radular teeth of indo-pacific molluscivorous species of conus
TRANSCRIPT

J. Moll. Stud. (1999), 65, 483–497 © The Malacological Society of London 1999
ABSTRACT
Species determination in the gastropod genus Conus,heretofore exclusively based on shell morphology andcolor pattern, has led to considerable uncertainty anddisagreement. We propose that qualitative and quan-titative radular tooth characters are potentially usefulin differentiating species as well as geographic subspecies and will improve the taxonomic base. Mol-luscivorous species of Conus, sometimes placed in thesubgenera Cylinder, Textilia, Darioconus, and thenominal subgenus, are taxonomically among the mostdifficult. We thus examined intra- and interspecificvariation in radular morphology of 11 of thesespecies, C. ammiralis, C. araneosus, C. bandanus, C.canonicus, C. episcopatus, C. marmoreus, C. nodulo-sus, C. omaria, C. pennaceus, C. textile, and C. victo-riae, and intra- and interregional variations in radularmorphology of C. pennaceus from three geographicregions. Taxonomically useful qualitative charactersinclude presence/absence of one or two barbs and ablade, and whether the row of denticles comprisingthe serration is continuous or interrupted. Usefulmetric characters include the ratios of first barb, second barb, blade, serration, shaft width and basewidth to tooth length, the ratio of tooth length to shelllength, the ratio of shaft width to base width, and thedegree of curvature of the teeth. Univariate analysisof variance (ANOVA and unplanned pairwise comparison tests) distinguished 53 of the 55 possiblespecies pairs from each other by at least one charac-ter. Multivariate analysis of variance (MANOVA)indicated statistically significant differences betweenthe species in the other two pairs. In C. pennaceus,ANOVA and unplanned pairwise tests differentiatedHawaiian from Indian Ocean samples, andMANOVA differentiated those from Maldives andSumatra. The discrete radular characters sort the 11species into three groups, and these are consistentwith the distribution patterns of the quantitative char-acters. Radular tooth characters are thus potentiallyuseful in differentiating species and subspecies andshould be combined with other character sets in generating future phylogenetic hypotheses.
INTRODUCTION
The predatory marine gastropod genus Conusis unusually diverse in number of species(.500), shell size (,2 cm to .20 cm), andbehavioural and ecological relationships (Kohn,1990). More than 300 species occur in the tropi-cal Indo-Pacific region (Röckel, Korn & Kohn,1995). While a few Conus species are more generalized, most are specialist predators onpolychaete annelids, other gastropods, or fishes,injecting paralytic venoms called conotoxinswith a hollow, needle-like radular tooth to capture prey (Kohn, 1956; Olivera, Rivier, Clark,Ramilo, Corpuz, Abogadie, Mena, Woodward,Hillyard & Cruz, 1990). Each tooth is struc-turally more complex than the individual teethof other molluscan radulas, is not attached to aradular membrane, and is used only once, as acombination harpoon and hypodermic venomdelivery system (Röckel et al., 1995; Olivera etal., 1990; Kohn, 1998).
Prior studies of Conus radular teeth havedemonstrated a remarkable range of morpho-logical diversity of teeth among species, haveclassified morphologically similar teeth intogroups, have associated functional morphologyof teeth with dietary specialization, and have in-vestigated ontogenetic changes in tooth morph-ology. Kohn, Nishi and Pernet (1999) reviewthese studies and provide the first descriptivetaxonomic character analysis of Conus radularteeth.
The need for accurate taxonomic analysis ofConus is increasing rapidly because:
(1) Presently, species are classified by shellcharacters. However, since the ranges ofwithin- and among-species variations inshell characters are poorly known, this clas-sification is subjective and there has beencontinuous uncertainty and disagreementconcerning it. For example, two of thespecies we consider, C. victoriae and C.nodulosus, have been synonymized by some
RADULAR TEETH OF INDO-PACIFIC MOLLUSCIVOROUSSPECIES OF CONUS : A COMPARATIVE ANALYSIS
MANAMI NISHI1 and ALAN J. KOHN2
Department of Zoology, University of Washington, Box 351800, Seattle, Washington 98195, U.S.A.(Received 23 October 1998; accepted 25 January 1999)
1 Present address: Department of Pathobiology, University of Pennsyl-vania, Philadelphia, Pennsylvania 19104-6008, U.S.A. e-mail:[email protected] 2 Author for correspondence. e-mail:[email protected]

484 MANAMI NISHI & ALAN J. KOHN
workers (e.g., Röckel et al., 1995) andassigned to different subgenera by others(e.g., Marsh & Rippingale, 1974).
(2) The evolutionary history of Conus is poorlyknown and taxonomic analysis of Conuscould provide potential phylogenetic infor-mation.
(3) Conus venoms are useful in neurobiologicaland medical research because of their selec-tive binding ability to specific receptors andion channels (Olivera et al., 1990). Sincevenoms vary within and among species anddifferent venoms have different biologicalactivities, correct identification of species isessential for venom analysis.
In this study, we first hypothesized that toothcharacters might provide useful objective taxo-nomic characters if the range of variation withinindividuals, among individuals within species,and among species could be clearly docu-mented. We examined the radular teeth of 11molluscivorous Conus species. All are morph-ologically similar and their radular teeth belongto Type 1b of Nybakken (1990; Kohn et al.,1999) (Fig. 1). We analyzed intra- and inter-specific variation in the morphological charac-ters of radular teeth defined in Kohn et al.(1999). We also hypothesized that if tooth char-acters successfully differentiate species, theywould also differentiate geographic subspecies,and we tested this by applying the same pro-cedures to the radular teeth of specimens of onespecies from three different regions.
MATERIALS AND METHODS
We examined radular teeth of 118 specimens of 11molluscivorous Conus species for intra- and inter-specific variation (Table 1). We also analysed C. pen-naceus, a taxonomically particularly enigmatic entity,for geographic variation in tooth form among popula-tions in Hawaii, Maldives and Sumatra. We deter-mined species by shell characters according to Röckelet al. (1995).
The specimens were preserved in 70% ethanol with 5% glycerine. We removed the radular sac fromthe body cavity of Conus with forceps under the dissecting microscope. For morphometric analyses,we removed 1–6 radular teeth per specimen from the
radular caecum or short arm of the radular sac, wherefully formed teeth are stored. the teeth were mountedon slides with Aquamount and analyzed under aZeiss-Nomarski compound microscope. For visualiza-tion of teeth and their component parts, we preparedteeth for scanning electron microscopy as describedby Kohn et al. (1999).
We analyzed teeth both qualitatively and quantita-tively. We determined the presence or absence of firstand second barbs (B1, B2), blade (BL), cusp (cu) andspur (sp), presence or absence of serration (se) andwhether serration was continuous or interrupted, andwhether teeth were straight or measurably (>1%)curved. Quantitative measurements included abso-lute tooth length (TL), straight-line distance fromtooth tip to base (STL), length from apex to each barb(B1L, B2L), length from apex to proximal end of blade(BLL), length of serration (SEL), width of shaft(SHW), and width of base (BAW). Kohn et al. (1999)define these attributes explicitly, and they are shownin Figs. 1 and 2.
We used the following metric characters of theradular tooth, also defined in Kohn et al. (1999) forthe analysis of intra- and interspecific variations inteeth:
(1) Relative tooth length (TL/SL): ratio of toothlength to shell length.
(2) Relative first barb length (B1L/TL): ratio of dis-tance from apex to first barb, to tooth length.
(3) Relative second barb length (B2L/TL): ratio ofdistance from apex to second barb, to toothlength.
(4) Relative blade length (BLL/TL): ratio of distancefrom apex to blade, to tooth length.
(5) Relative length of serration (SEL/TL): ratio of serration length to tooth length.
(6) Relative width of shaft (SHW/TL): ratio of shaftwidth to tooth length.
(7) Relative width of base (BAW/TL): ratio of basewidth to tooth length.
(8) Relative width of shaft to base (SHW/BAW): ratioof shaft width to base width.
(9) Curvature (TL/STL): ratio of actual tooth lengthto the straight-line distance between its apex andbase. Measurements to determine curvature weremade on camera lucida tracings of teeth mountedon slides. Serration was determined to be contin-uous or interrupted also from mounted teethviewed with transmitted light.
