random distribution of mammalian replication origins in matrix and total nuclear dna
TRANSCRIPT

36 Biochimica et Biophysica Acta, 783 (1984) 36-41 Elsevie~
BBA91387
R A N D O M D I S T R I B U T I O N OF M A M M A L I A N REPLICATION O R I G I N S IN MATRIX A N D TOTAL NUCLEAR D N A
MARIA TODOROVA and GEORGE RUSSEV *
Institute of Molecular Biology, Bulgarian Academy of Sciences, 1113 Sofia (Bulgaria)
(Received April 10th, 1984)
Key words: DNA repfication," Repfication origin, Nuclear matrix, (Rat, Mouse)
Nuclear matrices from mouse and rat tumour cells were isolated and characterized by their microscopic appearance, protein profiles and D N A content. They presented well-defined structures containing 15-20% of the nuclear protein and 1-3% of the nuclear DNA. Matrix DNAs were immobilized on nitrocellulose filters and hybridized to nick-translation 32p-labelled homologous D N A fragments containing the corresponding replication origins. As control total nuclear DNAs were also immobilized on filters and hybridized to origin-containing DNAs. The origin-containing DNAs hybridized to the same extent to both matrix and total DNAs, which showed that they contained the same proportion of origin sequences. In an alternative series of experiments, plasmids containing either rat or mouse replication origins were immobilized on filters and were hybridized with in vitro 32p-labelled matrix and total nuclear DNAs. Here again both matrix and total nuclear DNAs hybridized to the same extent with the origin-carrying plasmids, which showed that neither rat nor mouse matrix DNAs were enriched in D N A replication origin sequences.
Introduction
Several years ago Berezney and Coffey [1,2] described a specific and ubiquitous nuclear struc- ture that they called the nuclear matrix. Since then the nuclear matrix has been an object of numerous investigations and gradually a view has been formed that it contains a few characteristic pro- teins with molecular weights in the range between 60000 and 70000 [3-5] and that the nuclear D N A is organized in the form of huge loops anchoring to this structure [6-8]. It has further been shown that matrix D N A is enriched in newly replicated D N A [2,6,9,10], in actively transcribed genes [11-13] and in D N A replication origins [14], and
* To whom correspondence should be addressed. Abbreviations: Cot, the product of nucleotide concentration (mol/ l ) and time (s); PMSF, phenylmethylsulfonyl fluoride.
that the attachment sites of D N A to the nuclear matrix represent a specific class of repeated se- quences [15]. All these data show that the nuclear matrix is a very complex structure that plays an important role in both D N A replication and tran- scription. However, at the same time other investi- gators presented data that cast some doubt on this picture. Thus it has been demonstrated that the major nuclear matrix proteins are in fact the pro- teins of the long-known nuclear lamina [8,16-18], that nuclear matrix D N A has no specific char- acteristics [19], that the active genes are not prefer- entially bound to the matrix [20,21], and finally that the nuclear matrix itself is an artefact [22,23]. This second group of results deprives the matrix of any functional role other than to provide the structural framework of the cell nucleus.
Recently we developed a method for isolation of mammalian D N A fragments containing replica- tion origins [24,25]. In the present communication
0167-4781/84/$03.00 © 1984 Elsevier Science Publishers B.V.

we determined the abundance of the origin se- quences in mouse and rat matrix DNA. It was approximately the same as in the corresponding total nuclear DNAs, and this was interpreated to mean that nuclear matrix D N A was not specifi- cally enriched in D N A replication origins.
