purification of a novel eif-2α protein kinase from calf brain
TRANSCRIPT

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
February 14, 1990 Pages 1237-1244
PURIFICATION OF A NOVEL eIF-2a PROTEIN KINASE FROM CALF BRAIN
Albert0 Alc&za?+, Enrique MBndez*, M-Elena Martin' and Matilde Salinas*l
'Servicio de Bioquimica, Dpto. Investigacibn, and *Servicio de
Endocrinologia, Hospital Ram6n y Cajal, Ctra. Colmenar Km.9 28034
Madrid, Spain
Received December 8, 1989
A new eukaryotic initiation factor 2 kinase has been purified for the first time from calf brain cytosol. The purification of a nonabundant novel protein kinase activity, designated as PKI, that phosphorylates the a subunit of eukaryotic initiation factor 2 is described. The protein kinase activity was assayed using purified initiation factor 2 as a sub- strate and ,was purified by ammonium sulphate precipitation, con- ventional chromatography in heparin-Sepharose and phosphocel- lulose and by high performance size exclusion and anion exchange chromatographies. The protein kinase activity elutes in the region of 140,000 in the size exclusion chromatography and is as- sociated with two different polypeptides a and b, with relative molecular masses of 38,000 and 20,000 and an approximate ratio of 2.5-3.0:1 . The protein kinase does not phosphorylate casein or histones and it is independent of cyclic nucleotides. It can be classified as a serine kinase since the phosphorylation of the a subunit of eIF-2 is produced in serine residues. Under these con- ditions none of the kinase subunits are phosphorylated. B 1990 Academic Press, Inc.
Eukaryotic initiation factor 2 (eIF-2) is involved in the
initial steps of peptide chain initiation, which appear to be a
general control point in the short-term control of translation in response to stimuli such as hormones, interferon and/or viral in-
fection fertilization of oocytes
(1).
and environmental conditions
*To whom correspondence should be addressed.
Abbreviations: eIF-2a, a subunit 2; eIF-2. eukarvotic initiation inhibitor: DAI, -double-stranded eIF-2a kinase from the brain.
of eukaryotic initiation factor factor 2; HCI, hemin-controlled RNA activated inhibitor; PKI,
.m 1231
0006-291X/90 $1.50 Copyright 0 1990 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Phosphorylation of the smallest, or a, subunit of eIF-2
(eIF-2a) inhibits peptide initiation in mammalian reticulocytes.
This translational regulation of protein synthesis is mediated by
either one of two different substrate-specific, c-AMP independent protein kinases that phosphorylate the same site(s) (2) on eIF-2a and thereby block the release of GDP from the eIF-2 catalyzed by
its GDP exchange factor (1,2). One protein kinase is activated under conditions of heme deficiency and is known as the heme-
controlled inhibitor (HCI) (3) and the other is induced by inter- feron and is activated by double-stranded RNA (DA11 (4).
Protein kinase activities that can act on the a-subunit of
eIF-2 have been identified in a wide variety of cells and
tissues, though none have been characterized as extensively as have HCI and DA1 from reticulocytes (1,5,6,7,8,9, 10). Whilst the
eventual control mechanism at the initiation level may resemble
those seen in the reticulocyte, the physiological signals induc-
ing those effects in eIF-2a are almost certainly different. Then, the search for HCI or DA1 activity in other mammalian cells and
the characterization of new eIF-2a kinases is a matter of inten-
sive work in this field.
Results obtained over the last five years in our laboratory
have shown that eIF-2 factor phosphorylation and/or eIF-2 subcel-
lular distribution may be implicated in the regulation of the
ontogenetic-related changes in protein synthesis observed in
brain tissue (9,11,12). Here we describe the identification,
purification and partial characterization of a novel eIF-2a
kinase from calf brain. The possible modulators of this kinase,
the specific site(s) for eIF-2 that it phosphorylates and its "in
viva" regulatory role remain to be studied.
