intertidal invertebrate and algal communities on the rocky shores of the bay of morbihan, kerguelen...
TRANSCRIPT

P.S.Z.N. I: Marine Ecology, 8 (3): 207-220 (1987) 0 1987 Paul Parey Scientific Publishers, Berlin and Hamburg ISSN 0173-9565
Accepted: March 26,1987
Intertidal Invertebrate and Algal Com- munities on the Rocky Shores of the Bay of Morbihan, Kerguelen (South Indian Ocean) JOHN M. LAWRENCE & JAMES B. MCCLXNTOCK'
Department of Biology, University of South Florida, Tampa, Florida 33620, U. S. A.
I Present address: Department of Biology, University of Alabama at Birmingham; Birmingham, Alabama 35294, USA.
With 5 figures and 4 tables
Key words: Intertidal, communities, Kerguelen, South Indian Ocean.
Abstract. Both the emergent surfaces and tide pools of the rocky intertidal shores of the Bay of Morbihan, Kerguelen are depauperate in number of macroinvertebrate and macroalgal species. Diversity (H') was high and similar for macroinvertebrates in tide pools from the high to low intertidal and on the emergent surface exposed at extreme low-tide when calculated in terms of numbers of individuals. H' was lower when calculated in terms of energy and twice as high for the tide pool in the high intertidal than the lower tide-pools and the emergent surface. H' for macroalgae was 0 in the highest tide-pool and highest in the lower tide-pools. The densitykpecies- rank curves for the macroinvertebrates and the macroalgae are logarithmic but vary in linearity. Algal biomass was 37 % of the macroinvertebrate biomass on the emergent surface at the extreme low-tide, but 106% in the highest tide-pool. Twenty-three months after denudation of the tide pools, the ratios of total densities of the macroinvertebrates (ind . m-l) to the original densities were 48 % for the pool in the high intertidal, 25 ?6 for the middle tide-pool, and 4 % for the pool in the low intertidal. The diversity indexes for the macroinvertebrates were similar to the original ones. Recolonization at Kerguelen is slow and shows an intertidal gradient. The composition of the rocky intertidal community at Kerguelen results from the rigorous physical environment and the small number and functional types of species. In the absence of major predators on macroinvertebrates and of herbivores on macroalgae, interaction among the species present seems to be competitive in nature.
Problem
PAINE (1966) pointed out that although longitudinal and latitudinal gradients are ' well known geographically, they are poorly known ecologically. Most of the studies of the communities of the rocky intertidal have been done on north- temperate (CONNELL, 1972; DAYTON, 1971,1975; LUBCHENCO, 1978; LUBCHENCO & MENGE. 1975; MENGE, 1976; PAINE, 1966,1974) and south-temperate (BERRY,
U. S . Copyright Clearance Center Code Statement: 0173-9565/87/0803-0207$02.50/0

208 LAWRENCE & MCCLINTOCK
1982; JAM & MORENO, 1984; MAY er al., 1970; MCQUAID & BRANCH, 1984; MORENO & JARAMILLO, 1983; PAINE, 1971; UNDERWOOD, 1981; UNDERWOOD & JERNAKOFF, 1984) continental coasts. These regions are productive, with an abundance of species and individuals, and with many functional types (plants and animals) in common.
In addition to the preponderance of information about communities of temperate regions, almost all information pertains to emergent surfaces. The tide pools that frequently occur in the rocky intertidal are rarely studied. It is known that tide pools extend the distribution of species upwards, but that there is still an increased stress associated with increased height of the pools so that intertidal zonation occurs with tide pools as with the emergent surfaces (DOTY, 1957; LEWIS, 1964; NEWELL, 1979; STEPHENSON & STEPHENSON, 1972). These stresses can be qualitatively different from those associated with emergent surfaces (e. g., pH, oxygen concentration, salinity) and can have interactive effects with each other and with temperature (NEWELL, 1979). These physical factors have been considered to control primarily the abundance and distribu- tion of algal species in tide pools, but both biological and physical controls exist (LUBCHENCO, 1978; PAINE & VADAS, 1969; SZE, 1980; UNDERWOOD & JERNAKOFF, 1984).
The island of Kerguelen is isolated in the South Indian Ocean and subject to rigorous oceanic conditions. Its marine flora and fauna are known ~(ARNAUD, 1971) and the general zonation of the emergent surfaces and tide pools of the rocky intertidal has been described qualitatively (ARNAUD, 1974; BELLIDO, 1981; DELEPINE, 1963). Basic functional types of invertebrates which can control community structure (e. g. asteroid carnivores such as Pisaster and echinoid herbivores such as Srrongylocenfrorus) are missing from the rocky intertidal zone. Many of the algae found in the rocky intertidal of Chile and Southern Australia (JARA & MORENO, 1984; UNDERWOOD, 1981) are also absent at Kerguelen.
The purpose of the present paper is to quantitatively describe the macroin- vertebrate and algal components of the communities of the tide pools and emergent surfaces that occur at different levels of the rocky intertidal zone at Kerguelen. The basic question was whether the rigorous physical environment and the suite of species which exists at Kerguelen affects density, species richness, and species diversity.