We assumed different Conus species and theregional subgroups of C. pennaceus to be statisticallyindependent, since no hypotheses for species- andsubspecies-level phylogeny in Conus have been pro-
Figure 1. Scanning electron micrographs of entire radular teeth of molluscivorous Conus species, oriented withfirst barb to right, second barb to left. A, C. ammiralis; specimen 60 3 32 mm, from Majuro Atoll, MarshallIslands. B, C. canonicus; specimen 51 3 25 mm, from Pulo Bai, Batu Group, Sumatra, Indonesia. C, C. nodulo-sus; specimen 37 3 18 mm, from Long Island, Pelsaert Group, Houtman Abrolhos Islands, Australia. D, C. textile; specimen 35 3 16 mm, from Manuala, Ambon, Indonesia. E, C. omaria; specimen 38 3 18 mm, fromTabad Island, Madang Lagoon, Papua New Guinea. B1, first barb; B2, second barb; BA, base; cu, cusp; l, remnant of ligament; SH, shaft. Each scale bar 5 0.5 mm.
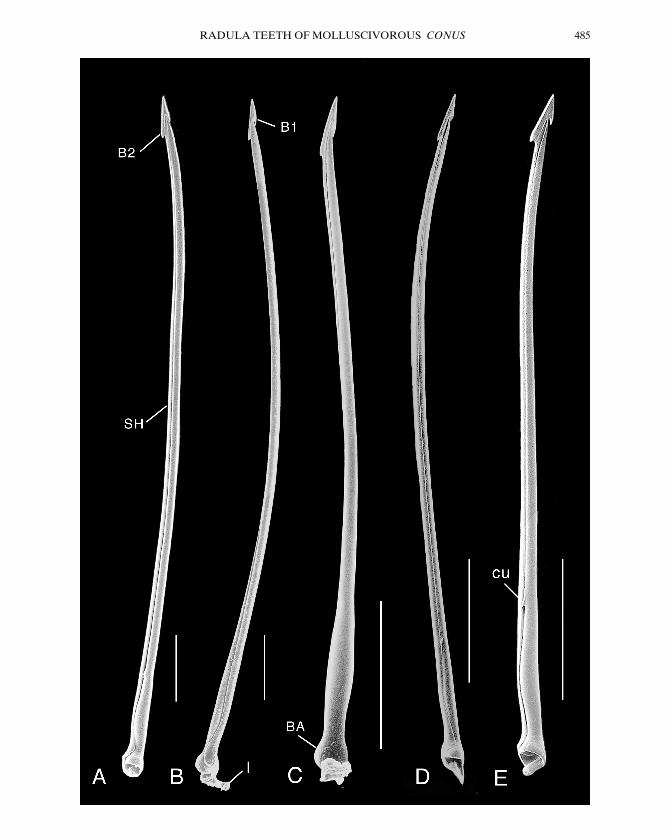
RADULA TEETH OF MOLLUSCIVOROUS CONUS 485

486 MANAMI NISHI & ALAN J. KOHN
posed. We examined the variation among teethwithin a specimen, among individuals within species,and among individuals from different geographicregions of C. pennaceus for each descriptive characterby calculating coefficients of variation (CV). We evaluated the distribution of each character amongspecies and among subgroups of C. pennaceus byusing single classification analysis of variance(ANOVA). To investigate which species or sub-species differed from each other by single characters,we carried out unplanned pairwise mean comparisontests between all possible pairs of taxa for each char-acter by using Kramer’s modification of Tukey’smethod (Sokal & Rohlf, 1981; Day & Quinn, 1989).For the preclassified species and the subgroups of C.pennaceus that were indistinguishable by pairwisecomparison tests, we carried out multivariate analysisof variance (MANOVA) to determine whether thesuite of all descriptive characters would distinguishthose species or subgroups, and whether multiple dis-criminant function analysis would correctly predictgroup membership.
Because each tooth had a second barb or a bladebut not both, and because only two of the three characters SHW/TL, BAW/TL and SHW/BAW are logi-cally independent, MANOVAs employed seven ofthe nine characters listed in Table 2. All statisticalanalyses were computed with the SYSTAT softwarepackage. In SYSTAT, MANOVAs use three types ofmultivariate test statistics, namely, Wilks lambda, theLawley-Hotelling trace and Pillai trace. We used thePillai trace, because it is more robust than otherswhen sample sizes are unequal and small (Tabachnick& Fidell, 1989). To test for correlations of the dif-ferent metric characters among species, we usedKendall’s coefficient of concordance, W (Sokal &Rohlf, 1981; see also Nunn & Smith, 1998). Differ-ences in specimen numbers for statistical analysesoccurred because some characters were either notmeasurable or not available for some specimens.
RESULTS
Discrete characters
All radular teeth of the molluscivorous Conusspecies we analyzed were curved, had eitherone barb and one blade or two barbs, eithercomplete or interrupted serration, and a cusp,but none had a spur. In all, the shaft diameterwas nearly uniform, that is without a waist.Analysis of these discrete radular tooth charac-ters (Table 2) led to classification of the 11species into the following three groups:
A. C. araneosus [Lightfoot], C. bandanusHwass and C. marmoreus Linnaeus haveteeth with one barb, one blade and continu-ous serration to the cusp (Kohn et al., 1999:Fig. 7).T
ab
le 1
.S
pec
ies,
nu
mb
ers
and
geo
gra
ph
ic s
ou
rces
of C
onus
spec
imen
s u
sed
for
rad
ula
r to
oth
an
alys
es. T
he
loca
lity
of o
ne
spec
imen
of C
.mar
mor
eus
is u
nkn
ow
n.
Mar
shal
lP
apu
aS
riR
edS
pec
ies
Haw
aii
Fiji
Isla
nd
sA
ust
ralia
New
Gu
inea
Ph
ilip
pin
esS
um
atra
Th
aila
nd
Lan
kaM
ald
ives
Sey
chel
les
Sea
To
tal
C. a
mm
iral
is L
inn
aeu
s4
004
C. a
rane
osus
[Lig
htf
oo
t]3
003
C. b
anda
nus
Hw
ass
500
5C
. can
onic
us H
was
s5
111
017
C. e
pisc
opat
us d
a M
ott
a1
1001
1C
. mar
mor
eus
Lin
nae
us
131
015
C. n
odul
osus
So
wer
by
500
5C
. om
aria
Hw
ass
13
51
009
C. p
enna
ceus
Bo
rn13
121
403
0C
. tex
tile
Lin
nae
us
42
15
21
015
C. v
icto
riae
Ree
ve4
004
118
RADULA TEETH OF MOLLUSCIVOROUS CONUS 487
B. C. ammiralis Linnaeus, C. canonicus Hwass,C. nodulosus Sowerby, C. textile Linnaeus,and C. victoriae Reeve have teeth with twobarbs and continuous serration (Fig. 1).
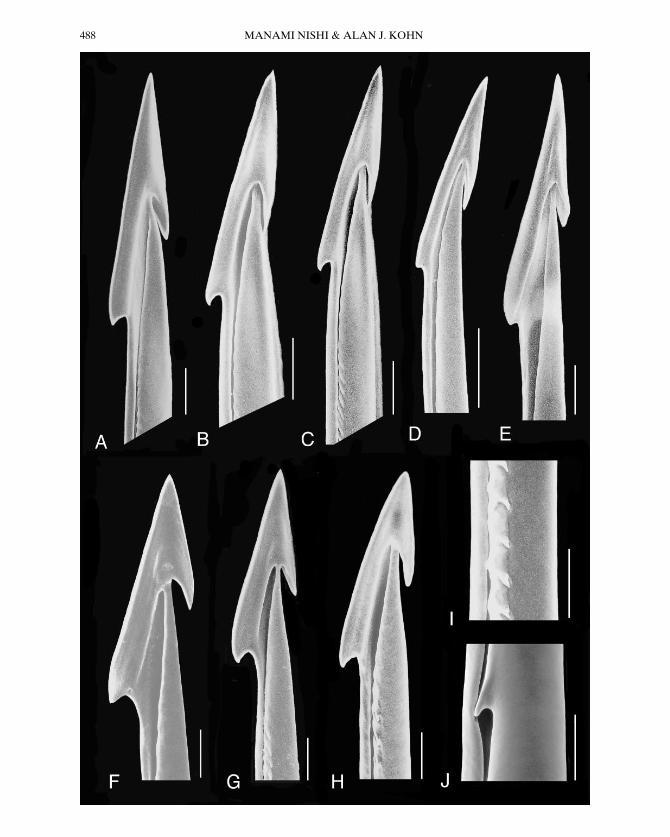
C. C. episcopatus da Motta, C. omaria Hwass,and C. pennaceus Born have teeth with twobarbs (Fig. 2), and the serration is inter-rupted by gaps.