Materials and Methods
Isolation of nuclei and nuclear matrix Ehrlich ascites tumour and Guerin ascites
tumour cells were propagated in vivo in commer- cially obtained albino mice and rats, respectively. At days 7-10 after inoculation ascites liquid was collected, cells were spun down and washed with 0.25 M sucrose/10 mM Tris-HC1 (pH 7.4)/3 mM MgC12- Unless otherwise stated all procedures were carried out at 4 ° C and in the presence of 1 mM PMSF (Serva). Cells were suspended in the same buffer and lysed with 0.5% (v /v ) Nonidet P-40 for 15 rain, the nuclei were sedimented by low-speed centrifugation and purified by ultracentrifugation through 1.6 M sucrose. Nuclei were suspended in 0.25 M sucrose/10 mM Tris-HC1 (pH 7.4)/3 mM MgCI 2 to approx. 100 #g D N A / m l and digested with RNAase A and DNAase I [17]. Ehrlich ascites tumour nuclei were first digested with 250 # g / m l of RNAase A (Sigma) at 37 °C for 30 min and then with 150 un i t s /ml of DNAase I (Sigma) added in the same reaction mixture at 37 o C for 5 min. Guerin ascites tumour nuclei were more dif- ficult to digest and they were treated with 250 # g / m l of RNAase A and 150 uni t s /ml of DNAase I simultaneously at 37 °C for 1 h. After comple- tion of the digestion the nuclear suspension was mixed with an equal volume of ice-cold 4 M NaCI and the nuclear matrices were pelleted down at 600 × g for 40 rain. They were extracted twice with 2 M NaC1/10 mM Tris-HC1 (pH 7.4)/0.2 mM MgCl 2 and finally washed in 10 mM Tris-HCl (pH 7.4)/0.2 mM MgCI 2.
Isolation of DATA, labelling and hybridization Total nuclear of matrix DNAs were isolated by
phenol /ch loroform (1:1) and chloroform extrac- tions after digestion of nuclei or matrices with Proteinase K (Merck) in the presence of 1 M NaC1/ I% SDS/50 mM Tris-HC1 (pH 7.4)/1 mM EDTA.
37
In order to determine DNA, cells were labelled for 2 h with [3H]thymidine (25 Ci /mmol , (~SSR) as previously described [24], and the radioactivity of D N A was determined. Using this figure the amount of residual matrix D N A was calculated from the radioactivity of the corresponding matrix preparations.
The D N A fractions containing rat and mouse replication origins were isolated from cells cross- linked with trioxsalen exactly as described in Refs. 24 and 25. D N A was labelled with [32p]dCTP (200-400 C i /mmol , Amersham) to approx. 5 • 107 c p m / p g by nick-translation [26]. Immobilization on nitrocellulose filters was as in Ref. 27 and hybridization as in Ref. 28.
The general approach for constructing and screening mouse and rat replication origin-con- taining plasmids is briefly outlined in the text to Fig. 4. The detailed procedure will be presented elsewhere (Vassilev and Russev, unpublished data; Anachkova, Nosikov and Russev, unpublished data).
Eiectrophoresis D N A was electrophoresed in 1.5% horizontal
agarose (Sigma) gels prepared and run in 40 mM Tris-acetate buffer (pH 7.8)/2 mM EDTA. Gels were stained with ethidium bromide and photo- graphed under ultraviolet light.
Proteins of nuclei and nuclear matrices were solubilized in 1% S D S / I % 2-mercaptoethanol/65 mM Tris-HC1 (pH 6.8) and electrophoresed in 10% polyacrylamide vertical slab gels as described in Ref. 29. Gels were stained with Coomassie brilliant blue (Serva).
