purification and of thermostable p-mannanase and …(nh4)2so4and0.25 mto 0 m(nh4)2so4. in the case...
TRANSCRIPT

Vol. 56, No. 11
Purification and Characterization of Thermostable P-Mannanase andoL-Galactosidase from Bacillus stearothermophilus
G. TALBOT AND J. SYGUSCH*
Departement de Biochimie, Faculte de Medecine, Universite de Sherbrooke,Sherbrooke, Quebec, Canada JJH 5N4
Received 18 April 1990/Accepted 17 August 1990
Bacillus stearothermophilus secretes ,-mannanase and oa-galactosidase enzymatic activities capable ofhydrolyzing galactomannan substrates. Expression of the hemicellulase activities in the presence of locust beangum was sequential, with mannanase activity preceding expression of aL-galactosidase activity. The hemicel-lulase activities were purified to homogeneity by a combination of ammonium sulfate fractionation, gelfiltration, hydrophobic interaction chromatography, and ion-exchange and chromatofocusing techniques. Thepurified P-D-mannanase is a dimeric enzyme (162 kilodaltons) composed of subunits having identical molecularweight (73,000). Maximal activity did not vary between pH 5.5 and 7.5. The P-D-mannanase activity exhibitedthermostabiity, retaining nearly full activity after incubation for 24 h at 70°C and pH 6.5. The enzyme
displayed high specificity for galactomannan substrates, with no secondary xylanase or cellulase activitydetected. Hydrolysis of locust bean gum yielded short oligosaccharides compatible with an endo mode ofsubstrate depolymerization. Initial rate velocities of the mannanase activity displayed substrate inhibition andyielded estimates for V.. and Km of 455 60 U/mg and 1.5 0.3 mg/ml, respectively, at 70°C and pH 6.5.The a-galactosidase activity corresponded to a trimeric enzyme (247 kilodaltons) having subunits of identicalmolecular weight (82,000). The a-galactosidase had maximal activity at pH 7 to 7.5 and retained full activityafter 24 h of incubation at 60°C. The enzyme had only limited activity on galactomannan substrates as
compared with hydrolysis of p-nitrophenyl a-D-galactose. Kinetics of p-nitrophenyl a-D-galactose hydrolysisyielded linear reciprocal plots corresponding to Vmax and Km of 195 + 10 U/mg and 0.25 0.02 mM,respectively, at 60°C and pH 7. The characterization of the mannanase activity is consistent with its potentialuse in enzymatic bleaching of softwood pulps.
The extraction of lignin from wood fibers is an essentialstep in bleaching of dissolving pulps. Pulp pretreatmentunder alkaline conditions hydrolyzes hemicelluloses cova-lently bound to lignin and thus facilitates subsequent re-moval of lignin. There is a drawback to alkaline treatment ofwood pulps, however, in that it creates an environmentalpollution problemn (8). The alternate use of hemicellulasesequally facilitates lignin removal in pulp bleaching and yieldsresults comparable to alkaline pretreatment (14). Conse-quently, to substitute within a pulp-bleaching sequence an
enzymatic pretreatment for the ultrahot alkaline extractionstage (14) offers the possibility of significant reduction inenvironmental pollution and thus is of considerable interestto the pulp and paper industry. To be feasible, however,enzymatic bleaching requires that hemicellulase treatmentnot impair pulp quality by attacking cellulose fibers.Softwoods from which the majority of pulps are derived
contain as much as 15 to 20% hemicellulose in the form ofgalactomannan (11). Hemicellulases having substrate speci-ficities for galactomannan constituents would make excellentcandidates for use in enzymatic bleaching of softwood pulps(14). Since pulping is best carried out at elevated tempera-tures, thermophilic hemicellulases could offer significantadvantages over mesophilic hemicellulases in terms of theirhigher intrinsic stability and catalytic efficiencies at suchelevated temperatures.
Bacillus strains are capable of utilizing lignocellulosefractions as carbon sources by secreting hydrolytic enzymeswhich depolymerize the hemicellulose and cellulose compo-
* Corresponding author.
nents (4, 9). This article describes the purification andcharacterization of two thermostable hemicellulases fromBacillus stearothermophilus. In the presence of galactoman-nan, B. stearothermophilus produces both a P-D-mannanaseand an a-D-galactosidase activity. Both enzymes retainactivity for more than 24 h at temperatures in excess of 60°C.The high specific activity at 70°C of the P-D-mannanasemakes the enzyme extremely attractive for use in enzymaticbleaching.
