels
(Hill et al., 2006). Environmental life-cycle analysis has revealed the negative impact
caused by the movement of agrichemicals such as nitrogen/phosphorous fertilizers
and pesticides (Vitousek et al., 1997). The competition between food and fuel crops
also remains a major concern (Geddes et al., 2011). Lignocellulosic feed stocks can
address these concerns. The net energy balance (NEB) is the difference between
energy inputs and outputs throughout the production chain, and NEB models for
lignocellulosic biomass estimate ratios of >4.0 which is a major improvement over
corn grain ethanol (1.25) and soybean diesel (1.93) (Hill et al., 2006). Mixtures of
grasses and fast-growing woody plants are promising candidates for the sustainable
production of biofuels from lignocellolosic biomass (Tilman et al., 2006).
The recalcitrance of the plant cell wall could be overcome by genetic engineering,
which is widely used to modify the properties of crops (Taylor et al., 2008). Strategies
to improve plants for biofuel production include increasing the accessibility of the cell
wall, and modification of the lignin biosynthesis pathway (Vanholme et al., 2008), e.g.
by modulating genes that control the ratio of guaiacyl and syringyl subunits (Chen
and Dixon, 2007) or the overall content of lignin (Jung et al., 2013a). Modification can
lead to unwanted side effects such as dwarf phenotypes or reduced water
conductivity and elasticity (Abramson et al., 2013). However, the modification of plant
polysaccharides by the expression of bacterial CBMs and plant GHs has already
achieved increases in yield, growth rate and porosity even though their exact role
and impact on cell wall biosynthesis is yet not fully understood (Shoseyov et al.,
2006) (Abramson et al., 2013).

Chapter 1: Introduction
12
4. Expression of recombinant glycosylhydrolases in plants
The expression of GHs in plants would reduce the costs of production and improve
the economic feasibility of biomass conversion (Sticklen, 2006; Taylor et al., 2008).
This concept was initially thought unfeasible because the enzymes would disrupt
vegetative growth. Indeed the impact of GHs on cell wall biosynthesis depends on
the species, tissue and the enzyme itself (Abramson et al., 2013; Rudsander et al.,
2008; Takahashi et al., 2009), resulting in both beneficial and deleterious effects
(Maloney and Mansfield, 2010; Park et al., 2003). Different strategies to produce GHs
in plants have therefore been tested in model species such as Arabidopsis and
tobacco (Sainz, 2011; Taylor et al., 2008), and in economically-relevant crops such
as rice and maize (Chou et al., 2011; Harrison et al., 2011; Hood et al., 2007).
However, the use of these crops for biofuel production competes with food and feed
resources, hence the recent focus on grasses and woody plants as alternatives (Jung
et al., 2013b; Vogel and Jung, 2001). Several endoglucanases and xylanases have
been successfully expressed in plants using regulatory elements that target different
tissues and subcellular compartments (Sainz, 2011; Taylor et al., 2008). Complex
enzymes with additional CBMs are often expressed in a truncated form which still
confers full activity on soluble substrates (Harrison et al., 2011; Sun et al., 2007). In
this context, endoglucanase Cel5A from Thermotoga maritime fused to CBM6 from
Clostridium stercorarium xylanase was stably produced in tobacco and showed an
improved efficiency for autohydrolysis (Mahadevan et al., 2011).
The stable accumulation of plant-derived GHs has been achieved by targeting
different subcellular compartments and tissues (Sainz, 2011; Taylor et al., 2008).
Subcellular targeting and tissue-specific expression are also promising for the
sequestration of potentially harmful enzymes during plant growth and development
(Taylor et al., 2008). Inducible expression systems can produce enzymes at similar
levels to constitutive systems while avoiding unwanted deleterious effects, and can
also regulate the amount of enzyme by adaptive induction (Lebel et al., 2008).

Chapter 1: Introduction
13
Table 1: Examples of recent progress in the heterologous expression of GHs in plants. Table
adapted and updated from Taylor et al. (2008)
Enzyme Source Host Targeting Properties
and comments
References
AcE1 endoglucanase Acidothermus
cellulolyticus
Tobacco Chloroplast Transplastomic,
modified CD leads to
high expression levels
(Ziegelhoffer et
al., 2009)
PttCel9A1 endoglucanase Populus tremula Arabidopsis Apoplast/
Cell wall
Modified cell wall
sugar contents
(Takahashi et
al., 2009)
AtCel1 endoglucanase
AaXEG xyloglucanase
HvXylI Xylanase
Tv6GAL galactase
RsAraf1
arabinofuranosidase
Arabidopsis
thaliana,
Aspergillus sp.,
Hordeum vulgare,
Raphanus sativus
Poplar Apoplast/
Cell wall
Altered fiber structure
and composition,
accelerated hydrolysis
of plant material
(Kaida et al.,
2009)
BglB β-glucosidase Thermotoga
maritima
Tobacco Cytoplasm,
chloroplasts
Thermostable
enzyme, preserved
activity after
lyophilization
(Jung et al.,
2010)
TmCel5A endoglucanase Thermotoga
maritima
Tobacco Chloroplast Optimized targeting
with RuBisCO small
subunit signal peptide
(Kim et al.,
2010)
CsXynB Xylanase Clostridium
stercorarium
BY2-cells,
Tobacco,
Rice
Secretion Bacterial signal
peptide is partially
functional in plants
(Kimura et al.,
2010)
endoglucanases, cutinase,
exoglucanase, lipase
pectate lyases, swollenin,
xylanase, acetyl xylan
esterase, β-glucosidase
Different bacteria
and fungi
Tobacco Chloroplast Transplastomic plants,
enzyme cocktails
derived from plat
crude extract
hydrolyzed filter paper,
pine wood or citrus
peel
(Verma et al.,
2010)
AtKOR endoglucanase Arabidopsis
thaliana
Poplar Apoplast/
Cell wall
Led to severe defects
in plant growth,
significantly impacts
the ultrastructure of
cellulose
(Maloney and
Mansfield,
2010)
DtXYnB Dictyoglomus
thermophilum
Maize Apoplast Intein modification
enables heat
activation
(Shen et al.,
2012)
Bsx xylanase
CsXynB xylanase
Bacillus sp.
Clostridium
stercorarium
Maize Apoplast Stunted growth
phenotype
(Gray et al.,
2011b)
AcE1 endoglucanase Acidothermus
cellulolyticus
Rice Apoplast Accelerated
Saccharification of the
transgenic plant
material
(Chou et al.,
2011)
TrMan1 mannase Trichoderma reesei Tobacco Chloroplast Transplastomic plant (Agrawal et al.,
2011)

Chapter 1: Introduction
14
AcE1 endoglucanase Acidothermus
cellulolyticus
Tobacco,
Maize
Accelerated
Saccharification of the
transgenic plant
material
(Brunecky et
al., 2011)
TmCel5A endoglucanase Thermotoga
maritima
Tobacco Chloroplast Engineered CBM for
enhanced hydrolysis
rate
(Mahadevan et
al., 2011)
TfBglC β-glucosidase
TfE3 exoglucanase
TfE4 endoglucanase
TfXeg74 xylanase
Thermobifida fusca Tobacco Chloroplast Transplastomic plants,
homoplasmic plants
showed severe effects
on plant health and
fertility
(Petersen and
Bock, 2011)
DtXynA endoxylanase
TlXynB endoxylanase
NtEGA endoglucanase
AcEGB endoglucanase
Dictyoglomus
thermophilum,
Thermomyces
lanuginosus,
Nasutitermes
takasagoensis,
Acidothermus
cellulolyticus
Maize not specified Transgenic stover
showed enhanced
properties when
utilized in SSF
process for ethanol
generation
(Zhang et al.,
2011)
AcE1 endoglucanase Acidothermus
cellulolyticus
Maize Seed embryo Cross breeding for
high enzyme level
accumulation
(Hood et al.,
2012)
TrXyn2 xylanase Trichoderma reesei Poplar Chloroplast Seed embryo due to
sequestration
(Kim et al.,
2012)
Cutinase
Swollenin
Fusarium solani
Trichoderma reesei
Tobacco Chloroplast (Verma et al.,
2013)
Thermophilic GHs with little activity below 50°C can be expressed functionally in
plants without adverse effects because they are inactive during the growth phase and
can be activated post-harvest by a temperature shift (Basu, 2001; Sainz, 2011;
Taylor et al., 2008). The in planta production of thermophilic GHs is also promising
because such enzymes facilitate lignocellulose hydrolysis under extreme conditions
e.g. thus increasing product and reactant solubility, increasing the speed of the
reaction and reducing the amount of enzyme that is needed (Bhalla et al., 2013). The
catalytic domain of endoglucanase E1 from A. cellulolyticus has been produced in
different plant species and showed great potential for biomass processing (Brunecky
et al., 2011; Chou et al., 2011; Hood et al., 2012) but the incorporation in an existing
pretreatment process (AFEX) revealed that there is still room for improvement
(Teymouri et al., 2004).
Plastid transformation could also facilitate the production of GHs in plants, because
proteins accumulate to high levels in chloroplasts without affecting vegetative growth,
there is no gene silencing, and transgene containment is achieved by the maternal

Chapter 1: Introduction
15
inheritance of chloroplast genomes (Scotti et al., 2012). The high-level expression of
individual functional GHs has been achieved (Gray et al., 2011a; Ziegelhoffer et al.,
2009) but these studies have focused predominantly on bacterial enzymes because
the environment of the chloroplast is similar to that of the bacterial cell. However, a
set of GHs from the fungus T. reesei has also been expressed in transplastomic
tobacco, demonstrating the potential of plastid-derived GHs for lignocellulose
hydrolysis (Verma et al., 2010). Remarkably high levels of T. fusca GHs have also
been expressed in transplastomic tobacco, and although there were severe
deleterious effects in homoplasmic plants, the heteroplasmic plants with wild-type as
well as transformed plastids could be grown to maturity (Petersen and Bock, 2011).
Achieving high yields of recombinant enzymes is one of the key challenges when
enzymes are expressed in plants to facilitate biomass conversion in situ.
Conservative estimates suggest that cellulase must represent at least 10% of TSP to
achieve complete hydrolysis, which is far higher than most reported yields (Sticklen,
2006). Approximately 4 kg of cellulase per hectare has been produced in maize
(Hood et al., 2007) but even this is insufficient for the complete conversion of the
stover (Sainz, 2011). Improved cellulase variants are helpful, but further increase in
productivity is also required (Hood et al., 2012).
Recent developments have shown that even low level expression of recombinant
GHs could contribute to an economically-feasible process for biomass conversion.
The impact of GHs on the structure and composition of the plant cell wall could be
used to make the plant biomass less recalcitrant. Maize and tobacco plants
expressing A. cellulolyticus E1cd showed more efficient saccharification and reduced
the severity of pretreatment (Brunecky et al., 2011). Similar approaches include the
expression of recombinant xylanases to enhance saccharification by altering the
structure of hemicellulose (Borkhardt et al., 2010; Shen et al., 2012; Zhang et al.,
2011). Even though the role of GHs in the construction of plant cell walls is not yet
fully understood and the performance of these transgenic plants has not been
evaluated in the field, the in planta production of GHs nevertheless offers hope for
sustainable bioprocessing.

Chapter 1: Introduction
16
Objectives of this study
The objectives of the study were to establish and evaluate different strategies for the
expression of heterologous cellulases in plants, focusing not only on enzyme
properties such as expression level and functionality, but also on their impact on the
plant development. The model system Nicotiana tabacum was used to generate
transgenic lines expressing different endoglucanase genes targeted to different
subcellular compartments, under the control of constitutive and inducible promoters.
The activity and biochemical characteristics of the enzymes were investigated in the
context of their bioconversion potential. The phenotype of the transgenic plants was
studied at the morphological level to determine the impact of cellulases on growth
and development, and also at the cellular and subcellular level to characterize the
impact on the cell wall. This study will provide new insight in the interdependency
between recombinant cellulase expression and the physiological characteristics of
the host plant.

Chapter 2: Differential targeting of a mesophilic endoglucanase
17
Chapter 2: Differential targeting of a mesophilic
endoglucanase
The results of this chapter have been or will be published as follows:
The Analysis of the biochemical characteristics of TrCel5A has been published in:
Klose H, Günl M, Usadel B, Fischer R, Commandeur U. (2013) Ethanol inducible
expression of a mesophilic cellulase avoids adverse effects on plant development.
Biotechnology for Biofuels, 6:53.
The transient localization studies of TrCel5A-mCherry fusion have been published in:
Garvey M, Klinger J, Klose H, Fischer R, Commandeur U (2013) Expression of recombinant
cellulase Cel5A from Trichoderma reesei in tobacco plants. Journal of Visualized
Experiments, in press
The comparison of different localizations of recombinant TrCel5A in transgenic tobacco is prepared to
be published in:
Klose H, Günl M, Usadel B, Fischer R, Commandeur U. Plant cell wall modification by
differential targeting of recombinant endoglucanase from Trichoderma reesei
The Trichoderma reesei genome contains 22 open reading frames (ORFs)
corresponding to known cellulolytic enzymes (Ouyang et al., 2006). This includes
eight genes representing different endoglucanases. TrCel5A (formerly EGII or EGIII)
is the second most abundant endoglucanase in the cell (Jäger et al., 2010; Ouyang
et al., 2006). It comprises a C-terminal catalytic domain from glycoside hydrolase
family 5, a Ser/Pro-rich linker and a family 1 carbohydrate-binding module (CBM)
(Saloheimo et al., 1988). Transient expression studies have already indicated that
this enzyme can be expressed in tobacco plants (Klose, 2009; Werthmann, 2009).
Here, the results were confirmed and analyzed in more detail so that the
endoglucanase could be produced in transgenic tobacco allowing its impact on plant
development to be investigated.
Results
Analyzing the transient expression of TrCel5A in tobacco plants
Constructs containing the TrCel5A sequence had already been prepared, and
included a plant codon-optimized leader peptide derived from the heavy chain of the
murine monoclonal antibody mAb24 for secretion to the apoplast, and a C-terminal
His6 tag for detection and purification. An additional construct was prepared

Chapter 2: Differential targeting of a mesophilic endoglucanase
18
containing a C-terminal KDEL tag to retain the recombinant protein in the
endoplasmic reticulum (ER). Both genes were controlled by the constitutive double
CaMV 35SS promoter. Transient expression was used to confirm the constructs were
functional before using them for the stable transformation of tobacco plants. A further
construct containing only the catalytic domain from codon 92 to 418 of TrCel5A was
cloned for comparison.
Figure 5: Schematic presentation of plant expression cassettes for the differential targeting of
TrCel5A. The CaMV promoter (P35SS) and terminator (pA35S) are indicated in light blue. The 5'-UTR
of chalcone synthase (CHS) and the His6 coding sequence (His6) are indicated in blue. The plant
codon-optimized leader peptide (LPH) derived from the heavy chain of the murine mAb24 is shown in
green. LPH achieves the secretion of the recombinant protein to the apoplast (A). The C-terminal
KDEL signal indicated in red retrieves the protein to the ER (B). Arrows show primer annealing sites to
amplify the CaMV 35SS expression cassette.
Transient expression of the holoenzyme followed by western blots of total soluble
protein (TSP) revealed two major bands with molecular weights of approximately 45
and 35 kDa, as well as additional intermediate degradation products (Figure 6). The
major bands were similar to the molecular weights of the holoenzyme (Ouyang et al.,
2006) and the catalytic domain (Nakazawa et al., 2008). Transient expression of the
catalytic domain TrCel5Acd produced one main product with a molecular weight of
approximately 35 kDa. Freezing and storage without protease inhibitors resulted in
additional small degradation products but these could be avoided by adding PMSF.
Even the addition of several different protease inhibitors did not prevent cleavage into
the two main fragments (Figure 6) although the serine protease-specific inhibitor E-64
prevented the appearance of the intermediate fragments (approximately 40 kDa).

Chapter 2: Differential targeting of a mesophilic endoglucanase
19
Figure 6: Transient expression of TrCel5A in N. tabacum. Western blot of 10 µg TSP per lane,
derived from frozen and stored crude plant extract following the transient expression of TrCel5AAP,
TRCel5AER and the catalytic domain TrCel5Acd (A). Extraction of recombinant TrCel5AAP in liquid
nitrogen with different protease inhibitors (B). NC: 10µg TSP from N. tabacum SR1 wild-type. PC:
purified His6-tagged phenylalanine ammonium lyase (PAL). Recombinant cellulase was
immunostained with the MαHis primary antibody and GAMFCAP as the secondary antibody.
Analyzing the localization of TrCel5A by transient expression in tobacco plants
The subcellular localization of TrCel5A was investigated by joining the C-terminus in-
frame with the mCherry fluorescent marker protein lacking an additional KDEL tag.
Transient expression was confirmed by the appearance of red fluorescence in the
infiltrated leaf areas and also a 70-kDa band on the corresponding Western blot. The
fusion proteins generated different degradation products depending on the presence
of a KDEL tag, e.g. full version (with the N-terminal CBM1) has only been obeserved
in the tagged version (Figure 7). Infiltrated leafs (representing each of the constructs)
showing red fluorescence were used to prepare 90-µm sections with a Leica
Vibrating Blade Microtome and the subcellular localization of the recombinant
proteins was determined by fluorescence microscopy. Fluorescence for TrCel5A-
mCherryAP is clearly located in the apoplastic space (Figure 8 A-C), whereas for
TrCel5A-mCherryER only a diffuse fluorescence signal was observed (Figure 8 D-E).

Chapter 2: Differential targeting of a mesophilic endoglucanase
20
Figure 7: Transient expression of TrCel5A-mCherry fusion in tobacco. Western blots show the
expression of TrCel5AmCherryAP (A) and TrCel5AmCherryER (B and C). Lanes 1, 3 and 5 were loaded
with 20 µg of total soluble protein from transient expression experiments. Lanes 2, 4 and 6 contain
purified and TCA-precipitated recombinant protein from 0.5 g of leaf material. NC: 20 µg of total
soluble protein from wild-type tobacco cv. SR1. The positive control (PC) in (A) and (B) is His6-tagged
PAL, and in (C) it is KDEL-tagged DsRed. The antibody system used in (A) and (B) is
MαHis5/GAMFcAP, and in (C) it is MαKDEL/GAMFc
AP. The presence of the 70-kDa fusion protein was
confirmed by detecting either the His6 tag or (in the case of TrCel5AmCherryER) the KDEL tag.
Figure 8: Localization of transiently-expressed TrCel5A-mCherry fusion in 90-µm sections of
tobacco leaf tissue. The LPH signal peptide promotes the secretion of TrCel5A-mCherry as indicated
by the strong red fluorescence in the apoplast (A-C). A C-terminal KDEL tag retrieves the recombinant
enzyme to the interior of the cell, as indicated by the strong but diffuse red fluorescence.
Chapter 2: Differential targeting of a mesophilic endoglucanase
21
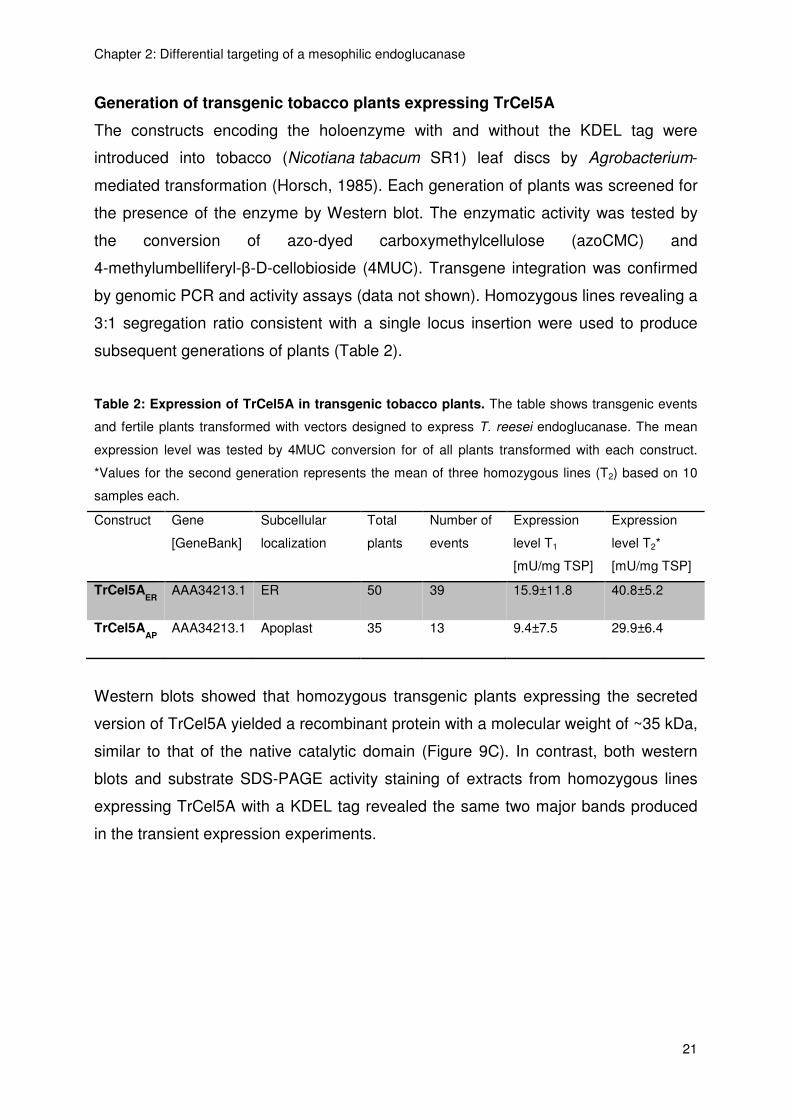
Generation of transgenic tobacco plants expressing TrCel5A
The constructs encoding the holoenzyme with and without the KDEL tag were
introduced into tobacco (Nicotiana tabacum SR1) leaf discs by Agrobacterium-
mediated transformation (Horsch, 1985). Each generation of plants was screened for
the presence of the enzyme by Western blot. The enzymatic activity was tested by
the conversion of azo-dyed carboxymethylcellulose (azoCMC) and
4-methylumbelliferyl-β-D-cellobioside (4MUC). Transgene integration was confirmed
by genomic PCR and activity assays (data not shown). Homozygous lines revealing a
3:1 segregation ratio consistent with a single locus insertion were used to produce
subsequent generations of plants (Table 2).
Table 2: Expression of TrCel5A in transgenic tobacco plants. The table shows transgenic events
and fertile plants transformed with vectors designed to express T. reesei endoglucanase. The mean
expression level was tested by 4MUC conversion for of all plants transformed with each construct.
*Values for the second generation represents the mean of three homozygous lines (T2) based on 10
samples each.
Construct Gene
[GeneBank]
Subcellular
localization
Total
plants
Number of
events
Expression
level T1
[mU/mg TSP]
Expression
level T2*
[mU/mg TSP]
TrCel5AER
AAA34213.1 ER 50 39 15.9±11.8 40.8±5.2
TrCel5AAP
AAA34213.1 Apoplast 35 13 9.4±7.5 29.9±6.4
Western blots showed that homozygous transgenic plants expressing the secreted
version of TrCel5A yielded a recombinant protein with a molecular weight of ~35 kDa,
similar to that of the native catalytic domain (Figure 9C). In contrast, both western
blots and substrate SDS-PAGE activity staining of extracts from homozygous lines
expressing TrCel5A with a KDEL tag revealed the same two major bands produced
in the transient expression experiments.