Intra- and interspecific analysis of quantitativecharacters
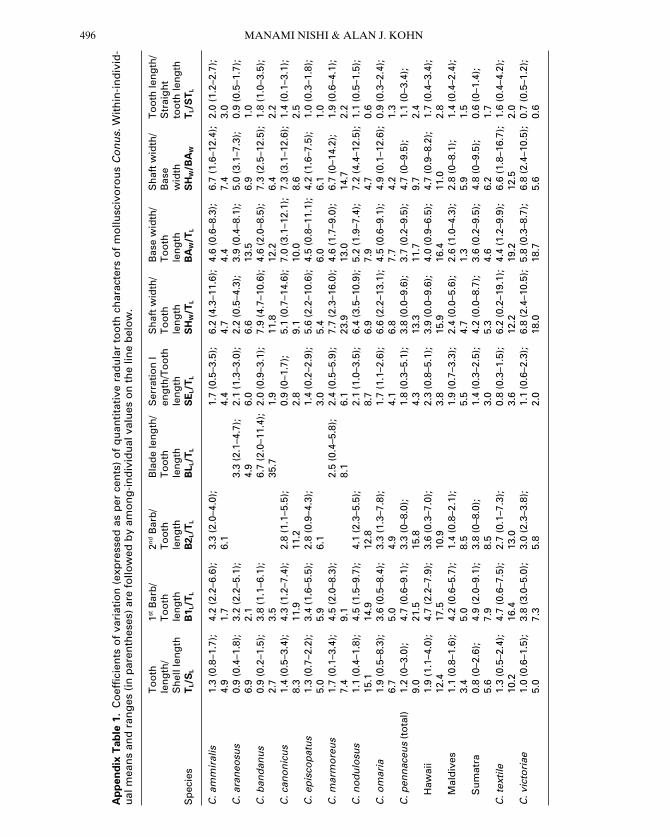
Variations in most metric tooth charactersamong teeth both within an individual andamong individuals within a species were small(average CV values of 0.6–7.7%) (AppendixTable 1). All quantitative tooth characters dif-fered significantly among species in ANOVAs(P < 0.001) (Table 3). In pairwise comparisontests, 96% (53 of 55) of all possible species pairsdiffered significantly with respect to one or moretooth characters (Appendix Table 2). BAW/TL
and SHW/BAW were used in MANOVAbecause BAW/TL distinguished species pairsmore strongly than SHW/TL (Appendix Table2).
All species within each species group sharethe same states of the four discrete characters,and if the model of the three species groupsdescribed in the previous section and based onthese states is valid, at least some of the quanti-
tative characters should fit the same expecta-tions. The distributions across species of five ofthese characters in fact supported the samespecies groups. The right portion of Table 2indicates that the rank orders of the values ofthe five metric characters segregate the speciesin Groups A and C. The species in Group Bwould be just as homogeneous were it not foranomalous values of C. nodulosus, discussedbelow. The seven metric characters present inall 11 species are highly significantly concordant(Kendall’s W 5 0.83; P , 0.001). For this testthe numerical rank orders of B1L/TL, BAW/TL,and SHW/TL were reversed to render them con-sistent with the other characters for the purposeof statistical analysis.
The three species comprising Group A byshared discrete characters grouped tightly withrespect to their quantitative character rankings.They have the shortest relative tooth length, theshortest relative serration length, the largestfirst barbs, and the widest bases relative to bothtooth length and shaft width (Table 2). Withinthis group, the teeth of Conus bandanus dif-fered significantly from those of C. araneosuswith respect to two quantitative characters (relative serration length and relative basewidth), and C. bandanus and C. marmoreusdiffered with respect to the former (AppendixTable 2). No single character distinguished C.araneosus from C. marmoreus, but MANOVA
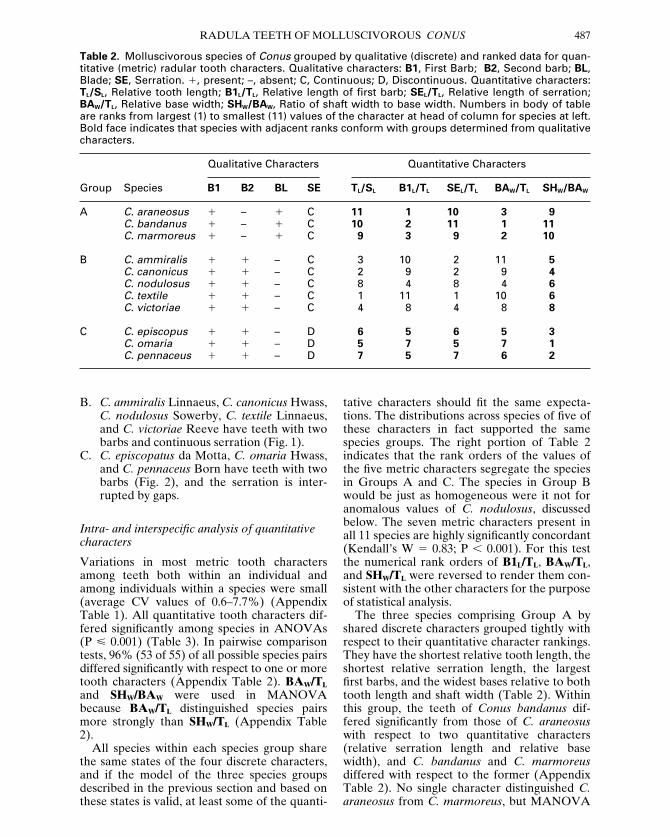
Table 2. Molluscivorous species of Conus grouped by qualitative (discrete) and ranked data for quan-titative (metric) radular tooth characters. Qualitative characters: B1, First Barb; B2, Second barb; BL,Blade; SE, Serration. 1, present; –, absent; C, Continuous; D, Discontinuous. Quantitative characters:TL/SL, Relative tooth length; B1L/TL, Relative length of first barb; SEL/TL, Relative length of serration;BAW/TL, Relative base width; SHW/BAW, Ratio of shaft width to base width. Numbers in body of tableare ranks from largest (1) to smallest (11) values of the character at head of column for species at left.Bold face indicates that species with adjacent ranks conform with groups determined from qualitativecharacters.