Material and Methods
1. Study site
The Kerguelen archipelago (495 , 70"E) is situated in the South Indian Ocean just within the antarctic convergence, approximately 4000 km southeast of Africa and 2000 km north of the Antarctic continent (Fig. 1). Grande Terre is the largest of over 300 islands which comprise the archipelago, and is ca. 140 km in width and length. The island is of volcanic origin., composed primarily of basalt which has been extensively modified by glaciation. The extreme isolation of the island contributes to its oceanic climate. Winds have a mean annual speed of 10m . sec-I and may reach speeds as great as 62 m . sec-' (ARNAUD, 1974). The number of d . a-' with winds exceeding 16m . sec-' can be as high as 333 (ARNAUD, 1974). The weather is typically cloudy with light rain 13 to 25 days each month and monthly precipitation of 27 to 117mm (HUREAU, 1970; ARNAUD, 1974).

69'E
Fig. 1. Map of Kerguelen. The ar- row points to Port-aux-Frangais in the Bay of Morbihan.
70'E
I ,$ KERGUELEN
I I Snow and hail fall at sea level, more frequently in winter but even in the austral summer. The air temperature is moderate at sea level, but may be as high as 20°C during the summer and, although rarely below O'C, as low as -8°C during the winter (HUREAU, 1970; ARNAUD. 1974).
The present study was conducted in the Bay of Morbihan, an extensive (ca. 100 km') semi- protected bay at the eastern end of the island. The shoreline of the bay has a variety of habitats including sand and cobble beaches, fjords, and extensive rocky-intertidal areas. Tidal cycles occur twlcr a day with an amplitude ranging from 30 to 210cm (HUREAU, 1970). Surface seawater temperatures range from 5 . 5 T in the summer to 2.1"C in the winter; surface salinities range from 31.9 to 33.6%0 (HUREAU, 1970). Although the bay is protected from open ocean swetls, high winds frequently generate surf conditions, particularly on the northern side which faces the prevailing winds.
The site of the transect was at the west end of the Promenade des Amerlocks, west of the Pte. de la Baignade (Fig. 1) on the northern coast of the bay, west of Port-aux-Frangais. The transect has the general features described for similar intertidal areas by DELdPrNE (1963), ARNAUD (1974), and BELLIDO (1981). Prevailing winds and waves strike this coast so that extremely high-energy wave action can occur. Occasional periods of dead calm result in the absence of wave activity. The transect (Figs. 2-4) was a broad, gently sloping rock shelf of ca. 45 m between extreme high-tide (EHT) and extreme low tide (ELT) (ca. 2M)cm vertical height). There were five stations on the transect including three tide-pools and two emergent surfaces.
In her study on the algae of tide pools, LUECHENCO (1978) selected three pools because they subjectively appeared to represent two extremes of the continuum in types of algae present. We selected three representative tide-pools at different levels because they were similar in size but showed distinct differences in-plant and animal composition. Pool 1 was closest to the edge of the shelf, 125 x 150cm (18,750cm') in area and 37cm in depth. Pool 2 was the middle tide-pool, 200 x 225cm (45,000cmZ) in area and 16cm in depth. Pool 3 was the highest tide-pool, 200 X 125cm (25,000 cm') in area and 6 cm in depth. At an ELT during moderate wave activity Pools 1 and 2 were isolated from the sea for ca. 3 h 30 min while Pool 3, because of orientation and neighboring relief, was isolated from the sea for only ca. 1 h. Maximal temperatures reached during these periods were 12.5"C, 17"C, and 17.5"C for Pools 1 , 2 , and 3, respectively. Minimal temperature during isolation at low tide in the austral winter 1983 were 1.5"C and 2°C for Pools 1 and 2, respectively (F. GUILBERT, pers. comm.). No ice forms in the pools during the winter.
One emergent-surface station was on the shelf below the lowest tide-pool and was exposed only at ELT. The second emergent-surface station was adjacent to the middle tide-pool and was exposed at moderate low tides.
2. Sampling and analyses
As UNDERWOOD (1981) pointed out, his and other similar studies on intertidal community structure restricted sampling to those species that were common, reliably identified, and quantified. The

210 LAWRENCE & MCCLINTOCK
EHT E LT
. P O O L 3 POOL 2 POOL 1
BOULDERS (depth 6 cm) (depth 1 6 c m ) ( d e p t h 3 7 c m ) SHELF
I , I I -t---b
(m) o 10 za 30 40 5 0
Fig. 2. Diagram of the transect across the rocky intertidal on the Promenade des Amerlocks west of Pte. de Baignade, Kerguelen showing the location of the stations, extreme high tide (EHT) and extreme low tide (ELT); height ca. 200cm.
numbers and size-frequency distributions of all macroinvertebrates except the numerous, very motile isopods were recorded at the species level in January 1983. The meiofauna of the tide pools (halicariens, tanaiids, small polychaetes, and nematodes; ARNAUD, 1974) were not considered.