MATERIALS AND METHODSMicroorganism. B. stearothermophilus was obtained from
the American Type Culture Collection (ATCC; catalog as-
cension no. 266).Materials. Locust bean gum, guar gum, mannan from
Saccharomyces cerevisiae, xylan (oat spelts), Sigmacell,carboxymethyl cellulose, melibiose, stachyose, raffinose,p-nitrophenyl ot-D-galactopyranoside, and other p-nitrophe-nyl derivatives were purchased from Sigma Chemical Co.Avicel (microcrystalline cellulose) was obtained from Fluka.Enzyme production. The bacterial culture was first ampli-
fied at 55°C in ATCC 266 liquid medium and then grown on
a defined minimal agar medium (12), using 1% (wt/vol) locustbean gum (Sigma) as carbon source.
Colonies grown on this agar medium were first amplifiedby vigorous shaking at 55°C in 3 ml of ATCC 266 liquidmedium. The overnight cultures were then further amplifiedunder identical conditions in 6 liters of the same mediumuntil growth had reached stationary phase.
After centrifugation at 10,000 x g for 20 min, the bacterialpellet was gently transferred into 3 liters of the definedminimal liquid medium (12) containing 1% (wtlvol) locust
3505
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1990, p. 3505-35100099-2240/90/113505-06$02.00/0Copyright © 1990, American Society for Microbiology
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

3506 TALBOT AND SYGUSCH
bean gum. Maximum hemicellulase activities occurredwithin 48 h and were monitored as described below.
Purification. Mannanase and a-galactosidase activities se-creted into the extracellular medium were harvested bycentrifugation of the culture broth to remove cells and solidresidues. The clarified supernatant was brought to 20%ammonium sulfate saturation and centrifuged at 4°C and40,000 x g for 15 min. All subsequent fractionation stepsinvolving ammonium sulfate were carried out at 4°C. Aftercentrifugation, the pellet was discarded and the supernatantwas brought to 50% (NH4)2SO4 saturation and pelleted againat 40,000 x g for 20 min. The a-galactosidase activity wasrecovered in the pellet and dissolved in 20 ml of 100 mMKH2PO4-150 mM KCl, pH 6.5. The supernatant was thenbrought to a 95% (NH4)2SO4 final saturation and centrifugedat 45,000 x g for 30 min. Mannanase activity was recoveredin the pellet and dissolved in 20 ml of 100 mM KH2PO4-150mM KCl, pH 6.5. Mannanase and oa-galactosidase fractionswere then each subjected to gel filtration, using SephacrylS-200 (Pharmacia) previously equilibrated in 100 mMKH2PO4-150 mM NaCl, pH 6.5.
Active fractions from the Sephacryl S-200 gel filtrationstep were pooled and further purified by ion-exchangechromatography: an anion exchanger, Mono Q HR 5/5(Pharmacia), previously equilibrated in 20 mM MES (mor-pholineethanesulfonic acid), pH 6.5, was utilized to purifymannanase activity, while a cation exchanger, Mono S HR5/5 (Pharmacia), equilibrated with 20 mM Bis-Tris, pH 6.0,was used for a-galactosidase purification. For both ionexchangers, enzyme activity was eluted by using a 0 to 1 MNaCl gradient at a flow rate of 1 ml/min. Fractions havingax-galactosidase activity were pooled and dialyzed against 25mM Bis-Tris, pH 6.3, buffer before being applied to achromatofocusing Mono P HR 5/20 column (Pharmacia)equilibrated with the same buffer. A pH gradient was devel-oped with 10% Polybuffer 74 (Pharmacia) and eluted a-ga-lactosidase activity at pH 5.0. To ensure solubility of a-ga-lactosidase activity, 4% (wt/vol) taurine (Sigma) was addedin buffers used for dialysis and chromatofocusing. Hydro-phobic interaction chromatography was then used to purifyboth mannanase and ot-galactosidase activities further. Man-nanase activity was dialyzed against 0.4 M (NH4)2SO4-50mM KH2PO4, pH 6.5, and absorbed onto a fast protein liquidchromatography phenyl-Superose HR 5/5 column (Pharma-cia). Retention of a-galactosidase activity on the hydropho-bic interaction column required only 0.25 M (NH4)2SO4made up in the same phosphate buffer. Enzymatic activitieswere eluted by using respective gradients of 0.4 to 0 M(NH4)2SO4 and 0.25 M to 0 M (NH4)2SO4. In the case of themannanase purification, activity could be detected in threedistinct fractions. The active fractions for both enzymeswere concentrated with polyethylene glycol 20,000 by plac-ing each enzyme in a dialysis bag which was in contact withthe polymer. Each active fraction was then applied to a gelfiltration column, Superose 12 HR 10/30 (Pharmacia), equil-ibrated with 100 mM KH2PO4-50 mM KCI, pH 6.5. Result-ing enzymatic activities were recovered in a single peak andcan be stored in 50% glycerol at -20°C for at least 6 monthswithout significant loss of activity.