Results
In a previous paper [30] we have demonstrated that some mouse chromosomal nonhistone pro- teins had preferential affinity towards D N A frag- ments containing the putative replication origins. These proteins had molecular weights in the range 60000-70000, which suggested that they might belong to the nuclear matrix. To check this possi- bility we decided to isolate nuclear matrices from mouse and rat tumour cells and to see whether they were enriched in the corresponding origin D N A sequences. For isolation of the nuclear
38
matrices we applied the procedure of K a u f m a n n et al. [17], since it efficiently el iminated the h n R N P
componen t of the matrices. According to their nomencla tu re these matrices were designated
nuclear ghosts. Care was taken to avoid any rup- t ion a n d / o r aggregation of both nuclei and matrices, and phase-contrast light microscopy
showed that they presented well-defined intact structures (Fig. 1). The nuclear matrices conta ined about 15-20% of the total nuclear protein and 1-3% of the nuclear DNA, depending on the degree of DNAase I digestion. When nuclei and
matrices had not aggregated dur ing the isolation procedure no long D N A fragments were observed and all matrix D N A represented low molecular
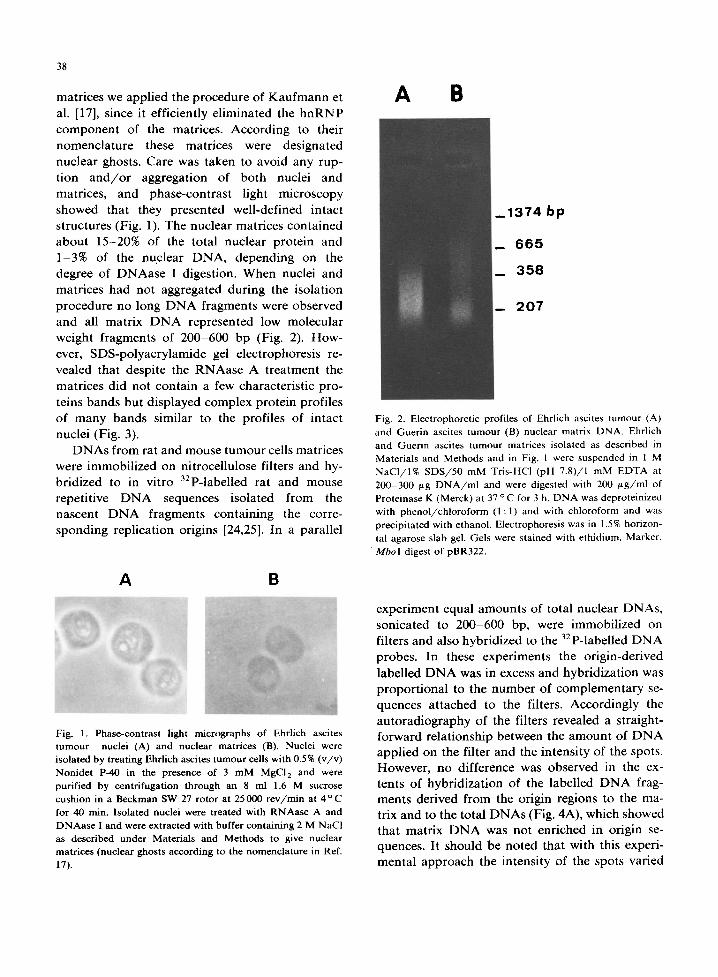
weight fragments of 200-600 bp (Fig. 2). How- ever, SDS-polyacrylamide gel electrophoresis re- vealed that despite the RNAase A t reatment the
matrices did not conta in a few characteristic pro- teins bands bu t displayed complex protein profiles of m a n y bands similar to the profiles of intact
nuclei (Fig. 3). D N A s from rat and mouse tumour cells matrices
were immobil ized on nitrocellulose filters and hy- bridized to in vitro 32p-labelled rat and mouse
repetitive D N A sequences isolated from the nascent D N A fragments conta in ing the corre- sponding replicat ion origins [24,25]. In a parallel
A B
_ 1 3 7 4 bp
_ 6 6 5
_ 3 5 8
_ 2 0 7
Fig. 2. Electrophoretic profiles of Ehrlich ascites tumour (A) and Guerin ascites tumour (B) nuclear matrix DNA. Ehrlich and Guerin ascites tumour matrices isolated as described in Materials and Methods and in Fig. 1 were suspended in 1 M NaCI/I% SDS/50 mM Tris-HC1 (pH 7.8)/1 mM EDTA at 200-300 ~tg DNA/ml and were digested with 200 gg/ml of Proteinase K (Merck) at 37 o C for 3 h. DNA was deproteinized with phenol/chloroform (1:1) and with chloroform and was precipitated with ethanol. Electrophoresis was in 1.5% horizon- tal agarose slab gel. Gels were stained with ethidium. Marker, M b o I digest of pBR322.