All macroifiertebrates in Pools 1 and 3 were counted and measured in both Janu:iry 1983 and December 1984. Pool 2 was divided into 2.5 X 2.5 cm quadrats and the numbers and sizes of all macroinvertebrates in 22 alternating quadrats were recorded in January 1983. As dono by LUB- CHENCO (1978) we did not attempt replicate quadrats within each tide pool. There are distinct microhabitats within tide pools (the edge of pool; the surfaces, edges, and undersides of submerged rocks within the pools; see PYEFINCH, 1943; D o n , 1957) to a much greater extent than on emergent intertidal surfaces. We decided it would be more appropriate to consider the organismal composi- tion of the tide pools as a whole as done by PAINE & VADAS (1969). All macroinvertebrates in Pool 2 were sampled in December 1984. All macroinvertebrates present in two and four 25 X 25cm quadrats on the lower and upper emergent-surface stations, respectively, were counted and measured only in 1983.
Individuals of all macroinvertebrate species over the size-ranges observed were collected and weighed to obtain size-weight regressions. These were used with the censuses and size-frequency distributions to obtain the biomass present for each species.
The algae were collected only in January 1983. The area of the tide pools and the lower emergent-surface station occupied by the encrusting alga Hildenbrandin lecanellieri wiis estimated. The biomass of H . lecanellieri from a 15 X 15 cm quadrat where the alga formed a continuous cover was used to calculate its total biomass in the tide pools. All other algae were completely removed from the three tide-pools and the lower emergent-surface station and weighed by species to obtain their biomass. No algae occurred on the higher emergent-surface station.
The macroinvertebrates and algae were lyophilized and homogenized. Their proximate composi- tion (ash, lipid, protein, carbohydrate) was measured by the methods used by LAWRENCE (1973). Energy equivalents were calculated using the energy coefficients of BRODY (1945).
Species richness (S) is considered to be the number of species present. Diversity was calculated
by the SHANNON-WIENER index (H’ = f pi . Inp,, where s = the number of species and pi =
the proportion of the total number of individuals consisting of the ith species) for each station on the basis of numbers of individuals for the macroinvertebrates and on the basis of kJ for both macroinvertebrates and algae. The indexes were calculated on the basis of species and riot on higher taxonomic or functional groups.
1-1

Fip.3. View of the transect from above at ELT.
Fig. 4. Rock shelf at ELT.

212 LAWRENCE B MCCWOCK
Results
1. Description of the communities in January 1!383
The species list and the numerical and energetic densities of the macroinverte- brates are given in Table 1. Only seven species occurred on the transect: five were suspension feeders (two species of bivalves, two species of holothuroids, and an anemone species), and two were grazers (one limpet and one chiton species). The higher emergent-surface station contained only the grazing limpet Kerguelenella lateralis. In contrast, five species occurred in the lower emergent- surface station. The species richness of the three tide-pools was similar to that of the lower emergent-surface station. The most evident zonation watj the inverse relation between the densities of Aulacomya ater and Mytilus edulis, particularly if one considers the lower emergent-shelf station as a continuation of the zonation. In addition, the holothuroids Pseudocnus laevigatus and F'entacucumis bouvetensis were essentially restricted to the lower emergent-shelf station.