Protein assay. Proteins were measured by using bicin-chonic acid protein assay reagent obtained from Pierce.Enzyme assays. Mannanase activity was estimated by the
3,5-dinitrosalicylic acid method as described previously (1),using locust bean gum as substrate. A 500-,lI assay contain-ing 200 RIu of a 0.5% (wt/vol) substrate suspension, 50 pl of500 mM phosphate buffer (pH 6.5), and the desired dilution
of enzyme was incubated for 5 min at 55°C. The reaction wasstopped by addition of 500 RI of 3,5-dinitrosalicylic acidsolution.
ox-Galactosidase was assayed in a 1-ml total volume bymixing 50 ,I of a 20 mM p-nitrophenyl a-D-galactopyrano-side solution with the appropriate enzyme dilution in a 20mM final concentration of phosphate buffer for 5 min at55°C. The reaction was stopped by addition of 30 RI of 2 NNaOH and incubated in a iced water bath to minimizehydrolysis of the substrate.A unit of mannanase or oa-galactosidase activity was
defined as the amount of enzyme which liberates 1 ,umol ofmannose or p-nitrophenol per min under the given assayconditions.Whenever indicated, error bars signify the standard error
about the mean value, which was calculated from a minimumof three different activity assays. Error bars were not shownwhenever the width of the error bar was less than the symbolsizes in the figures.
Polyacrylamide gel electrophoresis (PAGE). Molecularweight estimates of the purified enzymes were determinedon the basis of migration on 12.5% denaturing polyacryl-amide gels (5). Nondenaturating gel electrophoresis wascarried out with a 10 to 15% polyacrylamide gradient sepa-rating gel and molecular weight markers from Pharmacia.
Kinetic analysis. Kinetic constants and associated stan-dard deviations were evaluated by linear regression ofreciprocal velocity plots.
Product chromatography. Polyacrylamide gel, Bio-Gel P-4<400 mesh (Bio-Rad Laboratories), packed into a 90-cm HR16 gel filtration column (Pharmacia) was utilized for productsizing as described before (13). A constant temperature of55°C was maintained in the column jacket by circulation ofthermostated water. Eluant (water) was pumped through thecolumn at 0.25 ml/min. Differential refractive index wasmonitored with a Beckman Refractive Index Detector model156 connected to a Hewlett-Packard Integrator model 3392A. Samples were prepared by first boiling for 2 min (11) andthen centrifuging the suspension to discard insoluble mate-rial. Mannose, galactose, melibiose, and raffinose, havingdegrees of polymerization of 1, 1, 2, and 3, respectively,were utilized to calibrate retention times on the Bio-Gel P-4gel filtration column.
RESULTS AND DISCUSSION
Determination of hemicellulase activity expressed by B.stearothermophilus grown to stationary phase (10) in thepresence of 1% (wt/vol) locust bean gum indicated sequentialexpression of mannanase and a-galactosidase activities. B.stearothermophilus colonies grown previously on definedminimal agar medium secreted maximal mannanase activitywithin 2 days of inoculation. Mannanase activity was se-creted to approximately 0.9 U/ml of media, which is in thesame range as observed for B. subtilis TX1 (0.5 U/ml) (6).ao-Galactosidase activity secreted for the same period waspresent at levels of 0.02 U/ml of media, which is somewhatless than observed for B. stearothermophilus AT-7 (9).Maximal ot-galactosidase activity occurred only 5 days afterinoculation, suggesting that the ,-D-mannanase activity isfirst required to depolymerize the galactomannan to oli-gosaccharides before a-galactosidase activity is significantlyexpressed.
Purification and homogeneity. The purification steps of themannanase and a-galactosidase activities are summarized inTable 1. The purified cx-galactosidase activity was homoge-
APPL. ENVIRON. MICROBIOL.