Chapter 2: Differential targeting of a mesophilic endoglucanase
22
Figure 9: Analysis of transgenic tobacco expressing TrCel5A. Amplification of genomic DNA
prepared from wild-type and transgenic plants, using primers pSS-5 and pSS-3 which anneal within
the CaMV 35SS expression cassette (A) and primers spanning a highly conserved region in the
chloroplast genome as a positive control (Demesure et al., 1995) (B). Lanes: M, GeneRuler™ 1-kb
ladder; 1,2 TrCel5AAP; 3,4 TrCel5AER; 5,6 SR1 wild-type; 7 H2O; 8 control plasmid DNA pTRAkc-
TrCel5A-ER. Western blot of recombinant TrCel5A from different TrCel5AAP lines (lanes 1-3) and
TrCel5AER lines (lanes 4-6) (C). Lanes were loaded with 10 µg TSP from transgenic plants (1-6), wild-
type plants (7) and IMAC purified PAL-His6. The MαHis5/GAMFcAP antibody system was used in (C).
All transgenic lines produced TrCel5A degradation products, but small amounts of the full enzyme
were detected in the TrCel5AER lines. The expression level of TrCel5A in transgenic plants was
determined by the conversion of 4MUC (D). TrCel5AER plants showed approximately 1.25-fold higher
activity than TrCel5AAP plants.
Recombinant protein expression levels in transgenic tobacco plants were determined
by measuring the activity of total soluble proteins (TSP) against the substrates
azoCMC and 4MUC. T2 plants expressing the secreted version of TrCel5A achieved
a specific enzyme activity of up to 1.5 U mg-1 TSP on azoCMC (data not shown) and
35 nmol 4MU min-1 mg-1 on 4MUC (Figure 9D), whereas the ER-localized version
achieved a specific enzyme activity of up to 2.1 U mg-1 on azoCMC and
46 nmol 4MU min-1 mg-1 on 4MUC (Figure 9D).

Chapter 2: Differential targeting of a mesophilic endoglucanase
23
Biochemical characteristics of TrCel5A produced in planta
Figure 10: Enzymatic activity of TrCel5A at different pH values and temperatures. TrCel5A
activity was determined using the azoCMC assay for pH values between 3.0 and 7.0 (A) and
temperatures in the range 20–70°C (B). The maximum activity measured for each system (at pH 4.8
and 55°C, respectively) was set to 100%. For panel A, buffer systems are used as indicated in the
figure. For panel B, acetate buffer (pH 4.8) was used throughout.
Plant-derived TrCel5A was extracted and purified by IMAC to determine its
biochemical characteristics, such as its activity on azoCMC in different buffer
systems within the pH range 3.0–7.0 and the temperature range 20–70°C (Figure
10). The optimal conditions were pH 4.8 and 55°C. Under these conditions, 50% of
the enzyme remained active for 90 min. There was no significant activity against
highly crystalline substrates such as Avicel, but there was significant activity against
heterogeneous substrates including lichenan and barley-β-glucan (Table 3).
Table 3: Hydrolytic activity of TrCel5A against different polymeric carbohydrates. The quantity
of reducing sugars released in the reaction was determined using a PAHBAH assay. Activity against
carboxymethylcellulose (CMC) was set as 100%. N.D: no detectable activity.
CMC β-Glucan Lichenan PASC Avicel Xylan
Relative
activity [%] 100±2 120±3 136±5 22±2 N.D. N.D.

Chapter 2: Differential targeting of a mesophilic endoglucanase
24
Subcellular localization of TrCel5A in tobacco plants
The localization of TrCel5A in the transgenic tobacco plants was determined by
immunostaining and fluorescence microscopy. Thin sections [10 µm] of the vascular
bundles of tobacco leaves were prepared using a Leica Cryostat Microtome.
Figure 11: Subcellular localization of recombinant TrCel5AER in transgenic tobacco cells. Plants
expressing TrCel5AER were immunostained with the MαKDEL primary antibody and GAMFCFITC as the
secondary antibody. Images were obtained by fluorescence microscopy. The SR1 wild-type control (A)
was compared with the TrCel5AER transgenic plants (B). Intracellular fluorescent signals were
detected in the TrCel5AER transgenic plants in some cells (B), whereas control plants did not show
significant fluorescent staining (A).
When the sections were stained with either RαCell and GARFCFITC or MαKDEL and
GAMFCFITC, strong green fluorescence indicated the presence of the recombinant
enzyme in in some cells of the transgenic plants but not the wild-type control.
Fluorescence was more easily detected in the TrCel5AAP tissue despite the higher
recombinant protein expression levels in the TrCel5AER plants (Figure 12). As
expected, MαKDEL only detected the enzyme carrying the KDEL tag (in the
TrCel5AER tissues) and intracellular localization was confirmed (Figure 11).

Chapter 2: Differential targeting of a mesophilic endoglucanase
25
Figure 12: Subcellular localization of recombinant TrCel5A in transgenic tobacco cells. Plants
expressing variants of TrCel5A targeted to the apoplast and ER were immunostained with the RαCell
primary antibody and GARFCFITC as the secondary antibody. Images were obtained by fluorescence
microscopy. The SR1 wild-type control (A) was compared with TrCel5AER (B) and TrCel5AAP (C)
transgenic plants. Intracellular fluorescent signals were detected in the TrCel5AER tissues, whereas a
peripheral signal in the cell walls was detected in the TrCel5AAP tissues. Control plants did not show
any significant fluorescent staining (A).
Growth characteristics of the transgenic tobacco plants
The constitutive expression of TrCel5A and its localization in the apoplast reduced
the growth of tobacco plants and significantly delayed their development. The stem
length and germination rate were both reduced (Figure 13 and Figure 14). The
expression of TrCel5A with a KDEL peptide for retention in the ER resulted in a much
less significant difference in growth and development (Figure 13 and Figure 14). For
both of the constructs, a few transgenic plants displayed a curly leaf phenotype, and
in the TrCel5AAP plants this was accompanied by necrotic areas on the leaf (Figure
13C).

Chapter 2: Differential targeting of a mesophilic endoglucanase
26
Figure 13: Transgenic tobacco plants expressing TrCel5A. Eight-week-old N. tabacum SR1 wild-
type plants were grown in soil under standard conditions (A & D). TrCel5AAP plants showed a
significant reduction in the growth and germination rate (E), whereas TrCel5AER plants showed a near
normal growth and development phenotype (F). Several transgenic plants displayed a curly leaf
phenotype (B & C) which in the case of TrCel5AAP plants included additional necrotic lesions (C).

Chapter 2: Differential targeting of a mesophilic endoglucanase
27
Chemical analysis of tobacco cell walls
The structural carbohydrate composition of plants expressing differentially-targeted
TrCel5A was determined in leaf material from six-week-old tobacco plants.
Destarched alcohol-insoluble residues (AIRs) were hydrolyzed with weak acid (Foster
et al. 2010) and the solubilized carbohydrates were measured as alditol acetate
derivatives by GC/MS (Albersheim et al., 1967). The crystalline cellulose content of
the tobacco leaf material was determined by Updegraff hydrolysis (Updegraff, 1969a)
and an anthrone cellulose assay. Wild-type plants were compared with transgenic
lines expressing TrCel5A in the apoplast and ER.
Figure 14: Phenotypic analysis of transgenic tobacco plants. The crystalline cellulose content and
stem length was analyzed in six-week-old wild-type and TrCel5A transgenic tobacco plants grown in
soil. Asterisks indicate a significant decrease (P ≤ 0.01) in the crystalline cellulose content for
TrCel5AAP compared to wild-type plants whereas TrCel5AER plants show no significant difference (A).
The comparison of stem length between TrCel5A transgenic and wild-type plants showed that
TrCel5AAP lines with the lowest cellulose levels also has the shortest stems (B).
Transgenic lines accumulating TrCel5A in the apoplast showed dramatic changes in
the content of crystalline cellulose, which was significantly lower than in wild-type
plants, whereas transgenic plants accumulating TrCel5A in the ER did not show such
differences (Figure 14A). Analysis of the matrix phase polysaccharides also revealed
dramatic differences in the composition of simple sugars (Table 4). The TrCel5AAP
lines contained less glucose and more xylose in the matrix phase polysaccharides,
whereas the TrCel5AER lines showed only a marginal difference from the wild type

Chapter 2: Differential targeting of a mesophilic endoglucanase
28
levels. The amount of galactose increased in all the transgenic lines. Arabinose and
rhamnose levels were lower in the TrCel5AAP lines, but there was no change in the
level of mannose.
Table 4: Compositional analysis of matrix-phase polysaccharides in different transgenic
tobacco lines. Relative values [%] for sugar distribution in matrix polysaccharide fraction. Data
analyzed by two-sided Student’s t-test; * indicates a significant P < 0.01; + indicates a significant P <
0.05
Line Arabinose Glucose Galactose Mannose Rhamnose Xylose
Wild-type 20.41
±2.61
23.60
±8.42
16,85
±2.14
6.12
±1.10
17.90
±2.28
13.96
±1.81
TrCel5AAP-1 14.95*
±1.36
10.81 *
±1.27
32.71*
±6.71
5.31
±1.69
14.42+
±3.68
21.79*
±3.44
TrCel5AAP-2 16.63+
±1.86
13.90 *
±4.68
34.08*
±4.68
5.01
±1.18
13.43*
±1.69
16.94*
±1.93
TrCel5AER-1 20.72
±1.65
17.93+
±2.46
27.70*
±2.93
4.99+
±0.74
16.92
±1.55
13.91
±1.35
TrCel5AER-2 16.11*
±1.09
19. 74+
±2.59
33.11*
±2.37
4.73+
±1.56
13.36*
±1.13
11.04*
±1.10
TrCel5AER-3 18.96
±1.93
20.54+
±3.44
27.22*
±3.51
4.50*
±1.42
16.21
±1.85
11.97*
±1.19
Two of three TrCel5AER lines contained lower levels of xylose, and all three lines
contained slightly lower levels of mannose compared to wild-type plants. There were
no significant changes in the levels of arabinose and rhamnose in two of the
TrCel5AER but there was a significant reduction for both sugars in the remaining line.
The biochemical changes in the cell wall sugars of TrCel5AAP lines were associated
with the growth reduction phenotype, but as discussed above there was no
significant visible change in the TrCel5AER lines despite the modulation of sugar
levels.

Chapter 2: Differential targeting of a mesophilic endoglucanase
29
Hydrolysis of leafy biomass from transgenic tobacco
To determine the rate at which transgenic leaf material was hydrolyzed by
commercially-available cellulase preparations, tobacco plants of both varieties were
selected, with similar levels of TSP and TrCel5A activity (based on the release of
4MU) and compared to wild-type controls. Following enzymatic hydrolysis, the
crystalline cellulose remaining in the insoluble material was quantified.
Figure 15: Enzymatic hydrolysis of transgenic tobacco leaves. Leaf tissue from transgenic
tobacco lines TrCel5AAP and TrCel5AER, both expressing ~30 mU/mg TSP (based on 4MUC
conversion) were hydrolyzed with the appropriate amount of the commercial cellulose preparation
CTec2 (Zhang et al., 2011). Digestion was carried out at 55°C with constant shaking at 1000 rpm. The
TrCel5AAP leaves underwent more rapid hydrolysis than the TrCel5AER and wild-type leaves, but
eventually reached the same level of total cellulose hydrolysis.
The TrCel5AER transgenic leaves did not show any difference in the rate of hydrolysis
compared to wild-type leaves, regardless of the TrCel5A expression level. In
contrast, the TrCel5AP transgenic leaves were hydrolyzed more rapidly, although
ultimately the relative amount of solubilized crystalline cellulose was the same as that
in TrCel5AER and wild-type leaves (Figure 15).

Chapter 2: Differential targeting of a mesophilic endoglucanase
30
Discussion
Current research in the area of biofuels aims to achieve the economically feasible
production of cellulolytic enzymes for the conversion of lignocellulosic biomass into
soluble sugars. Plants offer a relatively inexpensive production platform for industrial
proteins (Fischer et al., 2004; Streatfield, 2007) and therefore plants have been used
to express a number of biomass-degrading enzymes (Table 1 and reviewed in Sainz,
2011; Taylor et al., 2008).
Several studies have demonstrated the potential of in planta recombinant cellulases
by achieving the accumulation of functional enzymes at high levels (Brunecky et al.,
2011; Hood et al., 2012; Petersen and Bock, 2011). However, several drawbacks are
associated with constitutive cellulase expression, including growth inhibition, changes
in leaf morphology, loss of cell wall integrity and reduced stress tolerance (Abdeev et
al., 2004; Petersen and Bock, 2011; Taylor et al., 2008).
Plants express endogenous GHs that play an important yet barely understood role in
cell wall synthesis (Lopez-Casado et al., 2008). Both the overexpression and
silencing of these GHs can have a negative impact on growth and cell wall
development (Maloney and Mansfield, 2010; Takahashi et al., 2009).
These issues have been addressed by targeting cell wall-degrading enzymes to
different subcellular compartments, which can increase the overall enzyme yields
(Buanafina et al., 2010; Dai et al., 2005; Mahadevan et al., 2011) as well as keeping
potentially harmful enzymes away from the cell wall (Kim et al., 2010; Ziegelhoffer et
al., 2009). We therefore compared transgenic plants in which the T. reesei mesophilic
endoglucanase TrCel5A was either secreted to the apoplast or retained in the ER.
Already in the transient studies the localization of TrCel5A has been monitored with
mCherry fusions of the enzyme, proving the correct targeting for TrCel5A-mCherryAP
in the apoplastic space and TrCel5A-mCherryER inside the cells. For transgenic
plants TrCel5A localization was confirmed by immunostaining of tissue thin sections,
which approved the result of the transient study, though the staining did not reflect
the expression level of the enzyme. Further studies using embedding techniques
before sectioning might lead to improved results.
Transient expression revealed that TrCel5A underwent proteolytic cleavage within
the linker region, probably caused by papain-like proteases in the apoplast (Beyene
et al., 2006) that are known to degrade T. reesei cellulases (van Tilbeurgh et al.,
1986). Adding a specific inhibitor of this protease class (E-64) during extraction

Chapter 2: Differential targeting of a mesophilic endoglucanase
31
reduced the amount of degradation, albeit not completely. However, the truncated
enzyme remained active against soluble substrates such as azoCMC and other
β-glucans with a β1,4-linked glucose backbone, whereas carbohydrates with different
linkages remained unaffected (Nakazawa et al., 2008). The apoplast targeting
strategy yielded the truncated and active form of TrCel5A alone, whereas retention in
the ER yielded a mixture of the truncated and full-length enzyme, although the latter
was a minority product. TrCel5A activity in the transgenic plants was determined by
measuring the conversion of 4MUC into 4MU, resulting in a maximum of
33 nmol 4MU min-1 mg-1 in the TrCel5AAP plants and 46 nmol 4MU min-1 mg-1 in the
TrCel5AER plants. This probably reflects the relative paucity in the number and
quantity of proteases in the ER lumen compared to the apoplast (Beyene et al.,
2006).
The optimal reaction conditions for the recombinant TrCel5A were found to be 55°C
and pH 4.8, highlighting the mesophilic origin of the enzyme. However, the enzyme
also showed remarkable residual activity under physiological conditions typical for
tobacco plants (20–30°C, pH 5.0) suggesting it potentially interferes with the
synthesis of cell wall cellulose during normal plant growth and development. Thus it
was not surprising that the TrCel5AAP transgenic plants grew more slowly and
germinated less frequently, whereas there was no significant difference between the
TrCel5AER and wild-type plants. Despite the differences between the transgenic
plants accumulating TrCel5A in different compartments, there was no significant
change in the overall amount of crystalline cellulose for TrCel5AER plants compared
to the wild-type lines. Cell wall cellulose is synthesized by cellulose synthase
complexes in the plasma membrane. After polymerization, the glucan chains are
extruded into the extracellular space (Endler and Persson, 2011) and therefore
endoglucanase expression targeted to the apoplast has the potential to interfere with
endogenous cellulose synthesis. This corresponds to our observation that the
content of crystalline cellulose is reduced in TrCel5AAP plants but not in TrCel5AER
plants. However the role of endoglucanases during the synthesis of cell wall cellulose
appears far more intricate than previously reported. Plant endoglucanases such as
KORRIGAN are necessary for the accumulation of sufficient cellulose and its
absence leads to severe deficit (Molhoj et al., 2001; Takahashi et al., 2009). Recent
studies have shown that the overexpression of KORRIGAN and other recombinant

Chapter 2: Differential targeting of a mesophilic endoglucanase
32
endoglucanases can also reduce the crystallinity of cellulose (Brunecky et al., 2011;
Maloney and Mansfield).
The analysis of matrix polysaccharide composition of transgenic and wild-type
tobacco plants revealed a significant reduction in the relative values of glucose,
arabinose and rhamnose in the TrCel5AAP lines, but much higher levels of galactose
and xylose. Plant GHs have been shown to remodel cell wall polysaccharides in the
apoplastic space and during the transport of polysaccharide precursors through the
Golgi apparatus (Minic, 2008). TrCel5A is a strict endoglucanase and is only known
to cleave β1,4-linkages between glucopyranose units, while showing no activity
against complex mixed glucans (Nakazawa et al., 2008). The degradation is
therefore likely to take place at an earlier stage of XyG synthesis, when the glucan
chains are still undecorated. The higher levels of galactose and xylose would directly
reflect the loss of glucose.
The relative level of glucose was also lower in the TrCel5AER lines, albeit not to the
extent seen in the TrCel5AAP lines. Galactose levels were also higher, as in the
TrCel5AAP plants, but xylose levels were lower in two of three tested lines. All
TrCel5AAP plants had lower levels of mannose and one line also showed lower levels
of arabinose and rhamnose. These changes in the matrix polysaccharide
composition for TrCel5AER lines were not associated with a visible phenotype in
contrast to the observed changes in the TrCel5AAP plants.
ER localization was used to prevent the enzyme from entering the Golgi apparatus
where most hemicellulose and pectin synthesis takes place (Moore and Staehelin,
1988; Staehelin and Moore, 1995). However, recent studies have shown that ER-
localized enzymes can still affect matrix polymer synthesis. The deferuloylation of
arabinoxylans in tall fescue can be achieved using a fungal ferulic acid esterase A
(FAEA). Even if the detected values for Golgi apparatus localization - the place of the
synthesis - were higher, significant cleavage was observed for ER retardation of the
enzyme (Buanafina et al., 2010). This may reflect the promiscuous behavior of
KDEL-tagged proteins, which can in some cases enter the Golgi network from where
they are later retrieved (Pelham, 1990).
XyG is the most abundant hemicellulose in the cell walls of dicotyledonous plants,
representing up to 30% by weight (Scheller and Ulvskov, 2010). In most dicots the
main form is fucogalacto-XyG, but arabinogalacto-XyG is more abundant in
solanaceous plants and monocots (Pauly et al., 2013; Ring and Selvendran, 1981).

