purification and characterization of a-amylase from
TRANSCRIPT

PURIFICATION AND CHARACTERIZATION OF a-AMYLASE
FROM BACTEROIDES AMYLOPHILUS STRAIN H-18
by
SHEIKH SAIF - UR - RAHMAN
B.Sc. (A.H.)-, The University of the Panjab, 1960 M.S.A., The University of British Columbia, 1965
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF
THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
in the Department
of
Animal Science
We accept this thesis as conforming to the
required standard
THE UNIVERSITY OF BRITISH COLUMBIA
September, 1970

In p r e s e n t i n g t h i s t h e s i s in p a r t i a l f u l f i l m e n t o f the r equ i r emen t s f o r
an advanced degree a t the U n i v e r s i t y o f B r i t i s h Co lumb i a , I ag ree tha t
the L i b r a r y s h a l l make i t f r e e l y a v a i l a b l e f o r r e f e r e n c e and S tudy .
I f u r t h e r ag ree tha t p e r m i s s i o n f o r e x t e n s i v e c o p y i n g o f t h i s t h e s i s
f o r s c h o l a r l y pu rposes may be g r an t ed by the Head o f my Department or
by h i s r e p r e s e n t a t i v e s . It i s u n d e r s t o o d t h a t c o p y i n g or p u b l i c a t i o n
o f t h i s t h e s i s f o r f i n a n c i a l g a i n s h a l l not be a l l o w e d w i t h o u t my
w r i t t e n p e r m i s s i o n .
Department
The U n i v e r s i t y o f B r i t i s h Co lumbia Vancouver 8, Canada
Date 5se^V,U-

Chairman, Professor W. D. Kitts
ABSTRACT
The research was undertaken to study the extracellular
a-amylase produced by the anaerobic rumen bacterium, Bacteroides
amylophilus strain H-18. Four active isoenzymes of a-amylase
were detected by disc electrophoresis and electrofocusing tech
niques. Isoelectric points as determined by electrofocusing
were pH 3.7, 4.5,,5.9 and 8.0. Isoenzymes were named 1, 2, 3
and 4 with respect to their increasing isoelectric points.
a-Amylase isoenzyme 1 was purified by DEAE-Sephadex and
G-200 techniques. Some of i t s general physio-chemical proper
ties were studied. It had maximum activity at pH 6.7, 44°C and
was stabilized by calcium ion. It was susceptible to thermal
denaturation in the absence of calcium. Various other metal ions
tested could not replace the calcium in regenerating maximum
activity. It was found by atomic absorption spectrophotometry
that a-amylase isoenzyme 1 contained 3 gram-atoms of calcium per
mole of enzyme. The estimated molecular weight by gel f i l t r a
tion technique was 45,000 Daltons. Amino acid analysis indica
ted the absence of cysteine, therefore, disulphide linkages were
not involved in maintaining the. tertiary structure. Tryptophan
appeared to be required for enzymic activity, as determined by
the N-bromosuccinamide oxidation technique.
The mode of action of a-amylase isoenzyme 1 was studied

using amylose and soluble starch as substrates. The products of
enzymatic degradation were analysed qualitatively by thin layer
chromatography. The•maltohexaose, maltoheptaose, maltoctaose,
maltonanaose and maltodecaose remained in the digest mixture for
sometime after the achroic point. The degree of multiple attack
was 2 , as calculated by determining the ratio of the reducing value
of the oligosaccharide fraction to that of polysaccharide fraction.
Antisera against a-amylase isoenzyme 1, produced in rabbits
by injection of a-amylase inFreund's complete adjuvant was found
to be mono-specific. The inhibition of a-amylase activity by
antibody and inhibitory effect of starch on the amylase-anti-
amylase system were'demonstrated. The effect of anti-amylase
(isoenzyme 1) globulin on amylases of diverse origin was studied
by the Ouchterlony double diffusion technique. These experiments
demonstrated antigenic determinants which were distinct from those
present on the a-amylase of hog pancreas, Bacillus subtilis and
Aspergillus oryzae. Immunoelectrophoretic analysis indicated the
presence of only a single antigenic component. Quantitative pre
cipitation studies gave a typical curve with one equivalence
point with an antibody to antigen ratio of 2.'31. N-Bromosuccin-
imide treated a-amylase (isoenzyme 1) exhibited similar immuno
chemical behaviour to the native enzyme, but completely lost i t s
catalytic activity. It is possible that catalytic and antigenic
sites were distinct. Urea treated a-amylase (isoenzyme 1) did
not show any precipitate with i t s specific antibody and thus,.ap
peared to have lost i t s antigenic structure.

Dedicated to my parents, teachers and friends
who helped, each in their own way.
i

TABLE OF CONTENTS
Chapter Page
I. INTRODUCTION . . . . . . . . . . . 1.
II. REVIEW OF THE LITERATURE . . . . . . 2
Amylase . . . . - 2
Nomenclature . . • . • 2
a-Amylases and their Sources . . . 3
Non-ruminant a-Amylase . . . • 3
Ruminant a-Amylase . . . . 4
Production and Induction of a-Amylase . . . . . . 4
Physical Properties of Crystalline a-Amylase 6
Primary Structure of a-Amylases . 6
Amino Acid Analysis 6
Functional Groups 6
Non-protein Constituents and Co-factors
in a-Amylase . . . . 9
a-Amylase as Metalloenzyme . . . . . . . . . 9
Secondary and Tertiary Structure of a-Amylase . . . . . 11
Quarternary Structure of a-Amylase . . . . . . . . 13
The Action Pattern of a-Amylase . . . . . . . . . 1 5
Immunochemical Study of a-Amylase • . • . . . 18
Mammalian a-Amylase 18
Microbial a-Amylase . . . . 18

v i
Chapter Page
III. MATERIALS AND METHODS . . . . . . . . . . . . . . 31
Chemicals • . . . . 31
Organisms . . 3 1
Maintenance of Bacteroides amylophilus Strain H-18 . . . . • . . . : . . . . . . . . 32
Growth Measurements of Bacteroides amylophilus Strain H-18 . . . . 3 2
Production and Purification of a-Amylase
from Bacteroides amylophilus Strain H-18 33
Production of a-Amylase . . . . . . . . • . . 33
Purification of a-Amylase on DEAE-Sephadex
A-50 and G-200 Sephadex • 33
Assay of a-Amylase • . . 34
Assay of Protease 35
Determination of Nitrogen . • 35
Determination of Protein . . . 35
Determination of Total Carbohydrate 36
Disc Gel Electrophoresis 36
Isoelectrofocusing . . . . . . . . . . . . . . 37
Charcoal-Celite Column Chromatography . . . • . • . . . 37
Paper Chromatography . . . . . . . . . . . . . 38
Thin Layer Chromatography . . . . . 39
Effect of Temperature • . ... . . . . . 3 9
Molecular Weight Determination . . . . 3 9
Calcium Content Determination . . . . . . . . . . 39 •

v i i
Chapter Page
Amino Acid Analysis . . • 40
Immunochemical Techniques . . 40 .
Production of Antibodies 40
Determination of Enzymic Inhibition • . 41 -
Immunodiffusion Characteristics 41 •
Protein. Determination in Antigen-Antibody Complex . . . • . . . 4 1
IV. RESULTS AND DISCUSSION . . . . . . . . . . . . . 44
Characterization of a-Amylase from Bacteroides
amylophilus Strain H-18 . . ' . . • . . . . . . . 4 4
Production and Purification of a-Amylase 44
Production of a-Amylase . . . 44
Purification of a-Amylase Isoenzyme 1 54
Catalytic Properties of a-Amylase
Isoenzyme 1 . . . . . . . . . . . . . . 66
Determination of Type of Amylase . . . . . . . 66
Effect of pH on a-Amylase Activity 66
Effect of pH on Enzymic Stability 66
Effect of Temperature on a-Amylase Activity . . . . • . . . . - . . . . • . . 75
Effect of Temperature on Enzymic
Stability . . . . . . . . . . . . . . 75
Amino Acid Determination . . . . 8 1
Calcium Determination . . . • . . . . . . . . 8 1

v i i i
Chapter Page .
Effect of Chemical Reagents on
Enzymic Activity . . . . . . : . . 8 3
Effect of Urea on a-Amylase Activity 83
Effect of EDTA and Metallic Ions
on a-Amylase Activity . 87
Functional Groups Determination . . . . . . . 9 0
Determination of Molecular Weight 93
Determination of Isoelectric Point . . ... . . 93
The Action Patternrof a-Amylase Isoenzyme 1 . . . - . . 101
Immunochemical Studies on a-Amylase Isoenzyme 1 113 Inhibition of Enzymic Activity by
Antibodies . 113 .
Ouchterlony Double Diffusion Analysis 116
Immunoelectrophoretic Analysis - . 116
Quantitative Precipitation Analysis 121
Effect of N-bromosuccinimide and Urea on Antigenicity 121
The Neutralization of Amylase-Antiamylase System by Starch . . . . . . . . - . . . ... 126
V. CONCLUSIONS . . . . . . . . . . . . . . . . .135

ix
Chapter Page
General Properties . . • . . . . . . . . . . . . 135
Action Pattern . . . . . . . . 136
Immunochemical Properties . . . . 136
LIST OF REFERENCES : . . . . . . . .
Chapter I and Chapter II 22
Chapter III . 42
Chapter IV:A . . . 96
B • . . . . . . .111
C 133

LIST OF TABLES
Table Page
I. ' The Sources of a-Amylase . 5
II. Physical Properties of a-Amylase .• . . . 7
III. Purification of B_. amylophilus Strain H-18 a-Amylase Isoenzyme 1 58
IV. Summary of the Optimum pH Range of a-Amylase from Various Sources 71
V. Summary of the Optimum pH Stability Range for
Various a-Amylases . . . . . . • 74
VI. Optimum Temperature for Various a-Amylases . . . . . . 78
VII. Calcium Contents of Various a-Amylases . . . . . . . . 82
VIII. Effect of Reducing, Oxidizing and SH-Inactivating
Agents on a-Amylase Isoenzyme 1 Activity . . . . ; . . 91
IX. Molecular Weight of Various a-Amylases 94
X. Isoelectric Points of Various a-Amylases 95
XI. Estimation of the Degree of Multiple Attack by B̂. amylophilus a-Amylase Isoenzyme 1 . . • 110
XII. Precipitation reaction of B_. amylophilus a-Amylase Isoenzyme 1 with i t s Antibody . . . . . . . . . . 122
XIII. Effect of N-bromosuccinimide on Antigenicity of a-Amylase Isoenzyme 1 . . . . . . . . . . . . . . 125
XIV. Effect of Urea on Antigenicity of a-Amylase. Isoenzyme 1 . . . . . . . . . . . . . . . . 129

LIST OF FIGURES
Figure Page
1. Growth curve and production of a-amylase from Bacteroides amylophilus strain H-18 . . . • . . . . 4 5
2. Linear relationship between the production of a-amylase and growth of Bacteroides amylophilus strain H-18 . . . . . . . . - 47
3. Effect of maltodextrin on the growth and production of a-amylase.from Bacteroides amylophilus strain H-18 . . . . . . . . . . . . . 49
4. Detection of 4 isoenzymes of a-amylase by disc electrophoresis . . . : . . . . : . . . . . . 52
5. Detection of 4 isoenzymes of a-amylase by electrof ocusing . 55
6. Flow sheet of methods for isolation of a-amylase isoenzyme 1 from Bacteroides amylophilus strain H-18 57
7. Chromatography.of Bacteroides amylophilus strain H-18 a-amylase on DEAE-Sephadex A-50 ,. . . . . . . . 60
8. Chromatography of Bacteroides amylophilus strain H-18 a-amylase isoenzyme 1 on Sephadex G-200 . . . . . . . . . . . . . . . . 62
9. Disc electrophoresis of a-amylase isoenzyme 1 64
10. Electrof ocusing of a-amylase isoenzyme 1 . 67.
11. Optimum pH for hydrolyzing starch . . . . . . . . . . 6 9
12. Effect of pH on the s t a b i l i t y of a-amylase . - . . . . . . 72
13. Optimum temperature for hydrolysing starch . . . . . . . 76

x i i
Figure Page
14. Thermal st a b i l i t y of a-amylase 79
15. Effect of urea on a-amylase activity . . . . . . . . . . 84
16. Effect of (A) EDTA and (B) metal ions after EDTA treatment.on reactivation of a-amylase 88
17. Thin layer analysis of the digestion of amylose by a-amylase isoenzyme 1.. . . . . . . . . . 102
18. Thin layer analysis of the digestion of starch by a-amylase isoenzyme 1 104
19. Neutralisation curve of a-amylase isoenzyme 1 with antiserum . . . 114
20. A diagramatic representation of immunodiffusion precipitation reaction between B_. amylophilus a-amylase isoenzyme 1 . . . . ... . 117
21. A diagramatic representation of Immunoelectrophoresis of j$. amylophilus a-amylase • isoenzyme 1 . . . . . . . . 119
22. Precipitation curve of a-amylase isoenzyme 1 with i t s antibody . . ... . . • . . 123
23. Precipitation curve of N-bromosuccinimide treated and native a-amylase with i t s antibody 127
24. Inhibitory effect of starch on a-amylase isoenzyme 1 and antiamylase system . , . . 131

ACKNOWLEDGEMENTS
The author wishes to express his sincere thanks to Professor
W. D. Kitts, Chairman, Department of Animal Science, for his supervis
ion, indispensable guidance and helpful criticism in the completion
of this study.
The author would also like to express his sincere gratitude and
appreciation for the valuable suggestions and other f a c i l i t i e s provided
by Dr. T. H. Blackburn, Associate Professor, Department of Micro
biology, and to Dr. S. Nakai, Associate Professor, Department of Food
Science.
The author wishes to express his thanks to graduate students,
Mr. L. E. Lesk, Department of Microbiology, Mr. R. J. Hudson, and Mr.
J. A. Shelford, Department of Animal. Science for their help in enzyme
preparation, immunological studies and amino acid analysis.
Thanks are due to a l l those in Canada and in Pakistan whose help
and encouragement were of immense importance during the course of this
study.
Appreciation is also expressed to Miss V. Curylo and Mrs. J. A.
Shelford for typing the manuscript.
Acknowledged with thanks, is the National Research Council of
Canada postgraduate scholarship and the University of British Columbia
Fellowship.

PURIFICATION AND CHARACTERIZATION OF a-AMYLASE
FROM BACTEROIDES.AMYLOPHILUS STRAIN H-18

CHAPTER I
INTRODUCTION
Much of the work on rumen bacteria has been devoted to the study
of•cellulolytic organisms and the mechanism by which cellulose is de
graded. With the recent emphasis on higher grain feeding to ruminants
i t has been important to study the breakdown.of starch in the rumen.
Though several species of amylolytic bacteria have been isolated from
the rumen and their incidence studied under a variety of dietary treat
ments (35), the amylolytic enzymes of these organisms have not been
studied in detail. It is hoped that greater knowledge of the production
and mode of action of a-amylase by rumen bacteria may•facilitate better
understanding of starch hydrolysis in the rumen. This is particularly
significant when i t has been.shown that non-amylolytic strains of Buty-
r i v i b r i o , Selenomonas and Eubacterium, which can ferment dextrin but not
starch, are present in the rumen in greater proportions than starch
digesters (35).
Bacteriodes amylophilus strain H-18 is a predominant starch d i
gester constituting 10 per cent of the total rumen bacterial flora (35)
and secreting an active a-amylase in specific laboratory growth madium
(6). Since l i t t l e is known about the a-amylase produced by B_. amylophilus,
this organism was selected for the present investigation. It may be men
tioned here that with the exception of members of genera Pseudombnas and
Vibrio, most organisms producing exoenzymes are Gram positive (78). 13.
amylophilus,,on the other hand, deviates from this general rule in being
Gram negative.

CHAPTER II
REVIEW OF LITERATURE
A. Amylases
Starch is an important source of dietary carbon and therefore i t
is not surprising to find amylases widely distributed in a l l Phyla.
Amylase causes the hydrolysis of amylose, amylopectin, glycogen and
their degraded products. In mammals the digestion of starch is initiated
by the action of salivary amylase and continued in the duodenum by the
action of amylase secreted by the pancreas and.the intestine.
Microbial amylases are extra-cellular, in nature and several micro
organisms continue to produce extra-cellular amylase even after the fer
mentation of starch is completed. It might be expected that the amylase
produced during the growth period of the organism hydrolyzes starch; and
sugar thus produced is utilized for the growth of micro-organisms. The
species of genus Bacillus (82) appear, however, to deviate from this
general rule, viz. , IS. stearothermophilus (112) and B̂. subtilis (82)
which produce extra-cellular amylase even during the stationary phase.
B. Nomenclature
Amylases were classified as a and 3 types by Khun (43) and
Ohlsson (70). The a and 8 amylases yield products which have a and $
configuration at C^ of the reducing sugar respectively. Freeman and.
Hopkin (26) have confirmed the configuration of the anomeric reducing
carbon atom released during the enzymatic hydrolysis of starch.