MATERIAL AND METHODS
eIF-2 factor purification.- Initiation factor 2 was purified from the ribosomal salt wash fraction from calf brain according to our recently described method (131, which consists of precipitation with ammonium sulphate followed by three chromatographic steps, in heparin-Sepharose, phosphocellulose and GDP-agarose. Gel electrophoresis.- The protein fractions from the different chromatographic steps and the phosphorylation reactions were analyzed by SDS-electrophoresis on polyacrylamide gels, using 12% of acrylamide and 0.12% of N,N'methylene-bisacrylamide as described (14). Gels were stained with silver reagent. eIF-2a kinase assay.- Protein kinase assay was performed in a final volume of 50 ul containing: 50 mM Hepes (N-2-hydroxyethyl- piperazine-N'-2-ethanesulfonic acid), pH 7.5; 150 mM potassium acetate; 5 mM magnessium acetate: 0.5 mM dithiothreitol; and 0.32 M sucrose. Before beginning the incubation 1 ug of purified eIF-2
1238

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
factor was added to the mixture as substrate for the enzyme and 0.2-2.0 pg of the protein fraction containing the kinase activity. After 2 min preincubation at 30°C, the reaction was started by the addition of [T- 92 PI ATP (25-50 @l, 5 VCi) and main- tained for 10 min. The incubation mixture was analyzed by SDS- electrophoresis as described above and the gels were washed, dried, and autoradiographed. Both, silver stained gels and autoradiographies were scanned at 450 and 800 nm respectively in a DU-8 Beckman spectrophotometer. Protein was determined by Bradford's method (15). Purification of the eIF-2a kinase.- The eIF-2a kinase was purified using the above described purification procedure for eIF-2 factor followed by two additional high performance size ex- clusion and anion exchange steps. 3.4 mg of the non-bound frac- tion from the GDP-agarose chromatography were used as a source of the brain eIF-2a kinase. The size-exclusion chromatography was performed on a Superose 6 HR lo/30 from Pharmacia by isocrating elution with buffer A (20 mM Tris-HCl, pH 7.6; 0.5 mM phenyl- methylsulphonyl fluoride; 7 mM 2-mercaptoethanol; 1.0 mM EDTA; 10 % glycerol, containing 50 mM Xl). The column was operated at room temperature at a flow rate of 0.3 ml/min and 1 ml fractions were collected on ice. The activity containing fractions (0.3 mg protein) were loaded on a protein anion exchange column (7,5 mm x 7.5 cm) (Protein Pack Glass DEAE-5PW from Waters) equilibrated with buffer A. The column was eluted with a KC1 lineal gradient and operated at 4OC at a flow rate of 0.3 ml/min. Fractions (0.9 ml) were collected and assayed for eIF-2a kinase activity. Both chromatographic steps were performed with Waters 650 equipment. Phosphoamino Acid Anavsis.- eIF-2 (1 pg) was phosphorylated by PKI (0.1 pg protein, fraction 49 from DEAE-5PW column 1 as described above, the excess of [~ZP]-ATP was eliminated by suces- sively diluting and concentrating the samples using microcon- centrators (Centricon 10 Amicon) down to a volume of 150 ~1. The samples were freeze-dried, and hydrolyzed in 6M HCl for 2h at 1lOOC. The hydrolates were derivatized with o-phthaladehyde (16) and then analyzed by RP-HPLC using a C-18 (150 x 4.6 cm) Ultrasphere-ODS Beckman column. A Gilson Spectra-G10 fluorescence detector equipped with 360 nm excitation and 425 nm emision fil- ters were used. Phosphoserine, phosphothreonine and phos- photyrosine were derivatized with o-phthalaldehyde and used as standards. Radioactivity was measured in 0.5 min samples by liq- uid scintillation counting. Samples of PKI (0.1 fig) and eIF-2 (1 clg) alone, were phosphorylated and processed in the same way as a background of the reaction.
RESULTS AND DISCUSSION
This report describes for the first time the purification of a new protein kinase which phosphorylate the a subunit of eIF-2 factor from an animal tissue " in vitro". During the purification of the eIF-2 factor from the postmicrosomal fraction
of calf brain, which followed the same procedure as that described for the factor from the ribosomal salt wash fraction (131, we observed that the protein fraction that did not bind to
the GDP-agarose column, still had eIF-2 factor. What is more, the a subunit of the factor was phosphorylated in the presence of
ATP-Mg, indicating the existence of an endogenous eIF-2a protein
1239

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0.4
0.3
0.2
OS1
0
Absorbance at 280 nm
Fraction number Figure 1. Fractionation by size-exclusion of the calf brain eIF- 2a kinase activity present in the non-bound fraction from the GDP-agarose column. Sample (3.4 mg), was applied to a Superose 6 column and eluted with an isocratic buffer as described in Material and Methods. The solid line represents absorbance at 28Onm and shaded bars indicate fractions where PKI activity is eluted. The autoradiography corresponding to the kinase assay performed as detailed in Material and Methods is inserted in the top-left corner of the figure. Aliguots of 15 pl from the dif- ferent fractions were used as a kinase source. G: 6 pg protein mixture from the non-bound fraction from GDP-agarose. The posi- tion of the a subunit of eIF-2 factor is marked by an arrow.