Chapter 2: Differential targeting of a mesophilic endoglucanase
33
In fucogalacto-XyG, galactose is transferred to the xylosyl residue and then
substituted with fucose, whereas in arabinogalacto-XyG galactose or arabinose can
be transferred to the xylosyl residue (Pauly et al., 2013). The scarcity of glucose
could then lead to the reduced levels of xylose in the matrix polysaccharides of
TrCel5AER plants. In contrast, xylose levels were normal in the TrCel5AAP lines.
Interestingly, galactose levels were elevated in all the plants. However, despite these
differences in biochemistry, there was no significant morphological or developmental
change in the TrCel5AER plants, which is consistent with previous observations of
mutant plants with changes in the XyG side chain composition (Jensen et al.). All
biochemical ana-lyses were carried out with plants grown in the greenhouse without
the addition of fertilizer. Therefore it is possible that the addition could reduce the
metabolic stress in the transgenic plants and diminish the observed changes.
However, if the same lines will be tested in field experiments, this stress is more likely
increased.
The remaining amount of crystalline cellulose was measured in the tobacco leaves
after digestion with a commercial cellulase preparation, revealing a two-fold greater
reduction in TrCel5AAP plants after 8 h compared to wild-type and TrCel5AER plants.
After 65 h however, all three lines contained the same relative amounts of residual
cellulose, showing that degradation was accelerated but not enhanced overall in the
TrCel5AAP plants. Quantitative effects were ruled out by carefully selecting transgenic
plants of both lines with similar overall enzyme expression levels, and in any case we
found no overall difference between the transgenic and wild-type lines.
Endoglucanases produced in planta have previously been shown to make the plant
cell wall less recalcitrant to hydrolysis (Brunecky et al., 2011). Accordingly, our
results confirmed a significant reduction in the levels of crystalline cellulose in
TrCel5AAP plants followed by accelerated hydrolysis.
The altered matrix polysaccharide fraction of the TrCel5AER plants appeared to have
little impact on the enzymatic hydrolysis of leaf material, which was similar to the
profile in wild-type leaves. This may reflect the relatively simple assay substrate,
which does not consider the impact of lignin due to its low abundance in leaves
compared to stems and roots (Van Dyk and Pletschke, 2012). The modification of cell
wall hemicellulose using hydrolytic enzymes can promote lignocellulose degradation
by reducing the amount of crosslinking between polymers (Buanafina et al., 2010;
Pogorelko et al., 2011; Zhang et al., 2011).

Chapter 2: Differential targeting of a mesophilic endoglucanase
34
In summary, the heterologous expression of the mesophilic endoglucanase TrCel5A
had a significant impact on the composition of plant cell wall polysaccharides,
although the precise changes were dependent on the subcellular localization of the
enzyme. Secretion to the apoplast reduced the levels of crystalline cellulose in the
cell wall and had a significant impact on plant growth and development, whereas
targeting to the ER resulted in compositional changes among the matrix-phase
polysaccharides but did not cause any adverse morphological or developmental
phenotypes.

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
35
Chapter 3: Ethanol-inducible endoglucanase expression in
tobacco
The comparison of inducible and constitutive expression of TrCel5A in transgenic tobacco has been
published in:
Klose H, Günl M, Usadel B, Fischer R, Commandeur U. (2013) Ethanol inducible
expression of a mesophilic cellulase avoids adverse effects on plant development.
Biotechnology for Biofuels, 6:53.
The high-level constitutive expression of GHs can affect the structure and
composition of plant cell walls by altering the quality and/or quantity of cellulose
(Chapter 2). The regulation of endoglucanase expression using inducible promoters
is therefore a promising approach to avoid the drawbacks caused by cellulase
accumulation during vegetative growth. Many different inducible expression systems
have been developed for plants (Bortesi et al., 2012; Bruce et al., 2000; Okuzaki et
al., 2011) and the Aspergillus nidulans alc platform has been widely used (Caddick et
al., 1998; Roslan et al., 2001; Tomsett et al., 2004). This platform comprises two
elements, the coding sequence for the transcription factor AlcR (controlled by the
CaMV 35S promoter), and the alcAmin 35S promoter, in which two AlcR binding sites
are joined to the CaMV 35S minimal promoter (Caddick et al., 1998). This promoter
shows negligible basal expression but strong induction ratio in most plants in
response to ethanol (Caddick et al., 1998; Roslan et al., 2001; Salter et al., 1998).
We used the alc platform to express the secreted form the TrCel5A endoglucanase
from T. reesei resulting in its accumulation in the apoplast of tobacco cells.
Results
Generation of transgenic tobacco plants expressing inducible TrCel5A
The cDNA encoding TrCel5A (lacking the native signal peptide sequence) was
placed under the control of the ethanol-inducible alcAmin35S promoter (Caddick et
al., 1998). The construct included a plant-codon-optimized signal peptide derived
from the heavy chain of the murine monoclonal antibody mAb24 for secretion to the
apoplast, and a C-terminal His6 tag for detection and purification. This construct was
then introduced into N. tabacum SR1 leaf discs by Agrobacterium-mediated

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
36
transformation (Horsch, 1985). Integration of the transgene was confirmed by
genomic PCR (Figure 16). After induction, each generation of plants was screened
for the presence and activity of the enzyme by Western blot and activity staining on
azoCMC and 4MUC gels. Positive T1 lines showing 3:1 transgene segregation
consistent with a single locus were selfed to produce the subsequent homozygous
generations.
Figure 16: Generation and analysis of transgenic tobacco expressing TrCel5A under the
control of an alcohol-inducible promoter. Schematic presentation of the plant inducible expression
cassette (A). The chimeric promoter (alcAmin35S) comprising the minimal CaMV35S promoter (±31 to
+5) fused to the upstream promoter sequences of alcA (Caddick et al., 1998) is shown in dark blue.
The terminator signal (pA35S) is shown in light blue. The chalcone synthase (CHS) gene 5'-UTR and
His6 tag sequence are indicated in navy blue. A plant-codon-optimized leader peptide (LPH) derived
from the heavy chain of the murine mAb24 is shown in green. Genomic DNA prepared from wild-type
and transgenic plants was tested by PCR using primers binding within the trcel5a gene (cel5a-seq)
and the pA35S terminator sequence (pss-3) (B) and primers binding in a highly-conserved region in
the chloroplast genome (C) (Demesure et al., 1995). Lanes: M, GeneRuler™ 1kb; 1-4,
alcR::TrCel5AAP lines; 5,6 SR1 wild-type; 7 water; 8 control plasmid pTRAkc-alcR-TrCel5A-AP.

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
37
Inducible TrCel5A expression in tobacco plants
The impact of alcohol induction on plant growth and TrCel5A expression was tested
on T2 alcR::TrCel5A transgenic plants and wild-type controls. The induction of
TrCel5A expression with 2% (v/v) ethanol led after 24 h to a maximum activity of
0.4 U mg–1 TSP on azoCMC and 5 nmol 4MU min-1 mg-1 (Figure 17). Further analysis
showed that significant expression could be achieved by inducing with 0.1% (v/v)
ethanol, but optimal expression levels required 2% (v/v) ethanol (Figure 17B).
TrCel5A expression did not increase when plants were induced with 5% (v/v) ethanol,
and higher concentrations were detrimental to both wild-type and transgenic plants,
resulting in physiological stress symptoms such as leaf yellowing. Significant basal
expression in the absence of ethanol was not detected.
Figure 17: Inducible TrCel5A expression in tobacco plants. Time course of ethanol induction in
six-week-old soil-grown T2 plants irrigated with ethanol solutions. Plants were induced at t0 by
irrigating with 2% (v/v) ethanol, and cellulase activity was monitored over a 96-h time course (A).
Transgenic lines were monitored for cellulase activity 24 h after watering with different ethanol
solutions. Values represent the mean of three plants per independent transgenic line (B). The effect of
sequential ethanol induction (C). Six-week-old soil-grown plants from the homozygous line
alcR::TrCel5A-3 were induced with 2% (v/v) ethanol at t0 and t48 (asterisks). Cellulase activity was
monitored over a six-day time course. Each data point represents the mean of three replicates. Wild-
type plants were monitored in parallel, and no activity was observed throughout the experiment.

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
38
To determine the time course of induction, TrCel5A activity was monitored for 4 days
following induction with 2% (v/v) ethanol (Figure 17A). Peak activity was observed
48 h after induction (7 nmol 4MU min-1 mg-1) followed by a significant decline.
Continuous or sequential induction has been shown to boost transgene expression
levels (Roslan et al., 2001), so a second induction step was introduced 48 h after the
first induction. This increased the enzyme activity significantly, to
27 nmol 4MU min-1 mg-1 (Figure 17C). This was lower than the enzyme activity
achieved in constitutive TrCel5A plants, but the two-step induction nevertheless
achieved an expression level within the same order of magnitude as the constitutive
promoter.
Growth characteristics and histology
The inducible expression of the secreted form of TrCel5A had no impact on the
growth or development of the transgenic tobacco plants compared to wild-type
controls (Figure 18). Plants with constitutive expression and secretion of TrCel5A as
described in chapter A, exhibited a significantly reduced growth and a delayed
development. The mature stems of the transgenic lines were 21–36% shorter than
controls and delayed flowering. Cross-sections of stems stained with calcofluor-white
showed no significant differences in cell structure between alcR::TrCel5A and wild-
type plants, whereas there was a slight increase in the number of small vessels in
TrCel5AAP plants (Figure 18).

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
39
Figure 18: Phenotype of transgenic tobacco plants. Cross sections of wild-type (A), TrCel5AAP (B)
and alcR::TrCel5AAP (C) stems (10x magnification) stained with calcofluor white and visualized under
UV light. There were no significant differences other than a marginal increase in the number of small
vessels (white arrow) in TrCel5AAP plants. Scale bars = 50 µm. Morphological phenotype of wild-type
(D), TrCel5AAP (E) and alcR::TrCel5AAP (F) plants grown under photoautotrophic conditions in soil.
TrCel5AAP plants were significantly shorter and had a lower germination rate compared to wild-type
and alcR::TrCel5AAP plants.

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
40
Chemical analysis of tobacco cell walls in alcR::TrCel5AAP plants
The impact of inducible TrCel5A expression on the structure of tobacco cell walls was
determined by measuring the amount of cellulose, alcohol-insoluble residues (AIRs)
and crystalline cellulose as described in Chapter 2.
Figure 19: Crystalline cellulose content in transgenic tobacco plants expressing TrCel5A. The
cellulose content was measured in six-week-old soil-grown transgenic tobacco plants transformed with
constitutive and inducible expression constructs, compared to wild-type controls (A). Values represent
the mean ± standard error of 10 plants per tobacco line (transgenic or wild-type). Asterisks indicate a
significant difference compared to the wild-type (P ≤ 0.01). Cellulase activity and cellulose content was
also measured in alcR::TrCel5AAP-3 plants after double induction with ethanol (B). A relative value of
100% cellulase activity represents the conversion of 27 nmol 4MU min-1 mg-1, whereas a value of
100% for cellulose content represents 140 µg glucose per mg alcohol insoluble residue (AIR). For all
panels, values represent the mean of three plants per independent transgenic line. Error bars show
standard deviations normalized against the wild-type controls.
Here wild-type plants were compared with three constitutive TrCel5AAP lines and two
inducible alcR::TrCel5AAP lines. As stated earlier, the constitutive transgenic lines
contained significantly lower levels of crystalline cellulose than the wild type plants
and the TrCel5AAP line with the highest enzyme activity also showed the greatest
reduction in crystalline cellulose (Figure 19A). In contrast, there was no difference in
crystalline cellulose levels between the inducible transgenic plants and wild-type
controls, even up to 120 h after the induction of cellulase activity (Figure 19B).

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
41
Discussion
As discussed in Chapter 2, the precise role of GHs in the development of plant cell
walls is not yet understood. The depletion and overexpression of GHs can each
affect plant growth and the formation of cell walls, and this represents a major
challenge when recombinant GHs are expressed in plants for biomass degradation
(Maloney and Mansfield, 2010; Park et al., 2003; Takahashi et al., 2009). In several
studies, the constitutive expression of biomass-degrading enzymes has been shown
to affect plant health, e.g. inhibiting growth, altering leaf morphology, reducing stress
tolerance and affecting the structure of the cell wall (Abdeev et al., 2003; Brunecky et
al., 2011; Petersen and Bock, 2011; Taylor et al., 2008). Therefore, alternative
strategies are required for the production of GHs such as cellulases in plants, without
affecting normal growth and development. These include sequestration into
subcellular compartments (Chapter 2) or the expression of enzymes that are inactive
under physiological conditions (Dai et al., 2000). However, the inducible expression
of cellulases could allow the expression of functional enzymes just prior to harvest,
thus avoiding detrimental effects during vegetative growth.
Ethanol-inducible expression cassettes are widely used in plant biotechnology (Ait-ali
et al., 2003), and here we combined the alcAmin 35S promoter, a fusion of the AlcR
binding site with the minimal CaMV 35S promoter (Caddick et al., 1998), to achieve
the inducible expression of TrCel5A, and used the doubled CaMV 35SS promoter as
a comparator for constitutive expression. Both promoters were used to express the
secreted form of the enzyme, which was shown to accelerate biomass degradation in
Chapter 2.
The one-step induction procedure with 2% (v/v) ethanol had no significant impact on
the health of wild-type tobacco plants and achieved a significant level of cellulase
activity, but after 24 h this was 4–6 times lower than achieved by constitutive
expression. Different induction times and ethanol concentrations were tested, but 2%
ethanol was found the optimal concentration for induction, in agreement with
previous reports (Roslan et al., 2001). TrCel5A activity peaked at 7 nmol 4MU min-1
mg-1 48 h after induction, compared to 35 nmol 4MU min-1 mg-1 achieved by
constitutive expression. We introduced a second induction step after 48 h because
previous reports have shown this can boost transgene expression (Roslan et al.,
2001). Accordingly, this boosted the maximum enzyme activity to
27 nmol 4MU min-1 mg-1 5 days after the first induction step. Although still lower than

Chapter 3: Ethanol-inducible endoglucanase expression in tobacco
42
the activity achieved by constitutive expression, it was in the same order of
magnitude (and was obtained without a negative impact on plant growth and
development) and was comparable to the levels achieved by other researchers
(Brunecky et al., 2011; Dai et al., 2005).
There was little difference in the crystalline cellulose content of the alcR::TrCel5AAP
transgenic plants before and after induction, suggesting that the endoglucanase only
reduces the crystallinity of the cell wall when cellulose is synthesized (evidently this
occurs during constitutive expression). This theory is also supported by the lower
endoglucanase activity of TrCel5A against more crystalline substrates, such as Avicel
and PASC (Table3). The truncation of the enzyme, which removes the CBM, could
also contribute to the lower affinity for crystalline substrates (Boraston et al., 2004).
Alternatively, the efficient hydrolysis of crystalline cellulose may require more than
one cellulolytic activity (Henrissat et al., 1985). These data suggest that the absence
of the CBM might also be advantageous for systems based on the constitutive
expression of TrCel5A. Potentially, the expression of CBMs alone could disrupt cell
wall architecture and interfere with plant development (Obembe et al., 2007), or act
as a microbe-associated molecular patterns (MAMPs) to trigger a defense
mechanism, such as the accumulation of sesquiterpene cyclase (Brotman et al.,
2008; Gaulin et al., 2006).
In summary, the constitutive expression of the mesophilic endoglucanase TrCel5A
has a negative impact on plant growth and development, but this can be avoided by
using the ethanol-inducible alcAmin35S system. Furthermore, double induction
resulted in the accumulation of more recombinant cellulase, approaching the levels
achieved by constitutive expression. We propose that the inducible expression of
biomass degrading enzymes in plants is beneficial because it allows controlled
applications such as expression during specific developmental stages or defined
expression levels. This system also uses an economical inducer and a comparatively
simple application process suitable for large-scale biomass degradation.

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
43
Chapter 4: Expression of a hyperthermophilic
endoglucanase in tobacco
Results presented in this chapter have been published in:
Klose H, Röder J, Girfoglio M, Fischer R, Commandeur U (2012) Hyperthermophilic
endoglucanase for in planta lignocellulose conversion. Biotechnology for Biofuels 5: 63.
Cellulases have been produced in a number of plant species (Dai et al., 2005;
Ziegelhoffer et al., 1999) and the isolated recombinant enzymes have been shown to
digest cell wall preparations (Kawazu et al., 1996; Montalvo-Rodriguez et al., 2000).
As discussed, the constitutive expression of mesophilic cellulases can inhibit the
growth of tobacco plants and generate abnormal morphological phenotypes reflecting
changes in the composition of the cell walls (Brunecky et al., 2011; Taylor et al.,
2008). Different strategies have been used in an attempt to avoid direct contact
between cellulases and cell wall polysaccharides during vegetative growth, including
targeting to intracellular compartments (chapter 2) and inducible expression (chapter
3). The degradation of plant cell walls during normal growth can also be avoided by
the expression of thermophilic or hyperthermophilic cellulases with extreme
temperature optima beyond the physiological range for mesophilic plantgrowth
(Montalvo-Rodriguez et al., 2000; Xue et al., 2003).
Extremophiles represent an ideal source for enzymes that can tolerate high
temperatures, extreme pH environments and high salt concentrations (Turner et al.,
2007; Unsworth et al., 2007). They may therefore provide thermophilic cellulases that
are compatible with standard pretreatment technologies for the conversion of
lignocellulose into fermentable sugars. For example, A. cellulolyticus endoglucanase
E1 has been expressed in transgenic tobacco plants that were subsequently
processed by AFEX pretreatment, and approximately 30% of the enzyme activity was
retained after treatment (Teymouri et al., 2004). Similarly, ionic liquids are efficient
lignocellulose solvents that can be combined with enzymatic hydrolysis, but the
enzymes must retain their activity in highly concentrated ionic solutions with a water
content of only 10–20% (Zavrel et al., 2009).
The genome of the hyperthermophilic archaeon Sulfolobus solfataricus encodes
three different endoglucanases named SSO1354, SSO1949 and SSO2534 (She et
al., 2001) that have already been shown to perform efficiently under harsh physical

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
44
and chemical conditions (Girfoglio et al., 2012; Huang et al., 2005; Limauro et al.,
2001; Maurelli et al., 2008). The transient expression of SSO1354 has already been
described in an earlier study (Röder, 2010). Here, stable SSO1354 expression in
N. tabacum was achieved and the ability of the enzyme to convert plant biomass into
fermentable sugars under different pretreatment conditions was determined, offering
a promising new approach for the bioconversion of lignocellulose in planta.
Results
Expression and downstream purification of SSO1354 in tobacco plants
The sso1354 gene was amplified from S. solfataricus genomic DNA without the
putative leader peptide as described elsewhere (Girfoglio et al., 2012). Ten potential
N-glycosylation sites (matching the consensus Asn X Ser/Thr) were identified using
the NetNGlyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc). The amplified
sequences were flanked by an N-terminal His6 tag and an optional C-terminal KDEL
sequence for ER retention, depending on the primers. The products were transferred
to the pTRAkc expression vector, which included an expression cassette containing
the Cauliflower mosaic virus (CaMV) double 35S promoter (P35SS), the 5'-UTR
translational enhancer from the Petroselinum hortense chalcone synthase gene and
a plant-codon-optimized LPH signal peptide for secretion to the apoplast (Figure 20).
Figure 20: Schematic representation of the SSO1354 expression cassettes including an
N-terminal His6 tag. The double CaMV promoter (P35SS) and terminator (pA35S) are shown in light
blue, the chalcone synthase 5'-UTR (CHS) in dark blue and the codon-optimized leader peptide from
the murine antibody mAb24 (LPH) is shown in green. The His6 and KDEL coding sequences are
indicated in red. LPH facilitates the secretion of the recombinant protein to the apoplast (A). An
additional C-terminal KDEL signal retains the protein in the ER lumen (B).