Despite the equivalent species richness of the three tide-pools and the lower emergent-surface station, there was a gradient in density, being lowest in the upper tide-pool and highest in the lower emergent-surface station. The gradient
Table 1. Numerical (ind . m-') and energetic (kJ . m-') densities and species richness (S) of invertebrates in the tide pools and on the emergent surfaces of the rocky intertidal at Kerguelen in January 1983.
Species Upper Middle Lower Lower Upper Tide Pool Tide Pool Tide Pool Emergent Emergent (3) (2) (1) Surface Surface
Numerical density (ind - m-I)
Aulacomya ater Mytilus edulis desolationb Pseudocnus laevigam Pentacuczimis bouvetensis Anemone sp. Heeniarthrum setulosum Kerguelenella lateralis sun1 Species richness ( S ) Energetic density (kJ . d) Aulacomya ater Mytilus edu2i.s desoiationis Pseudocnus laevigam Pentacucumis bouvetensis Anemone sp. Hemiarthrurn setulosum Kerguelenella lateralis Sum
6 86 0 0
58 23 10
183 5
37.3 490.9
0 0
116.8 0.8 7.5
653.3
1.02 10 0 0
13 259 58
442 5
1386.1 42.7 0 0
25.5 9.2
43.9 1507.4
259 8 4 6
128 29 0
434 6
3301.1 115.5
1.7 1.7
256.9 0.8 0
3677.7
1120 0
176 136 496 88 0
2016 5
12332.8 0
72.0 41.0
995.2 3.3 0
13144.7
0 0 0 0 0
53
53 1
0 0 0 0 0 0
40.1 40.1
9
Intertidal invertebrate and algal communities 213
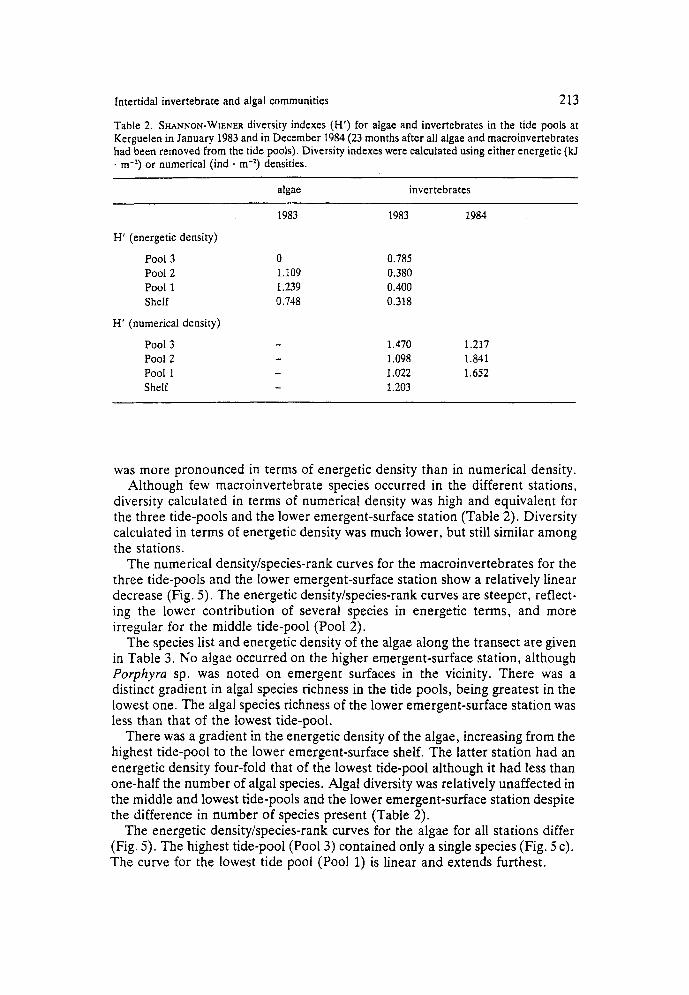
Table 2. SHANNON-WIENER diversity indexes (H’) for algae and invertebrates in the tide pools at Kerguelen in January 1983 and in December 1984 (23 months after all algae and macroinvertebrates had been removed from the tide pools). Diversity indexes were calculated using either energetic (ki . rn-z) or numerical (ind * m-*) densities.
algae invertebrates
H‘ (energetic density)
Pool 3 Pool 2 Pool 1 Shelf
H’ (numerical density)
Pool 3 Pool 2 Pool 1 Shelf
1983
0 1.109 1.239 0.748
1983 1984
0.785 0.380 0.400 0.318
1.470 1.217 1.098 1.831 1.022 1.652 1.203
was more pronounced in terms of energetic density than in numerical density. Although few macroinvertebrate species occurred in the different stations,
diversity calculated in terms of numerical density was high and equivalent for the three tide-pools and the lower emergent-surface station (Table 2). Diversity calculated in terms of energetic density was much lower, but still similar among the stations.
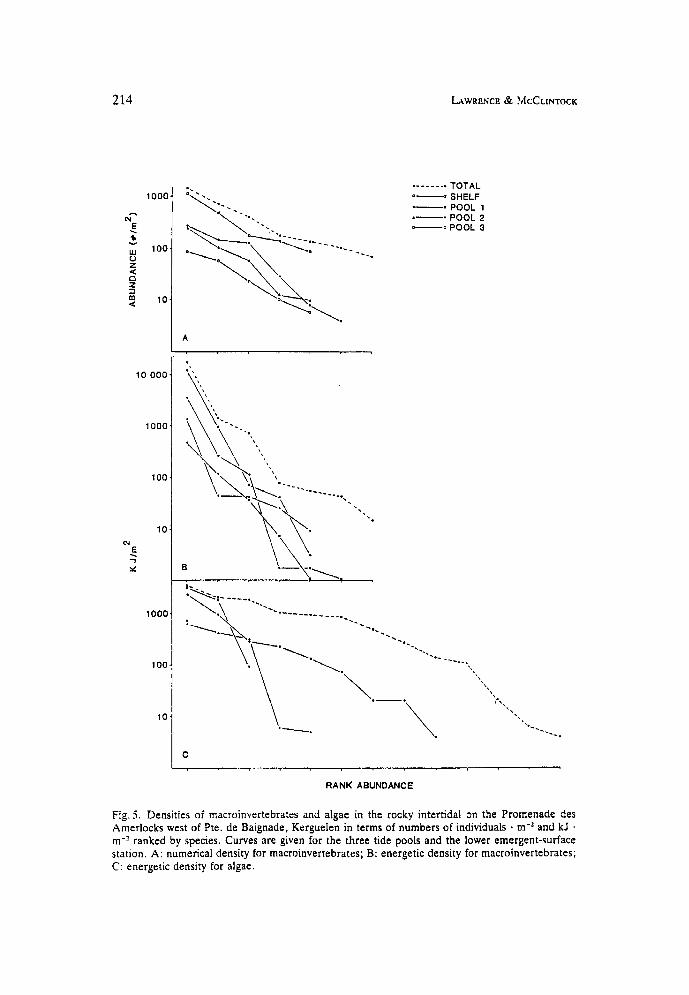
The numerical density/species-rank curves for the macroinvertebrates for the three tide-pools and the lower emergent-surface station show a relatively linear decrease (Fig. 5 ) . The energetic density/species-rank curves are steeper, reflect- ing the lower contribution of several species in energetic terms, and more irregular for the middle tide-pool (Pool 2).
The species list and energetic density of the algae along the transect are given in Table 3. No algae occurred on the higher emergent-surface station, although Porphyru sp. was noted on emergent surfaces in the vicinity. There was a distinct gradient in algal species richness in the tide pools, being greatest in the lowest one. The algal species richness of the lower emergent-surface station was less than that of the lowest tide-pool.