kinase. A sample (3.4 mg) of this protein fraction was loaded on
a Superose 6 column and protein kinase activity was identified by monitoring the incorporation of radioactive phosphate into
purified eIF-2 factor as can be seen in Fig. 1. The eIF-2a kinase
activity was eluted in fractions 16, 17 and 16 (eIF-2 factor co-
eluted in the same fractions) but fraction 16 was discarded be-
cause of its high endogenous phosphorylation, probably due to the presence of other protein kinases. Fractions 17 and 18, contain-
ing about 0.3 mg protein, were applied to a DEAE-5PW column and
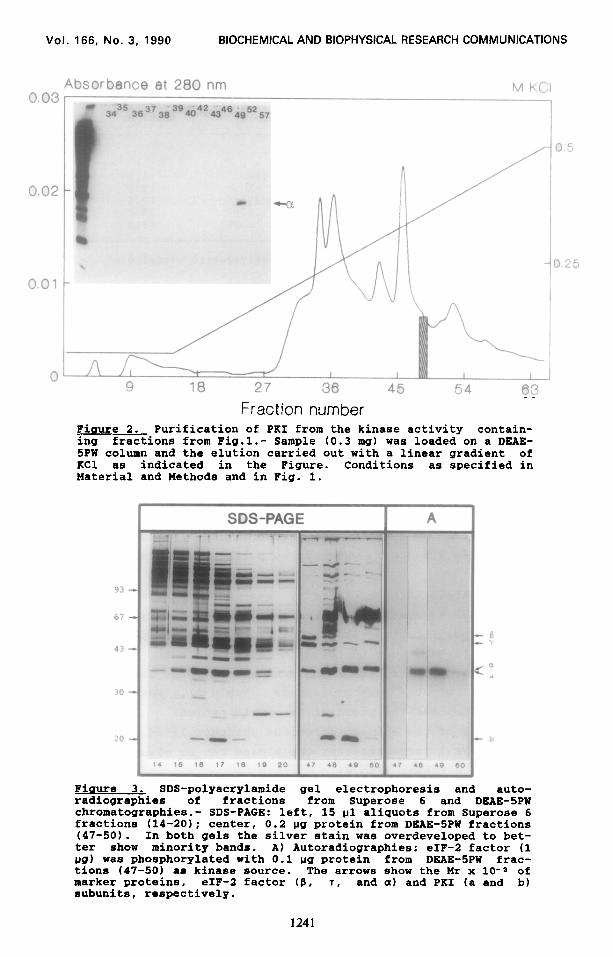
the activity was only detected in one of the fractions assayed (Fig.2). The fractions before and after fraction 49 were analyzed by gel electrophoresis and kinase activity. Although all of the fractions contained minority bands (Fig.31, the kinase activity was lower in fraction 48 than in fraction 49, even lower in frac-
tion 50 and undetectable in fraction 47 (in spite of containing
1240
Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0.03
0.02
0.01
0
Absorbance et 280 nm M KCI
0.5
0.25
54 63
Fraction number Figure 2. Purification of PKI from the kinase activity contain- ing fractions from Fig.l.- Sample (0.3 mg) was loaded on a DEAE- 5PW column and the elution carried out with a linear gradient of KC1 as indicated in the Figure. Conditions as specified in Material and Methods and in Pig. 1.
67
Figure 3. SDS-polyacrylamide gel electrophoresis and auto- radiographies of fractions from Superose 6 and DKAE-SPW chromatographies.- SDS-PAGE: left, 15 ul aliquots from Superose 6 fractions (14-20); center, 0.2 pg protein from DEAE-5PW fractions (47-50). In both gels the silver stain was overdeveloped to bet- ter show minority bands. A) Autoradiographies: eIF-2 factor (1 pg) was phosphorylated with 0.1 ug protein from DEAE-5PW frac- tions (47-50) as kinase source. The arrows show the Mr x 10-s of marker proteins, eIF-2 factor (K, T, and a) and PKI (a and b) subunits, respectively.