Qualitative Characters Quantitative Characters
Group Species B1 B2 BL SE TL/SL B1L/TL SEL/TL BAW/TL SHW/BAW
A C. araneosus 1 – 1 C 11 1 10 3 9C. bandanus 1 – 1 C 10 2 11 1 11C. marmoreus 1 – 1 C 9 3 9 2 10
B C. ammiralis 1 1 – C 3 10 2 11 5C. canonicus 1 1 – C 2 9 2 9 4C. nodulosus 1 1 – C 8 4 8 4 6C. textile 1 1 – C 1 11 1 10 6C. victoriae 1 1 – C 4 8 4 8 8
C C. episcopus 1 1 – D 6 5 6 5 3C. omaria 1 1 – D 5 7 5 7 1C. pennaceus 1 1 – D 7 5 7 6 2
488 MANAMI NISHI & ALAN J. KOHN
RADULA TEETH OF MOLLUSCIVOROUS CONUS 489
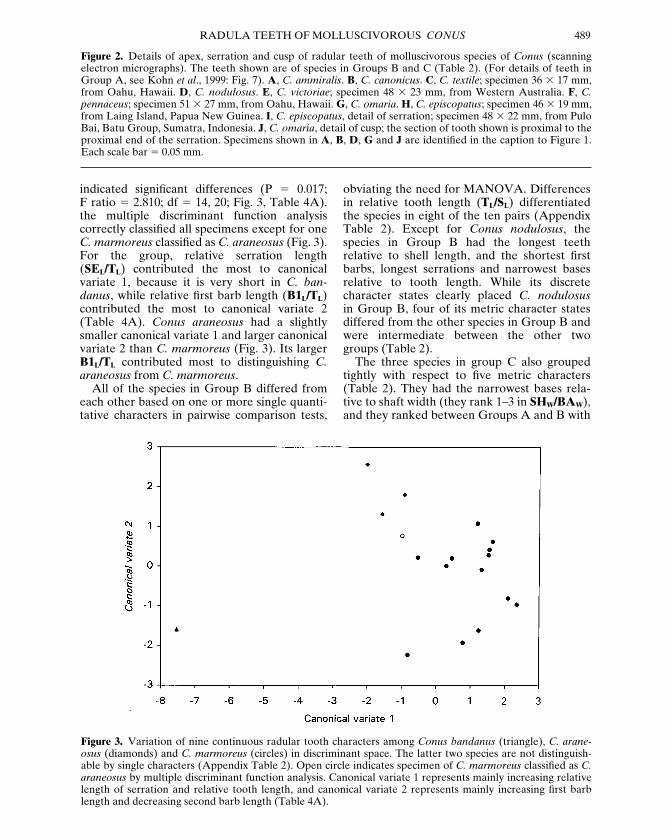
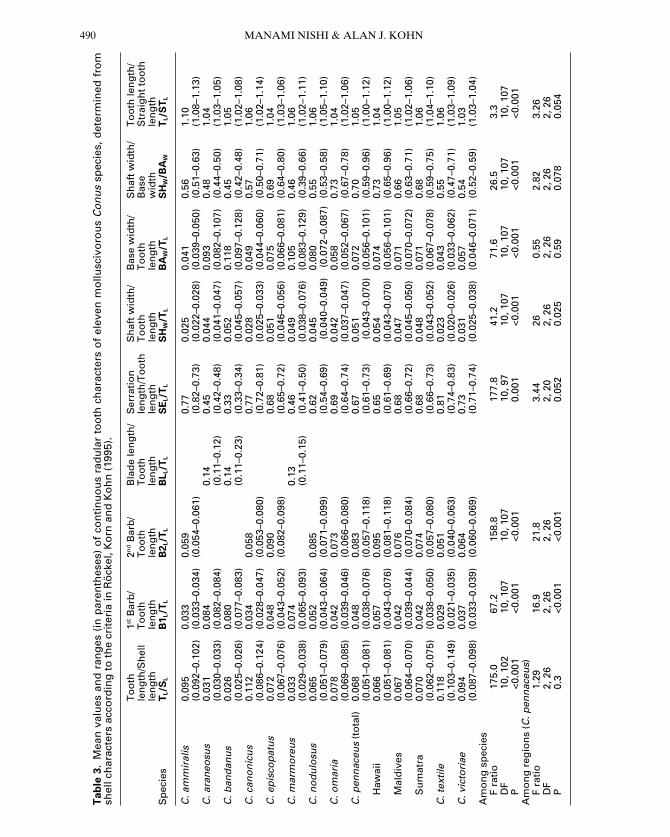
indicated significant differences (P 5 0.017; F ratio 5 2.810; df 5 14, 20; Fig. 3, Table 4A).the multiple discriminant function analysis correctly classified all specimens except for oneC. marmoreus classified as C. araneosus (Fig. 3).For the group, relative serration length(SEL/TL) contributed the most to canonicalvariate 1, because it is very short in C. ban-danus, while relative first barb length (B1L/TL)contributed the most to canonical variate 2(Table 4A). Conus araneosus had a slightlysmaller canonical variate 1 and larger canonicalvariate 2 than C. marmoreus (Fig. 3). Its largerB1L/TL contributed most to distinguishing C.araneosus from C. marmoreus.
All of the species in Group B differed fromeach other based on one or more single quanti-tative characters in pairwise comparison tests,
obviating the need for MANOVA. Differencesin relative tooth length (TL/SL) differentiatedthe species in eight of the ten pairs (AppendixTable 2). Except for Conus nodulosus, thespecies in Group B had the longest teeth relative to shell length, and the shortest firstbarbs, longest serrations and narrowest basesrelative to tooth length. While its discrete character states clearly placed C. nodulosusin Group B, four of its metric character statesdiffered from the other species in Group B andwere intermediate between the other twogroups (Table 2).
The three species in group C also groupedtightly with respect to five metric characters(Table 2). They had the narrowest bases rela-tive to shaft width (they rank 1–3 in SHW/BAW),and they ranked between Groups A and B with
Figure 2. Details of apex, serration and cusp of radular teeth of molluscivorous species of Conus (scanningelectron micrographs). The teeth shown are of species in Groups B and C (Table 2). (For details of teeth inGroup A, see Kohn et al., 1999: Fig. 7). A, C. ammiralis. B, C. canonicus. C, C. textile; specimen 36 3 17 mm,from Oahu, Hawaii. D, C. nodulosus. E, C. victoriae; specimen 48 3 23 mm, from Western Australia. F, C. pennaceus; specimen 51 3 27 mm, from Oahu, Hawaii. G, C. omaria. H, C. episcopatus; specimen 46 3 19 mm,from Laing Island, Papua New Guinea. I, C. episcopatus, detail of serration; specimen 48 3 22 mm, from PuloBai, Batu Group, Sumatra, Indonesia. J, C. omaria, detail of cusp; the section of tooth shown is proximal to theproximal end of the serration. Specimens shown in A, B, D, G and J are identified in the caption to Figure 1.Each scale bar 5 0.05 mm.
Figure 3. Variation of nine continuous radular tooth characters among Conus bandanus (triangle), C. arane-osus (diamonds) and C. marmoreus (circles) in discriminant space. The latter two species are not distinguish-able by single characters (Appendix Table 2). Open circle indicates specimen of C. marmoreus classified as C.araneosus by multiple discriminant function analysis. Canonical variate 1 represents mainly increasing relativelength of serration and relative tooth length, and canonical variate 2 represents mainly increasing first barblength and decreasing second barb length (Table 4A).
490 MANAMI NISHI & ALAN J. KOHN
Tab
le 3
.M
ean
val
ues
an
d r
ang
es (
in p
aren
thes
es)
of
con
tin
uo
us
rad
ula
r to
oth
ch
arac
ters
of
elev
en m
ollu
sciv
oro
us
Con
ussp
ecie
s, d
eter
min
ed f
rom
shel
l ch
arac
ters
acc
ord
ing
to
th
e cr
iter
ia in
Rö
ckel
, Ko
rn a
nd
Ko
hn
(19
95).
To
oth
1st
Bar
b/
2nd
Bar
b/
Bla
de
len
gth
/S
erra
tio
n
Sh
aft
wid
th/
Bas
e w
idth
/S
haf
t w
idth
/T
oo
th le
ng
th/
len
gth
/Sh
ell
To
oth
T
oo
th
To
oth
le
ng
th/T
oo
thT
oo
th
To
oth
B
ase
Str
aig
ht
too
th
len
gth
len
gth
len
gth
len
gth
len
gth
le
ng
thle
ng
thw
idth
le
ng
th
Sp
ecie
sT
L/S
LB
1L/T
LB
2L/T
LB
LL/T
LS
EL/T
LS
HW
/TL
BA
W/T
LS
HW
/BA
WT
L/S
TL
C. a
mm
iral
is0.
095
0.03
3 0.
059
0.
77
0.02
50.
041
0.56
1.
10
(0.0
92–0
.102
)(0
.033
–0.0
34)
(0.0
54–0
.061
)(0
.82–
0.73
)(0
.022
–0.0
28)
(0.0
39–0
.050
)(0
.51–
0.63
)(1
.08–
1.13
)C
. ara
neos
us0.
031
0.08
4 0.
14
0.45
0.
044
0.09
3 0.
48
1.04
(0
.030
–0.0
33)
(0.0
82–0
.084
)(0
.11–
0.12
)(0
.42–
0.48
)(0
.041
–0.0
47)
(0.0
82–0
.107
)(0
.44–
0.50
)(1
.03–
1.05
)C
. ban
danu
s0.
026
0.08
0 0.
14
0.33
0.
052
0.11
8 0.
45
1.05
(0
.025
–0.0
26)
(0.0
77–0
.083
)(0
.11–
0.23
)(0
.33–
0.34
)(0
.045
–0.0
57)
(0.0
97–0
.128
)(0
.42–
0.48
)(1
.02–
1.08
)C
. can
onic
us0.
112
0.03
4 0.
058
0.77
0.
028
0.04
9 0.
57
1.06
(0
.086
–0.1
24)
(0.0
28–0
.047
)(0
.053
–0.0
80)
(0.7
2–0.
81)
(0.0
25–0
.033
)(0
.044
–0.0
60)
(0.5
0–0.
71)
(1.0
2–1.
14)
C. e
pisc
opat
us0.
072
0.04
8 0.
090
0.68
0.
051
0.07
5 0.
69
1.04
(0
.067
–0.0
76)
(0.0
43–0
.052
)(0
.082
–0.0
98)
(0.6
5–0.
72)
(0.0
46–0
.056
)(0
.066
–0.0
81)
(0.6
4–0.
80)
(1.0
3–1.
06)
C. m
arm
oreu
s0.
033
0.07
4 0.
13
0.46
0.
049
0.10
5 0.