There was a gradient in the energetic density of the algae, increasing from the highest tide-pool to the lower emergent-surface shelf. The latter station had an energetic density four-fold that of the lowest tide-pool although it had less than one-half the number of algal species. Algal diversity was relatively unaffected in the middle and lowest tide-pools and the lower emergent-surface station despite the difference in number of species present (Table 2).
The energetic density/species-rank curves for the algae for all stations differ (Fig. 5). The highest tide-pool (Pool 3) contained only a single species (Fig. 5 c). The curve for the lowest tide pool (Pool 1) is linear and extends furthest.
214 LAWRENCE & MCCLINTOCK
.- - - - - -. TOTAL O- SHELF .-. POOL 1 .-. POOL 2 -0 POOL 3
I
I A
--._.
0,
I c RANN ABUNDANCE
Fig. 5 . Densities of macroinvertebrates and algae in the rocky intertidal on the Promenade des Amerlocks west of Pte. de Baignade, Kerguelen in terms of numbers of individuals . m-2 and kJ . m-? ranked by species. Curves are given for the three tide pools and the lower emergent-surface station. A: numerical density for macroinvertebrates; B: energetic density for macroinvertebrates; C: energetic density for algae.

Intertidal invertebrate and algal communities 215
Table 3. Energetic density (kJ * m-’) and species richness (S) of algae in the tide pools and the lower emergent-surface of the rocky intertidal at Kerguelen.
Species Upper Middle Lower Lower Tide Pool Tide Pool Tide Pool Emergent (3) (2) (1) Surface
Plumoriopsb eatoni 0 0 0 3034 Hymenema laciniata 0 0 0 1892 Ulva rigida 0 5 21 96 Codium difforme 0 0 2402 0 Macrocystis pyrifera 0 0 950 0 Durvillea antarctica 0 0 276 0 Ceramium rubrum 0 607 230 0 Myriogramme livida 0 0 138 0 Callophyllus tenera 0 423 71 0 Spongomorpha pacifica 0 0 21 0 Cladophora sp. 0 0 4 0 Caepidium antarcticum 0 6 0 0 ffildenbrandia lecanellieri 695 322 0 0 Sum 695 1363 41 13 5022 S 1 4 9 3
2. Description of the communities in December 1984
The species list and numerical density of macroinvertebrates that colonized the tide pools since denudation 23 months earlier (January 1983) are given in Table 4. The same species composition occurred in the highest tide-pool, but addi- tional species were present in very low densities in the lower two tide-pools. These included a gastropod, a crab, and an asteroid.
The total numerical densities, much less than the original values, were 48 %, 25 %, and 4 % in a gradient from the highest to the lowest tide-pool. This gradient in colonization occurred for both Mytilus edulis and Aulacomya ater. The diversity indexes of macroinvertebrates in the three tide-pools were high and similar to those found in 1983 (Table 2).
Discussion
Both physical and biological factors must be considered when analyzing the communities of the rocky intertidal at Kerguelen. The physical characteristics are primarily oceanic and meteorological and involve the relatively constant high winds that can become extreme, as well as relatively constant light rains and variable seasonal temperatures. The emergent surfaces can be subjected to extremely high levels of water activity, desiccation accentuated by the high winds, low salinity, frost during the winter, or high temperatures during the summer (ARNAUD, 1974). The biological characteristics are evident in the

216 LAWRENCE & MCCLINTOCK
Table 4. Numerical (ind . m-’) densities and species richness (S) of invertebrates in the tide pools in the rocky intertidal at Kerguelen in December 1984 (23 months after all algae and rnacroinverte- brates had been removed from the tide pools).
Species Upper Tide Pool ( 3 ) Middle Tide Pool (2) Lower Tide Pool (I)
Aulacomya ater iMytilur edulis desolutionir Anemone sp. Herniarthrum sendosum Kerguelenella lateralis Trachythyone cf. kerguelensb Halicurcinus planatus Asterina frigida Anuterias perrieri Trophon albolubratus Patinigera fuegiensis edgari Sum s
22 45 15 2 3 0 0 0 0 0 0
87 5
14 10
20 60
1.8
0.2 0.8 0.7 0 1.8 0
109 9
1.5 0.5 4.7 1.5 6.3 0.5 1.1 1.1 0.5 0 0.5
18 10
reduced number of intertidal species and of entire functional groups, probably the result of zoogeographical isolation and island effects.
The fauna and flora in the rocky intertidal at Kerguelen is depauperate compared to that of temperate and tropical regions (DAYTON, 1971; FEMINO & MATHIESON, 1980; LUBCHENCO, 1978; LUBCHENCO el al., 1984; MCQUAID & BRANCH, 1984; PAINE & VADAS, 1969; UNDERWOOD, 1981). Intertidal com- munities in the sub-antarctic which are depauperate in macroinvertebrate and algal species have been reported for nearby Heard Island (SMITH &. SIMPSON, 1985) and for Macquarie Island (KENNY & HAYSOM, 1962) and Marion Island (DE VILLIERS, 1976; BLANKLEY & GRINDLEY, 1985).