1241

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
TABLE I. Purification
Purification Total protein Total activity Specific activity Steps mc7 units' units/m%
Postmicrosomal Supernatant 7773 l *
Heparin-Sepharose 30.0 186 6.2 Phosphocellulose 6.25 61 9.8 GDP-agarose 3.40 35 10.4 Superose 6 0.25 a.9 35.6 DUE-5PW 0.007 0.69 99.0
1 unit: arbitrary unit estimated by densitometric scanning of autoradiograms.
l activity too low to quantitate.
endogenous eIF-2 factor which eluted in fractions 45-47)
(Fig.3A). From the study of both the electrophoretic and the
phosphorylation patterns of fractions from the Superose 6 and
DEAE-5PW chromatographies (Fig. 1,2 and 3) it can be concluded
that the protein kinase activity is associated with the two
majority bands present in fraction 49. The activity is then
present in a protein complex of two nonidentical polypeptides
called a and b with relative molecular masses of approximately
38,000(a) and 20,000 lb). The relative molecular mass of the
holoenzyme estimated from the Superose 6 column is about 140.000
and the ratio obtained by densitometric gel scanning for the a
and b bands is 2.5-3.0:1, which is compatible with a structure of
aab. The protein kinase activity was undetectable in the
postmicrosomal supernatant and increased progressively during the
successive purification steps (not shown). It was purified ap- proximately 16-fold over the heparin-Sepharose step, the first step in which phosphorylation could be estimated by densitometric quantitation of the autoradiograms (Table I). Work is now in progress to improve the yield of the purification.
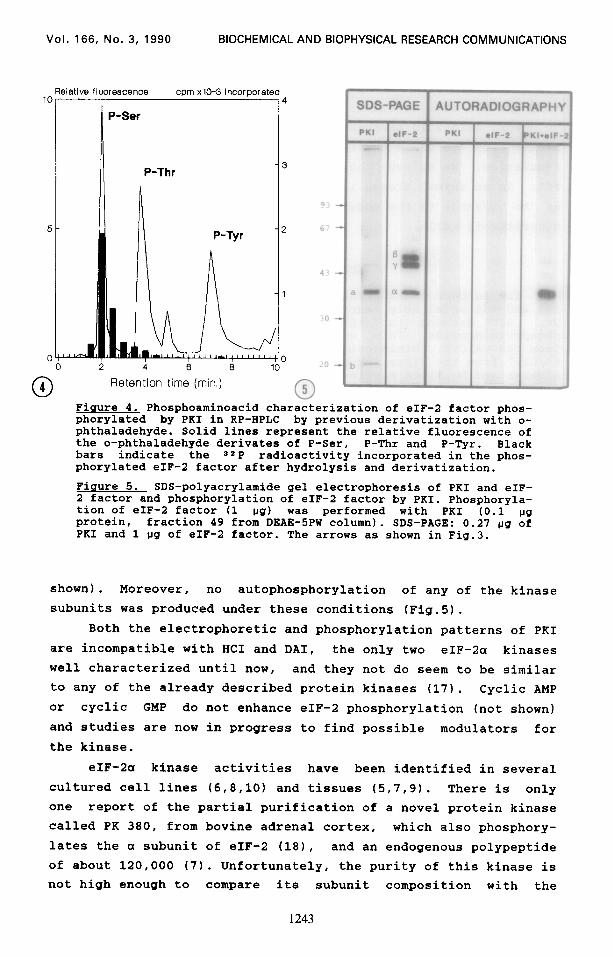
Analysis of the aminoacid residue phosphorylated in the a subunit of eIF-2 indicates that the kinase can be classified as a
serine protein kinase (Fig-l). The phosphorylation studies show that the kinase only phos-
phorylates the a subunit of eIF-2 factor in vitro (Fig.51, thereby indicating that it is not casein kinase II, protein kinase C nor CAMP-dependent protein kinase which phosphorylate the B subunit of the factor "in vitro" (131, and does not phos- phorylate exogenous substrates like casein, or histones (not
1242
Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Aatlve fluorescence cpm x10-3 Incorporated
Oil” 0
3r F ‘-Ser
P-Thr
I&
P-Tyr
l!JL 4 6 8
0 4 Retention time (min.)
4 SDS-PAGE 1 AUTORADlOGRAPHl
93
67
43
1
30
1 20
0 5
Figure 4. Phosphoaminoacid characterization of eIF-2 factor phos- phorylated by PKI in RP-HPLC by previous derivatization with o- phthaladehyde. Solid lines represent the relative fluorescence of the o-phthaladehyde derivates of P-Ser. P-Thr and P-Tyr. Black bars indicate the ==P radioactivity incorporated in the phos- phorylated eIF-2 factor after hydrolysis and derivatization.