46
1.06
(0
.029
–0.0
38)
(0.0
65–0
.093
)(0
.11–
0.15
)(0
.41–
0.50
)(0
.038
–0.0
76)
(0.0
83–0
.129
)(0
.39–
0.66
)(1
.02–
1.11
)C
. nod
ulos
us0.
065
0.05
2 0.
085
0.62
0.
045
0.08
00.
55
1.06
(0
.051
–0.0
79)
(0.0
43–0
.064
)(0
.071
–0.0
99)
(0.5
4–0.
69)
(0.0
40–0
.049
)(0
.072
–0.0
87)
(0.5
3–0.
58)
(1.0
5–1.
10)
C. o
mar
ia0.
078
0.04
2 0.
073
0.69
0.
042
0.05
8 0.
73
1.04
(0
.069
–0.0
85)
(0.0
39–0
.046
)(0
.066
–0.0
80)
(0.6
4–0.
74)
(0.0
37–0
.047
)(0
.052
–0.0
67)
(0.6
7–0.
78)
(1.0
2–1.
06)
C. p
enna
ceus
(to
tal)
0.06
8 0.
048
0.08
3 0.
67
0.05
10.
072
0.70
1.
05
(0.0
51–0
.081
)(0
.038
–0.0
76)
(0.0
57–0
.118
)(0
.61–
0.73
)(0
.043
–0.0
70)
(0.0
56–0
.101
)(0
.59–
0.96
)(1
.00–
1.12
)H
awai
i0.
066
0.05
7 0.
095
0.65
0.
054
0.07
4 0.
73
1.04
(0
.051
–0.0
81)
(0.0
43–0
.076
)(0
.081
–0.1
18)
(0.6
1–0.
69)
(0.0
43–0
.070
)(0
.056
–0.1
01)
(0.6
5–0.
96)
(1.0
0–1.
12)
Mal
div
es0.
067
0.04
2 0.
076
0.68
0.
047
0.07
1 0.
66
1.05
(0
.064
–0.0
70)
(0.0
39–0
.044
)(0
.070
–0.0
84)
(0.6
6–0.
72)
(0.0
45–0
.050
)(0
.070
–0.0
72)
(0.6
3–0.
71)
(1.0
2–1.
06)
Su
mat
ra0.
070
0.04
2 0.
074
0.68
0.
048
0.07
1 0.
68
1.06
(0
.062
–0.0
75)
(0.0
38–0
.050
)(0
.057
–0.0
80)
(0.6
6–0.
73)
(0.0
43–0
.052
)(0
.067
–0.0
78)
(0.5
9–0.
75)
(1.0
4–1.
10)
C. t
extil
e0.
118
0.02
9 0.
051
0.81
0.
023
0.04
3 0.
55
1.06
(0
.103
–0.1
49)
(0.0
21–0
.035
)(0
.040
–0.0
63)
(0.7
4–0.
83)
(0.0
20–0
.026
)(0
.033
–0.0
62)
(0.4
7–0.
71)
(1.0
3–1.
09)
C. v
icto
riae
0.09
4 0.
037
0.06
40.
73
0.03
1 0.
057
0.54
1.
03
(0.0
87–0
.098
)(0
.033
–0.0
39)
(0.0
60–0
.069
)(0
.71–
0.74
)(0
.025
–0.0
38)
(0.0
46–0
.071
)(0
.52–
0.59
)(1
.03–
1.04
)A
mo
ng
sp
ecie
sF
rati
o17
5.0
67.2
158.
817
7.8
41.2
71.6
26.5
3.3
DF
10, 1
0210
, 107
10, 1
0710
, 97
10, 1
0710
, 107
10, 1
0710
, 107
P<0
.001
<0.0
01<0
.001
0.00
1<0
.001
<0.0
01<0
.001
<0.0
01A
mo
ng
reg
ion
s (C
. pen
nace
us)
F ra
tio
1.29
16.9
21.8
3.44
260.
552.
823.
26D
F2,
26
2, 2
62,
26
2, 2
02,
26
2, 2
62,
26
2, 2
6P
0.3
<0.0
01<0
.001
0.05
20.
025
0.59
0.07
80.
054
RADULA TEETH OF MOLLUSCIVOROUS CONUS 491
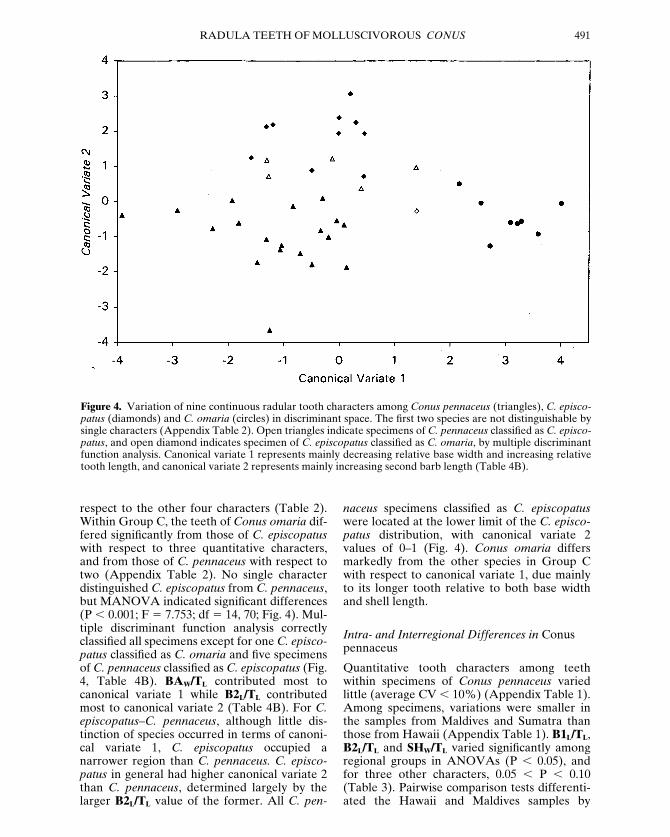
respect to the other four characters (Table 2).Within Group C, the teeth of Conus omaria dif-fered significantly from those of C. episcopatuswith respect to three quantitative characters,and from those of C. pennaceus with respect totwo (Appendix Table 2). No single characterdistinguished C. episcopatus from C. pennaceus,but MANOVA indicated significant differences(P , 0.001; F 5 7.753; df 5 14, 70; Fig. 4). Mul-tiple discriminant function analysis correctlyclassified all specimens except for one C. episco-patus classified as C. omaria and five specimensof C. pennaceus classified as C. episcopatus (Fig.4, Table 4B). BAW/TL contributed most tocanonical variate 1 while B2L/TL contributedmost to canonical variate 2 (Table 4B). For C.episcopatus–C. pennaceus, although little dis-tinction of species occurred in terms of canoni-cal variate 1, C. episcopatus occupied anarrower region than C. pennaceus. C. episco-patus in general had higher canonical variate 2than C. pennaceus, determined largely by thelarger B2L/TL value of the former. All C. pen-
naceus specimens classified as C. episcopatuswere located at the lower limit of the C. episco-patus distribution, with canonical variate 2 values of 0–1 (Fig. 4). Conus omaria differsmarkedly from the other species in Group Cwith respect to canonical variate 1, due mainlyto its longer tooth relative to both base widthand shell length.
Intra- and Interregional Differences in Conuspennaceus
Quantitative tooth characters among teethwithin specimens of Conus pennaceus varied little (average CV , 10%) (Appendix Table 1).Among specimens, variations were smaller inthe samples from Maldives and Sumatra thanthose from Hawaii (Appendix Table 1). B1L/TL,B2L/TL and SHW/TL varied significantly amongregional groups in ANOVAs (P , 0.05), andfor three other characters, 0.05 , P , 0.10(Table 3). Pairwise comparison tests differenti-ated the Hawaii and Maldives samples by
Figure 4. Variation of nine continuous radular tooth characters among Conus pennaceus (triangles), C. episco-patus (diamonds) and C. omaria (circles) in discriminant space. The first two species are not distinguishable bysingle characters (Appendix Table 2). Open triangles indicate specimens of C. pennaceus classified as C. episco-patus, and open diamond indicates specimen of C. episcopatus classified as C. omaria, by multiple discriminantfunction analysis. Canonical variate 1 represents mainly decreasing relative base width and increasing relativetooth length, and canonical variate 2 represents mainly increasing second barb length (Table 4B).