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
45
The expression vectors were introduced into A. tumefaciens strain GV3101 and
infiltrated into tobacco leaves. The infiltrated leaves were harvested 4 days later and
the TSP was extracted for analysis by SDS-PAGE, activity staining and Western blot.
Previous transient expression tests showed that enzyme variants containing a
C-terminal His6 tag could not be detected in the infiltrated plants (Röder, 2010).
However constructs carrying an N-terminal His6 tag either with or without the KDEL
sequence could be detected by means of Western blot. The ER-localized enzyme
showed the highest activity against azoCMC per gram leaf material and was selected
for expression in transgenic plants.
Figure 21: Substrate SDS-PAGE in the presence of 0.15% (w/v) CMC and extracts from different
SSO1354 transgenic plants. Lanes 1-12, T0 transgenic plants; lane 13, wild-type tobacco plants; lane
14, culture supernatant from K. lactis expressing SSO1354. Samples were incubated in 50 mM
sodium acetate buffer (pH 4.8) at 90°C for 15 min.
T0 transgenic plants expressing the ER-tagged enzyme were transferred to soil and
maintained in the greenhouse. The leaves were screened for enzyme activity using
the substrate SDS-PAGE method (Figure 21), and T1 seeds from positive candidates
were used to produce the next generation of plants. The leaves from the T1 plants
were tested using the substrate 4MUC to determine the activity of the recombinant
SSO1354 protein, which was comparably high, ranging from 92 to 488 nmol 4MU
min-1 mg-1 in the different transgenic plants (Figure 22). There was no difference in
morphology or growth behavior between the transgenic plants and wild-type controls
(Figure 22) and no differences in the biochemical composition of matrix-phase
polysaccharides or the content of crystalline cellulose (data not shown).

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
46
Figure 22: Tobacco plants expressing active SSO1354. SSO1354 activity in transgenic tobacco
lines (F1 to F6) based on the conversion of 4MUC (A). Values represent the mean of three in different
transgenic plants. Phenotypes of transgenic tobacco plant expressing SSO1354 (B) compared to
wild-type controls (C). Both lines were grown in soil under greenhouse conditions, and pictures were
taken six weeks after seeding. Monitoring different purification steps following SSO1354 expression in
tobacco with Western blot (D) and zymography (E) in the presence of 0.15 % (w/v) CMC. Lanes
contain 1 µg of purified SSO1354. Antibody system used in (D) is MαHis5 /GAMFcAP. The multiple
bands may represent alternative forms of N-glycosylation.
The recombinant SSO1354 was partially purified from crude plant extracts by thermal
precipitation. Heating to 85°C for 10 min caused 95% of the host cell proteins to
precipitate while SSO1354 remained soluble and active. Reducing sugars in the
extract, which could potentially interfere with subsequent enzyme activity assays,
were removed by passing the remaining extract through a PD10 desalting column.

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
47
Biochemical properties and catalytic activity of plant-derived SSO1354
The substrate profile of SSO1354 was determined by testing the enzyme against a
panel of β-glucans. The p-hydroxybenzoic acid hydrazide (PAHBAH) assay was used
to measure the release of reducing sugars, showing that SSO1354 was active
against CMC (arbitrary value set at 100% activity) and also barley β-glucan (136%
relative activity) and lichenan (130% relative activity), but not against pachyman
(Figure 23D).
Figure 23: Biochemical characterization of plant-derived SSO1354. The influence of temperature
on SSO1354 activity was measured using the azoCMC assay at 60–99°C (A). The maximum activity
measured at 90°C was set at 100%. The influence of buffer and pH on SSO1354 activity was
measured using the azoCMC assay at pH values between 2.0 and 7.0 (B). The maximum activity
measured at pH 4.5 was set at 100%. The influence of solvents and different salt concentrations on
SSO1354 activity (C) was measured using the azoCMC assay at optimal conditions (90°C, pH 4.5) by
adding different concentrations of salts and solvents. SSO1354 activity in the absence of salts and
solvents was set at 100%. The activity of SSO1354 was measured on different substrates (D). The
PAHBAH assay was used to determine the release of reducing sugars, and the amount reduced from
CMC was set at 100%.

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
48
The enzyme showed no significant activity against Avicel or oat xylan. This
suggested that SSO1354 has a β1,4-hydrolytic mode of action and is only active on
soluble substrates.
Further analysis using the azoCMC assay showed that SSO1354 has an optimum
temperature at 90°C, although it retains 79% residual activity at 99°C. The activity
was reduced substantially at lower temperatures, falling to ~30% activity at 70°C
(Figure 23A). The optimal pH was 4.5, although the enzyme retained 58% residual
activity at pH 2.7 and 62% residual activity at pH 6.5 (Figure 23B). There was only
minimal activity below pH 2.2 and above pH 7.0. Under optimal conditions (90°C, pH
4.5), the enzyme retained 50% of its activity after incubation for 100 min. SSO1354
also tolerated high salt concentrations, with 49.5% residual activity in the presence of
2.4 M NaCl, and 46.5% residual activity in the presence of 2.4 M CaCl2 (Figure 23C).
SSO1354 activity was inhibited by 15% (v/v) solutions of both protic solvents
(capable of donating protons) such as methanol and ethanol, and aprotic solvents
such as acetone and dimethylsulfoxide (DMSO). The enzyme retained 79% residual
activity in the presence of methanol, 75% in the presence of DMSO, 51% in the
presence of acetone and 48% in the presence of ethanol. Increasing the solvent
concentration to 30% (v/v) exacerbated the effect, with methanol and acetone
causing near-complete inhibition, DMSO leaving a residual activity of 14% and
ethanol leaving a residual activity of 16%.

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
49
SSO1354 activity in ionic liquids
As discussed above, SSO1354 cannot hydrolyze crystalline cellulose or cell wall
polysaccharides efficiently in aqueous solutions, but concentrated solutions (80–90%
v/v) of ionic liquids such as 1-ethyl-3-methylimidazolium acetate (EMIM-Ac) and 1,3
dimethylimidazolium dimethylphosphate (MMIM-DMP) can dissolve crystalline
cellulose substrates such as Avicel and cell wall polysaccharides, making them more
accessible to hydrolytic enzymes.
We therefore tested the activity of SSO1354 on Avicel that had been dissolved in
80% (v/v) EMIM-Ac and MMIM-DMP, by measuring the release of reducing sugars
and subtracting the quantity of sugars released by spontaneous hydrolysis in the
absence of the enzyme. The normalized release rates were 5.5 mg l-1 h-1 in EMIM-Ac
and 3.5 mg l-1 h-1 in MMIM-DMP (Figure 24). We also found that cell wall
polysaccharides dissolved in EMIM-Ac and MMIM-DMP were hydrolyzed by
SSO1354 with release rates of 1.8 mg l-1 h-1 for EMIM-Ac and 1.2 mg l-1 h-1 for
MMIM-DMP (Figure 24).
Figure 24: Activity of SSO1354 in high concentrations of ionic liquids. SSO1354 released
reducing sugars from 0.5% (w/v) Avicel (A) and tobacco cell wall polysaccharides (PCWS) (B)
dissolved in 80% (v/v) EMIM-Ac or MMIM-DMP. Assays were carried out at 90°C. The amount of
reducing sugar produced after ionic liquid precipitation was measured using the PAHBAH assay. The
sugars released by spontaneous hydrolysis in the absence of enzyme were measured in control
reactions and the values were subtracted from the test reactions. The release rate of reducing sugars
from dissolved Avicel was 5.5 mg l-1 h-1 in EMIM-Ac and 3.5 mg l-1 h-1 in MMIM-DMP, whereas the
release rate from dissolved cell wall polysaccharides was 1.8 mg l-1 h-1 for EMIM-Ac and 1.2 mg l-1 h-1
for MMIM-DMP.

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
50
Discussion
The conversion of lignocellulosic biomass into fermentable sugars is an important
step in the production of second-generation biofuels. A typical approach involves the
enzymatic hydrolysis of cellulose using microbial β-glucanases, but the efficiency of
saccharification is limited by the accessibility of the substrate, which must therefore
be pretreated (Galbe and Zacchi, 2007). Common cellulase preparations are derived
from mesophilic fungi and have usually an optimal activity around 50°C and pH 4–5
(Saddler and Gregg, 1998). In contrast, standard biomass pretreatment methods
involve harsh conditions that would invariably denature these enzymes, which means
they can only be added after pretreatment (Galbe and Zacchi, 2007). The production
of cellulases directly in the target biomass would be more economically and
environmentally sustainable than microbial fermentation, but the common mesophilic
enzymes are incompatible with current pretreatment regimens (Ragauskas et al.,
2006). Constitutively expressed and secreted mesophilic cellulases can also inhibit
normal plant growth, by reducing the integrity of the cell wall and therefore leaving
the plant vulnerable to environmental stress (Brunecky et al., 2011). These
challenges can be addressed by the sequestration of recombinant cellulases into
subcellular compartments with no direct access to cell wall components (Chapter 2)
or inducible expression when the plant is fully developed (Chapter 3).
The expression of cellulases from hyperthermophilic organisms is a promising
alternative because the temperature optima of these enzymes would ensure normal
plant growth and development at physiological temperatures and additionally enzyme
survival and activity under conventional pretreatment conditions. The sso1354 gene
from S. solfataricus encodes an endoglucanase from GH family 12 (She et al., 2001)
and does not contain a CBM. This enzyme is active at high temperatures and low pH
values, and is one of the first enzymes known to retain its activity in high
concentrations of ionic liquids. These features make SSO1354 an ideal candidate for
enzymatic biomass conversion directly in plants.
We demonstrated that SSO1354 can be functionally expressed in transgenic tobacco
plants by targeting the protein either for secretion into the apoplast or for retention in
the ER. The addition of a His6 tag to the N-terminus and an optional C-terminal KDEL
sequence had no adverse effects on enzyme activity. The Western blot revealed
different detectable forms of the protein, with molecular weights between 40 and 50
kDa (data not shown). The predicted molecular weight of the protein was 38 kDa, and

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
51
the discrepancy may reflect the presence of glycan residues on one or more of the
ten potential N-glycan acceptor sites.
Analysis of the morphological phenotype and the cell wall composition revealed no
significant differences between wild-type tobacco plants and transgenic lines
producing SSO1354, confirming that the enzyme remains inactive under
physiological conditions. The activity of plant-derived SSO1354 reached a maximum
of 0.5 µmol 4MU min-1 mg-1, reflecting its highly specific activity against 4MUC, but
this represents only a modest calculated yield of 1.1% TSP which is lower than other
cellulolytic enzymes in plant expression systems (Brunecky et al., 2011; Dai et al.,
2005; Ziegelhoffer et al., 2009).
Already known features of SSO1354 could be confirmed (Girfoglio et al., 2012;
Maurelli et al., 2008) and additionally, novel characteristics, such as the efficient
hydrolysis of substrates with β 1,4-linked glucose units in ionic liquids were
discovered. There was no hydrolysis of crystalline cellulose or substrates lacking β
1,4 linkages, nor did we observe the hydrolysis of substrates like oat xylan, which
comprises β 1,4-linked xylose, even though such activity has been reported
previously (Maurelli et al., 2008). This confirms that the enzyme is a strict β 1,4-
endoglucanase. The enzyme has a temperature optimum of 90°C, which is the
highest reported temperature for any cellulase expressed in plants and one of the
highest in any heterologous expression system (Bauer et al., 1999; Bronnenmeier et
al., 1995; Huang et al., 2005). The activity of the enzyme drops sharply as the
temperature declines, displaying only 25% activity at 70°C and virtually no activity at
temperatures below 30°C, suggesting there would be no hydrolysis of plant cellulose
under normal plant growth conditions. This is supported by the phenotype of the
transgenic plants which is not different compared to the wildtype and suggests that
post-harvest cellulose hydrolysis could be induced by a temperature shift.
SSO1354 is also advantageous because it tolerates a relatively broad pH range
(2.7–6.5, retaining at least 60% residual activity) whereas SSO1949 from the same
species has a sharp optimum at pH 1.8 (Huang et al., 2005). Under the optimal
conditions (pH 4.5 and 90°C), SSO1354 retains 85% activity after 1 h of incubation.
In addition to its remarkable thermostability and pH tolerance, SSO1354 also
tolerates moderate levels of organic solvents and high levels of salt.
Ionic liquids are very efficient in dissolving crystalline cellulose, however they are still
expensive (Abels et al., 2012; Zavrel et al., 2009). Promising results of their recycling

Chapter 4: Expression of a hyperthermophilic endoglucanase in tobacco
52
have been published recently (Abels et al., 2012). A unique feature of SSO1354 is its
activity in ionic liquids such as EMIM-Ac and MMIM-DMP. The hydrolysis rate was
3.5 mg l-1 h-1 in MMIM-DMP, showing this is a less suitable ionic liquid for SSO1354-
catalyzed biomass hydrolysis. SSO1354 was able to produce 5.5 mg l-1 h-1 of
reducing sugars from Avicel dissolved in EMIM-Ac, which is low compared to water
soluble substrates but still very promising for future optimization.
Although testing SSO1354 on artificial substrates confirmed its activity under a range
of conditions, plant biomass is a complex substrate with different components such
as cellulose, hemicelluloses and lignin, the latter acting as a significant inhibitor of
cellulases (hence the need for pretreatment) (Rahikainen et al., 2011). To investigate
the activity of SSO1354 against such complex substrates, we exposed the enzyme to
cell wall polysaccharides from tobacco leaves. This substrate is more heterogeneous
than crystalline cellulose but it has approximately 0.8% (dry weight) lignin content
and is therefore less complex than material derived from tobacco roots or stems
(Lagrimini et al., 1997). SSO1354 showed no significant activity against cell wall
polysaccharides in aqueous suspension, as determined by the release of reducing
sugars, but when the substrate was dissolved in ionic liquids the results were more
impressive. The enzyme released 1.8 mg l-1 h-1 of reducing sugars in EMIM-Ac and
1.2 mg l-1 h-1 in MMIM-DMP. These data indicate the possibility of combined
pretreatment and enzymatic hydrolysis.
The use of plants as an enzyme production system and subsequent utilization
requires two important criteria to be satisfied. The enzyme should not interfere with
cell wall synthesis or integrity and it must tolerate the extreme conditions found in
standard pretreatment processes. Hyperthermophilic cellulases like the
endoglucanase SSO1354 from S. solfataricus combine both features. This study
demonstrates the successful production of inactive SSO1354 at normal growth
conditions followed by the induction of enzyme activity at higher temperatures. To our
knowledge, SSO1354 is the first enzyme expressed in planta that remains active in
ionic liquids such as EMIM-Ac and MMIM-DMP, which are used for biomass
pretreatment. Plants have already been developed as a powerful platform for the
expression of biomass-processing enzymes, but the use of enzymes that function
optimally under harsh pretreatment conditions opens the way to develop combined
pretreatment and enzymatic hydrolysis strategies for the efficient conversion of
lignocellulosic plant biomass into fermentable sugars.

Chapter 5: Conclusion, Summary and Outlook
53
Chapter 5: Summary, Conclusion and Outlook
Lignocellulose is the major component of plant cell walls and is therefore an
abundant resource and a promising alternative to fossil fuel commodities. But plants
have evolved resistance to microbial and enzymatic attack, which makes them
recalcitrant to lignocellulosic conversion. Based on the complex structure and
composition of the lignocellulosic biomass, this recalcitrance is one the main barriers
preventing the large-scale industrial engineering of lignocellulose to produce fuels
and basic materials (Himmel et al., 2007).
Conversion strategies typically consist of three main steps: pretreatment (i) should
attenuate the recalcitrance of the substrate and therefore allow better access for the
enzymes used in the subsequent hydrolysis and saccharification steps (ii). The
resulting sugars are subsequently used for fermentation (iii) into fuels and
commodities. A significant reduction in enzyme costs, which still represent up to one
third of overall costs, is one of the most important goals for lignocellulose conversion
(Carroll and Somerville, 2009; Jørgensen et al., 2007). In planta production of
appropriate enzymes, e.g. GHs for lignocellulose conversion, is therefore a promising
option to improve the economic feasibility of such conversion processes (Sainz,
2011; Taylor et al., 2008).
Differential subcellular targeting plays an important role for in planta expression of
cellulases especially when functional enzymes are produced (Dai et al., 2005;
Ziegelhoffer et al., 2001). In chapter 2, the mesophilic endoglucanase TrCel5A from
the fungus T. reesei was produced in transgenic tobacco plants by targeting two
different subcellular compartments. A functional enzyme that was able to cleave
artificial substrates such as CMC and 4MU was produced by targeting either the
apoplast or the ER. However, the expression of TrCel5A was demonstrated to result
in an abnormal morphological phenotype and dramatic changes to the composition of
plant cell wall polysaccharides. The secreted enzyme reduced the content of
crystalline cellulose and inhibited both growth and germination, whereas keeping the
enzyme in the ER only altered the matrix phase polysaccharide composition without
visible effects on plant development, suggesting a more important role of the
crystalline cellulose for growth and development. Alternative strategies are therefore
required to produce cellulases in planta without a negative impact on growth and
development.

Chapter 5: Conclusion, Summary and Outlook
54
Inducible expression could be a promising alternative because potentially harmful
enzymes can be produced in fully-developed plants to avoid damage during
vegetative growth. In Chapter 3, the ethanol-inducible promoter system alcAmin35S
was used to produce TrCel5A endoglucanase. A single-step induction was inefficient
compared to constitutive expression but the optimization of the induction procedure
led yields of recombinant cellulase similar to the constitutive expression strategy.
Measurement of the composition of plant cell wall sugars in transgenic plants
carrying the induced expression cassette and phenotypical analysis were used to
characterize the impact of inducible TrCel5A expression on transgenic tobacco
plants. No significant difference was observed between transgenic lines with high
enzyme expression levels and wild-type controls. This underlines the advantage of
an induced expression strategy allowing a controlled application under different
expression conditions such as a specific developmental stage or defined expression
level without deleteriously impacting the plant development.
A high-level expression of functional cellulases is clearly beneficial for the
development of economically feasible in planta biomass conversion platforms, but the
enzymes must also survive the harsh conditions in the pretreatment step. Previous
studies already have shown that most enzymes do not survive common pretreatment
procedures like AFEX (Ragauskas et al., 2006; Teymouri et al., 2004).
Hyperthermophilic cellulases are more likely to remain active after or even during
pretreatment (Barnard et al., 2010; Blumer-Schuette et al., 2008). Accordingly, in
chapter 4, the successful production of S. solfataricus endoglucanase SSO1354 in
tobacco plants was described. Due to the enzyme´s inactivity under normal growth
conditions, SSO1354 was expressed without any morphological changes of the
plants or alterations in the plant cell wall sugar composition. However, the enzyme
can be activated when shifted to its optimal temperature, thus allowing full activity
during biomass conversion without the negative impact on growth and development
seen with mesophilic cellulases. Uniquely, it has been shown that SSO1354 can also
convert cellulose into smaller oligomers in the presence of ionic liquids, and can also
act on prepared cell walls in this environment. Plants therefore represent a powerful
platform for the expression of biomass-processing enzymes, including those that
function optimally under harsh pretreatment conditions. This offers new ways to
develop combined pretreatment and enzymatic hydrolysis strategies for the more
efficient conversion of lignocellulosic plant biomass into fermentable sugars.