Figure 5. SDS-polyacrylamide gel electrophoresis of PKI and eIF- 2 factor and phosphorylation of eIF-2 factor by PKI. Phosphoryla- tion of eIF-2 factor (1 IJS) was performed with PKI (0.1 l.lg protein, fraction 49 from DEAE-5PW column). SDS-PAGE: 0.27 ug of PRI and 1 ug of eIF-2 factor. The arrows as shown in Fig.3.
shown). Moreover, no autophosphorylation of any of the kinase
subunits was produced under these conditions (Fig.5).
Both the electrophoretic and phosphorylation patterns of PKI
are incompatible with HCI and DAI, the only two eIF-2a kinases
well characterized until now, and they not do seem to be similar
to any of the already described protein kinases (17). Cyclic AMP or cyclic GMP do not enhance eIF-2 phosphorylation (not shown)
and studies are now in progress to find possible modulators for
the kinase.
eIF-2a kinase activities have been identified in several
cultured cell lines (6,8,10) and tissues (5,7,9). There is only
one report of the partial purification of a novel protein kinase called PK 380, from bovine adrenal cortex, which also phosphory-
lates the a subunit of eIF-2 (181, and an endogenous polypeptide of about 120,000 (7). Unfortunately, the purity of this kinase is not high enough to compare its subunit composition with the
1243

Vol. 166, No. 3, 1990 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
kinase described in this paper, so it is impossible to assert if
they are the same or different protein kinases.
Work is now in progress to fully characterize the eIF-2a
kinase from the brain and to deepen our knowledge of its possible
regulation. We believe that the discovery of this eIF-2a kinase,
PKI, in an animal tissue is very important, as this will permit
us to advance in the understanding of translation regulation in
nonerythroid cells.
ACKNOWLEDGMENTS
The authors thank Fernando Soriano and C. Warren for techni- cal and editorial help,respectively, Dr. R. Martin de1 Rio and Dr. A. Herranz for their help in the analysis of phosphoaminoacids. This work was supported by grants B23/85, from the CAICYT, Ministerio de Educaci6n y Ciencia and 89/1047, 89/0167 from the FISss, Ministerio de Sanidad y Consumo. Albert0 Alcazar and M. Elena Martin acknowledge felowships from the FISss (86/490 and 89/1047 respectively).
REFERENCES
1. Pain, V.M. (1986) Biochem. J. 235, 625-637. 2. Ochoa, S. (1983) Arch. Biochem. Biophys. 223, 325-349. 3. Trachsel, Ii., Ranu R.S., and London, 1-M. (1978) Biochemistry
75, 3654-3658. 4. Ranu, R.S. (1980) Biochem. Biophys. Res. Commun. 97, 252-262. 5. Proud, C.G., and Pain, V.M. (1982) Febs Letters 143, 55-59. 6. Wong S.T., Mastropaolo, W., and Henshaw E.C. (1982) J. Biol.
Chem. 257, 5231-5238. 7. Kuroda, Y., Merrick, W.C., and Sharma, R.K. (1982) Arch.
Biochem. Biophys 213, 271-275. 8. Towle, C.A., Mankin, H.J., Avruch, J., and Treadwell, B.V.
(1984) Biochem. Biophys, Res. Commun. 121, 134-140. 9. Cal&s, C, Fando, J.L., Fernandez, T., Aldzar, A., and Sali-
nas (1985) Neurosci. Lett. 61, 333-337; 10. Clemens, M.J., Galpine, A., Austin, S.A., Panniers, R., Hen-
shaw, E.C., Duncan, R., Hersey, J.W.B., and Pollard, J.W. (1987) J. Biol. Chem. 262, 767-771.
11. Cal&s C., Fando J.L., Azuara, C., and Salinas M. (1986) Mech. Ageing. Dev. 33, 147-156.
12. Cal&s C., Salinas, M, Alcazar A., and Fando J.L. (1988) Neurosci. Lett. 87, 271-276.
13. Alcdzar A., Mendez, E., Fando, J.L., and Salinas M. (1988) Biochem. Biophys. Res. Commun. 153, 313-320.
14. Hames, B.D. (1986) in Gel electrophoresis of proteins (Hames, B.D., and Rickwood, D., eds.) pp. l-92. IRL Press, Oxford.
15. Bradford, M. (1976) Anal. Biochem. 72, 248-254. 16. Lerma, J., Herranz, A., Herreras, O., Abraira, V., and
Martin de1 Rio, R. (1986) Brain Research 384, 145-155. 17. Blackshear, P.J., Nairn, A.C., and Kuo, J.F. (1988) The FASEB
J. 22, 2956-2969. 18. Kuroda Y.. and Sharma R.K. (1980) Biochem., Biophys. Res.
Commun. 96, 601-610.
1244