A B
i!!l ........................ 'i
Fig. 1. Phase-contrast light micrographs of Ehrlich ascites tumour nuclei (A) and nuclear matrices (B). Nuclei were isolated by treating Ehrlich ascites tumour cells with 0.5% (v/v) Nonidet P-40 in the presence of 3 mM MgC12 and were purified by centrifugation through an 8 ml 1.6 M sucrose cushion in a Beckman SW 27 rotor at 25 000 rev/min at 4 o C for 40 min. Isolated nuclei were treated with RNAase A and DNAase I and were extracted with buffer containing 2 M NaCI as described under Materials and Methods to give nuclear matrices (nuclear ghosts according to the nomenclature in Ref. 17).
experiment equal amount s of total nuclear DNAs, sonicated to 200-600 bp, were immobil ized on filters and also hybridized to the 32p-labelled D N A
probes. In these experiments the origin-derived
labelled D N A was in excess and hybr idizat ion was propor t ional to the n u m b e r of complementary se- quences at tached to the filters. Accordingly the autoradiography of the filters revealed a straight- forward relat ionship between the amoun t of D N A applied on the filter and the intensi ty of the spots. However, no difference was observed in the ex- tents of hybr idizat ion of the labelled D N A frag- ments derived from the origin regions to the ma- trix and to the total D N A s (Fig. 4A), which showed that matr ix D N A was not enriched in origin se- quences. It should be noted that with this experi- menta l approach the intensi ty of the spots varied

A B C D
~.. 6 8 kD
39
f r o m e x p e r i m e n t to e x p e r i m e n t d u e to s o m e un -
c o n t r o l l e d d e t a c h m e n t o f t he s h o r t D N A frag-
m e n t s i m m o b i l i z e d o n the f i l ters d u r i n g the hy -
b r i d i z a t i o n a n d s u b s e q u e n t washes . M e a n w h i l e we
c l o n e d in p B R 3 2 2 ra t a n d m o u s e D N A f r a g m e n t s
c o n t a i n i n g the c o r r e s p o n d i n g r e p l i c a t i o n o r ig ins
a n d , to avo id the loss o f D N A in a s e c o n d ser ies o f
e x p e r i m e n t s , i m m o b i l i z e d o n f i l ters severa l such
p l a s m i d s a n d h y b r i d i z e d to t h e m m a t r i x D N A s
32p- label led by n i c k - t r a n s l a t i o n . A s con t ro l s ,
pa ra l l e l f i l ters c o n t a i n i n g the s a m e a m o u n t o f
p l a s m i d s w e r e h y b r i d i z e d wi th s o n i c a t e d in v i t ro
Fig. 3. Electrophoretic profiles of Ehrlich ascites tumour (A, B) and Guerin ascites tumour (C, D) total nuclear (A, C) and matrix (B, D) proteins. Ehrlich and Guerin ascites tumour matrices were isolated as described in the text and solubilized in sample buffer containing 1% SDS/1% 2-mercaptoethanol by heating in a boiling water bath for 5 rain. Electrophoresis was in 10% vertical polyacrylamide slab gels. Gels were stained with Coomassie. Arrow points the position of bovine serum albumin (68 kDa) eleetrophoresed in parallel.