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

THERMOSTABLE B. STEAROTHERMOPHILUS ENZYMES
TABLE 1. Purification of hemicellulase activities
Purification steps Total activity Total protein Sp act Yield Purification(U) (mg) (U/mg) (%) (fold)
MannanaseCrude extract 2,890 1,230 2.3 100 150-95% (NH4)2SO4 precipitation 1,620 380 4.3 56 1.9Gel filtration (Sephacryl S-200) 1,055 135 7.8 36 3.4Cation exchange (Mono S) 475 11 43 16 19HIC (phenyl-Superose)aA 55 0.99 55.5 1.9 24B 160 2.6 61.5 5.5 27C 50 0.88 56.8 1.7 25
Gel filtration (Superose 12)A 10.6 0.13 80 0.37 35B 130 1.3 100 4.5 43C 36 0.37 97 1.2 42
a-GalactosidaseCrude extract 52 1,380 0.04 100 120-50% (NH4),SO4 precipitation 51.7 162 0.32 98 8.5Gel filtration (Sephacryl S-200) 47 135 0.35 90 9.4Anion exchange (Mono Q) 47 25 1.9 90 51Chromatofocusing (Mono P) 30 7 4.4 58 117HIC (phenyl-Superose) 10.6 0.08 130 20 3,470Gel filtration (Superose 12) 5.9 0.037 160 12 4,275
a HIC, Hydrophobic interaction chromatography.
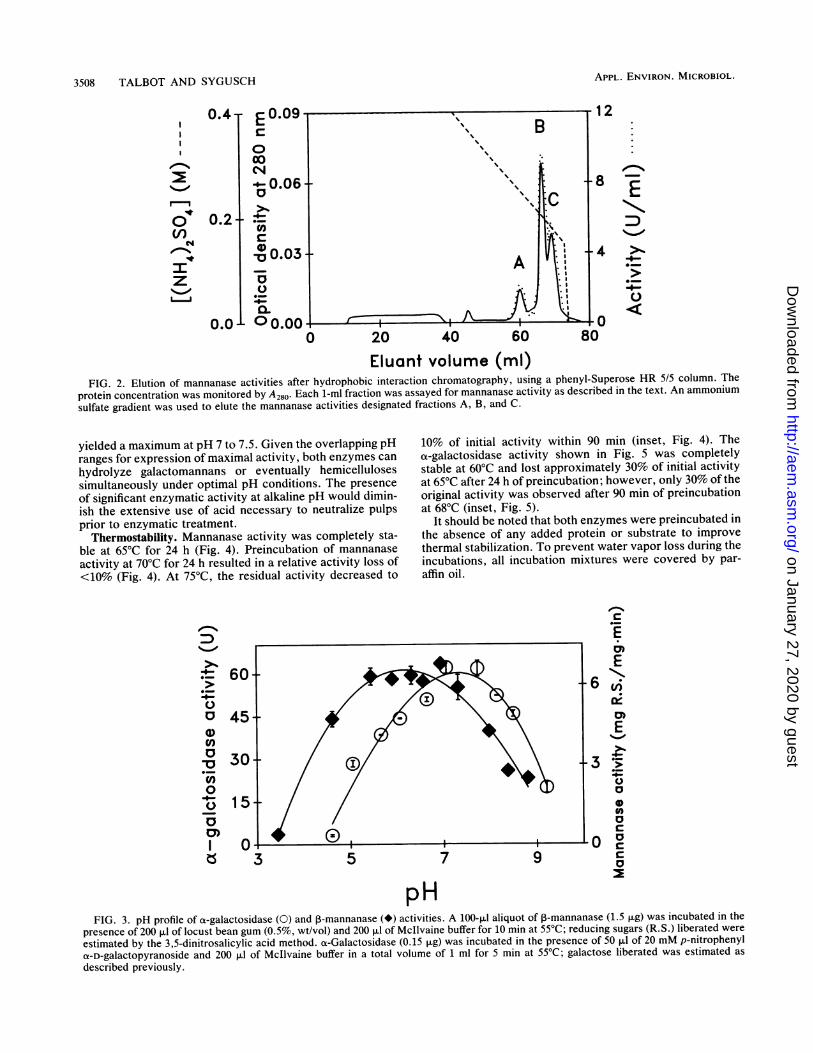
neous on the basis of sodium dodecyl sulfate (SDS)-PAGEand migrated according to a relative molecular weight of82,000 + 5,000 (Fig. 1). It is important to note that hydro-phobic interaction chromatography yielded three active frac-tions denoted A, B, and C (Fig. 2); fraction B containedabout 80% of total mannanase activity, while fractions A and
977.A7,47,7u
42.1 em
31.0-~',
21.5-
14.4- -
FIG. 1. SDS-PAGE (12.5%) of =-mannanase and ax-galactosi-dase. Lane A contains relative molecular weight (10') markers,phosphorylase b (97.4), albumin (66.2), ovalbumin (45), carbonicanhydrase (31), trypsin inhibitor (21.5), and lysozyme (14.4), whichwere purchased from Bio-Rad. Lanes B and C contain purifiedax-galactosidase (2 pLg) and P-mannanase (2 p.g), respectively.