3
The second important difference between a and g amylases is their
mode of attack on the substrate. a-Amylases, being endoenzymes, cleave
1—>-4 bonds located in the inner region of the substrate. Therefore, a-
amylases are expected to liberate products of varying chain lengths and
also rapidly decrease the viscosity and iodine staining capacity of
starch during enzymatic hydrolysis.
B-Amylases have been regarded as exo-amylases because they do not
rapidly decrease viscosity, and iodine staining of starch during starch
hydrolysis. Since 8-amylase is an exo-enzyme the penultimate bond at a
non-reducing chain end is the only bond available for enzymatic hydrolysis.
g-Amylase attacks in an exclusive manner and produces 8-maltose only.
Although the enzyme of 13. macerans produces cyclic schardinger dextrins
from starch, i t is s t i l l classified as an amylase (27). The enzyme from
13. macerans, like other a-amylases, renders starch achroic to iodine.
Robyt and French (80) reported that the enzyme of B_. polymyxa produces
mainly 8-maltose, but has the a b i l i t y to by-pass the 1—>-6 branch linkage
of glycogen.and amylopectin, thus indicating an a-amylase action pattern.
Amylases also differ in their action pattern on iodine-staining
polysaccharides. It is represented graphically by plotting the change
in blue value against the corresponding changes in reducing value during
starch or amylose hydrolysis, and various amylases follow their own
characteristic curves (44).
C. a-Amylases and their Sources
1. Non-ruminant a-Amylase
During the last twenty years a-amylases have been isolated,

4
purified and crystallized from a variety of sources. (Table I)
2, Ruminant a-Amylase •
Rumen micro-organisms capable of hydrolyzing starch include Strep
tococcus bovis, Bacteroides amylophilus, Bacteroides ruminicola, Siiccin-
imonas amylolytica and Selenomonas ruminantum (35).
A number of rumen c e l l u l o l y t i c micro-organisms also possess
amylolytic properties such as Clostridium lachheadii, some strains of
Bacteroides succinogenes and most strains of Butyrivibrio fibrisolvens
( 3 5 ) .
The amylolytic enzymes of rumen bacteria have not been character
ized except for those of Streptococcus bovis ( 1 1 0 ) and Clostridium
butyricum (32).
3. Production and Induction of a-Amylase
The production characteristics of a-amylase of B̂. subtilis and 13.
stearothermophilus have been studied by many workers which have often
seemed conflicting. J3. subtilis strain N produces extra-cellular a-
amylase predominantly after maximum c e l l growth has occurred (66). The
a-amylase of another strain of 13. subtilis ( 1 4 ) and of B_. stearothermo
philus ( 1 1 1 ) are formed during the logarithmic phase of growth parallel
ing the increase in c e l l mass. Yoshida and Tobita (115.) reported that
a-amylase is released into the medium during the stationary phase of
growth in a leucine requiring mutant of B_.' s u b t i l i s .
. Pseudomonas saccharophila produces inducible extra-cellular a-
amylase ( 4 9 ) . Markovitz and Klein ( 4 9 , 5 0 ) , Schiff et a l . ( 8 4 ) and

5
TABLE I
THE SOURCES OF a-AMYLASES
Source r- Reference
A. Mammalian
1. Human.saliva 25 2. Porcine pancreas. 8, 57 3. Rat pancreas 30 4. Human pancreas 20
B. Plant
1. Barley malt 22, 87 2. Sorghum malt 16
C. Bacterial
1. Bacillus subtilis 96 2. Bacillus stearothermophilus 10, 11 3. Bacillus macerans 88, 79 4. Bacillus polymyxa 80, 83 5. Pseudomonas saccharophila 51
D. Fungus
1. Aspergillus oryzae 21, 104 2. Aspergillus niger
102 j 3. Aspergillus candidus 82

6
Eisenstadt and Klein.(18,19) have presented evidence for the de novo syn
thesis and inducibility of a-amylase in P_. saccharophila. The kinetics
of enzyme formation was reported to be linear and the quantity of a-
amylase produced was proportional to the substrate concentration.
Welker and Campbell.(112) also studied the induction of a-amylase
of 15. stearothermophilus by maltodextrins. They observed that addition
of maltose,' maltotriose, maltotetraose ,'~maltopentaose and .maltohexaose to
a chemically defined medium resulted in a stimulation of the differential
rate of a-amylase production.
D. Physical Properties of Crystalline a-Amylase
Some of the general physical properties of a-amylase are summarized
in Table II.
E.- Primary Structure of the a-Amylases
1. Amino Acid.Analysis
Amino acid analyses have been reported for human salivary amylase
(62), porcine pancreatic amylase (9), 13. subtilis amylase (3,42), 13.
stearothermophilus amylase (12), and A. oryzae amylase (1,94).
The a-amylases of J3. subtilis do not contain cysteine and cystine.
_B. stearothermophilus a-amylase is unusual due to the absence of trypto
phan.
The amino acid sequence of a-amylases has not .been reported.
2. Functional Groups
In order to study the functional groups of a-amylases at least

7
TABLE II
PHYSICAL PROPERTIES OF a-AMYLASES
Properties Source
B.subt i l i s
(54,56, 96)
B.stearothermophilus (11,12)
P.Sacch-aro-phila (51)
A.ory-zae
(21, 104)
Barley malt
(61,87)
Porcine pancreas (8,57, 61)
Human saliva
(25,56, 58,59, 61)
Per cent nitrogen 16(24) 15 — 14.9(24) 13.0 15.9(24) 17.0
3ptimum pH 6.0 5.0
5.25-5.75
4.8-5.8 (24)
4.0-5.8 (4) 6.8 6.9
Optimum pH stab i l i t y range 4.8-8.5 4.5-8 5.5-8.5 4.9-9.1 7.0-8.5 4.8-11
Optimum temperature 40° 65° 40° 35° 37° 40°
Molecu^-lar weight
48,700 (23)
15,000 (47) _
51,000 (38)
59,500 45,000 (15) _
Isoelect r i c point 5.4 4.8 4.2 5.7 5.2-5.6 5.2-5.6
Absorbance % A 280 mu
25.3 (24)
19.7 (24) 26(24) 26(24)
Activation energy
(0-12°) 15,000 12° 11,000
14,000 (0-15°) 14,400 (15-40°) 8,500
10,650 7,050 13,500 13,500

8
two techniques have been,used; (a) the e f f e c t of pH on the'Michaelis
constant, Km, and the maximum v e l o c i t y , Vm and (b) chemical modification
of a-amylases.• E a r l i e r work using chemical modification of an enzyme
has given c o n f l i c t i n g reports regarding the p a r t i c i p a t i n g of functional
groups. At le a s t part of the reason f o r t h i s discrepancy i s the fac t
that the chemical reagent used had l i t t l e s e l e c t i v i t y and i s related
with many side chain groups.
Ono et a l . (73) investigated the e f f e c t of pH on the Km of EL
s u b t i l i s a-amylase.. Their r e s u l t s indicated that the apparent.rate con
stant, K^, of th i s enzyme diminished on both.the a l k a l i n e and acid side
of the optimum pH. This was ascribed to the formation of an anion and
a cation which were determined to PK value of 4.2 and 7.5. These PK
values along with the heat of i o n i z a t i o n indicated that the active groups
involved i n the cleavage of the bonds were a carboxylate ion and an imi-
dazolium ion. The apparent Michaelis constant (Km) was stable over the
pH range of 3.6 to 9.4, suggesting that the side chain.groups responsible
for the substrate binding must ion i z e outside the pH range studied. The
p o s s i b i l i t y that t y r o s y l groups may be involved was indicated because
the PK value of the phenolic hydroxyl group does not f a l l i n the range
studied.
Thoma et a l . (100) reported that the c a t a l y t i c groups of porcine
pancreatic a-amylase were l i k e l y to be,carboxylate and imidazolium ions.
The binding s i t e groups of t h i s a-amylase were d i f f e r e n t from those of 13.
s u b t i l i s shown by the fact that the Km changed with pH, i n d i c a t i n g that
at l e a s t two groups with PK values of 5.7 and 8.7 were responsible for
substrate binding.

9
L i t t l e and:Caldwell (46) inactivated porcine pancreatic a-amylase
by treating with ketene, phenylisocyanate, formaldehyde and nitrous acid,
and suggested that free amino groups were required for catalytic activity.
They also reported that p-chloromercuribenzoate, iodoacetamide and mer
curic chloride did not deactivate the a-amylase. Other work showed that
sulphydryl groups were not required for enzymic activity (7).
Ikenaka (36) treated A. oryzae a-amylase with dinitrobenzene sul-
phonate and fluoronitrobenzene and concluded from his results that the
phenolic group of tyrosine was necessary for enzymic activity. Ikenaka
(37) also reacted A. oryzae a-amylase with p-phenylazobenzoyl chloride
and suggested that e amino groups were required for enzymic action.
3. Non-protein Constituents and Co-factors in a-Amylases
The small quantity of carbohydrates present in A. oryzae a-amylase
are apparently not involved in the enzymic activity (2).
4. a-Amylase as a Metalloenzyme
a-Amylases so far investigated contain at least one atom of cal
cium per mole (105) which is apparently required for enzymic activity
(24,105). Since no other metals could be detected in significant amount,
except zinc in jB. subtilis a-amylase, i t has been suggested that a l l a -
amylases have certain sites to which calcium is attached specifically
(105). 13. subtilis a-amylase is quite unique because of the presence of
four atoms of calcium per mole of protein. It has been suggested that
the increased amount of calcium is required to maintain structural r i g i d
ity because the S-S linkage i s absent in 13. subtilis a-amylase.

10
Yamamoto and Fukumoto (114) reported partial regeneration of ca l
cium depleted subtilis a-amylase by the treatment of strontium, mag-,
nesium, barium and beryllium ions. Hsui et a l , (34) have suggested that
the reagent used by Yamamoto and Fukumoto (114) was not spectroscopically
pure, therefore reaction might be due to the contamination of calcium in
the reagent.
Calcium can be removed from a-amylases by dialysis against sodium
ethylenediamine tetra-acetate, by ammonium sulphate or by electrodialysis
(97). The treatment of enzyme by phosphate, oxalate and citrate failed
to lower the calcium content below 1 gram-atom per mole of enzyme (105).
Under appropriate condition of temperature, pH and ionic strength, the
removal of calcium i t s e l f did not cause an irreversible denaturation of
the enzyme (9,93,105). The calcium free a-amylases were highly suscep
tible to denaturation by heat, urea and acid (93) and also were attacked
easily by proteolytic enzymes (93).
Stein et a l . (97) and Fisher and Stein (24) have reported that
enzymic activity can be regenerated by the addition of calcium to cal
cium free a-amylases. However, enzymic activity could not be revived in
A. oryzae ct-amylase (97). This was thought to be due to the low isoionic
point of the enzyme (pH 4.2) as compared to other a-amylases (pH 5.2 to
5.4). In calcium free .B. subtilis and hog pancreas a-amylases i n s t a b i l
ity increased as pH increased (23). Fisher et a l . (23) reported that no
major structural changes occurred in calcium depleted a-amylase.
The exact role of calcium in the catalytic activity of a-amylases
is not known, but i t is indicated that calcium ions function in a number

11
of ways; (a) i t keeps the a-amylase molecule in compact and proper confor
mation for biological activity by forming a tight intramolecular metal
chelate structure, and (b) i t protects the native enzyme against extreme
pH, heat, and proteolytic enzymes (23,93,105).
Myrback (64) reported that chloride ions activated the pancreatic
and salivary a-amylases, whereas the data of Thoma et a l . (100) indicated
that chloride ion is not essential for porcine pancreatic,a-amylase. The
results of Muss (63) showed that 1-10 mM chloride gave maximum enzymic
activity for salivary a-amylase and protected i t against the detrimental
effect of high temperature and heavy metals. The optimum sodium chloride
concentration for porcine pancreatic a-amylase was 10 mM, and higher
concentrations than 10 mM would cause inhibition of enzymic activity.
Walker and Whelan (108) reported similar relationships between the a c t i
vity of human salivary a-amylase and chloride ion.
F. Secondary and Tertiary Structure of a-Amylase
The-secondary structure of a protein is believed to be due to the
folding of the polypeptide chains into a specific coiled structure. The
interrelationship and arrangement of the folded polypeptide chains into
specific layers of crystals are called tertiary structures of the protein.
It is understood that disulfide bonds, hydrogen bonds and hydrophobic
bonds maintain the secondary and tertiary structures of proteins.
Since IS. subtilis a-amylase does not have disulphide linkages' i t : is
expected to have different secondary and tertiary structures and behave
differently towards denaturing agents. Isemura and Imanishi (40) have

12
studied carefully the conformational changes in 13. subtilis a-amylase in
alkaline and urea solution. Their finding was that approximately 30 per
cent of a l l the phenolic hydroxyl groups ionize freely in alkaline pH
up to 11.5. The remaining groups appear to ionize irreversibly at apH
of 11.5 and therefore are likely to be buried in protein molecule. Also
at a high alkaline pH, the tertiary structure appears to be disrupted
irreversibly due to the dissociation of hydrogen bonds between carbox
ylate groups and phenolic hydroxyl groups. However, the enzymic•
activity was regenerated by dialysis after the disruption of hydrogen
bonds using 8 M urea.
Manning et a l . (47) reported large negative optical rotation on
15. stearothermophilus a-amylase and this was not significantly affected
by 8.0 M urea, 4.0 Mguanidine, or temperature as high as 75°C. No loss
of enzymic activity occurred under these conditions. It was concluded
that 15. stearothermophilus a-amylase is a well hydrated molecule and has
a semi-random or random c o i l in the native state (47). It was also sug
gested that secondary and tertiary structures might be maintained by
disulphide bonds (39).
Takagi and Toda (98) investigated the effect of alkaline pH on A.
oryzae a-amylase. Their observation on optical rotation and spectro-
photometric absorption with changes in enzymic activity indicated that
the modification in enzyme structure and activity was due to the dissoc
iation of hydrogen bonds which became disrupted at pH 10.5 by irreversible
ionization of phenolic hydroxyl groups of tyrosine. The activity of A.
oryzae a-amylase,- however, was regenerated after denaturation of a-amylase
by acid (99) and 8 M urea (37).

13
Isemura et a l . (41) reduced the four disulphide groups of A.
oryzae a-amylase by treating i t with sodium thioglycolate in 8.0 M urea
and found that this caused the unfolding of the linear polypeptide con
taining nine sulphydryl groups. The denaturation was reversible when
the enzyme was air-oxidized after the removal of urea and thioglycolate.
This regenerated preparation of a-amylase had 50 per cent.of the orig
inal activity.
Toda (101) studied the effect of proteolysis on A. oryzae a -
amylase and reported that modified derivatives of a-amylase had a lower
maximum velocity for the hydrolysis of amylose as compared to the native
enzyme. He suggested that the active site of the enzyme remained un
changed and that there.was an overall change in the molecular configura
tion by the formation of new secondary and tertiary structure.
G. Quarternary Structure of a-Amylase
The possession of quarternary structure of a protein implies that
a protein molecule can dissociate into two or more subunits each of which
retains i t s independent primary, secondary and tertiary structures. The
13. subtilis a-amylase in it s native form shows the phenomena of monomer-
dimer transformation. Vallee e_t a l . (105) and Stein (92) have reported
that B_. subtilis can be changed from 6S to 4S in the presence of EDTA,
and both 6S and 4S forms of the enzyme were homogeneous in the ultracen-
trifugation (95). Stein and Fisher (97) reported that other cation-
binding agents like citrate and oxalate produce heterogeneous sedimenta
tion patterns in I3_. subtilis a-amylase. ' The addition of zinc would

14
restore the dissociated amylase molecule into the homogeneous original
form. It was concluded that IL subtilis a-amylase existed in dimer form,
two units of monomer being crosslinked by an atom of zinc according to
Equation 1.
sequestering agent (Protein-Ca )-Zn-(Protein-Ca ) 2(Protein-Ca )+Zn [ l ] x x „ x
z,n
The above hypothesis was confirmed by treating the monomer form of enzyme
with to obtain a dimer. There was a direct correlation between the
release of zinc from ^~*Zn labelled enzyme when EDTA was added and a con
comitant conversion of the 6S into the 4S form of the enzyme (96).
Stein and Fisher (96) observed that other cations,such as Mn , Ni , I | | j | | _| |_ ^ |_ j |
Co and Cu gave some dimerization, while Mg , Ca , Ba and Si had no effect. A higher degree of association than dimerization was accom-
++ -9 plished when the concentration of Zn was increased to 2 X 10 M.
Isemura and Kakiuchi (39) studied the effect of pH on the sedimen
tation velocity of B>. subtilis a-amylase and"showed that Svedberg S de
creased from 6.23 to 4.45 as the pH was changed from 6.5 to 5.0 indicat
ing the involvement of the imidazole group in the dimerization process.
Isemura and Kakiuchi'(39) further explored the possible role of imidazole
groups in dimerization process by comparing the.sedimentation pattern of
the native and photo-oxidized B. subtilis a-amylase in the presence of
methylene blue. The sedimentation co-efficient was 6.2 to 4.45 for
native and photo-oxidized amylase respectively. In the case "of photo-
oxidized a-amylase, the amino acid analysis revealed that seven out of
twelve moles of h i s t i d y l residues were oxidized, while the other amino

15
acids residues were not affected. These results further supported the
hypothesis that the imidazole groups of h i s t i d y l residue are involved i n
monomer and dimer transformation of B_. subtilis a-amylase through the
chelating of zinc ions.
Stein and Fisher (95) reported that the pure crystalline a-amylases
from A. oryzae, human saliva and hog pancreas are normally present in the
4S forms which remain unchanged by the addition pf zinc ions or EDTA.
H. The Action Pattern of a-Amylase
The term "action pattern" refers here to the mechanism of cleavage
of 1-4 glucans by a-amylase. Meyer and Bernfeld (55) and Meyer and
Gonon (60) suggested that a l l a-amylases have the same action pattern
and that i t cleaved a 1-4 glycoside linkage in amylose, except those at
chain ends. Accordingly maltose.and maltotriose would be end products
of enzymatic digestion. Walker and Roberts (108) reported that the deg
radation of amylose into maltose and maltotriose.indicated semi-stable
end points, because the rate of hydrolysis of maltotriose was very low.
A further evidence used.by Meyer and his colleagues (55,60) that a l l a-
amylases have the same action pattern was the measurement of saccharo-
genic/dextrinogenic quotient, which gave similar values for different
amylases (33). This hypothesis was cr i t i c i s e d because the ratios were
taken at the same stage of amylolyses, and therefore differences could
not be expected. When saccharogenic and dextrinogenic ratios were deter
mined near the achroic point very wide differences were apparent between
various amylases (81). The plot of blue value against reducing value