492 MANAMI NISHI & ALAN J. KOHN
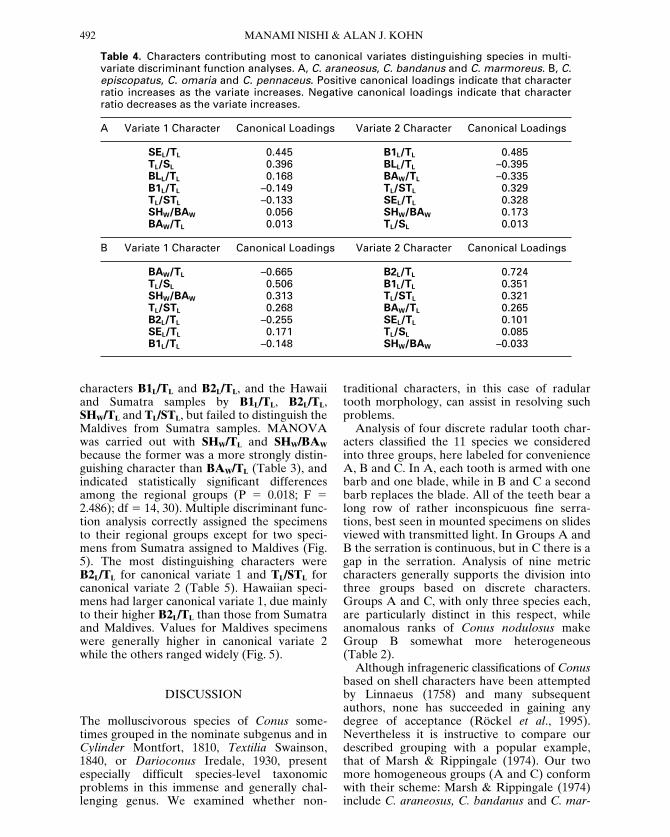
characters B1L/TL and B2L/TL, and the Hawaiiand Sumatra samples by B1L/TL, B2L/TL,SHW/TL and TL/STL, but failed to distinguish theMaldives from Sumatra samples. MANOVAwas carried out with SHW/TL and SHW/BAW
because the former was a more strongly distin-guishing character than BAW/TL (Table 3), andindicated statistically significant differencesamong the regional groups (P 5 0.018; F 52.486); df 5 14, 30). Multiple discriminant func-tion analysis correctly assigned the specimensto their regional groups except for two speci-mens from Sumatra assigned to Maldives (Fig.5). The most distinguishing characters wereB2L/TL for canonical variate 1 and TL/STL forcanonical variate 2 (Table 5). Hawaiian speci-mens had larger canonical variate 1, due mainlyto their higher B2L/TL than those from Sumatraand Maldives. Values for Maldives specimenswere generally higher in canonical variate 2while the others ranged widely (Fig. 5).
DISCUSSION
The molluscivorous species of Conus some-times grouped in the nominate subgenus and inCylinder Montfort, 1810, Textilia Swainson,1840, or Darioconus Iredale, 1930, presentespecially difficult species-level taxonomicproblems in this immense and generally chal-lenging genus. We examined whether non-
traditional characters, in this case of radulartooth morphology, can assist in resolving suchproblems.
Analysis of four discrete radular tooth char-acters classified the 11 species we consideredinto three groups, here labeled for convenienceA, B and C. In A, each tooth is armed with onebarb and one blade, while in B and C a secondbarb replaces the blade. All of the teeth bear along row of rather inconspicuous fine serra-tions, best seen in mounted specimens on slidesviewed with transmitted light. In Groups A andB the serration is continuous, but in C there is agap in the serration. Analysis of nine metriccharacters generally supports the division intothree groups based on discrete characters.Groups A and C, with only three species each,are particularly distinct in this respect, whileanomalous ranks of Conus nodulosus makeGroup B somewhat more heterogeneous(Table 2).
Although infrageneric classifications of Conusbased on shell characters have been attemptedby Linnaeus (1758) and many subsequentauthors, none has succeeded in gaining anydegree of acceptance (Röckel et al., 1995). Nevertheless it is instructive to compare ourdescribed grouping with a popular example,that of Marsh & Rippingale (1974). Our twomore homogeneous groups (A and C) conformwith their scheme: Marsh & Rippingale (1974)include C. araneosus, C. bandanus and C. mar-
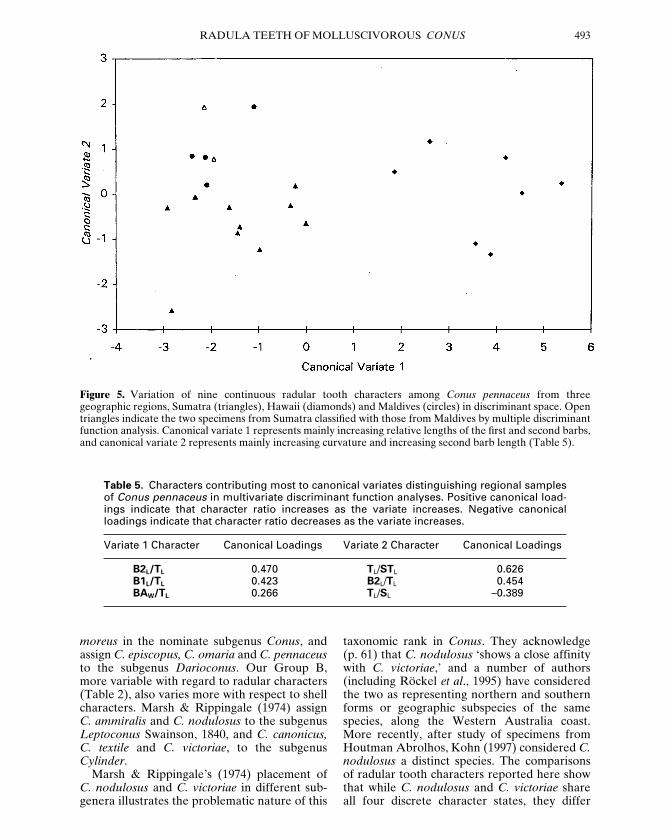
Table 4. Characters contributing most to canonical variates distinguishing species in multi-variate discriminant function analyses. A, C. araneosus, C. bandanus and C. marmoreus. B, C.episcopatus, C. omaria and C. pennaceus. Positive canonical loadings indicate that characterratio increases as the variate increases. Negative canonical loadings indicate that characterratio decreases as the variate increases.
A Variate 1 Character Canonical Loadings Variate 2 Character Canonical Loadings
SEL/TL 0.445 B1L/TL 0.485TL/SL 0.396 BLL/TL –0.395BLL/TL 0.168 BAW/TL –0.335B1L/TL –0.149 TL/STL 0.329TL/STL –0.133 SEL/TL 0.328SHW/BAW 0.056 SHW/BAW 0.173BAW/TL 0.013 TL/SL 0.013
B Variate 1 Character Canonical Loadings Variate 2 Character Canonical Loadings
BAW/TL –0.665 B2L/TL 0.724TL/SL 0.506 B1L/TL 0.351SHW/BAW 0.313 TL/STL 0.321TL/STL 0.268 BAW/TL 0.265B2L/TL –0.255 SEL/TL 0.101SEL/TL 0.171 TL/SL 0.085B1L/TL –0.148 SHW/BAW –0.033
RADULA TEETH OF MOLLUSCIVOROUS CONUS 493
moreus in the nominate subgenus Conus, andassign C. episcopus, C. omaria and C. pennaceusto the subgenus Darioconus. Our Group B,more variable with regard to radular characters(Table 2), also varies more with respect to shellcharacters. Marsh & Rippingale (1974) assignC. ammiralis and C. nodulosus to the subgenusLeptoconus Swainson, 1840, and C. canonicus,C. textile and C. victoriae, to the subgenusCylinder.
Marsh & Rippingale’s (1974) placement of C. nodulosus and C. victoriae in different sub-genera illustrates the problematic nature of this
taxonomic rank in Conus. They acknowledge(p. 61) that C. nodulosus ‘shows a close affinitywith C. victoriae,’ and a number of authors(including Röckel et al., 1995) have consideredthe two as representing northern and southernforms or geographic subspecies of the samespecies, along the Western Australia coast.More recently, after study of specimens fromHoutman Abrolhos, Kohn (1997) considered C.nodulosus a distinct species. The comparisonsof radular tooth characters reported here showthat while C. nodulosus and C. victoriae shareall four discrete character states, they differ
Figure 5. Variation of nine continuous radular tooth characters among Conus pennaceus from three geographic regions, Sumatra (triangles), Hawaii (diamonds) and Maldives (circles) in discriminant space. Opentriangles indicate the two specimens from Sumatra classified with those from Maldives by multiple discriminantfunction analysis. Canonical variate 1 represents mainly increasing relative lengths of the first and second barbs,and canonical variate 2 represents mainly increasing curvature and increasing second barb length (Table 5).