C represented, respectively, approximately 8 and 12% of thetotal mannanase activities. On gel filtration, each of themannanase activities was recovered in a single sharp peaknear the void volume of the Superose 12 gel filtrationcolumn. Migration of the mannanase B activity on SDS-PAGE (Fig. 1) as well as mannanase C activity showed theseactivities to be homogeneous and corresponded to identicalrelative molecular weights of 73,000 ± 6,000. The mannan-ase A activity migrated as two sharp bands on SDS-PAGE(data not shown), one of which migrated identically tomannanase B and C while the migration of the second bandcorresponded to a protein of lower molecular weight, 58,300+ 5,400. The mannanase A and C fractions were notcharacterized further.
Migration of the oa-galactosidase and mannanase B activ-ities on native gradient polyacrylamide (10 to 15%) electro-phoretic gels was consistent with the oa-galactosidase migrat-ing as a trimer of relative molecular weight 247,000 + 13,400,while mannanase B appeared to migrate as a dimer ofrelative molecular weight 162,200 ± 9,500 (data not shown).
Oligomerization of oa-galactosidases has also been ob-served for intracellular a-galactosidases from B. stearother-mophilus AT-7 (9) which have relative molecular weightsestimated from disc gel electrophoresis of 280,000 ± 30,000for a-galactosidase I (SDS-PAGE; molecular weight, 81,000+ 500) and 325,000 + 15,000 for ca-galactosidase II (SDS-PAGE; molecular weight, 84,000 + 500). Molecular weightconsiderations would tend to favor similarity of the purifiedgalactosidase activity with ca-galactosidase I (280,000 +
30,000). However, contrary to a-galactosidases I and II fromB. stearothermophilus AT-7 which is constitutively ex-pressed (9), the present a-galactosidase activity appears tobe induced since in the absence of locust bean gum nosignificant intracellular activity was detected.Optimal pH. The pH profile of mannanase activity showed
a broad plateau corresponding to maximal activity betweenpH 5.5 and 7.5 (Fig. 3). The corresponding pH profile foroa-galactosidase activity was considerably narrower and
VOL. 56, 1990 3507
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.3508 TALBOT AND SYGUSCH
0.4T
.5
0
v)I
zI
1
_0,40.2+
0.010 20 40
4-60
12
E
0-
C-)
80
Eluant volume (ml)FIG. 2. Elution of mannanase activities after hydrophobic interaction chromatography, using a phenyl-Superose HR 5/5 column. The
protein concentration was monitored by A280. Each 1-ml fraction was assayed for mannanase activity as described in the text. An ammoniumsulfate gradient was used to elute the mannanase activities designated fractions A, B, and C.
yielded a maximum at pH 7 to 7.5. Given the overlapping pHranges for expression of maximal activity, both enzymes canhydrolyze galactomannans or eventually hemicellulosessimultaneously under optimal pH conditions. The presenceof significant enzymatic activity at alkaline pH would dimin-ish the extensive use of acid necessary to neutralize pulpsprior to enzymatic treatment.
Thermostability. Mannanase activity was completely sta-ble at 65°C for 24 h (Fig. 4). Preincubation of mannanaseactivity at 70°C for 24 h resulted in a relative activity loss of<10% (Fig. 4). At 75°C, the residual activity decreased to
¢D-I-
-6.-
C-)
0C')0
-oC.)0CY)
10% of initial activity within 90 min (inset, Fig. 4). Theot-galactosidase activity shown in Fig. 5 was completelystable at 60°C and lost approximately 30% of initial activityat 65°C after 24 h of preincubation; however, only 30% of theoriginal activity was observed after 90 min of preincubationat 68°C (inset, Fig. 5).
It should be noted that both enzymes were preincubated inthe absence of any added protein or substrate to improvethermal stabilization. To prevent water vapor loss during theincubations, all incubation mixtures were covered by par-affin oil.