16
gave characteristic curves for different a-amylases. The difference was
possibly due to different chain lengths produced by the enzymic hydroly
sis of amylose by a-amylases (44). Subsequent studies based on paper
and column chromatographic techniques have revealed that a-amylases of
different origins produced low molecular weight products with molecular
size distribution characteristic of individual enzymes (17,76,79,110).
Robyt and French (81) reported that pancreatic and human salivary a-
amylase produced very similar end products from amylose. However, the
curves relating drop in blue values to the corresponding increase in the
reducing values were different. In the light-of these results these
authors did not accept the explanation offered by Kung et a l , (44) re
garding differences between various amylase curves relating drop in blue
values to corresponding increases in the reducing value. Bird and
Hopkins (5) reported another aspect of action pattern in which dif f e r
ences were observed even with a-amylase from the same source when the
substrate concentration was changed. These results indicated that
Meyer's hypothesis regarding equal rate of hydrolysis of a l l but end
linkage was not valid for various a-amylases.
Robyt and French (79), and Bird and Hopkins (5) have reported
that eventually a l l the amylose would be hydrolyzed to maltose and glu
cose by a-amylases through different action, patterns. The linear portion
of glycogen and amylopectin essentially follows the same fate as amylose
to produce maltose and glucose.
In the case of amylopectin, which is a branched polymer, the
limit dextrin produced by the action of salivary a-amylase was found to

17
contain 1—>k and•1—*6 bonds, ranging.from the pentasaccharide upwards,
and moreover these large molecules have two and three 1—̂ 6 bonds (68,76).
Wheal and Roberts (113) have suggested that salivary a-amylase cannot
cleave certain 1—>4 bonds in the v i c i n i t y of the 1—>6 branched points,
and this concept has been extended to other a-amylases as well (28).
Robyt and French (81) have considered the action pattern of a-
amylases on amylose in terms of single chain, multichain, and multiple
attack. They suggested that porcine, pancreatic, human salivary and A.
oryzae a-amylases follow multiple attack patterns during amylolysis and
they also calculated the degree df multiple attack by these enzymes.
Leach and Schoch (45) studied the action of various a-amylases on
starch granules and found that different types of starches have varying
degrees of susceptibility to amylases. In addition they observed no
correlation between granule size and the extent of. solubilization. Simi
lar differences have been reported by Walker and Hope (109) in the sus
ceptibility of starches of different origin to amylases. Their results
also indicated that porcine pancreatic and human salivary a-amylases
were adsorbed on the surface of the corn starch granules, while the
sweet potato B-amylase and A. oryzae a-amylase were not adsorbed. The
a-amylase from S_. bovis and C_. butyricum can also degrade corn starch
granules (109). Nordin and Kim (69) observed an apparent increase in
the amylose content during the i n i t i a l period of degradation of starch
granules, as measured by the potentiometric titration of bound iodine.
It was concluded that amylopectin must be degraded f i r s t , indicating that
i t constituted the external covering of starch granules. The location

18
of amylopectin with respect to amylose in starch granules is in agreement
with the hypothesis of Ulmann (103).
I. Immunochemical Study of a-Amylase
1. Mammalian a-Amylase
In recent years antibodies have been produced against a number of
mammalian enzymes, for example, phosphorylase (31), lactate dehydrogenase
(65), alkaline phosphatase (85) and ribonuclease (13). Antisera thus ob
tained have been used to compare and contrast enzymes from different
organs or species (31,48,86).
McGeachin and Reynolds (52) were the f i r s t workers to report that
mammalian a-amylase could act as an antigen to produce antibodies. They
used amylase antiserum to study the relationship of hog pancreatic amylase
to the amylase of other hog organs and to amylases of other species.
McGeachin (53) has reported immunological techniques for determining d i f
ferences and similarities among amylases of various species and also of'
a given species.
2. • Microbial a-Amylase
Wada (107) demonstrated that when crystalline Taka a-amylase was
injected into rabbits the antibody was formed against the enzyme. He
also studied the serological properties of the anti-sera produced and
found only a single homogenous antibody,. but this antibody could not i n
hibit enzyme activity completely. He further observed that starch and
starch hydrolysates inhibited the amylase-antiamylase reaction. It was
found that anti-Taka-amylase antibody specifically inhibited the activity

19 of a-amylase from Aspergillus species. On the other hand, a-amylase
activity of other molds, bacteria and a-amylase of a l l other sources
tested were unaffected (107). Heat denatured Taka-a-amylase did not i n
hibit the reaction between Taka-a-amylase and i t s antibody.
Nomura and Wada (67) obtained antibodies by injecting crystalline
—' subtilis a-amylase into rabbits. Antiserum produced in rabbits by
injection of crystalline amylase neutralized the enzymic activity to
about 90 per cent. A competitive inhibition of the action of antisera
by the substrate, starch and i t s hydrolysed products (67,108,109) was
also noted.
Onoue elt -al. (74) modified the 13. subtilis a-amylase by treating
i t with N-bromosuccinimide to study the molecular configuration between
modified and native a-amylase by immunochemical analysis. They reported
that anti-bacterial a-amylase gave almost identical precipitation curves
when treated with bacterial a-amylase and modified N-bromosuccinimide-
bacterial a-amylase. In.addition, by.using the agar-gel immunodiffusion
technique, they observed a single sharp precipitation line between N-
bromosuccinimide-bacterial a-amylase and anti-bacterial a-amylase, and
the precipitation line fused together with the. line between bacterial a-
amylase and anti-bacterial a-amylase. Onoue e_t a l . (74) reported that
N-bromosuccinimide treatment did not change the molecular configuration
of the enzyme and the loss of enzymic activity was due to oxidation of
one tryptophan residue. These results indicated that the catalytic site
of bacterial a-amylase might be different from that of the antigenic
site. Onoue ej: a l . (75) prepared purified antibodies against 13. subtilis

20
a-amylase and the purified antibodies neutralized the a-amylase activity
completely. The antibody in the antigen-antibody complex could not be
displaced by substrate. These results are not consistent with the ear
l i e r work of Nomura and Wada (67) in which they reported that 10 per cent
enzymic activity remained after antibody treatment. Onoue et a l . (75)
also demonstrated that the neutralizing a b i l i t y of papain treated anti
body was less than that of the intact antibody, though the papain digested
antibody had the capacity to combine with the antigen. ' It was suggested
from these results that the antibody affected the interaction of amylase
and starch by steric hindrance and therefore would be expected to decrease
when the molecular size of antibody is reduced. Okada,et a l . (71) re
ported that photo-oxidized a-amylase of .13. subtilis did not form a pre
cipitate with I3_. subtilis a-amylase antibody. It was further demonstra
ted that ih the presence of calcium, photo-oxidized 13. subtilis a -
amylase was not susceptible to proteinase indicating that photo-oxidation
did not grossly change the molecular configuration (29).
Okada et a l . (72) reported that a-amylase activity of both Taka-
amylase A and p-phenylazobenzyl-Taka-amylase A was inhibited up to the
same degree by anti-TakaTamylase A and by anti-p-phenylazobenzyl-Taka-
amylase A. It was suggested that antibody to the altered protein moiety
of p-phenylazobenzoyl-Taka-amylase A was produced (72). Okada et a l .
(72) further observed that the maltosidase activity of Taka-amylase A
was partially inhibited by anti-Taka-amylase A and anti-p-phenylazo-
benzyl-Taka-amylase A was ineffective to inhibit the enzyme activity.
Moreover, the maltosidase activity of p-phenylazobenzyl-Taka-amylase A

21
was not neutralized by anti-Taka-amylase A or anti-p-phenylazobenzyl-Taka-
amylase A. Since the neutralizing ab i l i t y of the antibody depends on the
molecular size of the substrate (starch, phenyl maltoside) i t was sugges
ted that the antibody inhibited enzymic activity by steric hindrance (72).
Sirisinha and Allen (90) used immunochemical methods to study the
structure of Aspergillus a-amylase. Urea treated native enzyme under
various conditions resulted in a preparation which gave a reaction partly
identical with the non-treated enzyme during immunodiffusion analysis.
Quantitative precipitation curves with urea treated enzyme preparation
indicated that only a partial loss of immunochemical reactivity occurred
even with prolonged treatment. The appearance of several bands of pre
cipitation with urea treated enzyme preparation suggested that various
intermediate states exist between the fully unfolded structure of protein
and the native protein (90). Immunochemical changes were also observed
with enzyme preparation treated with EDTA alone or in combination with
.1 M mercaptoethanol.
Sirisinha and Allen (91) reported marked differences regarding im
munochemical behaviour between urea treated and oxidized a-amylase from
A. oryzae. Although oxidized a-amylase would precipitate the same amount
of antibody, the efficiency of oxidized enzyme decreased per unit weight.
On the other hand, urea treated a-amylase would precipitate only a cer
tain portion of antibody from a specific antiserum. These authors also
suggested that antigenic sites are not involved with the catalytic activ
ity and the decreased activity shown by enzyme antibody complex is due to
the steric hindrance caused by attachment of antibody with respect to the
catalytic center.

REFERENCES I, . II
1. Akabori, S., T. Ikenda, H. Hanafusa and Y. Okada. 1954. Studies on taka-amylase A. II. Amino acid composition of taka-amylase A. J. Biochem. Tokyo. 41:803.
2. B. Maruo, M. Nomura and H. Mitsui. 1955. An attempt to study the mechanism of amylase action by the use of C^-labelled starch. J. Gen. App. Microbiol. L : l .
3. , Y. Okada, S.Fujiwara and K. J. Sugae. 1956. Studies on bacterial amylase. I. Amino acid composition of crystalline bacterial amylase from B_. subtilis N. J. Biochem. Tokyo. 43:741.
4. Bines, B. J. 1956. The a-amylase from barley malt. Ph.D. thesis, University of Wales.
5. Bird, R. and R. H. Hopkins. 1954. The action of some a-amylase on amylose. Biochem. J. 56:86.
6. Blackburn, T. H. 1968. Protease production by Bacteroides amylophilus Strain H 18. J. Gen. Microbiol.. 53:27.
7. Caldwell, M. L., C. E. Weill and R. S. Weill. 1945. Further studies of the essential groups of pancreatic amylase. J. Amer. Chem. Soc. 67:1079.
8. , M. Adams,. J. F. Rung and G. C. Toralballa. 1952. Crystalline pancreatic amylase. II. Improved method-for i t s preparation from hog pancreas glands and additional studies of it s properties. J. Amer. Chem. Soc. 74:4033.
9. , E. S. Dickey, V. M. Hanrahan, H. C. Kung., J. F. Kung and M. Misko. 1954. Amino acid composition of crystalline pancreatic amylase from swine. J. Amer. Chem. Soc. 76:143.
10. Campbell, L. L. 1954. Crystallization of alpha-amylase from a. thermophilic bacterium. J. Amer. Chem. Soc. 76:5256.
11. and P. D. Cleveland. 1961. Thermostable a-amylase of Bacillus.stearothermophilus. Crystallization and some general properties. J. Biol. Chem. 236:2952.

23
12. Campbell, L. L. :and G. B. Manning. 1961. Thermostable a-amylase of Bacillus stearothermophilus. III. Amino acid composition. J. Biol. Chem. 236:2962.
13. Carter, B. G., B. Cinader and C. A. Ross. 1961. Immunochemical analysis of the multiple forms of bovine ribonuclease. Ann. N. Y. Acad. Sci. 94:1004.
14. Coleman, G. and W. H. E l l i o t t . 1962. Studies on a-amylase formation by Bacillus s u b t i l i s . J. Biochem. 83:256.
15. Danielsson, C. E. 1947. Molecular weight of a-amylase. Nature. 160:899.
16. Dube, S. K. and P. Nordin. 1961. Isolation and properties of Sorgum a-amylase. Arch. Biochem. Biophys. 94:121.
17. . 1962. The action pattern of Sorghum a-amylase. Arch. Biochem. Biophys. 99:105.
18.. Eisenstadt, J.. M. and H. P. Klein. 1959. Sulphur incorporation into the a-amylase of P_. saccharophijia. J. Bacteriol. 77:661.
19. . 1961. Evidence for a de novo synthesis of the a-amylase of P_. saccharophilia. J. Bacteriol. 82:798.
20. Fischer, E. H., F. Duckert and P. Bernfeld. 1950. Isolement et c r i s t a l l i s a t i o n de 1'a-amylase de pancreas humain sur les enzymes amylolytiques XIV. Helv. Chim. Acta. 33:1060.
21. _ and R. DeMontmollin. 1951. Purification et c r i s t a l l i s a tion de l'a-amylase d'Aspergillus oryzae. Sur les enzymes amylolytiques. Helv. Chim. Acta. 34:1987;
22. and C. H. Haselbach. 1951. Contribution a l'etude de l'a-amylase de malt. Sur.les enzymes amylolytiques XVII. Helv. Chim. Acta. 34:325.
23. , W. N. Summerwell, J. M. Junge and E. A. Stein. . 1958. Calcium and molecular structure of a-amylases. Proc. 4th Inter. Congr. Biochem. Vienn. Pergamon Press. 8:124.
24. _ _ _ _ _ and D. E. Stein. 1960. "a-Amylase." In the enzyme. Ed. by P. D. Boyer, H. Lardy, and-K. Myrback. Vol. 4. Academic Press. New York and London.
25. and E. A. Stein. 1961. a-Amylase from human saliva. Biochemical Preparation. 8:27.

24
26. Freeman, N. M. and R. H. Hopkins. 1936. LXXII. The mechanism of degradation of starch by amylases. I I I . Mutorotation of f i s s i o n products. J . Biochem. 30:451.
27. French, D. 1957. The schardinger dextrins. Adv. Carbohydrate-Chem. 12:189.
28. _, M. L. Levine, E. Norberg, P. Nordin, J . Pazur and G. Wild. 1954. Studies on the schardinger dextrins. VII. Co-substrate s p e c i f i c i t y i n coupling reaction of Macerans amylase. J . Amer. Chem. Soc. 76:2387.
29. Hagihara, B., T. Nakayama, H. Matsubara and K. Okunuki. 1956. Denaturation and i n a c t i v a t i o n of enzyme proteins. I I . Denaturation and i n a c t i v a t i o n of b a c t e r i a l amylase. J . Biochem. 43:469. '
30. Heatley, N. G. 1958. Spontaneous c r y s t a l l i s a t i o n of amylase from pancreatic j u i c e of the r a t . Nature. 181:1069.
31. Henion, W. F. and E.-W. Sutherland. 1957. Immunological d i f f e r ences of phosphorylases. J . B i o l . Chem. 224:477.
32. Hobson, P. N. and Margaret Macpherson. 1952. Amylase of C l o s t r i -dium butyricum and a streptococcus i s o l a t e d from rumen of the sheep. J . Biochem. 52:671.
33. Hodge, J . E., E. M. Montgomery and G. E; Hiebert. 1948. Hydrolysis of the amylopectins from various starches with Beta-amylase. Cereal Chem. 25:19.
34. Hsui, J . , E.- H. Fisher and E. A. Stein. 1964. Alpha-amylases as calcium-metalloenzymes. II.- Calcium and the c a t a l y t i c a c t i v i t y . Biochemistry. 3:61.
35. Hungate, R. E. ' 1966. 'The rumen bacteria." In the Rumen and Its Microbes. By R. E. Hungate. Academic Press. New York and London.
36. Ikenaka, T. 1959; Chemical modification on taka-amylase A. I. Dinitrophenylation of taka-amylase A. J . Biochem. Tokyo. 46:177.'
37. . 1959. Chemical modification on taka-amylase A. I I . Phenylazobenzoylation of taka-amylase A. J . Biochem. Tokyo.-46:297.
38. Isemiira, T. and S. F u j i t a . 1957. Physicochemical studies on taka^ amylase A. I. Size and shape determination by the measurement of sedimentation constant, d i f f u s i o n c o e f f i c i e n t , and v i s c o s i t y . J . Biochem. Tokyo. 44:443.

25
39. Isemura, T. and K. Kakiuchi. 1962. Association and dissociation of bacterial a-amylase molecule. J. Biochem. Tokyo. 51:385.
40. and A. Imanishi. 1962. Reversibility of denaturation and molecular s t a b i l i t y of Bacillus subtilis a-amylase. J. = Biochem. Tokyo. 51:172.
41. , T. Takagi, Y. Maeda and K. Ytani. 1963. Recovery of the intact structure of taka-amylase A. after reduction of a l l disulphide linkages in 8 M urea. J. Biochem. Tokyo. 53:155.
42. Junge, J. M. , E.- A. Stein, H. Neurath and E. H. Stein. 1959. The amino acid composition of a-amylase from Bacillus s u b t i l i s . J. Biol. Chem. 234:556.
43. Kuhn, R. 1925. Der wirkungsmechanismus der amylasen; ein beitrag zum knofiguration sproblem der starke. Liebigs Ann. 443:1.
44. Kung., J. F. , V. M. Hanrahan and M. L. Caldwell. 1953. A comparison of the action of several alpha amylases upon a linear fraction from corn starch. J. Amer. Chem. Soc. 75:4438.
45.. Leach, H. W. and T. J. Schoch. 1961. Structure of the starch granules. II. Action of various amylases on granular starches. Cereal Chem. 38:34.
46. L i t t l e , J. E. and H. L. Caldwell. 1942; A study of the action of pancreatic amylase. J. Biol. Chem. 142:585.
47. Manning, G. C., L. L. Campbell and R. J. Foster. 1961. Thermostable a-amylase of Bacillus stearothermophilus. J. Biol. Chem. 236:2958.
48. Mansour, T. G., E. Bueding and A. B. Stavitsky. 1954. The effect of a specific antiserum on the activities of lactic dehydrogenase of mammalian muscle and of Schistosoma mansoni. Brit. J. Pharmacol. 9:182.
49. Markovitz, A. and H. P. Klein. 1955. Some aspects of induced biosynthesis of a-amylase of P_. saccharophilia. J. Bacterol. 70:641.
50. . . 1955. On the study of carbon for the i n duced biosynthesis of a-amylase in P_. saccharophilia. J. Bacterol. 70:649.
51. , and E. H. Fisher. 1956. Purification, crystals lization,.and properties of the a-amylase of Pseudomonas saccharophila. Biochem. Biophys. Acta. 19:267.