A B C
1 1 1 1 ̧̧ i O
0 ¸
2 2 2 i
i i! i i ! i i~
Fig. 4. Hybridization of isolated DNA fragments containing mouse and rat replication origin sequences to the corresponding total nuclear and matrix DNAs. A, Nascent DNA chains containing Ehrlich ascites tumour replication origins were isolated from trioxsalen-crosslinked cells and the repetitive DNA sequences present in these fragments were isolated after reassociation to Cot 5 and S 1 nuclease digestion [24,25]. They were labelled with [32p]dCTP by nick translation and hybridized to nitrocellulose filters on which 100 ng or 10 ng of either total nuclear Ehrhch ascites tumour DNA sonicated to 200-6000 bp (1) or matrix Ehrlich ascites turnout DNA (2) has been applied [27]. B. Rat DNA fragments were cloned in pBR322 and screened for colonies containing rat replication origins using the nascent DNA fragments isolated from trioxsalen-crosslinked Guerin ascites tumour cells [25] as probes. Two plasmids, one containing the replication origin sequences and the other containing unique DNA (arrows), were applied on
32 nitrocellulose filters in two concentrations (1 pg and 100 ng per dot) and were hybridized with in vitro P-labelled sonicated rat nuclear DNA (1) or rat matrix DNA (2). C. Ehrlich ascites tumour DNA fragments were cloned in pBR322 and screened for colonies containing Ehrlich ascites tumour replication origin sequences. Three such plasmids were isolated and applied on nitrocellulose filters (100 ng per dot). The filters were hybridized to either total sonicated in vitro 32p-labeiled mouse total nuclear (1) or matrix DNA (2). Filters were exposed to X-ray (ORWO) films in the presence of intensifying screens at - 70 o C for 24 h.

40
32p-labelled total D N A s (Fig. 4B,C). In this case the amount of immobilized D N A was in huge excess and any difference in the rates of hybridiza- tion of matrix and total D N A s would reflect dif- ferences in the concentrat ions of complementary sequences [27]. However, here again the origin- containing plasmids hybridized to the same extent with both total nuclear and matrix DNAs, which was interpreted to mean that matrix D N A was not enriched in D N A replication origin sequences.
Later the same experiments were repeated with matrix D N A s f rom rat and mouse liver instead of tumour matrices and exactly the same result was obtained (not shown).
Discussion
Current ideas are that in interphase nuclei D N A is organized in the form of replicon-size loops anchoring to a specific protein structure called the nuclear matrix or skeleton to which the newly synthesized D N A is being at tached (see Refs. 31 and 32). This was interpreted to mean that D N A replication takes place at fixed sites on the nuclear matrix, and stimulated speculations concerning mechanisms for eukaryotic D N A replication. All implied the seemingly obvious assumption that D N A replication origins both before the onset of D N A replication and after its completion, i.e., during the better part of the cell cycle, should remain permanent ly bound to the nuclear matrix. Recently this assumption was experimentally con- firmed by Aelen et al. [14], who deduced from their labelling kinetics experiments with Physarum that origins of replication or D N A sites very close to them were at tached to the nuclear matrix during the entire cell cycle.
In a previous paper [24] we have reported a new method for isolation of D N A fragments contain- ing mammal ian replication origins. It was based on the assumption that, in cells whose D N A had been in vivo crosslinked with trioxsalen, elonga- tion of the already initiated D N A chains should be effectively blocked while the initiation of D N A synthesis would not be seriously affected. This would lead to the synthesis and accumulat ion of short nascent D N A fragments containing replica- t ion origins, which could be released and sep- arated f rom the bulk of D N A in alkaline sucrose
density gradients. In a more recent paper [25] we applied this method to a variety of cell lines and were able to demonst ra te that in all cases short nascent chains had been synthesized as a result of semiconservative D N A replication. Still more re- cently, yeast cells were used to further evaluate and quanti tate the trioxsalen method. In this case the nascent chains synthesized between crosslinks were specifically enriched in ARS1 sequences [33,34], which directly proved the validity of the method for isolation of eukaryotic replication origins (unpublished results). In the present com- municat ion we used mouse and rat nascent frag- ments per se or cloned in plasmids as hybridiza- tion probes to study the distribution of the replica- tion origins and the adjacent D N A fragments be- tween total and matrix DNA. Our experiments failed to reveal any significant enrichment of either mouse or rat matrix D N A s in such sequences and we therefore concluded that the regions of initia- tion of mammal ian D N A replication were not permanent ly bound to the nuclear matrix. This finding is in agreement with the recent results of Cook and Lang [35], who also failed to detect any specific association of the initiation sites of D N A replication to the nuclear matrix. However, it should be pointed out that our results do not exclude the possibility of some dynamics in the D N A organization so that the replication origins due to begin replication shortly become associated to the nuclear matrix, while the vast majori ty of quiescent replication origins are scattered ran- domly along the D N A regardless of its a t tachment to the nuclear matrix.