Chapter 5: Conclusion, Summary and Outlook
55
The principal goal of in planta cellulase production is a high enzyme yield. Although
the achieved expression levels for TrCel5A in both strategies, constitutive and
inducible, are comparable to previous reports (Brunecky et al., 2011; Dai et al., 2005;
Ziegelhoffer et al., 2001), the expression level for SSO1354 is considerably lower.
However all obtained values are significantly lower than the amounts necessary for
complete self-conversion. Rough calculations, which ignore the requirement for
different activities and therefore different enzymes, estimate that ~15 kg of cellulase
would be required to convert 1 ton of biomass (Sainz, 2011). Even the best-
performing transgenic plants do not yet reach this goal (Hood et al., 2012). Potential
options for boosting in planta enzymes levels include plastid transformation, which
can produce impressive yields although homotransplastomic plants do often suffer
from sever damages that makes them unable to grow or flower in soil (Kim et al.,
2011; Petersen and Bock, 2011). However, there are approaches to significantly
increase the expression levels in transgenic plants like special breeding procedures
(Hood et al., 2012). Still the complete replacement of microbial enzymes by plant-
based production platforms seems unlikely in the short term, but in planta enzymes
could at least reduce the demand for microbial enzymes by contributing to the
biomass-degradation process thus increasing its economic feasibility. It is further
necessary to consider the loss of in planta produced enzyme during the pretreatment.
Even a comparably thermostable enzyme undergoes a dramatic loss of activity
during conventional AFEX pretreatment (Teymouri et al., 2004). A mesophilic
cellulase like TrCel5A would be completely inactivated. Enzymes from extremophilic
sources offer a more promising strategy to at least partially overcome the challenge
of inactivation by the combination of high temperature, high pressure, extreme pH,
and the presence of organic solvents used as pretreatment agents. Ionic liquids have
emerged as a powerful solvent for lignocellulose, though they are expensive and
highly toxic towards the microbes used for subsequent fermentation steps. The
endoglucanase SSO1354 is a promising enzyme for an application under these
harsh conditions. It is thus far unique in its ability to remain active in the presence of
high concentrations of ionic liquids, although the hydrolysis rate is comparatively low
and therefore unsuitable for economic conversion processes at the moment.
Besides high expression levels, characteristics like a good germination rate, plant
health and stress resistance are important for an industrial application of transgenic
plants producing cellulolytic enzymes. Most certainly the phenotypical alterations as

Chapter 5: Conclusion, Summary and Outlook
56
described here for TrCel5AAP plants would prevent an implementation as industrial
crop. The inducible expression of TrCel5A in fully developed plants or the post-
harvest activation of SSO1354 both led to plants with a normal phenotype. Therefore
both strategies seem to be promising for a later application.
Nevertheless cell wall modifications by heterologous expression of an endoglucanase
(E1 from A. cellulolyticus) have also been described as beneficial for subsequent
saccharification (Brunecky et al., 2011). The examined plants so far did not show an
abnormal phenotype as observed for TrCel5A in this study. This might be related to
different biochemical properties of the enzyme, though a detailed analysis of the cell
wall structure and composition is missing. However the effect, enabling an improved
saccharification, of both endoglucanses seems to be at least comparable. Also other
GHs, successfully implemented for in planta expression, have been leading to a
reduction of cell wall recalcitrance and therefore mitigating the pretreatment
conditions or reducing the amount of enzymes used in the saccharification (Shen et
al.; Zhang et al., 2011). Yet, there are no field studies for GHs expressing plants
concerning the impact of environmental stress. For the transgenic grass
Festuca arundinacea (tall fescue) with an altered esterification in the plant cell walls
an increased vulnerability to herbivore attack has been described (Buanafina and
Fescemyer, 2012). Future studies have to elucidate the effect of GHs in transgenic
plants in the field in order to design novel traits for industrial relevant crops.
In this study transgenic plants expressing recombinant cellulases have been
developed and analyzed. Novel insights have been obtained on the impact of such
enzymes on the plant especially the cell wall polysaccharides. Strategies have been
evaluated to avoid adverse effects on plant health. Developed analysis procedures
and the gained knowledge of this study established a promising basis for new
designed energy crops to contribute to novel sustainable and renewable resources.

Chapter 6: Materials and experimental procedures
57
Chapter 6: Materials and experimental procedures
Materials
Synthetic oligonucleotides
The oligonucleotide primers used to carry out the work described in this thesis are listed in
table 5.
Table 5: Oligonucleotide primers
Primer Sequence Length [bp] Description
cel5A fw TCC ATG GCA CAG CAG ACT GTC
TGG GGC
29 amplification of TrCel5A
adding NcoI site
cel5A rv TGC GGC CGC CTT TCT TGC GAG
ACA CG
26 amplification of TrCel5A
adding NotI site
cel5Acd fw AAC CAT GGA AGG GGT CCG ATT
TGC CGG CG
29 amplification of TrCel5A
catalytic domain adding
NcoI site
cel5A seq CTC TGG GCG CAT ACT GCA 18 sequencing TrCel5A
sso1354 His6 fw TCA TGA AAC ATC ACC ATC ACC
ATC ACG CGG CCG CTC AGT CTC
TCA GCG TTA AAC CCG T
58 amplification of sso1354
adding His6 and BspHI site
sso1354 rv AAC TCG AGT TAG AGG AGA GTT
TCA GAA AAG
30 amplification of sso1354
adding XhoI site
sso1354 KDEL rv AAC TCG AGC TAG AGC TCA TCT
TTG AGG AGA GTT TCA GAA AAG
42 amplification of sso1354
adding KDEL and XhoI site
mCherry fw GCG GCC GCA ATG GTG AGC
AAG GGC GAG GAG G
31 amplification of mCherry
adding NotI site
mCherry his6 rv CTC GAG CTA GTG ATG GTG ATG
GTG ATG AGC GGT CGC CTT GTA
CAG CTC GTC CAT G
55 amplification of mCherry
adding His6, NotI and XhoI
sites
alcAmin35S-fw AAG GAT CCA CCC GGG TGG CTA
GAA ATA TTT GCG ACT CTT CTG
42 amplification of alcAmin35s
adding BamHI and BstXI
sites
alcAmin35S rv AGC GGC CGC GTT TAA ACC AAT
TGG TCC TCT CCA AAT GAA ATG
AAC TTC C
49 amplification alcAmin35s
adding MfeI site
pSS-3 AGA GAG AGA TAG ATT TGT AGA
GA
23 3´ primer for pSS vector
pSS-5 ATC CTT CGC AAG ACC CTT CCT
CT
23 5´ primer for pSS vector

Chapter 6: Materials and experimental procedures
58
Plasmids
Plant expression vectors for transient expression and generation of transgenic plants were
based on pTRAk, a derivate of pPAM (GenBank no. AY027531). The vectors pTRAkc-AP
and pTRAkc-ER featured constitutive gene expression cassettes controlled by a duplicated
Cauliflower mosaic virus (CaMV) 35S promoter (P35SS). A plant-codon-optimized
signal peptide from the murine mAb24 heavy chain gene (Maclean et al., 2007) and the
sequence for a C-terminal His6 tag were also included. The ER retention signal (KDEL) was
also present in some constructs (Munro and Pelham, 1987). All vectors included a nos
expression cassette containing the nptII gene for kanamycin resistance.
Figure 25: Expression vector pTRAkc-rfp-his6-ERH for heterologous expression and ER
localization in plant cells. ColE1 ori, E. coli replicon; RK2 ori, A. tumefaciens replicon; bla, gene for
β-lactamase (AmpR); RB, right border; LB, left border; SAR, scaffold attachment region; pnos/pAnos,
nos expression cassette; p35SS/pA35SS, CaMV 35SS expression cassette; nptII, gene for neomycin
phosphotransferase II (KanR); DsRed, gene for Discosoma sp. red fluorescent protein; CHS, 5'-UTR
from P. hortense chalcone synthase gene; LPH, plant-codon-optimized signal peptide of murine
antibody mAb24; His6, hexahistidine tag; KDEL, signal for ER retention.
Bacterial strains
Escherichia coli strain DH5α [recA1, endA1, hsdR17 (rK-, mK+), ∆(lacZYA-argF)U169,
supE44, Φ80dlacZ∆M15, thi-1, gyrA96, relA1, deoR] was used for general cloning and
expression because of its modified recombination system (recA1) and the missing
endoglucanase (endA1).

Chapter 6: Materials and experimental procedures
59
For transient expression and stable transformation of tobacco, we used
Agrobacterium tumefaciens strain GV3101::pMP90RK (Koncz and Schell, 1986), which
contains the disarmed Ti helper plasmid pMP90RK conferring resistance against gentamicin
and kanamycin, and containing a RK2 origin of replication as well as virulence (vir) and
transfer (tra) genes essential for the horizontal transfer of T-DNA.
Plant material
Transient expression and stable nuclear transformation of tobacco plants was carried out in
Nicotiana tabacum cv. Petit Havana SR1.
Experimental procedures
Molecular biology procedures were carried out essentially as previously described
(Sambrook et al., 1989). Restriction endonucleases and DNA-modifying enzymes were used
according to the supplier’s instructions (New England Biolabs, Ipswich, MA USA).
Cloning
Construction of plant expression vectors for TrCel5A
The gene encoding TrCel5A (EGR51020.1, EMBL-CDS) was amplified by PCR from a cDNA
preparation of T. reesei QM9414 (kindly provided by Armin Merckelbach, Institute of
Molecular Biotechnology, RWTH Aachen University, Germany) using primers cel5A fw and
cel5A rv to amplify the whole gene except the first 21 codons comprising the fungal signal
peptide, or primers cel5Acd fw and cel5A rv to amplify the catalytic domain only. The purified
products were first transferred to the pCR2.1 vector (Invitrogen, Darmstadt, Germany) by TA-
cloning to generate the constructs pCR2.1 TrCel5A and pCR2.1 TrCel5Acd respectively.
Digestion with NcoI and NotI released a cassette which was then inserted into the NcoI-NotI
linearized vectors pTRAkc-TP for the whole enzyme and pTRAkc-ER for the catalytic domain
(Maclean et al., 2007). Additional constructs pTRAkc-AP-TrCel5A and pTRAkc-ER-TrCel5A
used in this study were prepared in earlier studies (Klose, 2009; Werthmann, 2009).
For the generation of TrCel5A/mCherry fusion constructs, the mCherry gene was amplified
from pmCherry-C1 (kindly provided by Christina Dickmeis, Institute of Molecular
Biotechnology, RWTH Aachen University, Germany) using primers mCherry fw and
mCherry rv. The purified products were digested with NotI and XhoI and inserted into the
similarly linearized vectors pTRAkc-AP-TrCel5A generating the construct pTRAkc-AP-
TrCel5AmCherry. Construct pTRAkc-ER-mCherry (Christina Dickmeis, Institute of Molecular
Biotechnology, RWTH Aachen University, Germany) was digested with NcoI and XhoI and
the resulting fragment was transferred into the NcoI-XhoI-linearized vector pTRAkc-AP-
TrCel5AmCherry to yield pTRAkc-ER-TrCel5AmCherry.

Chapter 6: Materials and experimental procedures
60
To generate the ethanol inducible expression construct (Caddick et al., 1998) the
alcAmin35S promoter was amplified from pTRAkt-alcR-alcA-stppc (Schirmacher, 2006) using
the primers alcAmin35S-fw and alcAmin35S-rv. The MfeI-PmeI fragment was transferred into
EcoRI-PmeI-linearized pTRAkc-AP-Cel5A and then subcloned as a BstXI-PmeI fragment into
FseI-PmeI-linearized pTRAkt-alcR (kindly provided by Dr. HJ Hirsch, Institute of Botany,
RWTH Aachen University, Germany).
Construction of plant expression vectors for SSO1354
The coding sequence of the SSO1354 catalytic domain (AAK41590.1, EMBL-CDS) was
amplified by PCR from S. solfataricus P2 genomic DNA (kindly provided by Raffaele Cannio,
Institute for Microelectronics and Microsystems, Naples, Italy) using the primers
sso1354 His6 fw and sso1354 rv or sso1354 KDEL rv to generate the His6-sso1354
products. The purified fragments were digested with BspHI and XhoI and inserted into the
NcoI/XhoI-digested pTRAkc-AP and pTRAkc-ER vectors. Additional constructs without a His6
tag (pTRAkc-AP-SSO1354oH and pTRAkc-ER-SSO1354oH were derived from earlier
studies (Röder, 2010).
Protein production in tobacco plants
Agroinfiltration of Nicotiana tabacum
Electrocompetent Agrobacterium tumefaciens cells, strain GV3101::pMP90RK GmR, KmR,
RifR (Koncz and Schell, 1986), were transformed by electroporation (Shen and Forde, 1989)
and selected on YEB medium plates (1% (w/v) Bacto tryptone, 1% (w/v) yeast extract, 0.5 %
(w/v) NaCl, 1.5% (w/v) agar, pH 7.0) supplemented with 100 mg ml-1 carbenicillin. Colonies
were transferred to liquid YEB medium containing kanamycin (50 mg ml-1), rifampicin (50 mg
ml-1) and carbenicillin (100 mg ml-1) for 36–40 h (26°C, 180 rpm) for selection. The
suspension was then supplemented with 10 µM acetosyringone, 10 mM MES (pH 5.6) and
10 mM glucose and the bacteria were incubated overnight. The OD600 of the culture was
adjusted to 1.0 with 2x infiltration medium (0.86% (w/v) MS salts, 10% (w/v) sucrose, 0.36%
(w/v) glucose, pH 5.6) and the adjusted suspension was supplemented with 200 µM
acetosyringone prior to incubation for approximately 45 min at room temperature. Before
infiltration, tobacco leaves were sprayed with water to open the stomata. The adaxial surface
of the leaves was scratched before the suspension was injected into the intercostal fields
using a syringe without a cannula.

Chapter 6: Materials and experimental procedures
61
Stable nuclear transformation of Nicotiana tabacum
Transgenic tobacco lines (Nicotiana tabacum L. cv. Petit Havana SR1) were established
using the leaf disc transformation method (Horsch, 1985). T0 plants were initially grown on
Murashige-Skoog medium containing 3% (w/v) sucrose, 100 mg l-1 kanamycin and 200 mg l-1
claforan, then transferred to soil in the greenhouse and selfed to produce T1 and T2
generation plants. The cell culture was kindly carried out by Flora Schuster, Institute of
Molecular Biotechnology, RWTH Aachen University.
Cultivation of tobacco plants
For the detailed analysis of recombinant protein expression and cell wall properties, plants
were grown in soil without the addition of fertilizer (Einheitserde classic, Patzer GmbH,
Sinntal-Altengronau) in the greenhouse at a constant 22°C (dark period) or 25°C (light
period) and 70% of relative humidity. Illumination was provided by Philips IP65 400 W lamps
(180 µmol s-1 m-2; λ= 400–700 nm) in a 16/8-h light/dark cycle with daily watering. The plants
were cultivated by Ibrahim Alamedi, Institute of Molecular Biotechnology, RWTH Aachen
University.
Ethanol-induced protein production in transgenic tobacco plants
Transgenic and wild-type tobacco plants were irrigated once with a defined volume of spilling
water containing concentrations of ethanol ranging from 0.1 to 10% (v/v). Induced plants
were kept separately from non-induced plants to avoid induction via ethanol vapor.
Isolation and analysis of plant genomic DNA
Genomic DNA was isolated from transgenic plants with the Wizard® Genomic DNA
Purification Kit (Promega, Mannheim, Germany). The transgene was detected by PCR using
the primers listed above.
Extraction and purification of recombinant proteins
Tobacco leaves from the transient expression experiments or transgenic plants were ground
in liquid nitrogen and homogenized in phosphate-buffered saline (PBS) at pH 7.0,
supplemented with 1 mM phenylmethylsulfonyl fluoride (PMSF). The extract was centrifuged
at 15,000 x g for 20 min at 4°C followed by filtration to remove suspended particles from the
extract.
The His6-tagged proteins were purified from the total soluble protein by Ni–NTA agarose
affinity chromatography (Qiagen, Hilden, Germany) following the manufacturer’s protocol.
Imidazole was removed using a Roti®Spin cut off column (Roth, Karlsruhe, Germany) with a
MWCO of 10 kDa. Total protein levels were determined using the Bradford method
(Bradford, 1976) with bovine serum albumin (Roth, Karlsruhe, Germany) as the standard.

Chapter 6: Materials and experimental procedures
62
Hyperthermophilic proteins without a tag were purified by removing plant bulk proteins in a
heat precipitation step at 85°C for 10 min and then separating small molecules such as
sugars from the recombinant enzyme by gel filtration with PD 10 Columns (GE Healthcare
Bioscience, Pittsburgh, PA USA).
Protein chemical methods
Protein separation by discontinuous SDS polyacrylamide gel electrophoresis
Protein samples were mixed 1:5 with reducing loading buffer (62.5 mM Tris-HCl (pH 6.8), 4%
(w/v) SDS, 0.05% (w/v) bromphenol blue, 10% (v/v) 2-mercaptoethanol, 30% (v/v) glycerol).
Separation was carried out under reducing conditions in 12% (w/v) polyacrylamide gels. Gels
were run in a Tris-glycine-SDS running buffer (25 mM Tris-HCl (pH 8.3), 0.2 M glycine,
0.25% (w/v) SDS) at a constant voltage of 200 V for 45 min. The gels were either stained
with 0.25% (v/v) Coomassie Brilliant Blue G-250, 50% (v/v) methanol, 10% (v/v) acetic acid
and then destained in 5% (v/v) methanol, 7.5% (v/v) acetic acid) or used for western blotting
(see below).
Protein analysis by Western blot
Proteins separated by SDS-PAGE were electrotransferred in blotting buffer (25 mM Tris,
192 mM glycine, 20% (v/v) methanol) for 60 min at a constant 100 V to a nitrocellulose
membrane, and blocked for 30 min at room temperature with 5% (w/v) skimmed milk
dissolved in PBS. His6-tagged proteins were detected with a polyclonal anti-His5 preparation
(Rockland Immunochemicals, Gilbertsville, USA) and a secondary, alkaline-phosphatase-
conjugated goat anti-mouse antibody (Dianova, Hamburg, Germany). T. reesei cellulases
were detected with polyclonal antibodies recognizing Trichoderma viride cellulase
(antibodies-online, Aachen, Germany) and a secondary alkaline-phosphatase-conjugated
goat anti-rabbit antibody (Dianova, Hamburg, Germany). The signal was visualized with
nitroblue tetrazolium chloride/5-bromo-4-chloro-3-indolyl phosphate p-toluidine salt
(NBT/BCIP; Roth, Karlsruhe, Germany).
Enzymatic assays
Protein analysis by activity staining
Protein samples were separated by SDS-PAGE in 12% polyacrylamide gels containing
0.15% (w/v) carboxymethylcellulose (CMC). The proteins were then renatured by washing
twice at room temperature for 15 min with 50 mM potassium acetate buffer (pH 4.8)
containing 20% (v/v) propan-2-ol followed by washing twice for 30 min with the same buffer
without propan-2-ol. The samples were incubated for 30 min in the potassium acetate buffer

Chapter 6: Materials and experimental procedures
63
at 50°C for mesophilic enzymes or 90°C for hyperthermophilic enzymes. The reactions were
stopped by incubating in 50 mM Tris-HCl (pH 7.5) for 30 min at room temperature. The gels
were stained for 30 min in 0.1% (w/v) Congo Red (Sigma-Aldrich, Seelze, Germany) and
destained in 1 M NaCl. To achieve a higher contrast between degradation and non-
degradation areas, the gel was incubated in 0.5% (v/v) acetic acid after destaining.
Activity against 4 methylumbelliferyl-β-D-cellobioside
The activity of cellulases against 4 methylumbelliferyl-β-D-cellobioside (4MUC) was carried
out as previously described (Ziegelhoffer et al., 2001). Each 5-µl sample aliquot was assayed
in 100 µl 50 mM sodium acetate (pH 4.8) containing 0.5 mM 4MUC in a 96-well plate. Plates
were covered with adhesive lids to prevent evaporation and incubated for 60 min either at
55°C or 90°C. The reaction was stopped by adding 100 µl 0.15 M glycine (pH 10.0). The
fluorescence was determined on a Tecan Infinite M200 (Tecan Deutschland GmbH,
Crailsheim) at 465 nm using an excitation wavelength of 360 nm. Fluorescence data
representing 4MUC activity was corrected by subtracting the average data from wild-type
crude plant extracts (n = 3). Conversion rates were calculated from corrected data based on
a series of 4MU standards (1–10 nM).
Activity against azo-CM-cellulose
The soluble chromogenic substrate azo-CM-cellulose (Megazyme, Bray, Ireland) was used
to determine the temperature and pH tolerance of the recombinant enzymes as previously
described (Jørgensen and Olsson, 2004), with five replicates taken at each measurement
point. Adsorption data were corrected by subtracting the average data from wild-type extracts
(n = 5). Temperature dependence was determined using 50 mM acetate buffer (pH 4.5) at
temperatures of 20–70°C. The pH tolerance was determined at 90°C using 50 mM citrate
buffer (pH 2.0–3.5), 50 mM acetate buffer (pH 3.5–5.5) and 50 mM phosphate buffer (pH
5.5–7.0). Salt tolerance was determined using 50 mM acetate buffer (pH 4.5) at 90°C
supplemented with up to 4.6 M NaCl.
Determination of reducing sugars
The reducing sugar content was measured using the p-hydroxybenzoic acid hydrazide
(PAHBAH) assay, by mixing one volume of sample with two volumes of PAHBAH reagent
(Lever, 1972). The solution was incubated at 100°C for 10 min and absorbance was
measured at 410 nm. Defined concentrations of glucose were used for calibration. Five
replicates were measured at each time point.
Determination of activity in ionic liquids
The conversion of pure cellulose or tobacco cell wall polysaccharides by the
hyperthermophilic endoglucanase was measured in high concentrations of ionic liquids. The

Chapter 6: Materials and experimental procedures
64
tobacco cell wall polysaccharides were prepared as previously described (Kawazu et al.,
1999). Therefore tobacco leaves were harvested and ground to powder under liquid nitrogen
followed by two washes with 25 ml distilled water and centrifugation (20,000 x g, 4°C, 15
min). Insoluble cell wall material was washed several times with methanol-chloroform (1:1)
until the green color disappeared completely. The organic solvent was then removed by
evaporation.
For activity determination, 0.5% (w/v) of the dried cell wall polysaccharides or Avicel was
dissolved in either 80% (v/v) 1-ethyl-3-methylimidazolium acetate (EMIM-Ac) or 80% (v/v)
1,3-dimethylimidazolium dimethylphosphate (MMIM-DMP). The aqueous-buffered enzyme
was added to a final concentration of 0.2 g l-1 and 20% (v/v) buffer in the reaction mixture.
The reaction mixture was incubated on a shaking platform at 90°C and samples were taken
at intervals to determine reducing sugar levels.
The reaction was stopped by cooling each sample and the cellulose was precipitated by
adding one volume of water followed by centrifugation (15,000 x g, RT, 2 min). Three phenol-
chloroform (1:1) extractions steps and one chloroform extraction were carried out to separate
the hydrolysis products and proteins from the ionic liquid. The reducing sugar content was
measured using the PAHBAH assay.
Chemical analysis of plant cell wall polymers
Extraction of alcohol-insoluble residues
Ten leaf tissue samples were taken from 6–7-week-old plants representing each transgenic
line and wild-type tobacco. To determine differences in the cellulose content, alcohol-
insoluble residues (AIRs) were prepared as previously described (Foster et al., 2010).
Approximately 50 mg per sample was ground to a fine powder under liquid nitrogen and plant
cell walls were isolated by washing with different organic solvents. Starch was removed by
hydrolysis with amylase and pullulanase (Sigma-Aldrich, Seelze, Germany). The remaining
AIRs were extracted with acetone, dried and weighed.
Analysis of matrix polysaccharides
The matrix polysaccharides were characterized as previously described (Albersheim et al.,
1967). We hydrolyzed 1–4 mg of AIR in 250 µl 2 M trifluoroacetic acid (TFA) for 90 min at
121°C. The cooled samples were centrifuged (10,000 x g, 4°C, 10 min) and 100 µl of the
supernatant was used for the alditol acetate derivatization procedure (Foster et al., 2010).
Insoluble material was washed three times with isopropanol and used to analyze the
crystalline cellulose content (see below). The cell wall matrix sugars were identified by
recording their mass profile or retention time. GC/MS was kindly carried out by Markus Günl
(Institute of Bio- and Geosciences, Forschungszentrum Jülich).