26
52. McGeachin, R. L. and.J. M. Reynolds. 1959. Difference in mammalian amylases demonstrated by enzyme inhibition with specific antisera. J. Biol. Chem.. 234:1456.
53. . 1968. Multiple molecular forms of amylase. Ann. N. Y. Acad. Sci. 151:208.
54. Menzi, R., A. Stein and E. H. Fisher. 1957. Proprietes de deux a-amylase de 13. s u b t i l i s . Sur les enzymes amylolytiques. Helv. Chim. Acta. 40:534.
55. Meyer, K. H. and P. Bernfeld. 1941. Recherches sur l'amidon XIV. La reaction coloree a l'iode de l'amidon et du glycogene. Helv. Chim. Acta. 24:389,
56. , M. Fuld and P. Bernfeld. 1947. Purification et c r i s t a l -lisation de 1'a-amylase de bacterie. Experentia. 3:411.
57. , E. H. Fisher and P. Bernfeld. 1947. Sur les enzymes amylolytiques (1). L'isolement de 1'a-amylase de pancreas'. Helv. Chim. Acta. 30:64.
58. , . 1948. Sur les enzymes amylolytiques. Isolement e t . c r i s t a l l i s a t i o n de 1'a-amylase de salive humaine. Helv. Chim. Acta. 31:2158.
59. , , A. Staub and P. Bernfeld. 1948. Proprietes de 1'a-amylase de salive humaine c r i s t a l l i s e e . Helv. Chim. Acta. 31:2165.
60. and W. F. Gonon. 1951. La degradation de l'amylose par les a-amylases. Helv. Chim. Acta. 34:294.
61. . 1952; The past and present of starch chemistry. Experentia. 8:405.
62. Muus,.J. 1954. The amino acid composition of human salivary amylase. J. Amer. Chem. Soc. 76:5163.
63. , F. P. Brockett and C. C. Connelly. 1956. The effect of various ions on the stability of crystalline salivary amylase in solution.-Arch. Biophys. 65:268.
64. - Myrback, K. 1926. Uber verbindungen einiger enzyme mit inak-. tiverenden stoffen. II. J. Physiol. Chem. 159:1.
65. Nisselbaum, J. A. and 0. Bodansky. 1960. Reaction of human tissue la c t i c dehydrogenases with antisera to human heart and liver l a c t i c dehydrogenases. J. Biol. Chem. 236:401.

27
66. Nomura, M. , J.- -Hosoda and H. Yoshikawa. 1958. Studies on amylase - formation by Bacillus, s u b t i l i s . VI .• The mechanism of amylase excretion arid cellular structure of Bacillus s u b t i l i s . J. Biochem. Tokyo. 45:737.
67. , and T. Wada. 1958. Studies on amylase formation by Bacillus s u b t i l i s . V. Immunochemical studies of amylase produced by Bacillus s u b t i l i s . J. Biochem. 45:629.
68. Nordin, P. and D. French. 1958. I. Phenyl-flavazole derivatives of starch dextrins. J. Amer. Chem. Soc;. 80:1445.
69. and Y. S. Kim. 1960. The reaction of.amylases with starch granules. J. Amer. Chem. Soc. 82:4604.
70. Ohlsson, E. 1930. Uber die beiden komponenten der malzdiastase, besonders mit rucksicht auf die mutarotation der bei der hydro-lyse der starke gebildeten product. J. Physiol. Chem. 189:17.
71. Okada, Y., S. Nakashima and Y.-Yamamura. 1963. Relationship between immunological memory and structure of bacterial a-amylase. J. Biochem. 54:99.
72. , Y. Matsuoka, T. Yagura, T. Kenda and Y. Yamamura. 1964. Immunochemical study of taka-amylase A and Phenylazobenzoyl taka-amylase A. J. Biochem. 55:446.
73. Ono, S.j.K. Hiromi and Y. Yoshikawa. 1958. Kinetics of hydro-l y t i c reaction catalyzed by crystalline bacterial a-amylase. Bull. Chem. Soc. Japan. 31:957.
74. Onoue, K., Y. Okada and Y. Yamamura. 1962. Modification of bact e r i a l a-amylase with N-brobosuccinimide. J. Biochem. 51:443.
75. , , S. Nakashima, K. Shimada and Y. Yamamura. 1963. Studies on enzyme-antienzyme .system. I. Immunochemical studies on Bacillus subtilis a-amylase. J. Biochem. 53:472.
76. Pazur, J. H., D. French and D. Knapp. 1950. Mechanisms of s a l i vary amylase action. Proc. Iowa Acad. Sci. 57:203.
77. Pollock, M. R. 1962. "Exoenzyme." In.the Bacteria. Ed. by I. C. Gunsalus and R. Y. Stanier. Vol. 4. Acad. Press. New York and London.
78. Roberts, P. J. -P., and W. J.. Whelan. 1960. The mechanism of carbohydrase action.. V. Action of human salivary a-amylase on amylopectin and glycogen. Biochem. J. 76:246.

28
79. Robyt, J. F. and D..French. 1963. Action pattern and specificity of an amylase from Bacillus s u b t i l i s . Arch. Biochem. Biophys. 100:451.
80. • 1964. Purification and action pattern of an amylase from Bacillus polymyxa. Arch. Biochem. Biophys. 104:338;
81. . 1967. Multiple attack hypothesis of c t-amylase action: action of porcine, pancreatic, human salivary and Aspergillus oryzae a-amylase. Arch. Biochem. Biophys. 122:8.
82. . and J. W. Whealn. 1968. "The a-amylase." In starch and ists derivatives. Ed. by J. A. Radley. Fourth Edition. Chapman and Hall Ltd., 11 New Fetter Lane, London EC4.
83. Rose, D. 1948. The amylase of Bacillus polymyxa. Arch. Biochem. 16:349.
84. Schiff, J. A., J. M. Eisenstadt and H. P. Klein. 1959. a-amylase formation in growing and non-growing cells of P_, saccharophila. J. Bacteriol. 78:124.
85. Schlamowitz, M. and 0. Bodansky. 1959. Tissue sources of human serum alkaline phosphatase as determined by immunochemical procedures. J. Biol. Chem. 234:1433.
86. . 1954. Specificity of dog intestinal phosphatase antiserum. J. Biol. Chem. 206:369.
87. Schwimmer, S. and A. K. Balls. 1949. Isolation and properties of crystalline a-amylase from germinated barley. J. Biol. Chem. 179:1063.
88. . and J. A. Garibaldi. 1952. Further studies on the production, .purification and properties of ..the Scharadinger dex-trinogenase of macerans. Cereal Chem. 29:108.
89. . 1953. Evidence for the purity of Schardinger dextrin-ogeriase. Arch. Biochem. Biophys. 43:108.
90. Sirisinha, S. and P. Z. Allen. 1965. Immunochemical studies on a-amylase. I.. Effect of denaturing agents and.proteolytic enzymes on the immunochemical reactivity of a-amylase from Aspergillus oryzae. Arch. Biochem. Biophys. 112:137.
91. . 1965. Immunochemical studies on a-amylase. II. Examination of immunochemical and enzymic activities of native and modified a-amylase from Aspergillus oryzae. Arch. Biochem. Biophys. 112:149.

29
92. Stein, E. A. 1957. Structure of s u b t i l i s a-amylase. Federat i o n Proc. 16:254.
93. and.E. H. Fischer. 1958. The resistance of a-amylase towards p r o t e o l y t i c attack. J . B i o l . Chem. 232:867.
94. , J . M. Junge and E. H. Fisher. 1960. The amino acid comp o s i t i o n of a-amylase from A s p e r g i l l u s oryzae. J . B i o l . Chem. 235:371.
95. and E..H. Fischer. 1960. B a c i l l u s s u b t i l i s a-amylase^ a z i n - p r o t e i n complex. Biochem. Biophys. Acta, 39:287.
96. ' . 1961. a-Amylase from B a c i l l u s s u b t i l i s . Biochemical Preparation. 8:34.
97. , J . Hsui and E. H. Fisher. 1964. Alpha amylase as c a l -cium-metalloenzymes. I. Preparation of calcium-free apoamy-lases by chelation and e l e c t r o d i a l y s i s . Biochemistry. 3:56.
98. Takagi, T. and H. Toda. 1960. Studies on the amphoteric propert i e s of taka-amylase A. I. Ionization of phenolyic hydroxyl groups. J . Biochem. Tokyo.8:781.
99. . 1962. Studies on the denaturation of taka-amylase A and on i t s r e v e r s i b i l i t y . J . Biochem. Tokyo. 52:16.
100. Thoma, J. A., J . Wakim and L. Stewart. 1963. Comparison of the ac t i v e s i t e s of alpha and beta amylase. Biochem. Biophys. Res. Comm. 12:350.
101. Toda, H. 1963. Enzymatic modification of phenylazobenzoyl-taka amylase A. J . Biochem. Tokyo. 53:425.
102. Tsuchiya, H. M. , J-. Corman and H. J . Koepsell. 1950. Production of mold amylases i n submerged culture. I I . Factors a f f e c t i n g the production of alpha-amylase and maltase by c e r t a i n A s p e r g i l l i . Cereal Chem. 27:322.
103. Ulmann, M. 1957. Bestimmung der chemischen natur der h i l l l e cines gerguollenen starkekornes. K o l l o i d . Z. 150:128.
104. Underkofler, L. A. and.D. K. Roy.. 1951. C r y s t a l l i z a t i o n of fungal alpha-amylase and l i m i t dextrinase. Cereal Chem. 28:18.
105. V a l l e e , B. L., E. A. Stein, W. N. Summerwell and.E. H. Fischer. 1959. Metal content of a-amylases of various o r i g i n s . J . B i o l . Chem. 234:2901.

30
106. Wada, T. and M. Nomura. 1958. An immunochemical study of microb i a l amylase (1). J. Biochem. 45:639.
107. . . 1959. An immunochemical study of microbial amylase (11) J. Biochem. 46:239.
108. Walker, G. J. and W. J. Whelan. 1960. The mechanism of carbo-hydrase action. VII. Stages in the salivary a-amylosis of amylose, amylopectin and glycogen. Biochem. J.. 76:257.
109. and P. M. Hope. 1963. The action of some a-amylases on starch granules. Biochem. J. 86:452.
110. -. - •;• 1965. The c e l l bound a-amylase of Streptococcus bovis. Biochem. J. 94:289.
111. Welker, N. E. and L. L. Campbell. 1963. Effect of carbon source on formation of a-amylase by Bacillus stearothermophilus. J. Bact; 86:681.
112. . 1963. Induction of a-amylase of Bacillus stearothermophilus. J. Bact. 86:687.
113. Whelan, W. J. and P. J. P. Roberts. 1952. Action of salivary a -amylase on amylopectin and glycogen. Nature. 170:748.
114. Yamamoto, T. and J. Fukumoto. 1960. Enzymatic properties of bacterial a-amylase reactivated with various alkaline earth metals. Bull. Agr. Chem. Soc.- Japan. 24:16.
115. Yoshida, A. and T. Tobita. 1960. Studies on the mechanism of protein,synthesis. Non-uniform incorporation of [c!4] leucine into a-amylase and the presence of a-amylase precursor. Biochem. Biophys. Acta. 37:513.

CHAPTER III
MATERIALS AND METHODS
A. Chemicals
The sources from which the substrates and chemicals were ob
tained are as follows: starch (British Drug House, Poole, England),
amylose (Stein-Hall and Co., New York, U.S.A.), maltose, technical and
reagent grade (Fisher Scientific Co., New Jersey, U.S.A.), bovine serum
albumin (Calbiochem, Los Angeles, California, U.S.A.), casein hydroly
sate, and a-amylases from Ii. s u b t i l i s , A. oryzae and hog pancreas (Sigma
Chemical Company, St..Louis, U.S.A.). A l l chemicals used during this
investigation were of the highest purity grade and were obtained through
Allie d Chemical Company Canada, Ltd., Vancouver, B.C., and Fisher
Scientific Co. Ltd., Vancouver, B.C.
DEAE Sephadex A-50 and Sephadex G-200 were purchased from
Pharmacia, Uppsala, Sweden.
B. Organism
The organism used in this investigation was Bacteroides amylo
philus strain H-18, kindly supplied by Dr. T. H. Blackburn, Department
of Microbiology, University of British Columbia, Vancouver 8, B.C.,
Canada. Blackburn and Hobson (2) isolated this strain from the rumen of
sheep.

32
C. Maintenance of Bacteroides amylophilus Strain H-18
The complete chemically defined basal medium used during this
investigation was that developed by Hungate (8). This medium contained
(g/1): K2HP04, 0.45; KH2P04, 0.45; (NH^SO^ 0.9; NaCl, 0.9; HgS04,
0.09; CaCl^, 0.09; resazurin, .001; L-cysteine hydrochloride, 0.5. The
resazurin and mineral solutions, or any additions to the medium were
placed in a screw capped bottle and d i s t i l l e d water added to give a
f i n a l volume of 900 ml. The medium was autoclaved for fifteen minutes
at 120°C and on removal of the bottle from the autoclave the cap was
immediately screwed tight. Fifty ml. of 1 per cent (w/v) L-cysteine
hydrochloride and f i f t y ml. of 10 per cent (w/v) sodium bicarbonate sol
ution were steam autoclaved separately at 100°C for fifteen minutes and
then added to the remaining medium under a stream of CO2. The f i n a l pH
of the medium was 6.7. A l l the dispensing of the medium and incubation
of the culture was done under oxygen-free CO^ as described by Blackburn
(3).
Stock cultures were maintained on nutrient agar slopes which
contained in addition to the basal medium 2 per cent agar and 0.5 per
cent each of maltose and casein. The cultures were stored at 4°C and
transfers were made each week to fresh slopes by stab innoculation under
an atmosphere of CO^. A l l cultures were grown at 38°C.
D. Growth Measurements of Bacteroides amylophilus Strain H-18
The growth of _. amylophilus was measured in a Bausch and Lomb

33
Spectronic 20 Colorimeter (Bausch and Lomb, Rochester, New York, U.S.A.)
at 660 nm.
E. Production and Purification of a-Amylase from Bacteroides amylophilus Strain H-18
The amylase was a by-product of protease purification undertaken
by Lesk (9) who kindly denoted the fraction III a-amylase (Figure 6) at
the point where i t was separated from the protease.
1. Production of a-Amylase
Twenty-nine l i t e r s of growth medium were prepared i n a 32 l i t r e
stainless steel milk can and inoculated with 1 l i t r e of log phase culture
of .B. amylophilus. After anaerobic incubation for 23 hours at 38°C the
can and i t s contents were cooled immediately with a waterhose. The c e l l
was removed by continuous flow centrifugation (8700 x g at 4°C) using a
Servall Centrifuge (Servall type SS-34, equipped with a KSB:R Servall
continuous flow adopter from Servall, Norwalk, Connecticut). The super
natant had a pH of 5.5 which was the optimum for the attachment to DEAE
Sephadex A-50 for purification.
2. Purification of a-Amylase on DEAE-Sephadex A-50 and G-200•Sephadex •
DEAE Sephadex A-50 (0.2 g dry weight/100 ml. of supernatant) was
gradually added to the supernatant and CO 2 was bubbled through i t for
twelve hours at 4°C to ensure proper mixing. The DEAE suspension was
allowed to settle; the supernatant decanted and the DEAE collected on a
sintered glass f i l t e r . The DEAE was mixed thoroughly in 500 ml. of 1 M

34
NaCl and centrifuged. The supernatant was then decanted and stored.
This procedure was repeated six times. The f i r s t five fractions (total
volume 2540 ml.) were pooled and dialyzed against 0.05 M phosphate
buffer (pH 7.0) overnight.
The dialysed preparation was further fractionated by chromatog
raphy on a 100 x 5 cm. column of DEAE Sephadex equilibrated with 0.05 M
phosphate buffer (pH 7.0).'
a-Amylase was eluted with linear gradient of 0.2 M to 1.0 M NaCl
in phosphate buffer (pH 7.0). The fractions were tested for a-amylase
activity and three fractions having enzymic activity were collected.
Volumes of 538 ml, ,500 ml and 840 ml were collected for fraction I, II,
and III respectively. Each fraction was dialysed against 0.05 M phos
phate buffer (pH 7.0) overnight at 4°C. Fraction III was concentrated
to 40 ml with a Diaflo U l t r a - f i l t r a t i o n Cell (Diaflo Model 50 Ultra
f i l t r a t i o n Cell from Amicon Co., Lexington, Mass., U.S.A.) equipped with
a Pm - 10 f i l t e r (exclusion limit;10,000 MW) under a pressure of 40 p.s.i.
The concentrated a-amylase fraction III was further purified on a 2.5 x
50 cm column of Sephadex G-200 which had been equilibrated and eluted
with 0.05 M phosphate buffer (pH 7.0). A l l of the a-amylase activity was
obtained in a single peak and the enzyme solution was concentrated by
pressure dialysis to 45.0 ml.
F. Assay of a-Amylase •
The a-amylase activity in the sample was assayed by determining
the amount of reducing sugars produced from starch or amylose using the

35
3.5 d i n i t r o s a l i c y l i c acid method of Fisher and Stein (5). The assay
medium consisted of 1.0 ml of properly diluted enzyme in an equal volume
of 2 per cent (w/v) soluble starch or amylose buffered to pH 6.7 with
0.2 M Tris and 0.1 M maleate. Unless otherwise indicated the time of
incubation was fifteen minutes at 44°C.
When i t was desired to calculate the degree of multiple attack,
the a-amylase activity was determined by the Nelson Copper method (16).
A unit of a-amylase activity was defined as the amount of enzyme
that would produce the equivalent of 1.0 mg. maltose in one minute under
standard.conditions.
The method of Robyt and Whelan (21) was used to determine the
blue values.
G. Assay of Protease
The determination of protease activity was done according to the
method of Blackburn (3).
H. Determination of Nitrogen
The indophenol colorimeter method of Nakai and Tsuchiya (14) was
used for nitrogen determination. Bovine serum albumin was used as the
standard.
I. Determination of Protein
The method of Lowrey et_ a l . (11) , was used to measure the protein
concentration in the extracts. Bovine serum was used as the standard.