Although the small asteroid Anmterias rupicolu penetrates the lower interti- dal in very protected regions of the Bay of Morbihan (LAWRENCE et (ul., 1983), neither carnivorous asteroids nor gastropods are present in the exposed interti- dal which could function as keystone species. In this aspect, the rocky intertidal at Kerguelen is similar to the northeastern United States where predators are least abundant on exposed shores with extreme water energy (MENGE, 1978, 1983). Macroinvertebrate carnivores do not seem important in determining the faunal and floral composition of the tide pools or emergent surfaces of the rocky intertidal at Kerguelen.
The gull Larus dominicanus preys upon the mussels in the upper intertidal and may restrict the mussels to protected locations there (ARNAUD, 1974). The gull also preys upon the large limpet Patinigera kerguefensis which occurs in the lower intertidal among the holdfasts of Durvillea anrurccica and subtidally, but does not seem to consume the small limpet Kerguelenella lateralis in large numbers (ARNAUD, 1974). We have no information on possible predation by fish during high tide.
Herbivores frequently determine intertidal community structure (JARA & MORENO, 1984; LUBCHENCO, 1978; MENGE & LUBCHENCO, 1981; PAINE & VADAS,

Intertidal invertebrate and algal communities 217
1969; SOUSA, 1980; SOUSA et af . , 1981; SZE, 1980; UNDERWOOD, 1981; UNDER- WOOD & JARNAKOFF, 1984). Only two small grazing herbivores occur in the exposed intertidal of the Bay of Morbihan. The chiton Hemiarfhrum setulosum and the limpet Kerguelenella lateralis graze on green-algal film in the intertidal at Macquarie Island, although a variety of macroscopic algae were eaten in the laboratory (SIMPSON, 1976). Herbivores do not seem important in determining the faunal and floral composition of the tide pools and emergent surfaces of the rocky intertidal at Kerguelen. However, the intertidal limpet Nacelfu defesserti feeds heavily on algal sporelings at Marion Island (BLANKLEY & GRINDLEY, 1985) and could influence algal community composition in this way.
The relative importance of the various species differs when their densities are expressed in numerical and energetic terms. MCQUAID & BRANCH (1984) pointed out that although numerical densities are important, patterns of energy flow through a community are of considerable significance in understanding trophic interactions.
The less rigorous conditions and the longer periods of water exchange important for suspension-feeding result in an increased biomass in the lower intertidal. The three suspension-feeders made up 99.9 % of the total macroin- vertebrate gravimetric biomass at the, lower emergent-surface station, with Aufacomya afer contributing 92 %. Subtidal energetic densities of A . afer off the coast of South Africa range from 1620kJ . m-* to 18400 kJ . m-? (FIELD e f af . , 1980). Energetic densities of two bivalve species on the Natal coast, Perna pernu and Crassosrrea rnargurifacea, were 6702.5 kJ - m-* and 3440.1 kJ . m-?, respect- ively (BERRY, 1982). Thus the biomass supported by conditions in the Bay of Morbihan are similar to those at the two above, greatly different environments.
Numerical and energetic densities of intertidal macroinvertebrates and algae at Kerguelen can be compared with those at Marion Island (BLANKLEY & GRINDLEY, 1985). A t Marion, the mean density of the chiton Herniarthrum serulosum is similar to densities at Kerguelen (188 ind - m-? and 6.6 kJ * m-’ vs 259 ind . m-2 and 9.2 kJ - m-?); that of the limpet Kerguefenelfa futeralis is similar in terms of individuals (57 ind m-? vs 58 ind 3 m-.) but lower in terms of energy (4.5 kJ - m-2 vs 43.9 kJ m-I); that of the holothuroid Psecidocnus fuevigatus is lower in both (8 ind * m-z and 11.3 kJ . m-? vs 117 ind - m-? and 72.0kJ * m-*). The intertidal studied by BLANKLEY & GRINDLEY (1985) differed from that studied here, because the asteroid Anasterias rupicofa was a conspicuous component of the community while no bivalves occurred. The Marion Island site would be more comparable to a community at Kerguelen found in a lower water-energy intertidal area with numerous small boulders.
Algal biomass was only 37 % of the macroinvertebrate biomass at the lower emergent-surface station. In contrast, the algal biomass (Hifdenbrandia fecanef- lieri) was 106 % of the macroinvertebrate biomass in the highest tide-pool (Pool l ) , 99 % in the intermediate pool (Pool 2), and 112 % in the lowest tide-pool (Pool 1). The biomass of the subtidal algae off the coast of South Africa ranged from 2370 kJ * m-’ to 8548 kJ * m-2 (FIELD et al., 1980), similar to the energetic densities of the algae in the lowest tide-pool (Pool 1) and the shelf of the intertidal in the Bay of Morbihan.
Low algal diversity occurred from the tide pools in the upper intertidal to the lowest station on the shelf which was exposed only at low tide. Similar

218 LAWRENCE & kV[CCL"TOCK
diversities occurred despite considerable changes in tidal level, species composi- tion, and abundances. As found by LUBCHENCO (1978), the low algal diversity can be correlated with the absence of herbivores.