Chapter 6: Materials and experimental procedures
65
Measurement of crystalline cellulose
The crystalline cellulose content was determined as previously described (Updegraff, 1969b)
after hydrolysis of the non-crystalline cellulosic polysaccharides with acetic and nitric acid.
The remaining crystalline cellulose residues were hydrolyzed with 72% (w/v) sulfuric acid and
glucose was measured using the anthrone assay (Scott and Melvin, 1953).
Histological methods
Immunolabeling of TrCel5A
10-µm sections were prepared from transgenic and wild type leaf material using a Leica
Cryostat Microtome (Leica, Wetzlar, Germany). The sections were incubated in acetone for
30 min at room temperature, blocked in PBS containing 1% (v/v) goat serum (Sigma-Aldrich,
Seelze, Germany) and incubated with the primary antibody diluted (1:50) in PBS containing
1% (v/v) goat serum for 45 min. T. reesei cellulases were detected with polyclonal antibodies
recognizing Trichoderma viride cellulase (1 mg/ml, antibodies-online, Aachen, Germany),
additionally KDEL fused cellulases were detected with monoclonal antibodies recognizing the
KDEL sequence (1 mg/ml, Enzo Life Science, Lörrach, Germany). The sections were then
washed three times in PBS containing 0.05% (v/v) Tween-20 (Roth, Karlsruhe, Germany)
and incubated with the secondary antibody diluted (1:200) in PBS containing 1% (v/v) goat
serum for 45 min. For secondary antibody fluorescein isothiocyanate-conjugated goat anti-
rabbit or goat anti-mouse antibodies (1.5 mg/ml, Dianova, Hamburg, Germany) were used.
After three further washes in PBS containing 0.05% (v/v) Tween-20, the sections were
analyzed using a Leica DRM light microscope (Leica, Wetzlar, Germany).
Conversion of transgenic tobacco leaf material
To estimate the glucan content, tobacco leaf material was hydrolyzed in 72% sulfuric acid for
45 min at room temperature and hydrolyzed carbohydrates were measured using the
anthrone assay (Scott and Melvin, 1953). For the conversion of leaf material, 50-100 mg of
fresh tobacco leaf tissue was ground to powder under liquid nitrogen and resuspended in
50 mM sodium acetate (pH 4.8) containing 15 Filter paper units (FPU) CTec2 (Novozymes,
Bagsvaerd, Denmark) per gram of glucans. The reaction was stopped by boiling for 10 min
followed by centrifugation (10,000 x g, RT, 10 min) To determine the rate of hydrolysis, the
remaining crystalline cellulose was measured as previously described (Foster et al., 2010;
Scott and Melvin, 1953). Final values were based three independent experiments each
comprising three replicates.

Chapter 6: Materials and experimental procedures
66
Statistical methods
Arithmetic mean of a statistical sample
μ = 1������
Equation 1: Calculation of the arithmetic mean µ of a statistical sample. Where a is defined as
the value and n is the number of values.
Standard deviation of a statistical sample
= � 1� − 1�(� − μ)�
Equation 2: Calculation of the standard deviation σ of a statistical sample. Where a is defined as
the value, µ is the arithmetic mean and n is the number of values.
Statistical difference between different groups of samples
One-way analysis of variance (ANOVA) followed by Student’s pairwise t test was used to
determine the significance of differences between groups, with a threshold of P ≤ 0.05 or
even P ≤ 0.01, using Microsoft Excel 2010.

References
67
References
Abdeev, R.M., Goldenkova, I.V., Musiychuk, K.A. and Piruzian, E.S. (2003) Expression of a
thermostable bacterial cellulase in transgenic tobacco plants. Russian Journal of Genetics 39,
300-305.
Abdeev, R.M., Musiichuk, K.A., Goldenkova, I.V., Sotchenkov, D.V., Salekhi Dzhuzani, G.R., Alyavina,
A.K., Zagoskina, N.V. and Piruzian, E.S. (2004) Morphology and Phytohormone Content in
Transgenic Tobacco Plants Expressing Bacterial Thermostable Cellulase. Russian Journal of
Plant Physiology 51, 642-647.
Abels, C., Redepenning, C., Moll, A., Melin, T. and Wessling, M. (2012) Simple purification of ionic
liquid solvents by nanofiltration in biorefining of lignocellulosic substrates. Journal of
Membrane Science 405–406, 1-10.
Abramson, M., Shoseyov, O., Hirsch, S. and Shani, Z. (2013) Genetic modifications of plant cell walls
to increase biomass and bioethanol production. In: Advanced Biofuels and Bioproducts (Lee,
J.W. ed) pp. 315-338. Springer New York.
Agrawal, P., Verma, D. and Daniell, H. (2011) Expression of Trichoderma reesei beta-mannanase in
tobacco chloroplasts and its utilization in lignocellulosic woody biomass hydrolysis. PLoS One
6, e29302.
Ait-ali, T., Rands, C. and Harberd, N.P. (2003) Flexible control of plant architecture and yield via
switchable expression of Arabidopsis gai. Plant Biotechnology Journal 1, 337-343.
Albersheim, P., Nevins, D.J., English, P.D. and Karr, A. (1967) A method for the analysis of sugars in
plant cell-wall polysaccharides by gas-liquid chromatography. Carbohydrate Research 5, 340-
345.
Alvira, P., Tomás-Pejó, E., Ballesteros, M. and Negro, M.J. (2010) Pretreatment technologies for an
efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresource
Technology 101, 4851-4861.
Amorim, H., Lopes, M., de Castro Oliveira, J., Buckeridge, M. and Goldman, G. (2011) Scientific
challenges of bioethanol production in Brazil. Applied Microbiology and Biotechnology, 1-9.
Atalla, R.H., Brady, J.W., Matthews, J.F., Ding, S.-Y. and Himmel, M.E. (2009) Structures of plant cell
wall celluloses. In: Biomass Recalcitrance pp. 188-212. Blackwell Publishing Ltd.
Atmodjo, M.A., Hao, Z. and Mohnen, D. (2013) Evolving views of pectin biosynthesis. Annual Review
of Plant Biology 64, 747-779.
Barnard, D., Casanueva, A., Tuffin, M. and Cowan, D. (2010) Extremophiles in biofuel synthesis.
Environmental Technology 31, 871-888.
Basu, S.S. (2001) Self-processing plants and plant parts. EP 1432798 B1.
Bauer, M.W., Driskill, L.E., Callen, W., Snead, M.A., Mathur, E.J. and Kelly, R.M. (1999) An
endoglucanase, EglA, from the hyperthermophilic archaeon pyrococcus furiosus hydrolyzes
beta -1,4 bonds in mixed-linkage (1-3),(1-4)-beta -D-glucans and cellulose. Journal of
Bacteriology 181, 284-290.

References
68
Beyene, G., Foyer, C.H. and Kunert, K.J. (2006) Two new cysteine proteinases with specific
expression patterns in mature and senescent tobacco (Nicotiana tabacum L.) leaves. Journal
of Experimental Botany 57, 1431-1443.
Bhalla, A., Bansal, N., Kumar, S., Bischoff, K.M. and Sani, R.K. (2013) Improved lignocellulose
conversion to biofuels with thermophilic bacteria and thermostable enzymes. Bioresource
Technology 128, 751-759.
Blumer-Schuette, S.E., Kataeva, I., Westpheling, J., Adams, M.W.W. and Kelly, R.M. (2008) Extremely
thermophilic microorganisms for biomass conversion: status and prospects. Current Opinion in
Biotechnology 19, 210-217.
Bodirlau, R., Teaca, C.-A. and Spiridon, I. (2010) Influence of ionic liquid on hydrolyzed cellulose
material: FT-IR spectroscopy and TG-DTG-DSC analysis. International Journal of Polymer
Analysis and Characterization 15, 460-469.
Boerjan, W., Ralph, J. and Baucher, M. (2003) Lignin biosynthesis. Annual Review of Plant Biology
54, 519-546.
Bonawitz, N.D. and Chapple, C. (2010) The genetics of lignin biosynthesis: Connecting genotype to
phenotype. Annual review of genetics 44, 337-363.
Boraston, A.B., Bolam, D.N., Gilbert, H.J. and Davies, G.J. (2004) Carbohydrate-binding modules:
Fine-tuning polysaccharide recognition. The Biochemical Journal 382, 769-781.
Borkhardt, B., Harholt, J., Ulvskov, P., Ahring, B.K., Jørgensen, B. and Brinch-Pedersen, H. (2010)
Autohydrolysis of plant xylans by apoplastic expression of thermophilic bacterial endo-
xylanases. Plant Biotechnology Journal 8, 363-374.
Bortesi, L., Rademacher, T., Schiermeyer, A., Schuster, F., Pezzotti, M. and Schillberg, S. (2012)
Development of an optimized tetracycline-inducible expression system to increase the
accumulation of interleukin-10 in tobacco BY-2 suspension cells. BMC Biotechnology 12, 40.
Bradford, M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72, 248-254.
Brodeur, G., Yau, E., Badal, K., Collier, J., Ramachandran, K.B. and Ramakrishnan, S. (2011)
Chemical and physicochemical pretreatment of lignocellulosic biomass: A review. Enzyme
Research 2011.
Bronnenmeier, K., Kern, A., Liebl, W. and Staudenbauer, W. (1995) Purification of Thermotoga
maritima enzymes for the degradation of cellulosic materials. Applied Environmental
Microbiology 61, 1399-1407.
Brotman, Y., Briff, E., Viterbo, A. and Chet, I. (2008) Role of swollenin, an expansin-like protein from
Trichoderma, in plant root colonization. Plant Physiology 147, 779-789.
Bruce, W., Folkerts, O., Garnaat, C., Crasta, O., Roth, B. and Bowen, B. (2000) Expression profiling of
the maize flavonoid pathway genes controlled by estradiol-inducible transcription factors CRC
and P. The Plant Cell Online 12, 65-79.
Brunecky, R., Selig, M., Vinzant, T., Himmel, M., Lee, D., Blaylock, M. and Decker, S. (2011) In planta
expression of A. cellulolyticus Cel5A endocellulase reduces cell wall recalcitrance in tobacco
and maize. Biotechnology for Biofuels 4, 1.

References
69
Buanafina, M.d. and Fescemyer, H. (2012) Modification of esterified cell wall phenolics increases
vulnerability of tall fescue to herbivory by the fall armyworm. Planta 236, 513-523.
Buanafina, M.M.d.O., Langdon, T., Hauck, B., Dalton, S., Timms-Taravella, E. and Morris, P. (2010)
Targeting expression of a fungal ferulic acid esterase to the apoplast, endoplasmic reticulum
or golgi can disrupt feruloylation of the growing cell wall and increase the biodegradability of
tall fescue (Festuca arundinacea). Plant Biotechnology Journal 8, 316-331.
Caddick, M.X., Greenland, A.J., Jepson, l., Krause, K.-P., Qu, N., Riddell, K.V., Salter, M.G., Schuch,
W., Sonnewald, U. and Tomsett, A.B. (1998) An ethanol inducible gene switch for plants used
to manipulate carbon metabolism. Nature Biotechnology 16, 177-180.
Cannio, R., Prizito, N., Rossi, M. and Morana, A. (2004) A xylan-degrading strain of Sulfolobus
solfataricus: isolation and characterization of the xylanase activity. Extremophiles 8, 117-124.
Carpita, N.C. and Gibeaut, D.M. (1993) Structural models of primary cell walls in flowering plants:
consistency of molecular structure with the physical properties of the walls during growth. The
Plant Journal 3, 1-30.
Carrasco, J.E., Sáiz, M., Navarro, A., Soriano, P., Sáez, F. and Martinez, J.M. (1994) Effects of dilute
acid and steam explosion pretreatments on the cellulose structure and kinetics of cellulosic
fraction hydrolysis by dilute acids in lignocellulosic materials. Applied Biochemistry and
Biotechnology 45-46, 23-34.
Carroll, A. and Somerville, C. (2009) Cellulosic biofuels. Annual Review of Plant Biology 60, 165-182.
Chaddad, F.R. (2010) UNICA: Challenges to deliver sustainability in the Brazilian sugarcane industry.
International Food and Agribusiness Management Review 13, 174-191.
Chen, F. and Dixon, R.A. (2007) Lignin modification improves fermentable sugar yields for biofuel
production. Nature Biotechnology 25, 759-761.
Cheng, Y.-S., Zheng, Y., Yu, C., Dooley, T., Jenkins, B. and VanderGheynst, J. (2010) Evaluation of
high solids alkaline pretreatment of rice straw. Applied Biochemistry and Biotechnology 162,
1768-1784.
Chou, H., Dai, Z., Hsieh, C. and Ku, M. (2011) High level expression of Acidothermus cellulolyticus
beta-1, 4-endoglucanase in transgenic rice enhances the hydrolysis of its straw by cultured
cow gastric fluid. Biotechnology for Biofuels 4, 58.
Chundawat, S.P.S., Beckham, G.T., Himmel, M.E. and Dale, B.E. (2011) Deconstruction of
lignocellulosic biomass to fuels and chemicals. Annual Review of Chemical and Biomolecular
Engineering 2, 121-145.
Cobucci-Ponzano, B., Aurilia, V., Riccio, G., Henrissat, B., Coutinho, P.M., Strazzulli, A., Padula, A.,
Corsaro, M.M., Pieretti, G., Pocsfalvi, G., Fiume, I., Cannio, R., Rossi, M. and Moracci, M.
(2010) A New Archaeal β-Glycosidase from Sulfolobus solfataricus. Journal of Biological
Chemistry 285, 20691-20703.
Cosgrove, D.J. (1997) Assembly and enlargement of the primary cell wall in plants. Annual Review of
Cell Developmental Biology 13, 171-201.
Dai, Z., Hooker, B.S., Anderson, D.B. and Thomas, S.R. (2000) Expression of Acidothermus
cellulolyticus endoglucanase E1 in transgenic tobacco: biochemical characteristics and
physiological effects. Transgenic Research 9, 43-54.

References
70
Dai, Z., Hooker, B.S., Quesenberry, R.D. and Thomas, S.R. (2005) Optimization of Acidothermus
cellulolyticus endoglucanase (E1) production in transgenic tobacco plants by transcriptional,
post-transcription and post-translational modification. Transgenic Research 14, 627-643.
Demesure, B., Sodzi, N. and Petit, R.J. (1995) A set of universal primers for amplification of
polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Molecular
Ecology 4, 129-134.
Endler, A. and Persson, S. (2011) Cellulose Synthases and Synthesis in Arabidopsis. Molecular Plant
4, 199-211.
Fairley, P. (2011) Introduction: Next generation biofuels. Nature 474, S2-S5.
Fischer, R., Stoger, E., Schillberg, S., Christou, P. and Twyman, R.M. (2004) Plant-based production
of biopharmaceuticals. Current Opinion in Plant Biology 7, 152-158.
Forsberg, Z., Vaaje-Kolstad, G., Westereng, B., Bunæs, A.C., Stenstrøm, Y., MacKenzie, A., Sørlie,
M., Horn, S.J. and Eijsink, V.G.H. (2011) Cleavage of cellulose by a CBM33 protein. Protein
Science 20, 1479-1483.
Foster, C.E., Martin, T.M. and Pauly, M. (2010) Comprehensive compositional analysis of plant cell
walls (lignocellulosic biomass) part II: carbohydrates. Journal of visualized experiments :
JoVE.
Galbe, M. and Zacchi, G. (2007) Pretreatment of lignocellulosic materials for efficient bioethanol
production. Advances in Biochemical Engineering/Biotechnology 108, 41-65.
Gaulin, E., Dramé, N., Lafitte, C., Torto-Alalibo, T., Martinez, Y., Ameline-Torregrosa, C., Khatib, M.,
Mazarguil, H., Villalba-Mateos, F., Kamoun, S., Mazars, C., Dumas, B., Bottin, A., Esquerré-
Tugayé, M.-T. and Rickauer, M. (2006) Cellulose binding domains of a phytophthora cell wall
protein are novel pathogen-associated molecular patterns. The Plant Cell Online 18, 1766-
1777.
Geddes, C.C., Nieves, I.U. and Ingram, L.O. (2011) Advances in ethanol production. Current Opinion
in Biotechnology 22, 312-319.
Gilkes, N.R., Warren, R.A., Miller, R.C. and Kilburn, D.G. (1988) Precise excision of the cellulose
binding domains from two Cellulomonas fimi cellulases by a homologous protease and the
effect on catalysis. Journal of Biological Chemistry 263, 10401-10407.
Girfoglio, M., Rossi, M. and Cannio, R. (2012) Cellulose degradation by Sulfolobus solfataricus
requires a cell anchored endo-β-1-4-glucanase. Journal of Bacteriology.
Graham-Rowe, D. (2011) Agriculture: Beyond food versus fuel. Nature 474, S6-S8.
Gray, B., Yang, H., Ahner, B. and Hanson, M. (2011a) An efficient downstream box fusion allows high-
level accumulation of active bacterial beta-glucosidase in tobacco chloroplasts. Plant
Molecular Biology 76, 345 - 355.
Gray, B.N., Bougri, O., Carlson, A.R., Meissner, J., Pan, S., Parker, M.H., Zhang, D., Samoylov, V.,
Ekborg, N.A. and Michael Raab, R. (2011b) Global and grain-specific accumulation of
glycoside hydrolase family 10 xylanases in transgenic maize (Zea mays). Plant Biotechnol J 9,
1100-1108.