36
J. Determination of Total Carbohydrates
Total carbohydrates were determined by phenol and sulphuric acid
procedure as described by Miller (13).
K. Disc Gel Electrophoresis
Disc gel electrophoresis apparatus was constructed by Mr. R. J.
Hudson and Mr. James A. Shelford, i n the Department of Animal Science
Laboratory, University of British Columbia, following the procedure of
Davis (4). Disc electrophoresis chemical r e f i l l pack containing standard
7 per cent acrylamide gel and premixed stock reagents, acrylamide, b i -
sacrylamide (N, N'-methylenebisacrylamide) and (N, N, N', N'-tetramethyl-
ethylene diamine) TEMED was purchased from Canalco, Rockville, Maryland,
U.S.A.
Disc gel electrophoresis was carried out in a standard gel (7 per
cent) according to the method described by Davis (4). The gels were
stacked at pH 8.9 and run at pH 9.5 during routine work. The enzyme
samples were run in duplicate. After gel electrophoresis for two hours
(5 mA per column) one gel was stained immediately for protein with Amido
black 10B. The second gel.was used to detect a-amylase activity by lay
ering the gel.on starch coated glass slides. The glass slides were
coated f i r s t with 1.0 per cent agar and then with 1.0 per cent starch
plus 1.0 per cent agar made in 2 M TRIS-Maleate buffer at pH 6.7. After
being superimposed with the gel, the slides were placed in petri dishes
and.incubated at 40°C for 15 minutes. After the incubation the gels were
removed and the slides dipped momentarily in Lugol's iodine solution to

37
stain the unhydrolysed starch. The clear band (S) indicating hydrolysis
of starch was visible against the blue stain produced by the starch.
L. Isoelectrofocusing
The LKB fraction collector with Uvicord (0.3 on light path), and
Ampholine Electrofocusing equipment (LKB Produkter AB, Stockholm, Brouma,
Sweden) were used.
The Ampholine column and sample were prepared according to the
instructions given in LKB 8100 Ampholine Instruction Manual. The gradient
mixer equipped with s t i r r e r motor was used to f i l l the Ampholine column,
No. 8102, capacity 440 ml. Low molecular weight ampholines in the range
pH 3.0 to 10.0 (4 per.cent).were used.
M. Charcoal-Celite Column Chromatography
The technique was essentially the same as described by Whistler
and Duro (22) using a charcoal-celite column to isolate and detect oligo
saccharides found in technical grade Maltose. A 50 per cent solution of
the sugar was autoclaved for 20 minutes at 40 lbs. (p.s.i.) and filtered
to remove the precipitate.
A chromatographic column (4.5 x 50 cm.) was f i l l e d to a height of
40 cm. with charcoal-celite mixture. The column was washed with 1.5
l i t e r s of 0.1 N HCl to remove basic ash; acid was removed by washing ex
haustively with d i s t i l l e d water. The sugar and oligosaccharides were
eluted by passing two l i t e r s of each of water, 5 per cent, 15 per cent,
30 per cent, and 95.per cent ethanol through the column. The effluent

38
was collected In 100 ml. fractions and sugars were detected by the 3.5
din i t r o s a l i c y l i c acid method. Qualitative detection of various sugars
and oligosaccharides was done by paper chromatography. The water frac
tion contained glucose and varying amounts of maltose and trisaccharides.
The 5 percent, 15 per cent, 30 per cent and 95 per cent ethanol fractions
contained maltose, trisaccharides, a mixture of t r i - , tetra- and penta
saccharides and a mixture of hexasaccharides and high molecular weight
oligosaccharides, respectively.
N. Paper Chromatography
Whatman No. 1 f i l t e r paper was used during this investigation.
The containers used were wide mouth cabinet with screw lids (Research
Specialties Co., New Jersey, U.S.A.).
This technique was essentially used to purify malto-oligosaccharide.
Resolution of the sugar and oligosaccharide was achieved by multiple
ascent technique (18) using solvent (10) n- butanol-pyridine-water
(6:4:3 v/v). It was found that an ascent of 30 cm. was sufficient to
separate the f i r s t seven members of homologous series of malto-oligosac
charide.
The spots on the paper chromatograms were detected by spraying
with aniline phosphate reagent (6) and.heating at 115°C for 20 minutes.
These spots were used as markers for sectioning the remaining portions
of the chromatograms. The individual sugars were extracted from the
paper and concentrated in vacuo. They were further dried with acetone '
and washed with a small amount of n- butanol. The syrup was dissolved

39
in 50 ml. of water and freeze-dried. The freeze-dried fractions were
stored in a vacuum desiccator. *
0. Thin Layer Chromatography
Desaga thin layer chromatography apparatus (Canadian Laboratory
Supplies Limited, Vancouver, B.C.) was used. The coating material was
Kieselgel G ( S i l i c a gel) or Kieselgur G (Merck and Co.).
A suitable technique for the separation of oligosaccharides by
TLC was developed (20) using the solvent systems, isopropanol-t^O-ethyl-
acetate (2:1:2 v/v) or 1-propanol-nitromethane-water (5:2:3) as reported
by Huber ___1. (7).
P. Effect of Temperature
The effect of temperature on a-amylase activity under various
treatment was studied in a thermal gradient apparatus. The temperature
range could be adjusted from 20° to 80°C.
Q. Molecular Weight Determination
Molecular weight estimation was done by gel chromatography accord
ing to the method of Andrews (1), on a Sephadex column (2.5 cm x 50 cm).
R. Determination of Calcium Content
SP 90 Atomic Absorption Spectrophotometer (Unicam Instrument, Ltd.,
York St., Cambridge, England) was used to determine the amount of calcium
in a-amylase. A known amount of a-amylase was dialysed against

40
demineralized water for 48 hours. The amount of Calcium was determined
according to the instructions given in the SP 90 spectrophotometer
manual.
S. Amino Acid Analysis
The amino acid analysis of the purified a-amylase was done on a
Phoenix Amino Acid Micro-Analyser (Model M-7800). The enzyme preparation
was f i r s t dialysed against deionized water for 48 hours at 5°C. Hydrol
ysis was performed in hydrolysis tubes containing 1.0 mg.of protein in
1.0 ml. of deionized water and 1.0 ml. of concentrated HCl. The hydrol
ysis tubes were put in an oven set at 110°± 1°C for a period of 24 hours.
Hydrochloric acid was removed by repeated evaporation under reduced pres
sure by a rotary evaporator. The residue was dissolved in 1.0 ml. of
sodium citrate buffer pH 2.2 and 0.95 ml. was f i n a l l y applied to the
column for amino acid analysis. A Piez-Morris (19) accelerated buffer
system was used for elution of the amino acids.
*T. Immunochemical Techniques
The various immunochemical techniques as detailed below were used
to study a-amylase immunochemistry and homogeneity.
1. Production of Antibodies
Antibodies against the a-amylase of _. s u b t i l i s , A. oryzae, hog
pancreas and B. amylophilus . (isoenzyme 1) were prepared in rabbits. Each
of the four rabbits received, by subcutaneous injection over a course of
three weeks, a total of 4.0 mg. of protein in complete Freund's adjuvant.

41
The animals were bled two weeks after the last injection to provide
immune serum. The-antisera was inactivated by heating and i t was stored
in frozen state (15).
2. Determination of Enzymic Inhibition
The determination of the percentage of the inhibited activity of
a-amylase was done according to the method of McGeachin and Reynolds
(12). Homologous normal rabbit serum was included as a control in a l l
experiments. Control with homologous normal rabbit serum showed no i n
hibition of a-amylase activity. The 3 to 5 dinitro s a l i c y l i c acid
method was used to measure the a-amylase activity.
3. Immunodiffusion Characteristics
Antigenic relationships were studied by Immunoelectrophoresis and
double diffusion in agar gel (17).
4. Protein Determination in Antigen-Antibody Complex
Protein of antigen-antibody precipitate was determined according
to the method of Lowry et a l . , using bovine albumin as standard (11).

REFERENCES III
1. Andrews, P. 1965. The gel f i l t r a t i o n behaviour of proteins related to their molecular weights over a wide range. Biochem. J. 96:595.
2. Blackburn, T. H. and P. N. Hobson. 1962. Further studies on the isolation of proteolytic bacteria from the sheep rumen. J. Gen. Microbiol. 29,69.
3. . . 1968. Protease production by Bacteriodes amylophilus Strain H 18. J. .Gen. Microbiol. 53:27.
4. Davis, B. J . ( 1964. Disc electrophoresis II. Method and application to human serum proteins. Ann. N.Y. Acad. Sci. 121:404.
5. Fisher^ E.- and E. A. Stein. 1961. a-Amylase from human saliva. Biochem. Preparation. 8:27.
6. Frahn, J. L. and J. A. Mills. 1959. Paper ionophoresis of carbo-nydrates. I. Procedures, and results for four electrolytes. Aust. J. Chem. 12:65.
7. Huber, C. N., H. D. Scobell and E. E. Fisher. 1968. Thin layer chromatography of the malto-oligosaccharides and megalo-saccharides with mixed support and multiple irrigation. Anal. Chem. 40:207.
8. Hungate, R. E. 1950. The anaerobic mesophilic c e l l u l o l y t i c bacteria. Bact. Rev. 14:1.
9. Lesk, E. M. • 1969. Purification and characterization of Proteolytic enzymes from _. amylophilus Strain H-18. M.Sc- Thesis. University of British Columbia, Vancouver, B.C.
10. Jeanes, A., C. W. Wise and R. J. Dimlee. 1951. Improved techniques in paper chromatography of carbohydrates. Anal. Chem. 23:415.
11. Lowry,. 0. H. ,• N. J. Rosebrough, A. L. Farr and R. J. Randall. 1951. Protein measurement with the f o l i n protein reagent. J. Biol. Chem. 193:265.

43
12. McGeachin, R. J. and J. M. Reynolds. 1959. Differences i n mammalian amylases demonstrated by enzyme inhibition with specific antisera. J. Biol. Chem. 234:1456.
13. Miller, G. L. 1960. Micro-column chromatographic method for analysis of oligosaccharides. Anal. Biochem. 2:133.
14. Nakai, S. and F. Tsuchiya. 1961. Improved method of nitrogen determination by indophenol reaction. Jap. Analy. 10:387.
15. Nomura, M. and T. Wada. 1958. Studies on amylase formation by Bacillus-subtilis. V. Immunochemical studies of amylase produced by Bacillus-subtilis. J. Biochem. 45:629.
16. Nelson, N, 1944. A photometric adaptation of the Somogyi method for determination of glucose. J. Biol. Chem. 153:375.
17. Ouchterlony, 0. 1968. "The techniques of double diffusion in two dimensions, and Immunoelectrophoresis." In Handbook of Immunodiffusion and Immunoelectrophoresis. Ann. Arbor Science Publishers, Michigan, 48106.
18. Pazur, J. and D. French. 1952. The action of transglucosidase of Aspergillus oryzae onmaltose. J. Biol. Chem. 196:265.
19. Piez, K. A. and L. Morris. 1960. A modified procedure for the automatic analysis of amino acids. Anal. Bio. Chem. 1:187.
20. Rahman, Sh. Saif-Ur-, C. R. Krishnamurti and W. D. Kitts. 1968. Separation of Cello-saccharides by thin layer chromatography. J. Chromat. 38:400.
21. Robyt, J. F. and W. J. Whelan. 1968. "The a-amylase." In the Starch and i t s Derivatives. Ed. by J . A. Radley, 4th Ed., Chapman and Hall Ltd. , London, EC4.
22. whistler, R. L. and D. F. Durso. 1950; Chromatographic separation of sugars on charcoal. J. Amer. Chem. Soc. 72:677.
\

CHAPTER IV
RESULTS AND DISCUSSION
A. Characterization of a-Amylase from Bac teroides amylophilus Strain H-18
1. Production a'nd Purification of a-Amylase
a. Production of a-Amylase
Figure 1 shows that a-amylase was produced extracellularly during
the logarithmic, and stationary phase of growth by 13. amylophilus. These
results are in agreement with the findings of Lesk (20). In this regard
]3. amylophilus is similar to .B. stearothermophilus which starts producing
a-amylase during the logarithmic.period of growth (51) , but.it is di f f e r
ent from B. subtilis which produces a-amylase during the stationary
phase of growth (36).
The growth curve (Figure 1) of 13. amylophilus is characteristic
of the other rumen bacteria (3). Seventy-eight and twenty-two per. cent
of a-amylase was released into the medium during the logarithmic and
stationary phase of growth respectively (Figure 1). During the mid-
logarithmic growth phase the' amount of a-amylase liberated was linear
(Figure 2). Since a-amylase did not contain cysteine (see IV:A:2:f) and
its production began during the logarithmic phase.of growth, i t has the
characteristic features of other extracellular enzymes (41).
The effect of maltodextrins on a-amylase formation is shown in
Figure 3. When maltodextrins were added to the growing culture which

45
Figure 1 Growth curve and production of a-Amylase
from Bacteroides amylophilus Strain H-18.
Growth curve,
o—o a-Amylase activity released into
the culture Supernatant.
O - - O Percentage of maximum a-Amylase
activity in culture Supernatant.

a-Amylase activity in Supernatant (units/ml)
to o
to
46
LO O
LO Ul
Ul
Ul
o c
t-i to O
to Ui
LO O
Lo Ul
to O O
ON o
00 o Percentage of Maximum Activity
(32.5 units/ml) J I J
o o
— J
4> 00 O.D,660

47
Figure 2 Linear relationship between the production
of a-Amylase and growth of Bacteroides
amylophilus Strain H-18.
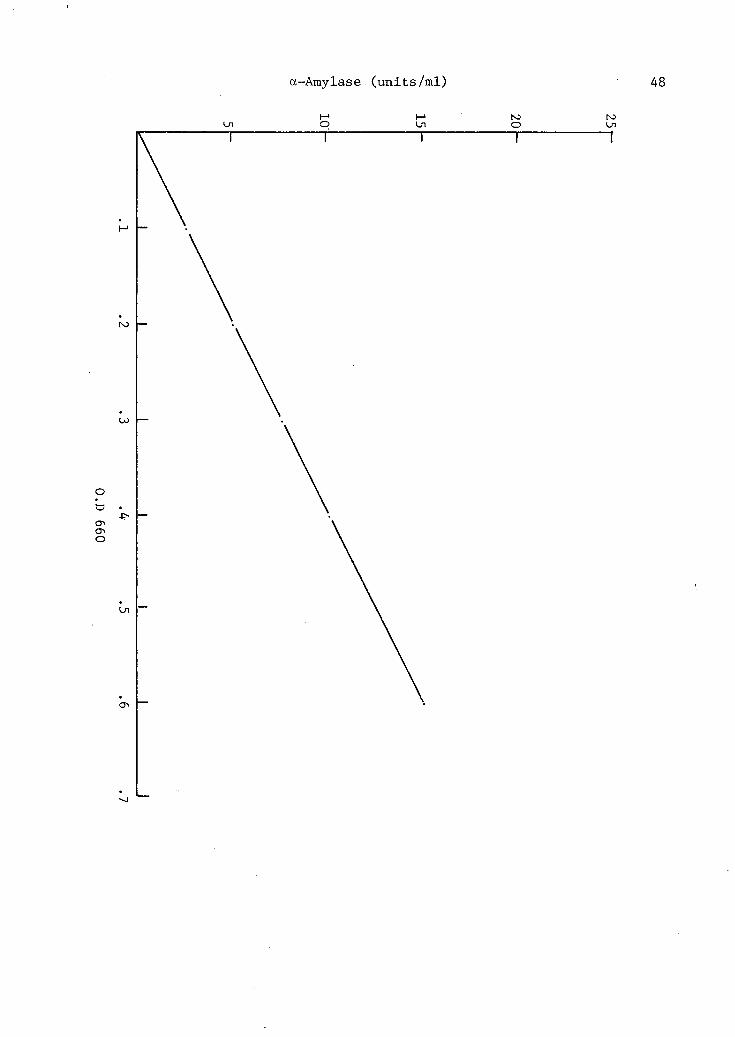
a -Amylase (units/ml)
t-1 1—1 NJ NJ Ul O Ul O Ul
O