Table 5. Characters contributing most to canonical variates distinguishing regional samplesof Conus pennaceus in multivariate discriminant function analyses. Positive canonical load-ings indicate that character ratio increases as the variate increases. Negative canonical loadings indicate that character ratio decreases as the variate increases.
Variate 1 Character Canonical Loadings Variate 2 Character Canonical Loadings
B2L/TL 0.470 TL/STL 0.626B1L/TL 0.423 B2L/TL 0.454BAW/TL 0.266 TL/SL –0.389

494 MANAMI NISHI & ALAN J. KOHN
significantly with respect to more than half ofthe quantitative characters, thus supportingtheir separation as distinct species. Of all thespecies in Group B (Table 2) C. nodulosus hasthe most aberrant radular teeth. The other fourspecies have the longest teeth relative to shellsize and the longest serration relative to toothlength, while those of C. nodulosus are inter-mediate between those of groups A and C withrespect to both characters. Also the other fourhave the shortest first barb and the smallestbase relative to tooth length, while C. nodulo-sus again differs in being intermediate betweenthe other two groups.
Within Group B, each species is distinct fromall the others by at least one radular tooth char-acter, and eight of the ten species pairs differwith respect to tooth length : shell length ratio(Appendix Table 2). The two most similarspecies in radular tooth characters as well asshell morphology and colour pattern are C. textile and C. canonicus (Appendix Table 2;Röckel et al., 1995). Unlike most other speciespairs, their relative tooth lengths are identical,and they differ only with respect to relative serration length, moderately but significantlylarger in C. textile (Table 3, Appendix Table 2).The radular teeth of one pair of species in eachof Groups A and C were too similar to be differentiated by any of the single charactersemployed, although multiple discriminant function analysis distinguished them. In bothgroups, the most similar radular teeth are notthose of the two species with most similar shellcharacters. In Group A, C. araneosus and C.bandanus have the most similar shells (Röckelet al., 1995), while C. araneosus and C. mar-moreus have the most similar teeth (AppendixTable 2). In Group C, the shells of C. omariaand C. pennaceus have long been the subject oftaxonomic confusion (Kohn, 1959; Röckel et al.,1995), while C. pennaceus and C. episcopatushave the most similar teeth (Appendix Table 2).
Within species, Conus pennaceus is known tobe conchologically very diverse both within andbetween local populations (Röckel et al., 1995).Shell characters are rather uniform in mostIndian Ocean populations while there is a widerange of variation in some regions, such asMozambique and Hawaii. Radular charactersthus conform with the regional patterns of shellcharacters, as coefficients of variation of toothcharacters were larger (.10%) among Hawai-ian specimens than in samples from Sumatraand Maldives (Appendix Table 1).
In conclusion, in the present study, radulartooth characters differentiated 11 molluscivo-
rous species as well as three regional popula-tions of C. pennaceus, although in some casesthis required multivariate statistical analyses.Discrete and metric radular tooth charactersalso assort the species into three groups, whichgenerally conform to some subdivisions of thegenus based on shell characters. Moreover,although application of molecular methods toConus systematics is just beginning, a prelimi-nary cladistic analysis of 18S rRNA genesequences, ((araneosus, marmoreus) ((canoni-cus, textile) (episcopus, pennaceus))) (Duda andPalumbi, in prep.) is also congruent with thegroups based on our radular analyses. Ourresults thus show that radular tooth charactersprovide potentially useful species- and sub-species-level taxonomic information. We believethese results are sufficiently promising thatanalyses of radular tooth morphometry shouldbe extended to the approximately 20 additionalknown species in the molluscivorous subgroupsof Conus. Analysis of such a more completedata set in concert with other morphologicalcharacters and with molecular genetic dataholds promise of generating and testing objec-tive hypotheses for understanding the phylo-geny of the genus.
ACKNOWLEDGEMENTS
This work was supported by a grant from HowardHughes Medical Institute to the University ofWashington Biology Program and by a grant from the University of Washington Royalty ResearchFund. Grants from the National Science Foundation,most recently BSR-8700523, supported collection of material. We thank Bruno Pernet for taking the scanning electron micrographs and Rebecca Price fortechnical assistance
REFERENCES
DAY, R.W. & QUINN, G.P. 1989. Comparisons oftreatments after an analysis of variance in ecology.Ecological Monographs, 59: 433-463.
KOHN, A.J. 1956. Piscivorous gastropods of the genusConus. Proceedings of the National Academy ofSciences, 42: 168-171.
KOHN, A.J. 1959. The Hawaiian species of Conus.Pacific Science, 13: 368-401.
KOHN, A.J. 1990. Tempo and mode of evolution ofConidae. Malacologia, 32: 55-67.
KOHN, A.J. 1997. Ecological and biogeographicattributes of Conus on the Indian Ocean’s southern-most coral reefs. In: The Marine Flora and Fauna ofthe Houtman Abrolos Islands, Western Australia;Proceedings of the Seventh International MarineBiological Workshop (F.E. Wells, ed.), 113-131.Western Australian Museum, Perth.

RADULA TEETH OF MOLLUSCIVOROUS CONUS 495
KOHN, A.J. 1998. Family Conidae. In: Mollusca: TheSouthern Synthesis. Fauna of Australia, vol. 5. (P.L.Beesley, G.J.B. Ross & A. Wells, eds) 852-854.CSIRO Publishing, Melbourne.
KOHN, A., NISHI, M. & PERNET, B. 1999. Snail spearsand scimitars: a character analysis of Conus radularteeth. Journal of Molluscan Studies, 68: 461-481.
LINNAEUS, C. 1758. Systema naturae per regna tria naturae, 10th ed. Stockholm.
MARSH, J.A. & RIPPINGALE, O.H. 1974. Cone shells ofthe world, 3rd edition. Jacaranda Press, Brisbane.
MORRISON, F.D. 1976. Multivariate statistical methods.McGraw-Hill, New York.
NUNN, C.L. & SMITH, K.K. 1998. Statistical analysesof developmental sequences: the craniofacial regionin marsupial and placental mammals. AmericanNaturalist, 152: 82-101.
NYBAKKEN, J. 1990. Ontogenetic change in the Conusradula, its form, distribution among the radulatypes, and significance in systematics and ecology.Malacologia, 32: 35-54.
OLIVERA, B.M., RIVIER, J., CLARK, C., RAMILO, C.A.,CORPUZ, G.C., ABOGADIE, F.C., MENA, E.E.,WOODWARD, S.R., HILLYARD, D.R. & CRUZ, L.J.1990. Diversity of Conus neuropeptides. Science,249: 257-263.
RÖCKEL, D., KORN, W. & KOHN, A.J. 1995. Manual of the living Conidae, Vol. I. Indo-Pacific Region.Verlag Christa Hemmen, Wiesbaden.
SOKAL, R.R. & ROHLF, F.J. 1981. Biometry, Secondedition. Freeman, San Francisco.
TABACHNICK, B.G. & FIDELL, L.S. 1989. Using multi-variate statistics. Harper & Row, New York.
496 MANAMI NISHI & ALAN J. KOHN
Ap
pen
dix
Tab
le 1
.C
oef
fici
ents
of
vari
atio
n (
exp
ress
ed a
s p
er c
ents
) o
f q
uan
tita
tive
rad
ula
r to
oth
ch
arac
ters
of
mo
llusc
ivo
rou
s C
onus
.Wit
hin
-in
div
id-
ual
mea
ns
and
ran
ges
(in
par
enth
eses
) ar
e fo
llow
ed b
y am
on
g-i
nd
ivid
ual
val
ues
on
th
e lin
e b
elo
w.
To
oth
1st
Bar
b/
2nd
Bar
b/
Bla
de
len
gth
/S
erra
tio
n l
Sh
aft
wid
th/
Bas
e w
idth
/S
haf
t w
idth
/T
oo
th le
ng
th/
len
gth
/T
oo
th
To
oth
T
oo
th
eng
th/T
oo
th
To
oth
T
oo
th
Bas
e S
trai
gh
t S
hel
l len
gth
le
ng
th
len
gth
le
ng
th
len
gth
le
ng
th
len
gth
w
idth
to
oth
len
gth
S
pec
ies
TL/S
LB
1L/T
LB
2L/T
LB
LL/T
LS
EL/T
LS
HW
/TL
BA
W/T
LS
HW
/BA
WT
L/S
TL
C. a
mm
iral
is1.