E
6a)
E
3 4-
0
Ca
pHFIG. 3. pH profile of at-galactosidase (0) and ,-mannanase (*) activities. A 100-p.l aliquot of P-mannanase (1.5 ,ug) was incubated in the
presence of 200 ,ul of locust bean gum (0.5%, wt/vol) and 200 p.l of Mcllvaine buffer for 10 min at 55°C; reducing sugars (R.S.) liberated wereestimated by the 3,5-dinitrosalicylic acid method. cx-Galactosidase (0.15 p.g) was incubated in the presence of 50 ,ul of 20 mM p-nitrophenyla-D-galactopyranoside and 200 .l1 of Mcllvaine buffer in a total volume of 1 ml for 5 min at 55°C; galactose liberated was estimated as
described previously.
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

THERMOSTABLE B. STEAROTHERMOPHILUS ENZYMES
e-4->1*
_
0
0*
0aEU)
50 1~~00 q1M'
0 0 0 1
P10Prencbatontim (rs
30 -
0 8 16
Preincubation time (hrs)FIG. 4. Thermal stability of P-D-mannanase. The enzyme (1.5
p.g) was preincubated in 50 mM phosphate buffer, pH 6.5, at 65°C(G), 70°C (*), and 75°C (0) for different periods of time and thenassayed for residual activity at 55°C.
Enzymatic kinetics: Km and Vmax. The kinetics of p-nitro-phenyl ct-D-galactopyranoside hydrolysis by ot-galactosidaseyielded a linear reciprocal velocity plot. A Vmax of 195 + 10U/mg and a Km of 0.25 + 0.02 mM were obtained at 60°C andpH 7.0 for the purified ao-galactosidase. The Km value issimilar to values obtained for ot-galactosidases I (0.47 mM)and 11 (0.53 mM) purified from B. stearothermophilus AT-7(9). Kinetics of locust bean gum depolymeration by mannan-ase was examined at 70°C and pH 6.5. Shown in Fig. 6 is a
reciprocal velocity plot of the kinetic characterization. Inhi-bition by substrate appears to occur at high substrate con-centrations since velocity decreases with increasing sub-strate concentration. Extrapolation of reciprocal velocities
_\_
-4-
-4--00*)
a)
1OOK
501
300 8 16 24
Preincubation time (hrs)FIG. 5. Thermal stability of cx-galactosidase. The enzyme (0.15
p.g) was preincubated in 20 mM phosphate buffer, pH 7.0 at 60°C(O), 65°C (O), and 68°C (0) for different periods of time and thenassayed for residual activity at 55°C as described in the text.
3% 8--
04-
> 2-
-1 0 1 2
1/[LB Gum] (ml/mg)FIG. 6. Reciprocal velocity plot of mannanase activity as a
function of locust bean (LB) gum concentration. Velocities weredetermined at 70°C by incubating 50 pul of enzyme (0.75 p.g), 400 p.lof substrate (different concentrations), and 50 p.1 of 0.5 M KH2PO4,pH 6.5, buffer and stopping the reaction at various times asdescribed previously.
from low concentrations (c5 mg/ml) yielded a Vmax of 455 ±60 U/mg, which is comparable to the specific activity of B.subtilis TX-1, 514 U/mg (6). All other mannanases studied todate have lower Vmax values ranging from 43.5 (6) to 238 (2)U/mg. The estimated Km value (1.5 + 0.3 mg/ml) is compa-rable to those resulting from characterization of other man-nanases (8).
Evaluation of kinetic parameters for both enzyme activi-ties were carried out under conditions of linearity as afunction of both time and enzyme concentration.
Specificity. The a-galactosidase substrate specificity sum-marized in Table 2 shows only very limited hydrolysis ofpolymeric substrates when compared with hydrolysis ofp-nitrophenyl cx-D-galactopyranoside. The low activity byot-galactosidase toward polymeric substrates is not inconsist-ent with the hypothesis that expression of mannanase activ-ity is first required to depolymerize locust bean gum intomore efficient oligomeric substrates for ot-galactosidase. Theapparent preference for short oligomeric substrates by the
TABLE 2. os-Galactosidase and P-D-mannanasesubstrate specificities
Enzyme Substrate Relative activity(%)
a-Galactosidase pNP a-D-galactopyranosidea 100Melibiose 3Stachyose 2.3Guar gum 1.4Locust bean gum 0.21Raffinose 0.19pNP P-D-galactopyranoside 0
P-D-Mannanase Locust bean gum 100Guar gum 15 + 3Mannan from S. cerevisiae 0pNP P-D-mannoside 0Xylan (oat spelts) 0Sigmacell 0Avicel 0Carboxymethyl cellulose 0
" pNP, p-Nitrophenyl.
%-..1001
0
o. 0.1 . .