49
Figure 3 Effect of Maltodextrin on the growth and
production of a-Amylase from Bacteroides
amylophilus Strain H-18.
Maltose
o—o Maltotriose
A - — A Maltotetraose
O—O Maltopentaose
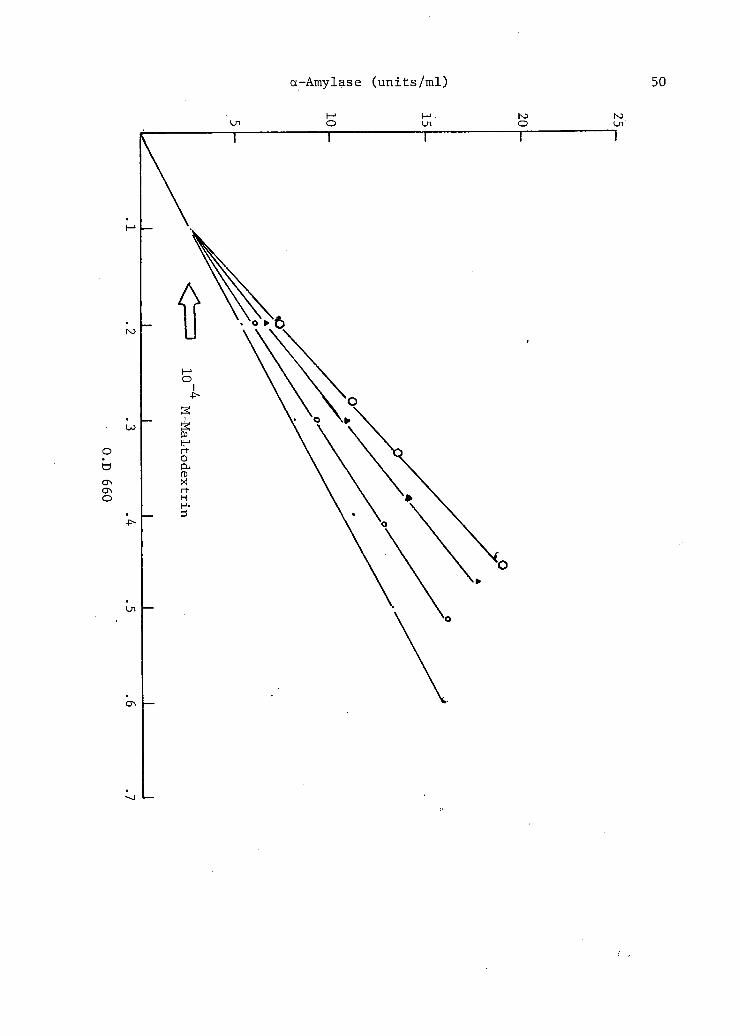
a-Amylase (units/ml) 50

had just entered the logarithmic phas.e, there was an increase in the
production of amylase. However when maltodextrins were replaced by glu
cose, sucrose, and cellobiose, there was no change in the amount of
amylase produced. These results, are essentially in agreement with the
findings of Blackburn (2) and Hungate (19) in which the a 1-4 linked
glucose polymers were the only carbohydrate substrates metabolized by
B_. amylophilus.
Under the experimental conditions used I5_. amylophilus cells
appeared to be permeable to maltose and maltotriose. After incubation
for 6 hours with maltodextrins, maltose was the only sugar detected extra
cellular ly by thin layer chromatography, and glucose was never found. It
was not investigated to find i f I3_. amylophilus takes up other malto-
oligosaccharides directly like Micrococcus Sp 40 (53) or hydrolyses them
to maltose or maltotriose. These results indicate that 1$. amylophilus
cells are permeable to maltose and maltotriose and therefore the maltose
and maltotriose up-take systems in this micro-organism are constitutive
like Micrococcus Sp 40 (53) but unlike that of E_. cold (52) which is
adaptive.
The results in Figure 1 indicated fluctuations of the production
of a-amylase into the. medium during the stationary phase of growth. This
suggested that there may be more than one a-amylase produced.by IS.
amylophilus. In order.to assess this hypothesis,.a-amylase was analysed
by disc electrophoresis and electrofocusing techniques.. Four active iso
enzymes of a-amylase (Figure 4) accompanied by 12 other bands of protein
were detected by disc electrophoresis. Four peaks of activity were also

52
Figure 4 Detection of 4 Isoenzyme of a-Amylase by
disc electrophoresis.
Illustration A. The- separation of 4 isoenzyme of a-Amylase by disc electrophoresis on acrylamide gel. The direction of migration was from the top of the figure.
Illustration B. The starch slide after i n cubation at 38°C for 15 minutes with acrylamide gel. After incubation the unhydrolysed starch was stained with Lugol's iodine. The clear bands indicating hydrolysis of starch was visible against the blue stain produced by the starch and iodine complex.

Stained acrylamide gel A
Amylase B
Amylase D
Amylase A
Amylase C
Stained slide with 1% starch B

54
detected by iso-electrofocusing (Figure 5). Isoelectric points as de
termined by electrofocusing were pH 3.7, 4.5, 5.9 and 8.0. The iso
enzymes were named 1, 2, 3 and 4 with respect to their increasing iso
electric points (Figure 5).
The reasons why this organism produces four a-amylases is not .
clear at the present time, but possibilities may be suggested. Since
they have different isoelectric points they should have different amino
acid compositions. It has been reported recently that different a-,
amylases have different a f f i n i t i e s towards various starches (5,28).
Evidently .B. amylophilus is a very versatile organism and may control
secretion of different isoenzymes depending upon the nature of starch in
the diet of the animal.
b. Purification of a-Amylase Isoenzyme 1
The methods used for purification of a-amylase isoenzyme 1 are
summarized in Figure 6 and the results are presented in Table III. The
stepwise purification process was conducted as follows:
Step 1 - A good quantity of a-amylase was obtained by growing the
culture for 23 hours. In order to measure the protein
content correctly, a sample of supernatant was dialysed
against 0.05 M phosphate buffer pH 7.0 to remove cysteine
and tryptose peptides (20).
Step 2 - The enzyme solution was concentrated by DEAE-Sephadex batch
operation. This step reduced the volume of the enzyme sol
ution from 29000 ml to 2540 ml, and gave a 5 fold purifica
tion.

55
Figure 5 Detection of 4 isoenzymes of a-Amylase (x—x)
by electrofocusing with superimposed pH curve
(.—.). The pi values of separated components
are obtained by taking the pH of the corres
ponding fraction at the maximum activity. The
pi value of the components were 3.7, 4.5, 5.9
and 8.2. The isoenzymes were named 1, 2, 3
and 4 with respect to their increasing iso
electric points. The figures above the enzyme
activity peaks give the pi values of a-Amylase
isoenzymes.

Relative activity (%)
56
< o M C B fD
pH Gradient

57
24 hours culture supernatant 29000.0 ml
DEAE-Sephadex batch operation. The enzyme was eluted from DEAE-Sephadex slurry with 1.0 M NaCl on a Buchner ;funnel
2540.0 ml
Dialysis overnight•against 0.05 M phosphate buffer, pH 7.0 2600.0 ml
DEAE-Sephadex column chromatography. Enzyme was eluted with a linear gradient of 0.2 M to 1.0 M NaCl in 0.05 M
phosphate buffer, pH 7.0
Fraction I 27-56 (Fig. 7)"* 538.0 ml
Fraction II 57-84 (Fig.7)* 500.0 ml
This indicates Fraction number in Figure VII .
Fraction III 85-131 (Fig.7)*
a-amylase Isoenzyme 1. Total volume collected, 840.0 ml. It was reduced by pressure dialysis to 40 ml.
Sephadex G-200 column chromatography. Total volume collected, 760.0 ml. It was reduced by pressure d i alysis to 45 ml.
Figure 6 Flow sheet of methods for the isolation
of a-amylase isoenzyme 1 from Bacteroides
amylophilus Strain H-18.

58
TABLE III
PURIFICATION OF a-AMYLASE ISOENZYME 1 FROM B. AMYLOPHILUS STRAIN H-18
Procedure
Volume ml
Concentration units/
ml
Total Units (xlO-3)
Protein mg/ml
Specific Activity (units/ mg protein
Yield %
P u r i f i cation
24 hour superr-natant 29000.0 4.0 116.00 0.30 13.3 100.00 1
DEAE-Sephadex batch operation 2540.0 40.0 101.60 0.60 66.6 87.5 5.00
Dialysis against P0 4
buffer 2600.00 35.0 91.00 0.50 70.0 78.4 5.26
DEAE-Sephadex Column Fraction No. I l l 840 75.0 65.52 0.15 550.00 56.00 41.35 .
Sephadex G-200 Column Fraction and Pressure Dialysis 45 1000 45.00 0.7 1428.5 38.8 107.4

59
Step 3 - The enzyme solution was dialysed against 0.05 M phosphate
buffer pH 7.0, to remove NaCl. The step due to some unknown
reasons decreased the enzymic activity. Since the amount
of protein present decreased from 0.60 mg to 0.50 mg per
ml, the decrease in enzymic activity may.be due to denatur-
ation of the a-amylase protein.
Step 4 - The enzymic preparation was fractionated by DEAE-Sephadex
chromatography using a linear sodium gradient (Figure 7).
At this stage the volume of enzyme solution was reduced to
840 ml and purification obtained was 41 fold. The solution
was further concentrated to 40.00 ml by pressure dialysis.
Step 5 - The enzyme solution obtained in Step 4 was subjected to gel
f i l t r a t i o n on Sephadex G-200. An elution profile of a-
amylase activity is shown in Figure 8. A l l of a-amylase
activity, was obtained in a single peak. At this stage the
recovery was 38.8 per cent and purification obtained was
107- fold. This enzyme preparation was free from protease
activity. The enzyme solution was concentrated by pressure
dialysis to 45.0 ml, freeze-dried and stored at 4°C for
further use.
Step 6 - The purity of this preparation was checked by disc electro
phoresis. A single band of.protein, was obtained indicating
homogeneity of a-amylase protein (Figure 9).
Step 7 - To further assess the homogeneity of this a-amylase, this
enzyme was subjected to isoelectrofocusing. A single sharp

60
Figure 7 Chromatography of Bacteroides amylophilus
Strain H-18 a-amylase on DEAE-Sephadex A-50.
Enzyme was eluted with a linear NaCl gradient
(0.2 to 0.75 M) in phosphate buffer (0.05 M,
pH 7.0).
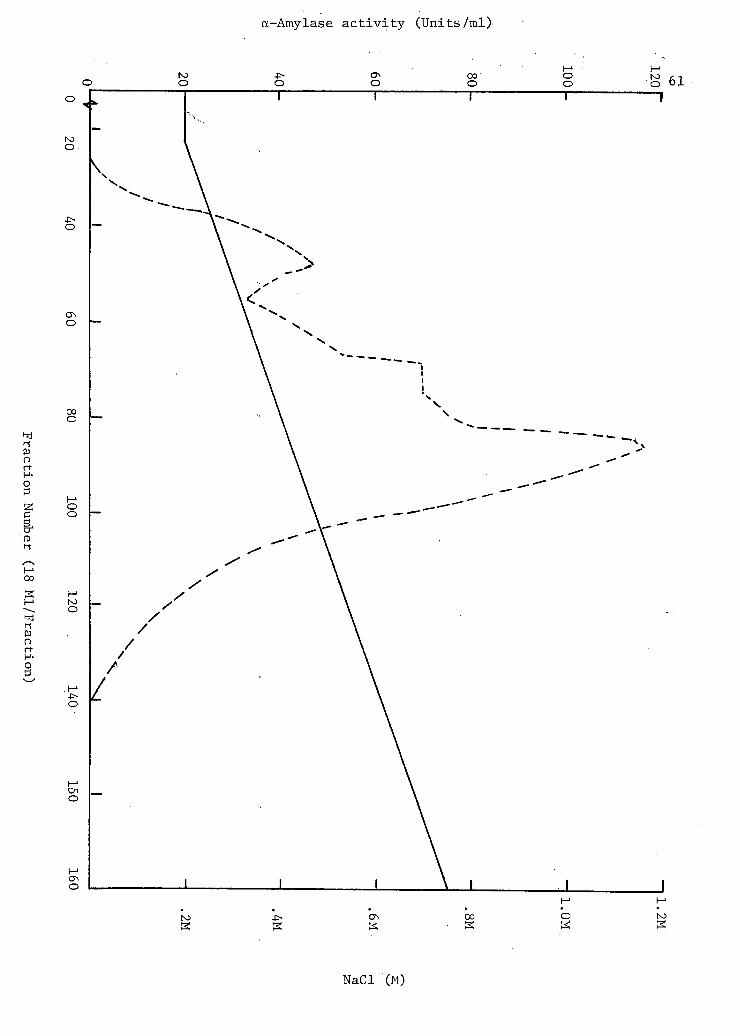
a-Amylase activity (Units/ml)
NaCl (M)

62
Figure 8 Chromatography of Bacteroides amylophilus
Strain H-18 a-amylase isoenzyme 1 on Seph
adex G-200. Enzyme was eluted with phos
phate- buffer (0.05 M, pH 7.0).

700,
cn vO
600
500
400
•300
200
100 / /
/
50 100 150 200 250 300 350 400 - z 1
450
Fraction Number (5.0 ml/Fraction)

64
Figure 9 Disc electrophoresis of a-Amylase
Isoenzyme 1.
Illustration A. Stained disc gel.
Illustration B. Densiometric tracing.

65
+
td

66
peak was obtained (Figure 10). Therefore-it is concluded
from these results that a-amylase isoenzyme 1 is a single
homogeneous protein.
2. Catalytic Properties of a-Amylase Isoenzyme 1
a. Determination of Type of Amylase
To determine whether the amylase in question is of the a or 3
type the enzymic digestion of starch and amylose was examined by thin
layer chromatography. Thin layer analysis revealed the. presence of high
molecular weight reducing dextrin and a series of malto-oligosaccharides
(see IV:B). These results indicate that the amylase isoenzyme 1 studied
is of the a-type.
b. Effect of pH on.a-Amylase Activity
The pH profiles of the activity of a-amylase are shown in Figure
11. The pH of the maximum enzymic activity was found to be 6.7 at 44°C.
The optimum pH values for a-amylases from other sources as reported in
the literature are in the acid region between 4.5 and 7.0. These re
sults are summarized in Table IV.
c. Effect of pH on Enzymic Stability
As seen in Figure 12, the a-amylase is stable in the pH range of
6.2 to 7.6. The stable pH i s narrow on both acidic and alkaline side.
The pH s t a b i l i t i e s of a-amylases from other sources are reviewed in
Table V for purposes of comparison.

67
Figure 10 Electrofocusing of a-Amylase Isoenzyme 1
(x—x) with superimposed pH gradient
(.—.)• The figure above the enzyme
activity peak gives the pi value of
a-Amylase.Isoenzyme 1.

pH Gradient

69
Figure 11 Optimum pH for hydrolyzing starch. The
enzymatic a c t i v i t y was determined at 44°C
for 15 minutes a f t e r incubation with
starch at respective pH values.

Relative Activity (%)

71
TABLE IV
SUMMARY OF THE OPTIMUM pH RANGE OF a-AMYLASE FROM VARIOUS SOURCES
Source of a-amylases Optimum References pH range
Porcine pancreas 6.8 4,31,34
Monkey small intestine 6.8 . 43
Human saliva 6.9 32,34
Aspergillus oryzae 4.8-5.8 12
Bacillus subtilis 6.0 29
Pseudomonas saccharophila 5.25-5.75 27
Bacillus stearothermophilus 5.0 6,7
4.5-6.5 37
Streptococcus bovis 4.6-6.1 17
Clostridium butyricum 5.5-6.5 17
B. polymyxa 6.2-7.5 42
Bacteroides amylophilus 6.7 S.U.R.

72
Figure 12 E f f e c t of pH on the s t a b i l i t y of a-amylase. The buffer s o l u t i o n used was 0.02 M T r i s -maleate (pH 5.8 to.8.6). To 1.0 ml of each of the above buffer solutions 0.2 ml of a 1 per cent s o l u t i o n of the enzyme ( i n d i s t i l l e d water) was added and the mixture was kept at 37°C for 24 hours. Af t e r adjusting the pH to 6.7, the f i n a l volume was made up to 4.0 ml. The enzymic a c t i v i t y was determined before and a f t e r treatment and the percentage of the a c t i v i t y which remained was calculated.
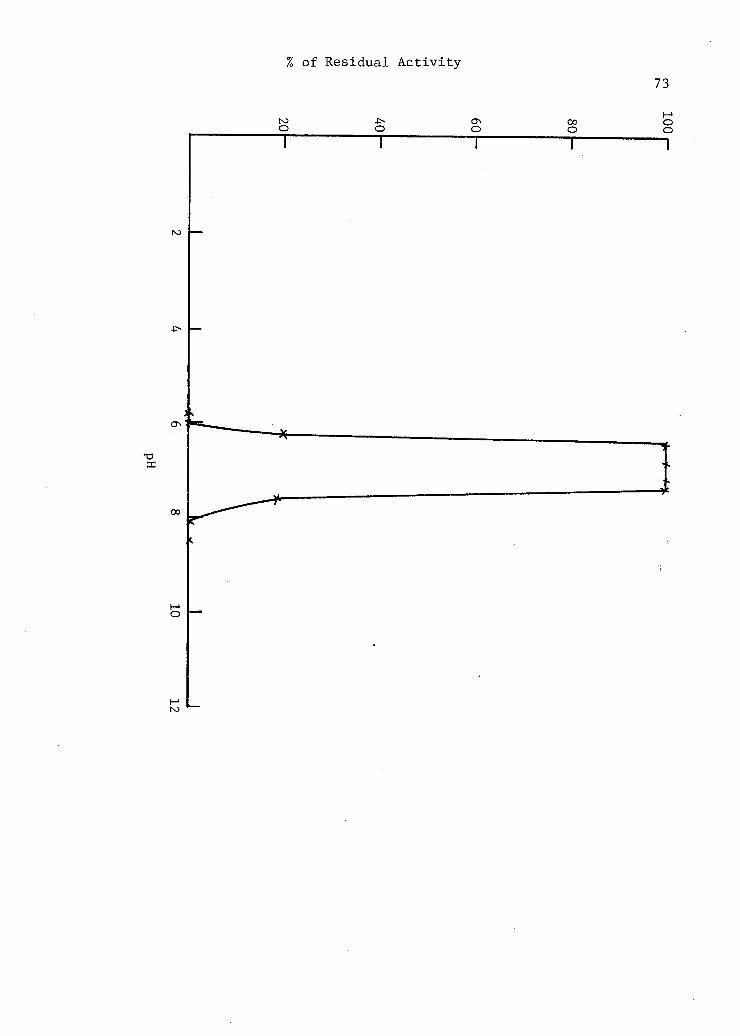
% of Residual Activity 73
O O ON O 00
o O O

74
TABLE V
SUMMARY OF THE OPTIMUM pH STABILITY RANGE FOR VARIOUS a-AMYLASES
Source of a-amylases pH sta b i l i t y range
References
Barley malt 4.9-9.1 34
Porcine pancreas 7.0-8.5 4,34
Human saliva 4.8-11 13,32,34
Aspergillus oryzae 5.5-8.5 10,49
Bacillus stearothermophilus 6-11 37
Pseudomonas saccharophila 4.5-8 27
Bacillus subtilis 4.8-8.5 30.46
Bacteroides amylophilus 6.2-7.6 S.U.R.