References
1 Berezney, R. and Coffey, D.S. (1974) Biochem. Biophys. Res. Commun, 60, 1410-1417
2 Berezney, R. and Coffey, D.S. (1975) Science 198, 291-293 3 Berezney, R. and Coffey, D.S. (1977) J. Cell Biol. 73,
616-637 4 Berezney, R. (1980) J. Cell Biol. 85, 641-650 5 Berezney, R. and Buchholtz, L. (1981) Biochemistry 20,
4995-5002 6 Pardoll, D., Vogelstein, B. and Coffey, D.S. (1980) Cell 19,
527-536 7 Lebkowski, J. and Laemmli, U.K. (1982) J. Mol. Biol. 156,
309-324 8 Hancock, R. and Hughes, M. (1982) Biol. Cell. 44, 201-212 9 Hunt, B. and Vogelstein, B. (1981) Nucleic Acids Res. 9,
349-363

10 Berezney, R. and Buchholtz, L. (1981) Exp. Cell Res. 132, 1-13
11 Robinson, S., Nelkin, B. and Vogelstein, B. (1982) Cell 28, 99-106
12 Robinson, S., Small, D., Idzera, R., McKnight, G. and Vogelstein, B. (1983) Nucleic Acids Res. 11, 5113-5130
13 Giejek, E.M., Tsai, M.J. and O'Malley, B.O. (1983) Nature 306, 607-609
14 Aden, J.M.A., Opstelten, R.J.G. and Wanka, F. (1983) Nucleic Acids Res. 11, 1181-1195
15 Razin, S.V., Mantieva, V.L. and Georgiev, G.P. (1979) Nucleic Acids Res. 7, 1713-1735
16 Peters, K. and Commings, D. (1980) J. Cell Biol. 86, 135-155
17 Kaufman, S., Coffey, D. and Shaper, J. (1981) Exp. Cell Res. 132, 105-132
18 Lebkowski, J. and Laemmli, U.K. (1982) J. Mol. Biol. 156, 325-344
19 Basler, J., Hastie, N., Pietras, D., Matsui, S.-I., Sandberg, A. and Berezney, R. (1981) Biochemistry 20, 6921-6929
20 Kuo, T.M. (1982) Biochemistry 21, 321-326 21 Ross, D., Yen, R.-W. and Chae, Chi-B. (1982) Biochemistry
21,764-771
41
22 Hadlaczki, G., Sumner, A. and Ross, A. (1981) Chro- mosoma 81, 557-567
23 Oalcheva-Gargova, Z., Petrov, P. and Dessev, G. (1982) Eur. J. Cell Biol. 28, 155-159
24 Russev, G. and Vassilev, L. (1982) J. Mol. Biol. 161, 77-87 25 Anachkova, B., Todorova, M., Vassilev, L. and Russev, G.
(1984) Eur. J. Biochem. 141,105-108 26 Rigby, P., Dieckmann, M., Rodes, C. and Berg, P. (1977) J.
Mol. Biol. 113, 237-251 27 Kafatos, F., Jones, C. and Efstratiadis, A. (1979) Nucleic
Acids Res. 7, 1541-1551 28 Grunstein, M. and Hogness, D. (1975) Proc. Natl. Acad.
Sci. USA 72, 3961-3965 29 Laemmli, U.K. (1970) Nature 227, 680-685 30 Anachkova, B. and Russev, G. (1983) Biochim. Biophys.
Acta 740, 369-372 31 Hancock, R. and Boulikas, T. (1982) Annu. Rev. Cytol. 79,
165-214 32 Hancock, R. (1982) Biol. Cell. 46, 105-122 33 Stinchcomb, D.T., Struhl, K. and Davis, R.M. (1979) Na-
ture 282, 39-43 34 Tschumper, G. and Carbon, J. (1980) Gene 10, 157-166 35 Cook, P,R. and Lang, J. (1984) Nucleic Acids Res. 12,
1069-1075