Pe50C
Eix 25I i
0.0 0.5 1.0 1.5Preincubation time (hrs)
VOL. 56, 1990 3509
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
3510 TALBOT AND SYGUSCH
X 6 -a)
a) 4
0-
0L-
#'4--a)
1 -2_l0-0
175
Retention time (
295
DP 2
DP 1 DP 3
min) DP
A I\AA
415 535
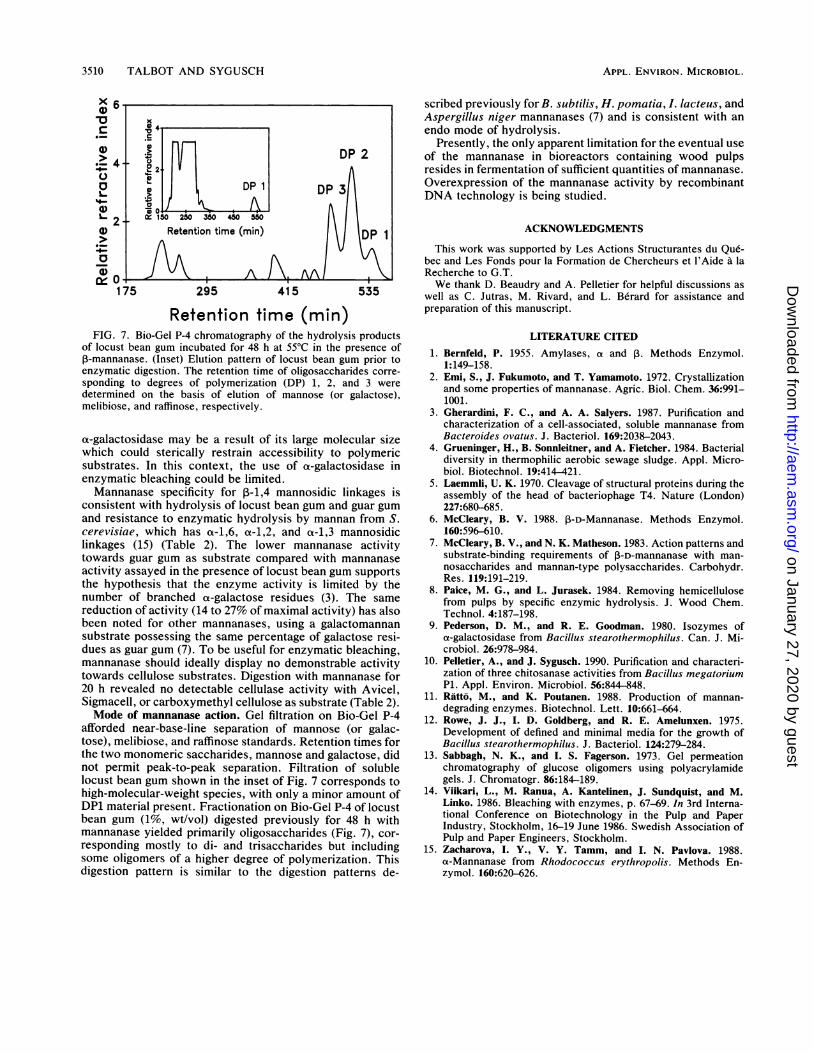
Retention time (min)FIG. 7. Bio-Gel P-4 chromatography of the hydrolysis products
of locust bean gum incubated for 48 h at 55°C in the presence of,-mannanase. (Inset) Elution pattern of locust bean gum prior toenzymatic digestion. The retention time of oligosaccharides corre-sponding to degrees of polymerization (DP) 1, 2, and 3 weredetermined on the basis of elution of mannose (or galactose),melibiose, and raffinose, respectively.