3 (0
.8–1
.7);
4.
2 (2
.2–6
.6);
3.
3 (2
.0–4
.0);
1.
7 (0
.5–3
.5);
6.
2 (4
.3–1
1.6)
; 4.
6 (0
.6–8
.3);
6.
7 (1
.6–1
2.4)
; 2.
0 (1
.2–2
.7);
4.
91.
76.
14.
44.
74.
47.
43.
0C
. ara
neos
us0.
9 (0
.4–1
.8);
3.
2 (2
.2–5
.1);
3.
3 (2
.1–4
.7);
2.
1 (1
.3–3
.0);
2.
2 (0
.5–4
.3);
3.
9 (0
.4–8
.1);
5.
0 (3
.1–7
.3);
0.
9 (0
.5–1
.7);
6.
92.
14.
96.
06.
613
.56.
91.
0C
. ban
danu
s0.
9 (0
.2–1
.5);
3.
8 (1
.1–6
.1);
6.
7 (2
.0–1
1.4)
; 2.
0 (0
.9–3
.1);
7.
9 (4
.7–1
0.6)
; 4.
6 (2
.0–8
.5);
7.
3 (2
.5–1
2.5)
; 1.
8 (1
.0–3
.5);
2.
73.
535
.71.
911
.812
.26.
42.
2C
. can
onic
us1.
4 (0
.5–3
.4);
4.
3 (1
.2–7
.4);
2.
8 (1
.1–5
.5);
0.
9 (0
–1.7
);
5.1
(0.7
–14.
6);
7.0
(3.1
–12.
1);
7.3
(3.1
–12.
6);
1.4
(0.1
–3.1
);
8.3
11.9
11.2
2.8
9.1
10.0
8.6
2.5
C. e
pisc
opat
us1.
3 (0
.7–2
.2);
3.
4 (1
.6–5
.5);
2.
8 (0
.9–4
.3);
1.
4 (0
.2–2
.9);
5.6
(2.2
–10.
6);
4.5
(0.8
–11.
1);
4.2
(1.6
–7.5
);
1.0
(0.3
–1.8
);
5.0
5.9
6.1
3.0
5.4
6.0
6.1
1.0
C. m
arm
oreu
s1.
7 (0
.1–3
.4);
4.
5 (2
.0–8
.3);
2.
5 (0
.4–5
.8);
2.
4 (0
.5–5
.9);
7.
7 (2
.3–1
6.0)
; 4.
6 (1
.7–9
.0);
6.
7 (0
–14.
2);
1.9
(0.6
–4.1
);
7.4
9.1
8.1
6.1
23.9
13.0
14.7
2.2
C. n
odul
osus
1.1
(0.4
–1.8
);
4.5
(1.5
–9.7
);
4.1
(2.3
–5.5
);
2.1
(1.0
–3.5
);6.
4 (3
.5–1
0.9)
; 5.
2 (1
.9–7
.4);
7.
2 (4
.4–1
2.5)
; 1.
1 (0
.5–1
.5);
15
.114
.912
.88.
76.
97.
94.
70.
6C
. om
aria
1.9
(0.5
–8.3
);
3.6
(0.5
–8.4
);
3.3
(1.3
–7.8
);
1.7
(1.1
–2.6
);
6.6
(2.2
–13.
1);
4.5
(0.6
–9.1
);
4.9
(0.1
–12.
6);
0.9
(0.3
–2.4
);
6.7
5.0
4.9
4.1
6.8
7.7
4.2
1.3
C. p
enna
ceus
(to
tal)
1.2
(0–3
.0);
4.
7 (0
.6–9
.1);
3.
3 (0
–8.0
);
1.8
(0.3
–5.1
);
3.8
(0.0
–9.6
);
3.7
(0.2
–9.5
);
4.7
(0–9
.5);
1.
1 (0
–3.4
);
9.0
21.5
15.8
4.3
13.3
11.7
9.7
2.4
Haw
aii
1.9
(1.1
–4.0
);
4.7
(2.2
–7.9
);
3.6
(0.3
–7.0
);
2.3
(0.8
–5.1
);
3.9
(0.0
–9.6
);
4.0
(0.9
–6.5
);
4.7
(0.9
–8.2
);
1.7
(0.4
–3.4
);
12.4
17.5
10.9
3.8
15.9
16.4
11.0
2.8
Mal
div
es1.
1 (0
.8–1
.6);
4.
2 (0
.6–5
.7);
1.
4 (0
.8–2
.1);
1.
9 (0
.7–3
.3);
2.
4 (0
.0–5
.6);
2.
6 (1
.0–4
.3);
2.
8 (0
–8.1
);
1.4
(0.4
–2.4
);
3.4
5.0
8.5
5.5
4.7
1.3
5.9
1.5
Su
mat
ra0.
8 (0
–2.6
);
4.9
(2.0
–9.1
);
3.8
(0–8
.0);
1.
4 (0
.3–2
.5);
4.
2 (0
.0–8
.7);
3.
6 (0
.2–9
.5);
4.
8 (0
–9.5
);
0.6
(0–1
.4);
5.
67.
98.
53.
05.
34.
66.
21.
7C
. tex
tile
1.3
(0.5
–2.4
);
4.7
(0.6
–7.5
);
2.7
(0.1
–7.3
);
0.8
(0.3
–1.5
);
6.2
(0.2
–19.
1);
4.4
(1.2
–9.9
);
6.6
(1.8
–16.
7);
1.6
(0.4
–4.2
);
10.2
16.4
13.0
3.6
12.2
19.2
12.5
2.0
C. v
icto
riae
1.0
(0.6
–1.5
);
3.8
(3.0
–5.0
);
3.0
(2.3
–3.8
);
1.1
(0.6
–2.3
);
6.8
(2.4
–10.
5);
5.8
(0.3
–8.7
);
6.8
(2.4
–10.
5);
0.7
(0.5
–1.2
);
5.0
7.3
5.8
2.0
18.0
18.7
5.6
0.6

RADULA TEETH OF MOLLUSCIVOROUS CONUS 497
Ap
pen
dix
Tab
le 2
.In
div
idu
al c
on
tin
uo
us
rad
ula
r ch
arac
ters
th
at s
ign
ifica
ntl
y (a
5.0
5) d
iffe
ren
tiat
e m
emb
ers
of
Con
ussp
ecie
s p
airs
fro
m e
ach
oth
er,
bas
ed o
n A
NO
VA
an
d T
uke
y-K
ram
er u
np
lan
ned
co
mp
aris
on
tes
ts.
Key
to
ch
arac
ter
nu
mb
ers:
1,
TL/S
L;
2, B
1L/T
L;
3, B
2L/T
L;
4, B
LL/T
L;
5, S
EL/T
L;
6,S
HW
/TL; 7
, BA
W/T
L; 8
, SH
W/B
AW
; 9,T
L/S
TL.
Sp
ecie
sC
. ara
neos
usC
. ban
danu
sC
. mar
mor
eus
C. a
mm
iral
is
C. c
anon
icus
C. n
odul
osus
C. t
extil
eC
. vic
tori
aeC
. epi
scop
atus
C. o
mar
iaC
. pen
nace
us
C. a
rane
osus
5, 7
1–7
1–7
1–5
1–7
1–5,
71–
5, 7
, 81–
5, 7
, 81–
5, 7
, 8C
. ban
danu
s5
1–7
1–8
1–5,
71–
71–
71–
5, 7
, 81–
5, 7
, 81–
5, 7
, 8C
. mar
mor
eus
1–7
1–8
1–5,
71–
81–
71–
5, 7
, 81–
5, 7
, 81–
5, 7
, 8C
. am
mir
alis
11–
3, 5
–71
91–
3, 5
–91,
5, 6
, 8, 9
1–3,
5–9
C. c
anon
icus
1–3,
5–7
51
1–3,
6–8
1, 2
, 5, 6
, 9 1
–3, 5
–8C
. nod
ulos
us1–
3, 5
–71–
3, 5
, 75,
81,
5, 8
, 95,
8C
. tex
tile
1, 5
1–3,
5, 7
, 81–
3, 5
–7, 8
1–3,
5–8
C. v
icto
riae
1, 3
, 7, 8
1, 9
1–3,
5–8
C. e
pisc
opat
us3,
6, 7
C. o
mar
ia1,
7