References
71
Günl, M. and Pauly, M. (2011) AXY3 encodes a α-xylosidase that impacts the structure and
accessibility of the hemicellulose xyloglucan in Arabidopsis plant cell walls. Planta 233, 707-
719.
Harris, P.J. and Stone, B.A. (2009) Chemistry and Molecular Organization of Plant Cell Walls. In:
Biomass Recalcitrance pp. 61-93. Blackwell Publishing Ltd.
Harrison, M.D., Geijskes, J., Coleman, H.D., Shand, K., Kinkema, M., Palupe, A., Hassall, R., Sainz,
M., Lloyd, R., Miles, S. and Dale, J.L. (2011) Accumulation of recombinant cellobiohydrolase
and endoglucanase in the leaves of mature transgenic sugar cane. Plant Biotechnology
Journal 9, 884-896.
Henrissat, B., Driguez, H., Viet, C. and Schulein, M. (1985) Synergism of cellulases from Trichoderma
reesei in the degradation of cellulose. Nature Biotechnology 3, 722-726.
Hill, J., Nelson, E., Tilman, D., Polasky, S. and Tiffany, D. (2006) Environmental, economic, and
energetic costs and benefits of biodiesel and ethanol biofuels. Proceedings of the National
Academy of Sciences 103, 11206-11210.
Himmel, M.E., Ding, S.-Y., Johnson, D.K., Adney, W.S., Nimlos, M.R., Brady, J.W. and Foust, T.D.
(2007) Biomass recalcitrance: Engineering plants and enzymes for biofuels production.
Science 315, 804-807.
Hood, E.E., Devaiah, S.P., Fake, G., Egelkrout, E., Teoh, K., Requesens, D.V., Hayden, C., Hood,
K.R., Pappu, K.M., Carroll, J. and Howard, J.A. (2012) Manipulating corn germplasm to
increase recombinant protein accumulation. Plant Biotechnology Journal 10, 20-30.
Hood, E.E., Love, R., Lane, J., Bray, J., Clough, R., Pappu, K., Drees, C., Hood, K.R., Yoon, S.,
Ahmad, A. and Howard, J.A. (2007) Subcellular targeting is a key condition for high-level
accumulation of cellulase protein in transgenic maize seed. Plant Biotechnology Journal 5,
709-719.
Horn, S., Vaaje-Kolstad, G., Westereng, B. and Eijsink, V.G. (2012) Novel enzymes for the
degradation of cellulose. Biotechnology for Biofuels 5, 45.
Horsch, R.B. (1985) A simple and general method for transferring genes into plants. Science 227,
1229-1231.
Huang, Y., Krauss, G., Cottaz, S., Driguez, H. and Lipps, G. (2005) A highly acid-stable and
thermostable endo-beta-glucanase from the thermoacidophilic archaeon Sulfolobus
solfataricus. The Biochemical Journal 385, 581-588.
Ilmberger, N., Meske, D., Juergensen, J., Schulte, M., Barthen, P., Rabausch, U., Angelov, A.,
Mientus, M., Liebl, W., Schmitz, R. and Streit, W. (2011) Metagenomic cellulases highly
tolerant towards the presence of ionic liquids—linking thermostability and halotolerance.
Applied Microbiology and Biotechnology, 1-12.
Jäger, G. and Büchs, J. (2012) Biocatalytic conversion of lignocellulose to platform chemicals.
Biotechnology Journal 7, 1122-1136.
Jäger, G., Girfoglio, M., Dollo, F., Rinaldi, R., Bongard, H., Commandeur, U., Fischer, R., Spiess, A.
and Buchs, J. (2011) How recombinant swollenin from Kluyveromyces lactis affects cellulosic
substrates and accelerates their hydrolysis. Biotechnology for Biofuels 4, 33.

References
72
Jäger, G., Wu, Z., Garschhammer, K., Engel, P., Klement, T., Rinaldi, R., Spiess, A. and Buchs, J.
(2010) Practical screening of purified cellobiohydrolases and endoglucanases with alpha-
cellulose and specification of hydrodynamics. Biotechnology for Biofuels 3, 18.
Jensen, J.K., Schultink, A., Keegstra, K., Wilkerson, C.G. and Pauly, M. (2012) RNA-Seq analysis of
developing Nasturtium seeds (Tropaeolum majus): Identification and characterization of an
additional galactosyltransferase involved in xyloglucan biosynthesis. Molecular Plant 5, 984-
992.
Jørgensen, H., Kristensen, J.B. and Felby, C. (2007) Enzymatic conversion of lignocellulose into
fermentable sugars: challenges and opportunities. Biofuels, Bioproducts and Biorefining 1,
119-134.
Jørgensen, H. and Olsson, L. (2004) Production of cellulases by Penicillium brasilianum IBT 20888—
Effect of substrate on hydrolytic performance. Enzyme and Microbial Technology 38, 381-390.
Jung, J.H., Vermerris, W., Gallo, M., Fedenko, J.R., Erickson, J.E. and Altpeter, F. (2013a) RNA
interference suppression of lignin biosynthesis increases fermentable sugar yields for biofuel
production from field-grown sugarcane. Plant Biotechnology Journal, n/a-n/a.
Jung, S., Kim, S., Bae, H., Lim, H.-S. and Bae, H.-J. (2010) Expression of thermostable bacterial β-
glucosidase (BglB) in transgenic tobacco plants. Bioresource Technology 101, 7144-7150.
Jung, Y.H., Cho, H.J., Lee, J.-S., Noh, E.W., Park, O.K. and Kim, K.H. (2013b) Evaluation of a
transgenic poplar as a potential biomass crop for biofuel production. Bioresource Technology
129, 639-641.
Kaida, R., Kaku, T., Baba, K.i., Oyadomari, M., Watanabe, T., Nishida, K., Kanaya, T., Shani, Z.,
Shoseyov, O. and Hayashi, T. (2009) Loosening xyloglucan accelerates the enzymatic
degradation of cellulose in wood. Molecular Plant 2, 904-909.
Kawazu, T., Ohta, T., Ito, K., Shibata, M., Kimura, T., Sakka, K. and Ohmiya, K. (1996) Expression of
a Ruminococcus albus cellulase gene in tobacco suspension cells. Journal of Fermentation
and Bioengineering 82, 205-209.
Kawazu, T., Sun, J.L., Shibata, M., Kimura, T., Sakka, K. and Ohmiya, K. (1999) Expression of a
bacterial endoglucanase gene in tobacco increases digestibility of its cell wall fibers. Journal of
Bioscience and Bioengineering 88, 421-425.
Kilpeläinen, I., Xie, H., King, A., Granstrom, M., Heikkinen, S. and Argyropoulos, D.S. (2007)
Dissolution of wood in ionic liquids. Journal of Agricultural and Food Chemistry 55, 9142-9148.
Kim, J., Kavas, M., Fouad, W., Nong, G., Preston, J. and Altpeter, F. (2011) Production of
hyperthermostable GH10 xylanase Xyl10B from Thermotoga maritima in transplastomic plants
enables complete hydrolysis of methylglucuronoxylan to fermentable sugars for biofuel
production. Plant Molecular Biology 76, 357-369.
Kim, S., Kim, Y.-O., Lee, Y., Choi, I., Joshi, C.P., Lee, K. and Bae, H.-J. (2012) The transgenic poplar
as an efficient bioreactor system for the production of xylanase. Bioscience, Biotechnology,
and Biochemistry 76, 1140-1145.
Kim, S., Lee, D.-S., Choi, I., Ahn, S.-J., Kim, Y.-H. and Bae, H.-J. (2010) Arabidopsis thaliana Rubisco
small subunit transit peptide increases the accumulation of Thermotoga maritima

References
73
endoglucanase Cel5A in chloroplasts of transgenic tobacco plants. Transgenic Research 19,
489-497.
Kimura, T., Mizutani, T., Sun, J.-L., Kawazu, T., Karita, S., Sakka, M., Kobayashi, Y., Ohmiya, K. and
Sakka, K. (2010) Stable production of thermotolerant xylanase B of Clostridium stercorarium
in transgenic tobacco and rice. Bioscience, Biotechnology, and Biochemistry 74, 954-960.
Klose, H. (2009) Expression mikrobieller cellulolytischer Proteine in Pflanzen. MSc Thesis RWTH
Aachen University.
Koncz, C. and Schell, J. (1986) The promoter of T-DNA gene 5 controls the tissue-specific expression
of chimaeric genes carried by a novel type of Agrobacterium binary vector. Molecular and
General Genetics MGG 204, 383-396.
Kumar, R., Mago, G., Balan, V. and Wyman, C.E. (2009) Physical and chemical characterizations of
corn stover and poplar solids resulting from leading pretreatment technologies. Bioresource
Technology 100, 3948-3962.
Kumar, R., Singh, S. and Singh, O. (2008) Bioconversion of lignocellulosic biomass: biochemical and
molecular perspectives. Journal of Industrial Microbiology & Biotechnology 35, 377-391.
Lagrimini, L.M., Gingas, V., Finger, F., Rothstein, S. and Liu, T. (1997) Characterization of antisense
transformed plants deficient in the tobacco anionic peroxidase. Plant Physiology 114, 1187-
1196.
Lebel, E.G., Heifetz, P.B., Ward, E.R. and Uknes, S.J. (2008) Transgenic plant expressing a cellulase.
US Patent 7361806 B2.
Lehmann, C., Sibilla, F., Maugeri, Z., Streit, W.R., Dominguez de Maria, P., Martinez, R. and
Schwaneberg, U. (2012) Reengineering CelA2 cellulase for hydrolysis in aqueous solutions of
deep eutectic solvents and concentrated seawater. Green Chemistry 14, 2719-2726.
Lever, M. (1972) A new reaction for colorimetric determination of carbohydrates. Analytical
Biochemistry 47, 273-279.
Lewis, N.S. and Nocera, D.G. (2006) Powering the planet: Chemical challenges in solar energy
utilization. Proceedings of the National Academy of Sciences 103, 15729-15735.
Liang, C., Fioroni, M., Rodríguez-Ropero, F., Xue, Y., Schwaneberg, U. and Ma, Y. (2011) Directed
evolution of a thermophilic endoglucanase (Cel5A) into highly active Cel5A variants with an
expanded temperature profile. Journal of Biotechnology 154, 46-53.
Limauro, D., Cannio, R., Fiorentino, G., Rossi, M. and Bartolucci, S. (2001) Identification and
molecular characterization of an endoglucanase gene celS from the extremely thermophilic
archaeon Sulfolobus solfataricus. Extremophiles 5, 213-219.
Lopez-Casado, G., Urbanowicz, B.R., Damasceno, C.M.B. and Rose, J.K.C. (2008) Plant glycosyl
hydrolases and biofuels: a natural marriage. Current Opinion in Plant Biology 11, 329-337.
Lynd, L.R., Weimer, P.J., van Zyl, W.H. and Pretorius, I.S. (2002) Microbial cellulose utilization:
Fundamentals and biotechnology. Microbiology and Molecular Biology Reviews 66, 506-577.
Maclean, J., Koekemoer, M., Olivier, A.J., Stewart, D., Hitzeroth, I.I., Rademacher, T., Fischer, R.,
Williamson, A.-L. and Rybicki, E.P. (2007) Optimization of human papillomavirus type 16
(HPV-16) L1 expression in plants: comparison of the suitability of different HPV-16 L1 gene

References
74
variants and different cell-compartment localization. The Journal of General Virology 88, 1460-
1469.
Mahadevan, S., Wi, S., Kim, Y., Lee, K. and Bae, H.-J. (2011) In planta differential targeting analysis
of Thermotoga maritima Cel5A and CBM6-engineered Cel5A for autohydrolysis. Transgenic
Research 20, 877-886.
Maloney, V.J. and Mansfield, S.D. (2010) Characterization and varied expression of a membrane-
bound endo-β-1,4-glucanase in hybrid poplar. Plant Biotechnology Journal 8, 294-307.
Marzialetti, T., Valenzuela Olarte, M.B., Sievers, C., Hoskins, T.J.C., Agrawal, P.K. and Jones, C.W.
(2008) Dilute acid hydrolysis of loblolly pine: A comprehensive approach. Industrial &
Engineering Chemistry Research 47, 7131-7140.
Maurelli, L., Giovane, A., Esposito, A., Moracci, M., Fiume, I., Rossi, M. and Morana, A. (2008)
Evidence that the xylanase activity from Sulfolobus solfataricus Oα is encoded by the
endoglucanase precursor gene (sso1354 ) and characterization of the associated cellulase
activity. Extremophiles 12, 689-700.
McIntosh, S. and Vancov, T. (2010) Enhanced enzyme saccharification of Sorghum bicolor straw
using dilute alkali pretreatment. Bioresource Technology 101, 6718-6727.
Michael, W., May, W. and Hong, H. (2007) Life-cycle energy and greenhouse gas emission impacts of
different corn ethanol plant types. Environmental Research Letters 2, 024001.
Minic, Z. (2008) Physiological roles of plant glycoside hydrolases. Planta 227, 723-740.
Molhoj, M., Ulvskov, P. and Dal Degan, F. (2001) Characterization of a functional soluble form of a
Brassica napus membrane-anchored endo-1,4-beta -glucanase heterologously expressed in
Pichia pastoris. Plant Physiology 127, 674-684.
Montalvo-Rodriguez, R., Haseltine, C., Huess-LaRossa, K., Clemente, T., Soto, J., Staswick, P. and
Blum, P. (2000) Autohydrolysis of plant polysaccharides using transgenic hyperthermophilic
enzymes. Biotechnology and Bioengineering 70, 151-159.
Moore, P. and Staehelin, L.A. (1988) Immunogold localization of the cell-wall-matrix polysaccharides
rhamnogalacturonan I and xyloglucan during cell expansion and cytokinesis in Trifolium
pratense L.; implication for secretory pathways. Planta 174, 433-445.
Mosier, N., Wyman, C., Dale, B., Elander, R., Lee, Y.Y., Holtzapple, M. and Ladisch, M. (2005)
Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresource
Technology 96, 673-686.
Munro, S. and Pelham, H.R.B. (1987) A C-terminal signal prevents secretion of luminal ER proteins.
Cell 48, 899-907.
Nakazawa, H., Okada, K., Kobayashi, R., Kubota, T., Onodera, T., Ochiai, N., Omata, N., Ogasawara,
W., Okada, H. and Morikawa, Y. (2008) Characterization of the catalytic domains of
Trichoderma reesei endoglucanase I, II, and III, expressed in Escherichia coli. Applied
Microbiology and Biotechnology 81, 681-689.
Obembe, O.O., Jacobsen, E., Timmers, J., Gilbert, H., Blake, A.W., Knox, J.P., Visser, R.G. and
Vincken, J.P. (2007) Promiscuous, non-catalytic, tandem carbohydrate-binding modules
modulate the cell-wall structure and development of transgenic tobacco (Nicotiana tabacum)
plants. Journal of Plant Research 120, 605-617.

References
75
Okuzaki, A., Konagaya, K.-i., Nanasato, Y., Tsuda, M. and Tabei, Y. (2011) Estrogen-inducible GFP
expression patterns in rice (Oryza sativa L.). Plant Cell Reports 30, 529-538.
Ouyang, J., Yan, M., Kong, D. and Xu, L. (2006) A complete protein pattern of cellulase and
hemicellulase genes in the filamentous fungus Trichoderma reesei. Biotechnol Journal 1,
1266-1274.
Park, Y.W., Tominaga, R., Sugiyama, J., Furuta, Y., Tanimoto, E., Samejima, M., Sakai, F. and
Hayashi, T. (2003) Enhancement of growth by expression of poplar cellulase in Arabidopsis
thaliana. Plant Journal 33, 1099-1106.
Pauly, M., Gille, S., Liu, L., Mansoori, N., Souza, A., Schultink, A. and Xiong, G. (2013) Hemicellulose
biosynthesis. Planta, 1-16.
Pelham, H.R.B. (1990) The retention signal for soluble proteins of the endoplasmic reticulum. Trends
in Biochemical Sciences 15, 483-486.
Percival Zhang, Y.H., Himmel, M.E. and Mielenz, J.R. (2006) Outlook for cellulase improvement:
screening and selection strategies. Biotechnology Advances 24, 452-481.
Petersen, K. and Bock, R. (2011) High-level expression of a suite of thermostable cell wall-degrading
enzymes from the chloroplast genome. Plant Molecular Biology.
Peterson, R. and Nevalainen, H. (2012) Trichoderma reesei RUT-C30 – thirty years of strain
improvement. Microbiology 158, 58-68.
Pogorelko, G., Fursova, O., Lin, M., Pyle, E., Jass, J. and Zabotina, O. (2011) Post-synthetic
modification of plant cell walls by expression of microbial hydrolases in the apoplast. Plant
Molecular Biology 77, 433-445.
Popper, Z.A. (2008) Evolution and diversity of green plant cell walls. Current Opinion in Plant Biology
11, 286-292.
Pottkämper, J., Barthen, P., Ilmenberger, N., Schwaneberg, U., Schenk, A., Schulte, M., N, I. and
Streit, W. (2009) Applying metagenomics for the identification of bacterial cellulases that are
stable in ionic liquids. Green Chemistry 11, 957-965.
Ragauskas, A.J., Williams, C.K., Davison, B.H., Britovsek, G., Cairney, J., Eckert, C.A., Frederick,
W.J., Hallett, J.P., Leak, D.J., Liotta, C.L., Mielenz, J.R., Murphy, R., Templer, R. and
Tschaplinski, T. (2006) The path forward for biofuels and biomaterials. Science 311, 484-489.
Rahikainen, J., Mikander, S., Marjamaa, K., Tamminen, T., Lappas, A., Viikari, L. and Kruus, K. (2011)
Inhibition of enzymatic hydrolysis by residual lignins from softwood—study of enzyme binding
and inactivation on lignin-rich surface. Biotechnology and Bioengineering, n/a-n/a.
Ring, S.G. and Selvendran, R.R. (1981) An arabinogalactoxyloglucan from the cell wall of Solanum
tuberosum. Phytochemistry 20, 2511-2519.
Röder, J. (2010) Heterologe Expression des Cellulasegens sso1354 aus Sulfolobus solfataricus in
Nicotiana tabacum. BSc Thesis RWTH Aachen Univesity.
Rosgaard, L., Pedersen, S., Langston, J., Akerhielm, D., Cherry, J.R. and Meyer, A.S. (2007)
Evaluation of minimal Trichoderma reesei cellulase mixtures on differently pretreated Barley
straw substrates. Biotechnology Progress 23, 1270-1276.
Roslan, H.A., Salter, M.G., Wood, C.D., White, M.R.H., Croft, K.P., Robson, F., Coupland, G.,
Doonan, J., Laufs, P., Tomsett, A.B. and Caddick, M.X. (2001) Characterization of the

References
76
ethanol-inducible alc gene-expression system in Arabidopsis thaliana. The Plant Journal 28,
225-235.
Rudsander, U.J., Sandstrom, C., Piens, K., Master, E.R., Wilson, D.B., Brumer Iii, H., Kenne, L. and
Teeri, T.T. (2008) Comparative NMR analysis of cellooligosaccharide hydrolysis by GH9
bacterial and plant endo-1,4-β-glucanases. Biochemistry 47, 5235-5241.
Saddler, J.N. and Gregg, D.J. (1998) Ethanol production from forest product wastes. In Forest
Products Biotechnology; Bruce, A, Palfreyman, J W, Eds; Taylor & Francis Ltd: London, 1998,
183-207.
Saha, B.C., Iten, L.B., Cotta, M.A. and Wu, Y.V. (2005) Dilute acid pretreatment, enzymatic
saccharification and fermentation of wheat straw to ethanol. Process Biochemistry 40, 3693-
3700.
Sainz, M. (2011) Commercial Cellulosic Ethanol: The Role of Plant-Expressed Enzymes. In: Biofuels
(Tomes, D., Lakshmanan, P. and Songstad, D. eds), pp. 237-264. Springer New York.
Saloheimo, A., Lehtovaara, P., Penttila, M., Teeri, T., Stahlberg, J., Johansson, G., Pettersson, G.,
Claeyssens, M., Tomme, P. and Knowles, J. (1988) EGIII, a new endoglucanase from
Trichoderma reesei: The characterization of both gene and enzyme. Gene 63, 11-21.
Salter, M.G., Paine, J.A., Riddell, K.V., Jepson, I., Greenland, A.J., Caddick, M.X. and Tomsett, A.B.
(1998) Characterisation of the ethanol-induciblealcgene expression system for transgenic
plants. The Plant Journal 16, 127-132.
Sambrook, J., Fritsch, E. and Maniatis, T. (1989) Molecular Cloning - A Laboratory Manual.
Sánchez, C. (2009) Lignocellulosic residues: Biodegradation and bioconversion by fungi.
Biotechnology Advances 27, 185-194.
Saville, B.A. (2011) Pretreatment Options. In: Plant Biomass Conversion pp. 199-226. John Wiley &
Sons, Inc.
Scheller, H.V. and Ulvskov, P. (2010) Hemicelluloses. Annual Review of Plant Biology 61, 263-289.
Schirmacher, J. (2006) Ein Ethanol induzierbarer Promotor zur Reduktion der Photorespiration in C3-
Pflanzen, Diplomarbeit. Institut für Biologie 1, RWTH Aachen.
Scott, T.A. and Melvin, E.H. (1953) Determination of dextran with anthrone. Analytical Chemistry 25,
1656-1661.
Scotti, N., Rigano, M.M. and Cardi, T. (2012) Production of foreign proteins using plastid
transformation. Biotechnology Advances 30, 387-397.
She, Q., Singh, R.K., Confalonieri, F., Zivanovic, Y., Allard, G., Awayez, M.J., Chan-Weiher, C.C.-Y.,
Clausen, I.G., Curtis, B.A., De Moors, A., Erauso, G., Fletcher, C., Gordon, P.M.K., Heikamp-
de Jong, I., Jeffries, A.C., Kozera, C.J., Medina, N., Peng, X., Thi-Ngoc, H.P., Redder, P.,
Schenk, M.E., Theriault, C., Tolstrup, N., Charlebois, R.L., Doolittle, W.F., Duguet, M.,
Gaasterland, T., Garrett, R.A., Ragan, M.A., Sensen, C.W. and Van der Oost, J. (2001) The
complete genome of the crenarchaeon Sulfolobus solfataricus P2. Proceedings of the National
Academy of Sciences 98, 7835-7840.
Shen, B., Sun, X., Zuo, X., Shilling, T., Apgar, J., Ross, M., Bougri, O., Samoylov, V., Parker, M.,
Hancock, E., Lucero, H., Gray, B., Ekborg, N.A., Zhang, D., Johnson, J.C.S., Lazar, G. and