75
d. Effect of Temperature on a-Amylase Activity
The temperature profiles of the activity of a-amylase isoenzyme 1
are shown in Figure 13. Temperature•for the maximum activity is 44°C
as compared to a-amylases for other sources (Table VI).
e. Effect of Temperature on Enzymic Stability
Figure 14 (curve B) illustrates that a-amylase isoenzyme 1 re
tained 100 per cent of i t s original activity after heat treatment up to
42°C for 15 minutes. In this regard thermophilic a-amylase from B.
stearothermophilus retained 100 per cent of i t s original activity at
65°C for 15 minutes and the mesophilic a-amylase from B_. subtilis main
tained 100 per cent of i t s original activity at 43°C for 15 minutes (38).
It was found that a-amylase isoenzyme 1 contained 3 gram atoms of
calcium per mole of enzyme (see IV:A:2). a-Amylases of various origins
have been shown to contain calcium which is essential i n the catalytic
activity and stabilization of the enzyme molecule (12,18,22,50). There
fore the effect of calcium ions on the st a b i l i t y of a-amylase isoenzyme
1 was studied for comparative purposes. In the presence of 0.02 M
CaC^, the thermal s t a b i l i t y of a-amylase isoenzyme 1 increased from
42°C to 58°C (Figure 14, Curve A). This result indicates that the
property of a-amylase isoenzyme 1 is similar to that of other a-amylases.
Pretreatment of a-amylase isoenzyme 1 with 0.02 M EDTA decreased
the thermal s t a b i l i t y of the enzyme (Figure 14, Curve C). Increased
susceptibility of a-amylase to temperature was caused by the non
availability of calcium which was chelated by EDTA. Similar results have

76
Figure 13 Optimum temperature for hydrolysing starch.
The reaction mixture contained 1 ml of a
2 per cent solution of starch at pH 6.7 and
0.3 mg of enzyme in 0.02 M Tris-maleate
buffer (pH 6.7). The-reaction was carried
out at various temperatures for 10 minutes.

Relative Activity (%) 77

78
TABLE VI
OPTIMUM TEMPERATURE FOR VARIOUS a-AMYLASES
Source of a-amylase Optimum Temperature
References
Barley malt 35°C 3 4
Porcine pancreas 37°C 4,31,34
Human saliva 40 °C 1 3 , 3 2 , 3 3 , 3 4
Aspergillus oryzae 40°C 10,49
Bacillus subtilis 40°C 2 9 , 3 0 , 4 6
43°-58°C 37
Bacillus stearothermophilus 65°C . 6,7
65°-73°C 37
Streptocuccus bovis ,; 48°C 17
Clostridum butyricum 48°C 17
Bacteroides amylophilus 44°C S.U.R.

79
Figure 14 Thermal st a b i l i t y of a-amylase. Enzyme (0.03 mg/ml) was treated at various temperatures as indicated. After 15 minutes each solution was immediately cooled. The residual activity was determined and percentage of the activity which remained was calculated with respect to each treatment. The various treatments were' as follows:
Curve A. 0.02 M CaCl 2 in 0.02 M Tris-maleate buffer, pH 6.7.
Curve B. 0.02 M Tris-maleate buffer, pH 6.7.
Curve C. Enzyme was dialysed against 0.22 M EDTA in Tris-maleate buffer, pH 6.7, 20°C.

% of Residual Activity 80

81
been reported by EDTA treatment for human saliva, hog pancreas, B. sub-
t i l i s and A. oryzae a-amylases (50), _. stearothermophilus a-amylase
(38) and a-amylase from monkey small intestine (43 ) .
f. Amino Acid Determination
The amino acid analysis of a-amylase isoenzyme 1 was principally
done to determine the presence or absence of cysteine and cystine. The
amino acid analysis showed complete absence of cysteine and cystine.
Therefore disulphide linkages and sulphydryl groups are not involved in
maintaining the enzymic activity and tertiary structure of a-amylase
molecules. The absence of sulphydryl groups is in agreement with the
finding that p-chloromercuribenzoate did not inactivate the enzyme
(IV:A:2). It is also interesting to note that protease from B. amylo
philus H-18 is completely void of cysteine and cystine (20). In this
regard a-amylase isoenzyme 1 is comparable with _. subtilis a-amylase
which does not contain cystine (25). Bacterial a-amylase from B_. stear-
othermophilus does contain cysteine, but completely lacks tryptophan
(6 ) . Pollock (41) has reported the absence of disulphide linkages as a
characteristic feature of various bacterial exqenzymes.
g. Determination of Calcium
a-Amylases isolated from various sources have been reported to
contain a few atoms of firmly bound calcium (Table VII). When _. sub
t i l i s a-amylase was dialysed continuously against chelating agents, (a)
calcium was not removed completely, indicating i t was bound very firmly;
(b) and there was a reversible loss of enzymic activity when calcium

82
TABLE VII
CALCIUM CONTENTS OF VARIOUS a-AMYLASES
Source of a-amylase Amount of Calcium Present*
References
Human saliva 1-2 50
Hog pancreas 1-2 50
A. oryzae 2-3 50
B. subtilis 3.0 50
Bacteroides amylophilus 3.0 S.U.R.
The results are reported in. gram-atom per mole (50,000 g)
of enzyme.

83
contents were lowered below 1 gram atom per mole of enzyme, thus, indicat
ing a functional, role, of calcium (50) . The findings of Hsiu.et al_. • (18) ,
are similar for 13. subtilis a-amylase and human salivary a-amylase.
By spectrophotometric analysis i t was found that purified a-
amylase isoenzyme 1 from B. amylophilus contains 3 gram-atom of calcium
per mole of a-amylase. This value is in the range of reported values
for the other a-amylases (Table VII). When a-amylase isoenzyme 1 was
dialysed against 0.02 M EDTA (Figure 14), the enzyme retained 30 per
cent of i t s original activity and the calcium content at this stage was
0.8 gram atom per mole of a-amylase. When I3_; subtilis a-amylase was
dialysed against 0.01 M EDTA for 50 hours, the enzyme retained 40 per
cent of i t s original activity and calcium content after dialysis was 0.4
gram-atom per mole of enzyme (50) . These results indicate that while
isoenzyme 1 binds calcium more strongly than 13. subtilis a-amylase, i t
is less stable at low calcium levels than I3_. subtilis a-amylase.
h. Effect of Chemical Reagents on Enzymic Activity
(i) Effect of Urea on a-Amylase Activity
The effect of urea on a-amylase activity is shown in Figure 15.
The results (Curve A) indicate the remaining enzymic activity at d i f f e r
ent concentrations of urea. In 8.0 M urea the activity was completely
inhibited. After removal of urea by dialysis against Tris-maleate buffer
of pH 6.7, partial regeneration of the enzyme was obtained (Curve B,
Figure 15). After incubation in 8.0 M urea (pH 8.5) at 30°C for 30 min
utes, 13. subtilis a-amylase lost .40 per cent of i t s original activity,

84
Figure 15 Effect of urea on a-amylase activity.
Curve A. Various concentrations of urea as indicated were added to standard assay mixture and the percentage of the activity remaining was calculated.
Curve B. Solutions of enzymes (0.03 mg/ml), containing various concentrations of urea were kept at 37°C for 16 hours at pH 7.0. After dialysis in cold against 0.02 M Tris-maleate buffer (pH 6.7) for 24 hours, the. percentage of the remaining activity was calculated.


86
whereas B. stearothermophilus a-amylase lost 10 per cent of the original
activity (38). Imanishi et a l . (23) recovered about:80 per cent of the
original enzymic activity of _. subtilis a-amylase after treating with
8.0 M urea containing EDTA. Maximum recovery of a-amylase isoenzyme 1
was 68 per cent in 1.0 M urea. It appears from the results that a-amylase
isoenzyme 1 was more sensitive to urea under the experimental conditions
used. During the urea treatment the temperature was held at 37°C for 16
hours, which was higher than that used by other workers as noted above.
_. aeruginosa protease was completely inhibited in a solution of 8.0 M
urea, after treatment-at 37°C for 16 hours and reactivation could not be
achieved after a removal of urea (35) . Treatment of a-amylase from
monkey intestine with 5.0 M urea resulted in inactivation which was appar
ently irreversible (43). Fukushi e_ a l . (14) , also recovered 60 to 90
per cent of denatured B_. subtilis a-amylase activity at pH 8.5 in 8.0 M
urea at room temperature. These workers further reported that changes
in optical rotatory dispersion and spectral shift took place much more
rapidly, reaching almost f i n a l values immediately after the onset of
regeneration. Therefore, denatured protein after the removal of urea
rapidly resumed a three dimensional structure closely resembling that of
the native protein. This refolding is followed by a slower intra
molecular rearrangement to a characteristic native structure responsible
for a native and biologically active protein. Activity obtained was re
ported to be due to partial regeneration of enzyme molecules having the
same specific activity as that of native a-amylase (14). The failure of
regeneration may be due to the chemical binding of cyanate which is

87
present in urea with, amino groups to.yield carbamyl derivatives (8,44).
The other possible causes of irreversible inactivation of denatured
a-amylase reported by others may be due to incorrect refolding and inter-
molecular aggregation (23) , and partially due to proteolytic degradation
of the unfolded enzyme molecule by proteolytic contaminants (23,45). Re
generation of the enzymic activity also depends on the pH value and
ionic strength of the solution and concentration of enzyme (23).
( i i ) Effect of EDTA and Metallic Ions on a-Amylase Activity
As seen in Figure 16, treatment with EDTA reduced the enzymic
activity to 30 per cent of the original activity. By the addition of
various metals the enzymic activity was regenerated (Figure 16, B).
Treatment with calcium restored the enzymic activity completely while
magnesium reactivated the enzymic activity up to 90 per cent. It may be
noted that dialysis had no effect on the a-amylase isoenzyme activity.
The activity remained constant during the period of dialysis.
Treatment with EDTA removed the calcium from the various a-amy
lases, thus decreasing their activity (18,50). Addition of calcium re
sulted i n the restoration of enzymic activity (18,48). In this regard
the results presented in this thesis are similar to the findings of
other workers (18,38,46). The evidence has been presented that the ca l
cium atom is necessary in maintaining the catalytically active conforma
tion of the amylase molecule (18). The role of calcium becomes important
particularly in .B. subtilis a-amylase which lacks intramolecular disul
phide linkage and free sulfhydryl groups (1,25). In such cases the

88
Figure 16 E f f e c t of (A) EDTA and (B) metal ions a f t e r
EDTA treatment on r e a c t i v a t i o n of a-amylase.
The enzyme s o l u t i o n was dialysed against 0.02
M EDTA i n 0.02 M tris-maleate b u f f e r (pH 7.0)
at 5°C for 100 hours. EDTA was removed by
further d i a l y s i s for 24 hours against various
metal solutions as indicated i n 0.02 M t r i s -
maleate buffer (pH 6.7) and the enzyme a c t i v
i t y was determined.

% of the A c t i v i t y Remained 89
t o O O
ON
o oo o
t-c c /—\ bd g • X) C O tn Mi r-1 t o
M l fD S •~J (D fo • H r t H o fD M
I
o (S3 g
W o 1-3 >
Reactivation (%)
t o o o O N
o oo o c
"7
o • o o n 4>
r o
n • 01 o O 4S M g
ho
3 O n *~ M g
g •
CM O
M g ISJ

90
active conformation of protein may be maintained by certain intramolecu
lar non-covalent linkages rather than usual disulphide bridges (18).
Actually the non-existence of disulphide linkage appears to be character-,
i s t i c of various bacterial exoenzymes (41). The role.of calcium in
maintaining enzymic activity has also been suggested in several other
bacterial exoenzymes, viz. proteinase (16) and ribonuclease (18). In
this regard a-amylase isoenzyme 1 is an exo-enzyme, lacks disulphide
linkage and free sulfhydryl groups and requires calcium for i t s activity.
Imanishi (22) has reported that removal of calcium does not cause detec
table changes in protein conformation and suggested that calcium ions
may be located on.the surface of the protein. It appears, therefore^
that calcium forms a tight metal-chelate structure with the protein
molecule to maintain a proper configuration for biological activity
(18,50). Takagi and Iseumura (48) also indicated the role of calcium
ions in refolding the reduced taka-a-amylase A. As noted in Figure 16
(B), the treatment with cobalt, zinc and magnesium could not regenerate
the enzymic activity completely. The reason may be that certain metals
could not form-a correct metal chelate structure with protein, thus re
sulting in unstable or incomplete secondary and tertiary structures,
which did not have complete biological activity.
i . Functional Groups Determination
The results on the effect of reducing, oxidizing and SH-inactivat-
ing agents are presented in Table VIII. The a-amylase activity, remained
unaffected after the treatment with reducing agents, viz . , cysteine,
sodium cyanide, sodium thioglycolate, mercaptoethanol.

91
TABLE VIII
EFFECT OF REDUCING, OXIDIZING AND SH-INACTIVATING AGENTS ON a-AMYLASE ISOENZYME 1 ACTIVITY
Reagents Concentration* (M)
Residual Activity (%)
Cysteine 5 x 10"3 100.00
Sodium cyanide 5 x IO - 3 100.00
Sodium thioglycolate 5 x IO - 3 102.00
Mercaptoethanol 5. x IO - 3 100.00
p-Chloromercuribenzoate 5 x IO - 3 98.00
Monoiodoacetic acid 5 x IO - 3 92.0
Potassium permanganate IO"3 0
N-Bromosuccinimide 5 x IO - 3 0
Solutions of enzyme (6 units/ml) containing various concen-r trations of different reagents at pH 7.0 are kept for one hour at 37°C. The percentage of the activity which remained after the treatment was calculated.

92
Further the enzymic activity is not inactivated by specific SH-
inactivating agents such as p-chloromerciiribetizoate and monoiodoacetic
acid. These results indicate the noninvolvement of sulfhydryl groups in
enzymic reaction mechanisms, and are in agreement with the finding that
a-amylase isoenzyme 1 did not contain cystine. The slight inhibition may
be due to the reaction with methionine, serine or imidazole-group of
histidine. Since the inhibition observed was slight these.amino acids
may be located near the active center.
It is to be noted that enzymic activity was completely lost by ox
idizing agents viz., potassium permanganate and N-bromosuccinimide.
Although potassium permanganate is a non-specific oxidizing agent, N-
bromosuccinimide is more specific with controlled conditions in i t s
reaction with tryptophan. (39). Okada et^.a_. (39) used N-bromosuccinimide
to oxidize tryptophan and suggested that i t was involved ih the active
center of ._. subtilis ar-amylase. It has been reported by Sugae (47)
that B. subtilis a-amylase lost i t s activity when an azo-group.was intro
duced into the a-amylase molecule. He suggested that a peptide group in
the neighbourhood of a particular tyrosine residue which was modified by
the azo group was closely related with the activity of bacterial ct-.
amylase. In general, the active center includes, besides the catalytic
site, the grouping conforming to the substrate specific for the enzyme
(40); the two groups are sufficiently close to each.other. It is possible
that the active center of bacterial a-amylase may have two groups,.try
ptophan (39) and tyrosine (47). Yamato (54) indicated that.the trypto
phan and tyrosyl group a-amylase obtained from B, amyloliquefaciens

Fukumoto was essential for i t s enzymic activity. Ikenda (21) reported
that in case of A. oryzae, the phenolic group of tryosine was essential
for enzymic activity. The activity of a-amylase isoenzyme was lost with
the treatment.of N-bromosuccinimide (Table VIII). It is suggested from
this result that tryptophan is essential for the catalytic a b i l i t y of
a-amylase isoenzyme 1.
j . Determination of Molecular Weight
The estimated molecular weight for a-amylase isoenzyme i s 45,000.
The'molecular weight of various a-amylases are given in Table IX.
k. Determination of Isoelectric Point
Isoelectric point as determined by electrofocusing technique was
pH 3.7. Isoelectric points of other a-amylases are also in the acidic
region as indicated in Table X.