a-galactosidase may be a result of its large molecular sizewhich could sterically restrain accessibility to polymericsubstrates. In this context, the use of a-galactosidase inenzymatic bleaching could be limited.Mannanase specificity for 1-1,4 mannosidic linkages is
consistent with hydrolysis of locust bean gum and guar gumand resistance to enzymatic hydrolysis by mannan from S.cerevisiae, which has ot-1,6, a-1,2, and a-1,3 mannosidiclinkages (15) (Table 2). The lower mannanase activitytowards guar gum as substrate compared with mannanaseactivity assayed in the presence of locust bean gum supportsthe hypothesis that the enzyme activity is limited by thenumber of branched a-galactose residues (3). The samereduction of activity (14 to 27% of maximal activity) has alsobeen noted for other mannanases, using a galactomannansubstrate possessing the same percentage of galactose resi-dues as guar gum (7). To be useful for enzymatic bleaching,mannanase should ideally display no demonstrable activitytowards cellulose substrates. Digestion with mannanase for20 h revealed no detectable cellulase activity with Avicel,Sigmacell, or carboxymethyl cellulose as substrate (Table 2).Mode of mannanase action. Gel filtration on Bio-Gel P-4
afforded near-base-line separation of mannose (or galac-tose), melibiose, and raffinose standards. Retention times forthe two monomeric saccharides, mannose and galactose, didnot permit peak-to-peak separation. Filtration of solublelocust bean gum shown in the inset of Fig. 7 corresponds tohigh-molecular-weight species, with only a minor amount ofDP1 material present. Fractionation on Bio-Gel P-4 of locustbean gum (1%, wt/vol) digested previously for 48 h withmannanase yielded primarily oligosaccharides (Fig. 7), cor-
responding mostly to di- and trisaccharides but includingsome oligomers of a higher degree of polymerization. Thisdigestion pattern is similar to the digestion patterns de-
scribed previously for B. subtilis, H. pomatia, I. lacteus, andAspergillus niger mannanases (7) and is consistent with anendo mode of hydrolysis.
Presently, the only apparent limitation for the eventual useof the mannanase in bioreactors containing wood pulpsresides in fermentation of sufficient quantities of mannanase.Overexpression of the mannanase activity by recombinantDNA technology is being studied.
ACKNOWLEDGMENTS
This work was supported by Les Actions Structurantes du Qud-bec and Les Fonds pour la Formation de Chercheurs et l'Aide A laRecherche to G.T.We thank D. Beaudry and A. Pelletier for helpful discussions as
well as C. Jutras, M. Rivard, and L. Berard for assistance andpreparation of this manuscript.
LITERATURE CITED1. Bernfeld, P. 1955. Amylases, a and ,B. Methods Enzymol.
1:149-158.2. Emi, S., J. Fukumoto, and T. Yamamoto. 1972. Crystallization
and some properties of mannanase. Agric. Biol. Chem. 36:991-1001.
3. Gherardini, F. C., and A. A. Salyers. 1987. Purification andcharacterization of a cell-associated, soluble mannanase fromBacteroides ovatus. J. Bacteriol. 169:2038-2043.
4. Grueninger, H., B. Sonnleitner, and A. Fietcher. 1984. Bacterialdiversity in thermophilic aerobic sewage sludge. Appl. Micro-biol. Biotechnol. 19:414-421.
5. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
6. McCleary, B. V. 1988. P-D-Mannanase. Methods Enzymol.160:596-610.
7. McCleary, B. V., and N. K. Matheson. 1983. Action patterns andsubstrate-binding requirements of ,-D-mannanase with man-nosaccharides and mannan-type polysaccharides. Carbohydr.Res. 119:191-219.
8. Paice, M. G., and L. Jurasek. 1984. Removing hemicellulosefrom pulps by specific enzymic hydrolysis. J. Wood Chem.Technol. 4:187-198.
9. Pederson, D. M., and R. E. Goodman. 1980. Isozymes ofa-galactosidase from Bacillus stearothermophilus. Can. J. Mi-crobiol. 26:978-984.
10. Pelletier, A., and J. Sygusch. 1990. Purification and characteri-zation of three chitosanase activities from Bacillus megatoriumP1. Appl. Environ. Microbiol. 56:844-848.
11. Ratto, M., and K. Poutanen. 1988. Production of mannan-degrading enzymes. Biotechnol. Lett. 10:661-664.
12. Rowe, J. J., I. D. Goldberg, and R. E. Amelunxen. 1975.Development of defined and minimal media for the growth ofBacillus stearothermophilus. J. Bacteriol. 124:279-284.
13. Sabbagh, N. K., and I. S. Fagerson. 1973. Gel permeationchromatography of glucose oligomers using polyacrylamidegels. J. Chromatogr. 86:184-189.
14. Viikari, L., M. Ranua, A. Kantelinen, J. Sundquist, and M.Linko. 1986. Bleaching with enzymes, p. 67-69. In 3rd Interna-tional Conference on Biotechnology in the Pulp and PaperIndustry, Stockholm, 16-19 June 1986. Swedish Association ofPulp and Paper Engineers, Stockholm.
15. Zacharova, I. Y., V. Y. Tamm, and I. N. Pavlova. 1988.a-Mannanase from Rhodococcus erythropolis. Methods En-zymol. 160:620-626.
APPL. ENVIRON. MICROBIOL.
on January 27, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from