References
77
Raab, R.M. (2012) Engineering a thermoregulated intein-modified xylanase into maize for
consolidated lignocellulosic biomass processing. Nature Biotechnology 30, 1131-1136.
Shen, W.-j. and Forde, B.G. (1989) Efficient transformation of Agrobacterium spp. by high voltage
electroporation. Nucleic Acids Research 17, 8385.
Shoseyov, O., Shani, Z. and Levy, I. (2006) Carbohydrate binding modules: biochemical properties
and novel applications. Microbiology and Molecular Biology Reviews 70, 283-295.
Staehelin, L.A. and Moore, I. (1995) The plant golgi apparatus: Structure, functional organization and
trafficking mechanisms. Annual Review of Plant Physiology and Plant Molecular Biology 46,
261-288.
Sticklen, M. (2006) Plant genetic engineering to improve biomass characteristics for biofuels. Current
Opinion in Biotechnology 17, 315-319.
Streatfield, S. (2007) Approaches to achieve high-level heterologous protein production in plants.
Plant Biotechnol J 5, 2 - 15.
Sun, F. and Chen, H. (2008) Enhanced enzymatic hydrolysis of wheat straw by aqueous glycerol
pretreatment. Bioresource Technology 99, 6156-6161.
Sun, Y. and Cheng, J. (2002) Hydrolysis of lignocellulosic materials for ethanol production: a review.
Bioresource Technology 83, 1-11.
Sun, Y., Cheng, J.J., Himmel, M.E., Skory, C.D., Adney, W.S., Thomas, S.R., Tisserat, B., Nishimura,
Y. and Yamamoto, Y.T. (2007) Expression and characterization of Acidothermus cellulolyticus
E1 endoglucanase in transgenic duckweed Lemna minor 8627. Bioresource Technology 98,
2866-2872.
Takahashi, J., Rudsander, U.J., Hedenström, M., Banasiak, A., Harholt, J., Amelot, N., Immerzeel, P.,
Ryden, P., Endo, S., Ibatullin, F.M., Brumer, H., del Campillo, E., Master, E.R., Vibe Scheller,
H., Sundberg, B., Teeri, T.T. and Mellerowicz, E.J. (2009) KORRIGAN1 and its Aspen
homolog PttCel9A1 decrease cellulose crystallinity in Arabidopsis stems. Plant and Cell
Physiology 50, 1099-1115.
Taylor, L.E., 2nd, Dai, Z., Decker, S.R., Brunecky, R., Adney, W.S., Ding, S.Y. and Himmel, M.E.
(2008) Heterologous expression of glycosyl hydrolases in planta: a new departure for biofuels.
Trends in Biotechnology 26, 413-424.
Teymouri, F., Alizadeh, H., Laureano-Pérez, L., Dale, B. and Sticklen, M. (2004) Effects of ammonia
fiber explosion treatment on activity of endoglucanase from Acidothermus cellulolyticus in
transgenic plant. Applied Biochemistry and Biotechnology - 116.
Tilman, D., Reich, P.B. and Knops, J.M.H. (2006) Biodiversity and ecosystem stability in a decade-
long grassland experiment. Nature 441, 629-632.
Tomme, P., Van Tilbeurgh, H., Pettersson, G., Van Damme, J., Vandekerckhove, J., Knowles, J.,
Teeri, T. and Claeyssens, M. (1988) Studies of the cellulolytic system of Trichoderma reesei
QM 9414. Analysis of domain function in two cellobiohydrolases by limited proteolysis.
European Journal of Biochemistry 170, 575-581.
Tomsett, B., Tregova, A., Garoosi, A. and Caddick, M. (2004) Ethanol-inducible gene expression: first
step towards a new green revolution? Trends in Plant Science 9, 159-161.

References
78
Turner, M.B., Spear, S.K., Huddleston, J.G., Holbrey, J.D. and Rogers, R.D. (2003) Ionic liquid salt-
induced inactivation and unfolding of cellulase from Trichoderma reesei. Green Chemistry 5,
443-447.
Turner, P., Mamo, G. and Karlsson, E. (2007) Potential and utilization of thermophiles and
thermostable enzymes in biorefining. Microbial Cell Factories 6, 9.
Tyner, W.E. (2008) The US ethanol and biofuels boom: Its origins, current status, and future
prospects. BioScience 58, 646-653.
Unsworth, L.D., Van Der Oost, J. and Koutsopoulos, S. (2007) Hyperthermophilic enzymes − stability,
activity and implementation strategies for high temperature applications. FEBS Journal 274,
4044-4056.
Updegraff, D.M. (1969a) Semimicro determination of cellulose in biological materials. Analytical
Biochemistry 32, 420-424.
Updegraff, D.M. (1969b) Semimicro determination of cellulose in biological materials. Anal Biochem
32, 420-424.
Van Dyk, J.S. and Pletschke, B.I. (2012) A review of lignocellulose bioconversion using enzymatic
hydrolysis and synergistic cooperation between enzymes—Factors affecting enzymes,
conversion and synergy. Biotechnology Advances 30, 1458-1480.
van Tilbeurgh, H., Tomme, P., Claeyssens, M., Bhikabhai, R. and Pettersson, G. (1986) Limited
proteolysis of the cellobiohydrolase I from Trichoderma reesei: Separation of functional
domains FEBS Letters 204, 223-227.
Vancov, T., Alston, A.-S., Brown, T. and McIntosh, S. (2012) Use of ionic liquids in converting
lignocellulosic material to biofuels. Renewable Energy 45, 1-6.
Vanholme, R., Morreel, K., Ralph, J. and Boerjan, W. (2008) Lignin engineering. Current Opinion in
Plant Biology 11, 278-285.
Verma, D., Jin, S., Kanagaraj, A., Singh, N.D., Daniel, J., Kolattukudy, P.E., Miller, M. and Daniell, H.
(2013) Expression of fungal cutinase and swollenin in tobacco chloroplasts reveals novel
enzyme functions and/or substrates. PLoS One 8, e57187.
Verma, D., Kanagaraj, A., Jin, S., Singh, N.D., Kolattukudy, P.E. and Daniell, H. (2010) Chloroplast-
derived enzyme cocktails hydrolyse lignocellulosic biomass and release fermentable sugars.
Plant Biotechnology Journal.
Vitousek, P.M., Mooney, H.A., Lubchenco, J. and Melillo, J.M. (1997) Human domination of earth's
ecosystems. Science 277, 494-499.
Vogel, K.P. and Jung, H.-J.G. (2001) Genetic modification of herbaceous plants for feed and fuel.
Critical Reviews in Plant Sciences 20, 15-49.
Voget, S., Steele, H.L. and Streit, W.R. (2006) Characterization of a metagenome-derived halotolerant
cellulase. Journal of Biotechnology 126, 26-36.
Werthmann, V. (2009) Untersuchungen zur Expression der Cellulase EGL2 und CBH2 aus Hypocrea
jecorina in Pflanzen. BSc Thesis RWTH Aachen Univesity.
Xue, G.P., Patel, M., Johnson, J.S., Smyth, D.J. and Vickers, C.E. (2003) Selectable marker-free
transgenic barley producing a high level of cellulase (1,4-beta-glucanase) in developing
grains. Plant Cell Reports 21, 1088-1094.

References
79
Yang, B. and Wyman, C.E. (2008) Pretreatment: the key to unlocking low-cost cellulosic ethanol.
Biofuels, Bioproducts and Biorefining 2, 26-40.
Zavrel, M., Bross, D., Funke, M., Büchs, J. and Spiess, A.C. (2009) High-throughput screening for
ionic liquids dissolving (ligno-)cellulose. Bioresource Technology 100, 2580-2587.
Zhang, D., VanFossen, A., Pagano, R., Johnson, J., Parker, M., Pan, S., Gray, B., Hancock, E.,
Hagen, D., Lucero, H., Shen, B., Lessard, P., Ely, C., Moriarty, M., Ekborg, N., Bougri, O.,
Samoylov, V., Lazar, G. and Raab, R.M. (2011) Consolidated pretreatment and hydrolysis of
plant biomass expressing cell wall degrading enzymes. Bioenerg Res 4, 276-286.
Zhang, Y.-H.P. and Lynd, L.R. (2004) Toward an aggregated understanding of enzymatic hydrolysis of
cellulose: Noncomplexed cellulase systems. Biotechnology and Bioengineering 88, 797-824.
Ziegelhoffer, T., Raasch, J.A. and Austin-Phillips, S. (2001) Dramatic effects of truncation and sub-
cellular targeting on the accumulation of recombinant microbial cellulase in tobacco. Molecular
Breeding 8, 147-158.
Ziegelhoffer, T., Raasch, J.A. and Austin-Phillips, S. (2009) Expression of Acidothermus cellulolyticus
E1 endo-beta-1,4-glucanase catalytic domain in transplastomic tobacco. Plant Biotechnology
Journal 7, 527-536.
Ziegelhoffer, T., Will, J. and Austin-Phillips, S. (1999) Expression of bacterial cellulase genes in
transgenic alfalfa (Medicago sativa L.), potato (Solanum tuberosum L.) and tobacco
(Nicotiana tabacum L.) Molecular Breeding 5, 309-318.
Žifčáková, L. and Baldrian, P. (2012) Fungal polysaccharide monooxygenases: new players in the
decomposition of cellulose. Fungal Ecology 5, 481-489.

Appendix
80
Appendix
Abbreviations
4MUC 4-methylumbelliferyl cellobioside
Ac acetate
AFEX ammonia fiber expulsion
AIR alcohol insoluble residues
BGL beta-glucosidase
CaMV cauliflower mosaic virus
CBH cellobiohydrolase
CBM carbohydrate binding module
cd catalytic domain
CDH cellobiosdehydrogenase
CMC carboxymethylcellulose
Da Dalton
DMP dimethylphosphate
DMSO dimethylsulfoxide
EB energy balance
EG endoglucanase
EMIM 1-ethyl-3-methylimidazolium
ER endoplasmic reticulum
FAEA ferulic acid esterase A
FITC fluorescein isothiocyanate
GAX glucoronoarabinoxylan
GC gas chromatography
GH glycosyl hydrolase
GT glycosyl transferase
GX glucoronoxylan
IL ionic liquid
M molar
MAMP microbe associated molecular pattern
MMIM 1,3-dimethylimidazolium
MS mass spectrometry
MU methylumbelliferone
ORF open reading frame
PAGE polyacrylamide gel electrophorese
PAHBAH 4-hydroxybenzoic acid hydrazide
PAL phenylalanine ammonium lyase
PCWS plant cell wall polysaccharides
PGA polygalacturonic acid

Appendix
81
PMO polysaccharide monooxygenases
PMSF phenylmethylsulfonyl fluoride
RG I rhamnogalacturonan I
rpm revolutions per minute
SDS sodium dodecylsufate
TSP total soluble protein
UDP uridine diphosphate
UTR untranslated region
v volume
w weight
XyG xyloglucan

Appendix
82
Register of equipment
Name Type Manufacturer
"-20°C premium "-20°C freezer Liebherr, Germany
Herafreeze "-80°C freezer Heraeus, Germany
Infinite 200 96-well plat reader Tecan, Germany
Waring blender blender Warring, USA
Allegra-X15R centrifuge Beckmann Coulter, Germany
Avanti J 301 centrifuge Beckmann Coulter, Germany
5414 D centrifuge Eppendorf, Germany
Mikrofuge R centrifuge Beckmann Coulter, Germany
Multiporator electroporation device Eppendorf, Germany
Herolab E.A.S.Y Win32 gel documentation Herolab GmbH, Germany
Grant QBT heating device Cambridge, GB
Heraeus B12 incubator Heraeus, Germany
Innova 4340 incubator/shaker New Brunswick Scinetific, Germany
Kuhner ShakerX incubator/shaker Kuhner AG, CH
Leica DRM+DCF microscope Leica, Germany
Olympus Bx4I microscope Olympus, Germany
Leica VT1000S vibratome Leica, Germany
Leica CM3050S cryo microtome Leica, Germany
Primus 96 Plus PCR cycler Eurofins MWG Operon, Germany
Masterflex L/S peristaltic pump Cole Parmer, Germany
Nikon CoolPix 5400 photographic camera Niko, Germany
Biophotometer photometer Eppendorf, Germany
Heraeus HS9 sterile bench Heraeus, Germany
Ikamag REC-G stirrer IKA Labortechnik, Germany
Thermomixer comfort temperature controlled mixer Eppendorf, Germany
Herolab UVT-20M 302nm UV transilluminator Herolab GmbH, Germany
G-560E vortex Scientific inductries, USA

Appendix
83
Table of figures
Figure 1: Global demographic development. .......................................................................................... 2
Figure 2: The primary cell wall of a growing cell as found in most flowering plants. ............................... 4
Figure 3: Schematic representation of hemicellulose polysaccharides................................................... 6
Figure 4: Current overview on fungal enzymatic degradation of cellulose. ............................................. 9
Figure 5: Schematic presentation of plant expression cassettes for the differential targeting of TrCel5A.
........................................................................................................................................................ 18
Figure 6: Transient expression of TrCel5A in N. tabacum. ................................................................... 19
Figure 7: Transient expression of TrCel5A-mCherry fusion in tobacco. ............................................... 20
Figure 8: Localization of transiently-expressed TrCel5A-mCherry fusion in 90-µm sections of tobacco
leaf tissue. ..................................................................................................................................... 20
Figure 9: Analysis of transgenic tobacco expressing TrCel5A. ............................................................. 22
Figure 10: Enzymatic activity of TrCel5A at different pH values and temperatures. ............................. 23
Figure 11: Subcellular localization of recombinant TrCel5AER in transgenic tobacco cells. .................. 24
Figure 12: Subcellular localization of recombinant TrCel5A in transgenic tobacco cells. ..................... 25
Figure 13: Transgenic tobacco plants expressing TrCel5A. ................................................................. 26
Figure 14: Phenotypic analysis of transgenic tobacco plants. .............................................................. 27
Figure 15: Enzymatic hydrolysis of transgenic tobacco leaves. ............................................................ 29
Figure 16: Generation and analysis of transgenic tobacco expressing TrCel5A under the control of an
alcohol-inducible promoter. ............................................................................................................ 36
Figure 17: Inducible TrCel5A expression in tobacco plants. ................................................................. 37
Figure 18: Phenotype of transgenic tobacco plants. ............................................................................. 39
Figure 19: Crystalline cellulose content in transgenic tobacco plants expressing TrCel5A. ................. 40
Figure 20: Schematic representation of the SSO1354 expression cassettes including an N-terminal
His6 tag. .......................................................................................................................................... 44
Figure 21: Substrate SDS-PAGE in the presence of 0.15% (w/v) CMC and extracts from different
SSO1354 transgenic plants. .......................................................................................................... 45
Figure 22: Tobacco plants expressing active SSO1354. ...................................................................... 46
Figure 23: Biochemical characterization of plant-derived SSO1354. .................................................... 47
Figure 24: Activity of SSO1354 in high concentrations of ionic liquids. ................................................ 49
Figure 25: Expression vector pTRAkc-rfp-his6-ERH for heterologous expression and ER localization in
plant cells. ...................................................................................................................................... 58

Appendix
84
Scientific publications
Publications
Klose H, Röder J, Girfoglio M, Fischer R, Commandeur U (2012) Hpyerthermophilic
endoglucanase for in planta lignocellulose conversion. Biotechnology for Biofuels 5:
63.
Klose H, Günl M, Usadel B, Fischer R, Commandeur U. (2013) Ethanol inducible
expression of a mesophilic cellulase avoids adverse effects on plant development.
Biotechnology for Biofuels, 6:53.
Garvey M, Klose H, Fischer R, Lambertz C, Commandeur U (2013) Cellulases for
biomass degradation: comparing recombinant cellulase expression platforms. Trends
in Biotechnology, 10: 581-593.
Garvey M, Klinger J, Klose H, Fischer R, Commandeur U (2013) Expression of
recombinant cellulase Cel5A from Trichoderma reesei in tobacco plants. Journal of
Visualized Experiments, in press.
Poster presentations
Klose H, Wennekamp S, Commandeur U (2011) Heterologous expression of fungal
cellulases in plants. First International Working on Walls (WoW) Symposium, 01.-
03.2011, Vancouver, Canada.
Klose H, Röder J, Girfoglio M, Fischer R, Commandeur U (2011) In planta
expression of hyperthermophilic β-glucanase. 10th International Symposium on
Biocatalysis, 02.-06.10.2011, Giardini Naxos (ME), Italy
Klose H, Röder J, Girfoglio M, Fischer R, Commandeur U (2012) Expression of
hyperthermophilic β-glucanase in plants. NSERC Bioconversion Network
Pretreatment Workshop, 04.-06.06.2012, Vancouver, Canada

Appendix
85
Klose H, Klinger J, Röder J, Girfoglio M, Fischer R, Commandeur U (2012)
Hyperthermophilic β-glucanase for advanced in planta biomass conversion. 6th
International Congress on Biocatalysis, 02.-06.10.2012, Hamburg
Schmidt T, Przyklenk M, Girfoglio M, Becker J, Klose H, Fischer R, Commandeur U
(2012) Synergistic effects of combined cellulolytic enzymes active in pure ionic
liquids. 6th International Congress on Biocatalysis, 02.-06.2012, Hamburg.
Klose H, Lambertz C, Günl M, Commandeur U (2013) Inducible expression of
mesophilic cellulases in tobacco. 9th International Conference on Renewable
Resources & Biorefineries, 05.-07.06.2013, Antwerp, Belgium

Appendix
86
Acknowledgement
The work of the last years was a magnificent and challenging experience for me.
During this time many people supported me, both directly and indirectly, with my
academic career. Here, I would like to thank them for their precious help.
I am grateful to Prof. Dr. Rainer Fischer for the opportunity to work at the Institute for
Molecular Biotechnology and his support during my PhD-work.
Dr. Ulrich Commandeur I would like to thank for the allocation of this interesting topic
and for all the lively debates.
To Prof. Dr Björn Usadel I am thankful for the willingness to take the co-review of my
thesis and all the valuable aid in understanding cell wall chemistry.
A big thank goes to Dr. Flora Schuster for her excellent work in the plant cell culture,
Dr. Marcus Günl for his help with the measurement of cell wall sugars, Ibrahim
Alameidi for his care in cultivating the plants, Maria Krämer and Dr. Dmitrij
Hristodorov for their kind help with my histological experiments, Dr. Richard Twyman
for the critical review of my thesis, Dr. Michele Girfoglio for his help with the
extremophilic enzymes and all the delightful discussions (not only on science), Dr.
Megan Garvey for her precious aid with my manuscripts and her invaluable
willingness for scientific discussion especially in the last months.
I am grateful to Karo Richter and Tini Dickmeis not only for sharing office and lab with
me in the last five years but also being such wonderful colleagues!
A big thanks goes to all the coworkers of the Institute (present and former) with whom
I have spent such great times in the last years!
All this work would not have been possible without the constant aid and
understanding from my family, especially my parents, to whom I would like to express
my deepest gratitude.

Appendix
87
Declaration
I hereby declare that all information in this document has been obtained and
presented in accordance with academic rules and ethical conduct. I also declare that,
as required by these rules and conduct, I have fully cited and referenced all material
and results that are not original to this work.
_____________________________
Holger Klose, Aachen 18.12.2013

Appendix
88
Curriculum vitae
Personal data
Name: Holger Klose
Gender: Male
Date of birth: 17th Aug, 1981
Place of birth: Lüdenscheid
Nationality: German
Marital status: Single
Employment
01/2014 Institute of Biology I (Botany/Molecular Genetics), RWTH
Aachen University, Aachen
Doctorate
11/2009 – 12/2013 RWTH Aachen University, Aachen,
Doctoral studies: Biotechnology
Education
10/2007 – 10/2009 RWTH Aachen University, Aachen, Germany
Master of Science in Biotechnology/Molecular
Biotechnology
10/2002 – 09/2007 RWTH Aachen University, Aachen, Germany
Bachelor of Science in Biotechnology/Molecular
Biotechnology
08/1992 – 06/2001 Bergstadt Gymnasium, Lüdenscheid, Germany