The diversity indexes of macroinvertebrates based on numerical densities are low at all stations. They are much less than those found by LUBCHENCO er al. (1984) for the rocky intertidal of the Pacific coast of Panama except for the highest intertidal level. Both high-intertidal situations are stressful and charac- terized by a reduced number of species and densities. The diversity indexes at Kerguelen are much lower when calculated in terms of energetic density than when calculated in terms of numerical density.
The densityhpecies-rank curves all show a variation of densities among the macroinvertebrates and algal species. As with the diversity indexes., shapes of curves for macroinvertebrates depend on whether the units are densities in terms of individuals or energy. The change in density is much greater when expressed energetically than in terms of individuals, probably reflecting an area/ volume relation. Densityhpecies-rank curves of complex communities should show the lognormal distribution according to WARWICK (1984). Inclusion of the densities of isopods, arnphipods, and polychaetes would change the shape of the curves.
The low densities of macroinvertebrates 23 months after the tide pools had been denuded indicate that colonization and return to the original conditions is a slow process. There should be no lack of propagules: individuals of all species were abundant both in adjacent intertidal areas and subtidally. The slowness of reestablishment of the colonies may reflect the physical stress of the intertidal, even in the tide pools. However, the greater colonization of the upper tide-pools than the lower does not fit the expected zonation of stress in the tide pools.
Summary
The characteristics of the intertidal communities at Kerguelen result from the isolated location of the island and the rigorous oceanic conditions, as well as from the interaction of the species. Few species are present and some functional groups commonly associated with rocky intertidal areas are not present. A distinct zonation of both invertebrate and algal species occurs. Among algae, the number of species as well as biomass was greatest in the lowest tid.e pool. In contrast, the number of invertebrate species varied little with the level of the tide pools although the biomass again was greatest in the lowest tide pool.
Acknowledgements
This work was supported by NSF grant DPP-8108992 to the senior author and by the administration of the Terres Australes et Antarctiques Franqaises. We thank C. J . DAWES for identifying the algae; A . GUILLE for identifying the invertebrates; F. GUILBERT for making supplementary observations during the austral winter; G. BRANCH, A . GUILLE, R. PAINE, R. SIMPSON. and G. VANELARKOM for helpful comments on a draft of the paper; and the staff and personnel at Port-aux-Franqtis for their courtesy and assistance.

Intertidal invertebrate and algal communities 219
References
ARNAUD, P. M., 1974: Contribution a la bionomie marine benthique des regions antarctiques et subantarctiques. Tethys, 6: 480430.
BELLIDO, A,, 1981: Les biocenoses du littoral rocheux aux isles Kerguelen. Com. Nat. Franc. Recherch. Antarct., 51: 81-92.
BERRY, P. F., 1982: Biomass and density of detritivores on a littoral rocky reef on the Natal coast, with an estimate of population production for the ascidian Pyura stolonifera. Invest. Rep. Oceanogr. Res. Inst., 53: 1-15.
BLANKLEY, W. 0. & J. R. GRINDLEY, 1985: The intertidal and shallow sublittoral food web at Marion Island. In: W. R. SIEGRIED, P. R. CONDY & R. M. LAWS (Eds.), Antarctic Nutrient Cycles and Food Webs. Springer-Verlag, Berlin: 630-636.
BRODY, S., 1945: Bioenergetics and growth. Hafner Publishing Co., Inc., New York; 1023pp. CONNELL, J. H., 1972: Community interactions on marine rocky intertidal shores. Annu. Rev. Ecol.
Syst., 3: 169-192. DAYTON, P. K., 1971: Competition, disturbances and community organization: The provision and
subsequent utilization.of space in a rocky intertidal community. Ecol. Monogr., 41: 3.51-389. - -, 1975: Experimental evaluation of ecological dominance in a rocky intertidal algal community.
Ecol. Monogr., 45: 137-159. DELBPINE, R., 1963: Un aspect des etudes de biologie marine dans les Iles Australes FranFaises.
Com. Nat. Franc. Recherch. Antarct., 3: 1-22. DE VILLIERS, A. F.. 1976: Littoral ecology of Marion and Prince Edwards Islands (Southern Ocean).
S . Afr. J. Antarct. Res. (Suppl.), 1: 1-10. DOTY, M.S., 1957: Rocky intertidal surfaces. In: J. W. HEDGPETH (Ed.). Treatise on Marine
Ecology and Paleoecology, Vol. 1. The Geological Society of America, New York: 535-385. FEMINO, R. J. & ‘4. C . MATHIESON, 1980: Investigations of New England marine algae. IV. The
ecology and seasonal succession of tide pool algae at Bald Head Cliff, York, Maine, USA. Bot. Mar., 23: 319-332.
FIELD, J. G., C. L. GRIFFITHS, R. J. GRIFFITHS, N. JARMAN, P. ZOUTENDYK, B. VELIMIROV& A. BOWES, 1980: Variation in structure and biomass of kelp communities along the southwest Cape coast. Trans. R. SOC. S . Afr., 44: 145-203.
HUREAU, J.-C., 1970: Biologie comparCe de quelques poissons antarctiques (Nofotheniidae). Bull. Inst. Ockanogr., Monaco, 68: 1-244.
JARA, H. F. & C. A. MORENO, 1984: Herbivory and structure in a midlittoral rocky community: a case in southern Chile. Ecology, 65: 28-38,
KENNY, R. & N. HAYSOM, 1962: Ecology of rocky shore organisms at Macquarie Island. Pac. Sci.,
LAWRENCE, J. M., 1973: Level, content, and caloric equivalents of the lipid, carbohydrate, and protein in the body components of Luidia clafhrata (Echinoderrnaia: Asteroidea: Platyasterida) in Tampa Bay. J. Exp. Mar. Biol. Ecol., 11: 263-274.
- -, J. B. MCCLINTOCK & A. GUtLLE, 1983: Anasferias rupicola (VERRILL) dans la zone intertidale de la baie du Morbihan (archipel de Kerguelen). Symbioses, 15: 213.
LEWIS, J. R., 1964: The Ecology of Rocky Shores. The English Universities Press Ltd., London; 323 pp.
LUBCHENCO, J., 1978: Plant species diversity in a marine intertidal community: Importance of herbivore food preference and algal competitive abilities. Am. Nat., 112: 23-39.
- - & B. A. MENGE, 1978: Community development and persistence in a low rocky intertidal zone. Ecol. Monogr., 5 9 67-94.
- -, B. A. MENCE, S. D. GARRITY, P. J. LUBCHENCO, L. R. ASKENAS. S. D. GAINES, R. EMLET, J. LUCAS & S . STRAUSS, 1984: Structure, persistence, and role of consumers in a tropical rocky intertidal community (Taboguilla Island, Bay of Panama). J. Exp. Mar. Biol. Ecol., 78: 23-73.
MAY, V., I.BENNETT & T.E. THOMPSON, 1970: Herbivore-algal relationships on a coastal rock platform (Cape Banks, N. S . W.). Oecologia, 6: 1-14.
MCQUAID, C. D . & G. M. BRANCH, 1984: Influence of sea temperature, substratum and wave exposure on rocky intertidal communities: an analysis of faunal and floral biomass. Mar. Ecol. Prog. Ser., 19: 145-151.
MENCE, B.A., 1976: Organization of the New England rocky intertidal community: role of predation, competition, and environmental heterogeneity. Ecol. Monogr., 46: 355-393.
16: 245-263.

220 LAWRENCE & MCCLINTOCK
- -, 1978: Predation intensity in a rocky intertidal community. Oecologia, 34: 1-16. - -, 1983: Components of predation intensity in the low zone of the New England rocky intertidal
region. Oecologia, 58: 141-155. - - & J. LUBCHENCO, 1981: Community organization in temperate and tropical rocky intertidal
habitats: Prey refuges in relation to consumer pressure gradients. Ecol. Monogr., 51: 42-50, MORENO, C. A. & E. JARMILLO, 1983: The role of grazers in the zonation of intertidal macroalgae of
the Chilean coast near Valdivia. Oikos, 41: 73-76. NEWELL, R. C., 1979: Biology of Intertidal Animals. Marine Ecological Surveys Ltd., Faversham;
781 pp. PAINE, R.T., 1966: Food web complexity and species diversity. Am. Nat., 100: 65-75. - - , 1971: A short-term experimental investigation of resource partitioning in a New Zealand rocky
intertidal habitat. Ecology, 52: 1096-1106. - -, 1974: Intertidal community structure: Experimental studies on the relationship between a
dominant competitor and its principal predator. Oecologia, 15: 93-120. - - & R. L. VADAS, 1969: The effects of grazing by sea urchins, Sfrongylocenfrofus spp., on benthic
algal populations. Limnol. Oceanogr., 14: 710-719. PYEFINCH, K. A., 1943: The intertidal ecology of Bardsey Island, North Wales, with special reference
to the recolonisation of rock surfaces and the rockpool environment. J. Aninr. Ecol., 12:
SIMPSON, R. D., 1976: Physical and biotic factors limiting the distribution and abundance of littoral molluscs on Macquarie Island (sub-antarctic). J. Exp. Mar. Biol. Ecol., 21: 11-49.
SMITH, J. M. B . & R. D. SIMPSON, 1985: Biotic zonation on rocky shores of Heard Island. Polar Biol., 4: 89-93.
SOUSA, W . P., 1980: The responses of a community to disturbance: The importance of successional age and species' life histories. Oecologia, 45: 72-81.
- -, S. C.' SCHROETER & S. D. GAINES, 1981: Latitudinal variation in intertidal algal community structure: the influence of grazing and vegetative propagation. Oecologia. 48: 297-307.
STEPHENSON, T. A. & A. STEPHENSON, 1972: Life Between Tidemarks on Rocky Shores. W. H. Freeman and Company, San Francisco; 425 pp.
SZE, P., 1980: Aspects of the ecology of macrophytic algae in high rock pools at the of Shoals (USA). Bot. Mar., 23: 313-318.
UNDERWOOD, A. J. , 1981: Structure of a rocky intertidal-community in New South Wales: Patterns of vertical distribution and seasonal changes. J. Exp. Mar. Biol. Ecol., 51: 57-85.
- - & P. JERNAKOFF, 1984: The effects of tidal height, wave-exposure, seasonality and rock pools on grazing and the distribution of intertidal macroalgae in New South Wales. J. Exp. Mar. Biol.
WARWICK, R. M., 1984: Species size distributions in marine benthic communities. Oecologia, 61:
82-108.
Eco~. , 75: 71-96.
32-41.