94
TABLE IX
MOLECULAR WEIGHT OF VARIOUS a-AMYLASES
Source Molecular Weight References
Barley malt 59,500 34
Porcine pancreas 45,000 9
Aspergillus oryzae 51,000 24
Bacillus subtilis 48,700 11
Bacillus saccharophila 15,600 26
Bacteroides amylophilus 45,000 S.U.R. •
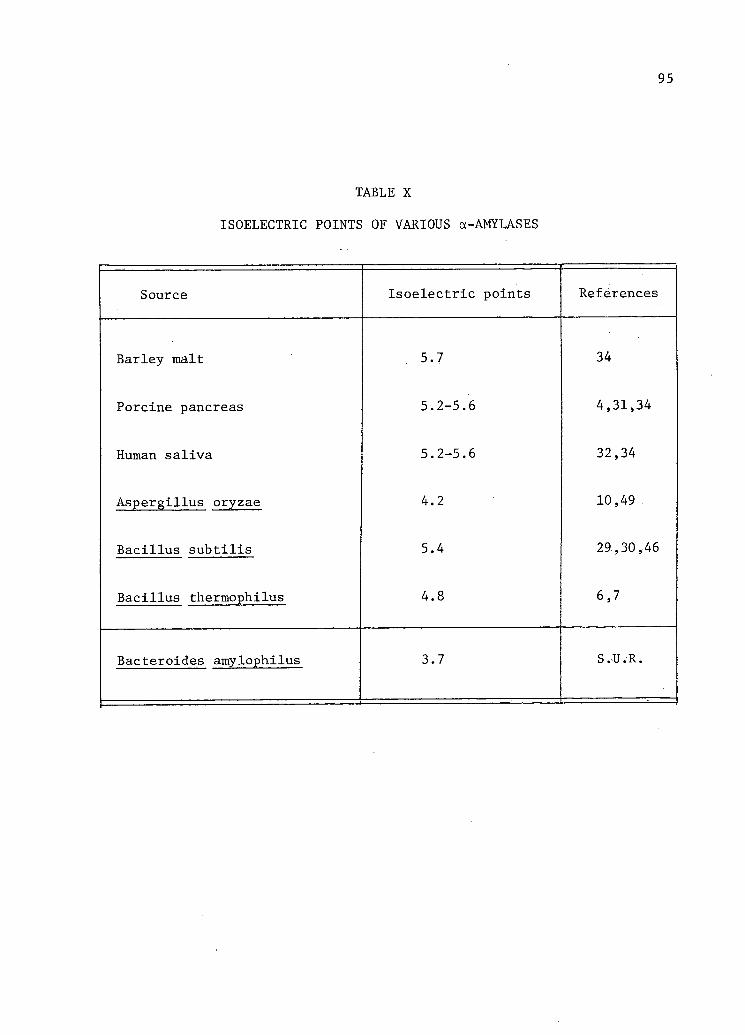
95
TABLE X
ISOELECTRIC POINTS OF VARIOUS a-AMYLASES
Source Isoelectric points References
Barley malt 5.7 34
Porcine pancreas 5.2-5.6 4,31,34
Human saliva 5.2-5.6 32,34
Aspergillus oryzae 4.2 10,49
Bacillus subtilis 5.4 29,30,46
Bacillus thermophilus 4.8 6,7
Bacteroides amylophilus 3.7 S.U.R.

REFERENCES IV:A
1. Akabori, S., Y. Okada, S. Fujiwara and K. J. Sugae. 1965. Studies on bacterial amylase. I. Amino acid composition of crystalline bacterial amylase from B_. subtilis N. J. Biochem. 43:741.
2. Blackburn, T. H. 1968; Protease production by Bacteroides amylophilus strain H-18. J. Gen. Microbiol. 51:27.
3. Bryant, M. P. and I. W. Robinson. 1961. Some nutritional requirements of genus Ruminococcus. Apply Microbiol. 9:91.
4. Caldwell, M. L., M. Adams, J. F. Kung and G. C. Toralballa. 1952. Crystalline pancreatic amylase. II. Improved method for i t s preparation from hog pancreas glands and additional studies of it s properties. J. Amer. Chem. Soc. 74:4033.
5. Clary, J. Jj G. E. Mitchell, Jr. and C O . L i t t l e . 1968. Action of bovine and ovine a-amylases on various starches. J. Nut. 95:469.
6. Campbell, L. L. and G. B. Manning. 1961. Thermostable a-amylase of Bacillus stearothermophilus. III. Amino acid composition. J. Biol. Chem. 236:2962.
7. and P. D. Cleveland. 1961. Thermostable a-amylase of Bacillus stearothermophilus. Crystallization and some general properties. J. Biol..Chem. 236:2952.
8. Cole, R. D. 1961. On the transformation of insulin in concentrated solution of urea. J. Biol. Chem. 236:2670.
9. Danielsson,. C.' E. 1947. Molecular weight of a-amylase; Nature. 160:899.
10. Fischer, F. H. and R. DeMontmollin. 1951. Purification et crysta l l i s a t i o n de 1'a-amylase d.'Aspergillus oryzae. Sur les enzymes amylolytiques. Helv. Chim. Acta. 34:1987. •
11. , W. N. Summerwell, J. M. Junge and E. A. Stein. 1958. Proceedings of Symposium VIII, IVth International Congress of Biochemistry, Vienna, Pergamon Press.

97
12. Fischer, E. H. and E. A. Stein. ' 1960. "a-Amylases." In the enzymes.-Ed. P. D. Boyer, H. Lardy, and K. Myrback. Vol. 4. Academic Press.
13. . 1964 a-Amylase from human saliva. Biochemical Preparation. 8:27.-
14. Fukushi, T., A. Imanishi and T. Isemura. 1968. Changes in enzymat i c activity and conformation during regeneration of native bacterial amylase from denatured form. J. Biochem. 63:409.
15. Hagihara, B. 1954. Crystalline bacterial amylase and proteinase. Ann. Rep. Sci. Osaka Univ. 2:35.
16. . 1960. "Bacterial and mold.proteases." In the enzyme. Ed. by P. D. Boyer, H. Lardy, and K. Myrback. Academic Press Inc. New York. Biochem. Biophys. Acta. 52:176.
17. Hobson, P. N. and M. Macpherson. 1952. Amylases of Clostridium butyricum and a streptococcus isolated from rumen of the sheep. J. Biochem. 52:671.
18. Hsin, J.-, E. H. Fisher and E. A. Stein. 1964. Alpha-amylase as calcium-metalloenzyme. II. Calcium and catalytic activity. Biochem. 3:61.
19. Hungate, R. E. - 1966. "The rumen bacteria." In the rumen and i t s microbes. By R. E. Hungate. Academic Press. New York and London.
20. Lesk, E. M. 1969. Purification and characterization of proteolytic enzymes from _. amylophilus strain H-18. M.Sc. Thesis. University of British Columbia, Vancouver, B.C.
21. Ikenda, T. 1959. Chemical modification on taka-amylase A. II. Phenylazobenzoylation of taka-amylase A. J. Biochem. 46:297.
22. Imanishi, A. 1966. Calcium binding by bacterial a-amylase. J. Biochem. 60:381.
23. , K. Kakiuchi and T. Isemura. 1963. Molecular stability and reversibility of denaturation of _. subtilis a-amylase. II. Regeneration of urea denatured enzyme by removal or dilution pf urea. J. Biochem. 54:89.
24. Isemura, T. andS. Fujita. 1957. Physicochemical studies on taka-amylase A. I. - Size and shape determination by the measurement of sedimentation, diffusion coefficient and viscosity. J. Biochem. 44:443.

98
25. Junge, J. M. , E. A. Stein, J. Neurath and E. H. Fischer. 1959. The amino acid composition of a-amylase from Bacillus s u b t i l i s . J. Biochem. 234:556.
26. Manning, G. B., L. L. Campbell and R. J. Foster. 1961. Thermostable a-amylase of Bacillus stearothermophilus. II. Physical properties and molecular weight. J. Biol. Chem. 236:2958.
27. Markovitz, A., H. P. Klein and E. H. Fisher. 1956. Purification, crystallization, and.properties of the a-amylase of Pseudomonas saccharophila. Biochem. Biophys. Acta. 19:267.
28. Meites, S. and S. Rogols. 1968. Serum amylases, isoenzymes, and pancreatitis. I. Effect of substrate variation. Clin. Chem. 14:1176.
29. Menzi, R., E. A. Stein and E. H. Fisher. 1957. Proprietes de deux • a-amylase de .B. s u b t i l i s . Sur les enzymes amylolytiques. Helv. Chim. Acta. 40:534.
30. Meyer, K. H., M. Fuld and P. Bernfeld. 1947. Purification et c r i s -t a l l i s a t i o n de 1'a-amylase de bacterie. Experentia. 3:411.
31. . , E. H. Fischer and P. Bernfeld. 1947. Sur les enzymes amylolytiques (1). L'isolement de l'a-amylase de pancreas. Helv. Chim. Acta. '30:64.
32. , , A. Stauband P. Bernfeld. 1948. Proprietes de l'a-amylase de salive humainine c r i s t a l l i s e e . Helv. Chim. Acta. 31:2165.
33. , , . 1948. Sur les enzymes amylolytiques. Isolement et c r i s t a l l i s a t i o n de l'a-amylase.de salive humaine. Helv. Chim. Acta. 31:2158.
34. . 1952. The'past and present of starch chemistry. Experentia. 8:405.
35. Morihara, K. 1963. Pseudomonas aeruginosa proteinase. I. Purification and general properties. Biochem. Biophys. Acta. 73:113.
36. Nomura, M., B. Maruo and S.Akabori. 1956. Studies on amylase formation by Bacillus s u b t i l i s . I. Effect of high concentration of polyethylene glycol on amylase formation by Bacillus s u b t i l i s . J. Biochem. 43:143.
37. Ogasaharaj K. , A. Imanishi and T. Isemura. 19.70. Studies orither-mophilic a-amylase from Bacillus stearothermophilus. I. Some' general and physico-chemical properties of thermophilic a-amylase. J. Biochem. 67:65.

99
38. Ogasahara; K. , A. Imanishi and T. Isemura. 1970. Studies on thermophilic a-amylase.from Bacillus stearothermophilus. II. Thermal st a b i l i t y of thermophilic a-amylase. J. Biochem. 67:77.
39. Okada, Y. , K. Onoue, S.Nakashima and Y.Yamamura. 1963. Studies on the enzyme-antienzyme system. II. N-bromosuccinimide modified bacterial a-amylase. J. Biochem. 54:477.
40. Pechere, J. F. and H. Neurath. 1957. "Proteolytic enzyme." In Symposium on protein structure.. Ed. by A. Neuberger. International Union of Pure and Applied Chemistry. Paris.
41. Pollock, M. R. 1962. "Exoenzyme." In the Bacteria. Ed. by I. C. Gunsalus and R. Y. Stainer. Vol. 4. New York and London. Academic Press, Inc.
42. Rose, D. 1948. The amylase of Bacillus polymyxa. Arch. Biochem. 16:349.
43. Seetharam, B., N. Swaminathan and A. N. Radhakrishna. 1969. Purification and properties of a-amylase from monkey small intestine. Indi. J. Biochem. 6:51.
44. Stark, G. R., W. H. Stein and S. Moore. 1960. Reaction of the cyanate present in aqueous urea with amino acids and protein. J. Biol. Chem. 235:3177.
45. Stein, E.A., and E. H. Fisher. 1958. The resistance of a-amylase towards proteolytic attack. J. Biol. Chem. 232:867.
46. . 1961. a-Amylase from Bacillus s u b t i l i s . Biochemical preparation. 8:34.
47. Sugae, K. I960; . Studies on bacterial a-amylase. V. Chemical modification of-bacterial amylase and coupling of taka-amylase A with p. sulfobenzene-dizonium-chloride. J. Biochem. 48:790.
48. Takagi, T. and T. Isemura. 1965. Necessity of calcium for the regeneration of reduced Taka-amylase A. J. Biochem. 57:89.
49. Underkofler, L. A. and D. K. Roy. 1951. Crystallization of fungal alpha-amylase and limit destrinase. Cereal Chem. . 28:18.
50. Vallee, B. L., E. A. Stein, W. N. Summerwell and E. H. Fisher. 1950. Metal content of a-amylases of various origins. J. Biol. Chem. 234:2901.
51. Welker, N. E. and L. L. Campbell. 1963. Effect of carbon source on formation of a-amylase by Bacillus stearothermophilus. J. Bact. 86:861.

100
52. Wiesmeyer, H. and M. Cohn. 1960. The characterization of the pathway of maltose u t i l i z a t i o n by Escherichia Coli. III. A description of the concentrating mechanism. Biochem. Biophys. Acta. 39:440.
53. Williams,. P. J. and J. J. McDonald. 1966. Permeability of a.micro-coccal c e l l to maltose and some related sugars. J. Canad. Microbiol. 12:1213.
54. Yamamoto. T. 1955. Studies on sensitive groups of.crystalline bacterial a-amylase. Bull. Agr. Chem. Soc. Japan. 19:121.
55. . 1955. Studies on the a-amylase destroying enzyme. Part I. Occurrence and some properties of the enzyme. Bull. Agr. Chem. Soc. Japan. 19:22.

101
B. The Action Pattern of a-Amylase Isoenzyme 1
In this section the mode of action of _. amylophilus a-amylase
isoenzyme 1 was examined with two substrates, amylose and.soluble starch.
The products of digestion were examined qualitatively by thin layer
chromatography. The importance of the iodine-staining characteristics
is reported in terms of possible mechanisms for a-amylase action. The
differences and similarities of a-amylase isoenzyme 1.action are discussed
with respect to other a-amylases.
The progress of hydrolysis of amylose and starch is shown
chromatographically in Figures 17 and 18. Figure 17 represents hydrol
ysis beyond the achroic point and this was reached by about thirty min
utes. In Figure 18 hydrolysis was also extended beyond the achroic
point. The reference samples in each case were obtained by partial acid
hydrolysis of amylose. : Due to the following two observations, the spots
were regarded as linear oligosaccharides of maltose series:
I. The experimental spots from enzymatic digests had a corres
ponding Rf with regard to reference sample (3).
II. The introduction of branch point (4) retards the mobility i n
such a way that linear oligosaccharides are separated from
branched oligosaccharides of equal D.P. For example, a
branched dextrin of four glucose units f a l l s at a point inter
mediate in distance between maltopentaose arid maltotetraose
These experimental results suggest that significant amounts of
branched oligosaccharides were not present at the achroic point. The

102
Figure 17 Thin layer analysis of the digestion of Amylose.by a-Amylase Isoenzyme 1. The f i r s t column contained reference compounds, obtained by the acid hydrolysis of Amylose. The remaining columns contained.the products of the progressive hydrolysis of Amylose taken from the digestion mixture at various time intervals as indicated.on the chromatograms. The chromato-gram was obtained by the method of Rahman et a l . (11) with a solvent system, n-propanol ethyl acetate-water (6:1:3,v/v).
C. Control of pure sample.
DP. Degree of polymerization.

« • — , • •
5 10 15 20 25 30 3 5 u 0
Tine (Minute)

104
Figure 18 Thin layer analysis of the digestion of starch by an a-Amylase Isoenzyme 1. The f i r s t column contained reference compounds, obtained by the acid hydrolysis of Amylose. The remaining columns contained the products of the progressive hydrolysis of Amylose taken from the digestion mixture at various time intervals as indicated on the chromatogram. The chromato-gram was obtained by the method of Rahman et a l . (11) and solvent system was 1-propanol--nitromethane-water (5:2:3) as reported by Huber ejt.al. (8).
C. Control of pure sample.
DP. Degree of polymerization.


106
appearance of thin layer chromatogram developed with the digestion pro
ducts of amylose and starch were the same, and could not be differeritia-
ted from one another. It is suggested from these results that the pro
duction of higher malto-dextrin is a characteristic pattern of B.
amylophilus a-amylase isoenzyme 1.
Figures 17 and 18 show the complete maltodextrin spectrum from
D.P. 2 to 14. The early products in the digest are maltodextrin mixtures
from D.P. 5 to 14. There are, f i r s t of a l l , traces of maltopentaose,
maltohexaose and maltoheptaose, and predominance of higher saccharides.
Maltose appeared only after twenty-five minutes of digestion and i t s
amount increased steadily as.enzymatic reaction progressed. Maltodextrin
of D.P. 12 to 14 started disappearing after the achroic point, and corres
pondingly the intensity of maltodextrin to D.P. 7 to 11 started decreasing.
No glucose could be detected during the course of enzymatic reaction.
This can be contrasted and compared with the characteristic maltodextrin
spectrum produced by other a-amylases. B_. subtilis a-amylase produces
mainly maltotriose, maltotetraose and maltoheptaose during early stages
of amylose, amylopectin and starch hydrolysis (12,14) and near the
achroic point maltose, maltotriose and very.small quantities of malto
triose, maltopentaose, maltohexaose and maltoheptaose.(2). During a pro
gressive course salivary amylase produced increased amounts of maltose,
maltotriose-and trace amounts of maltopentaose, maltohexaose.and higher
oligosaccharides (2). Main end products are maltose, maltotriose and a
small amount of glucose. B_. polymyxa a-amylase produces maltose mainly,
however there are trace amounts of glucose and maltotriose formed as

107
well (13). Hanrahn and Caldwell (7) reported that with high concentra
tion of enzyme and.if a sufficient time of hydrolysis were allowed,
Aspergillus oryzae a-amylase degraded amylose completely into glucose
and maltose. The early stage products of the action of porcine pan
creatic a-amylase on starch and amylose were found to be maltose and
maltotriose and i t has been suggested that mode of action of this enzyme
is similar to that of human salivary a-amylase (14). The results of
enzymatic hydrolysis shown in Figures 17 and 18 indicate that only
certain series of maltodextrin could be separated by thin layer chroma
tography, as reducing spots were present at the point of application on
the chromatogram.
In the present, investigation the changes in the polysaccharide-
iodine spectrum were studied during hydrolysis of starch and amylose by
a-amylase to understand the mode of attack of substrate.- Since the blue
colour of iodine-starch complex depends upon the degree of polymeriza
tion of straight chains of the polymer (16), a decrease in absorption
during progressive enzymatic hydrolysis has been taken as a decrease in
the average degree of polymerization of the substrate. During the enzy
matic hydrolysis of amylose and starch, a decline in the absorption was
recorded. In each case the extent of this decline was comparable to the
results reported by Swanson (16), for salivary a-amylase. It is suggested
that in a qualitative way the dextrinizing action of JB; amylophilus a-
amylase isoenzyme 1 is comparable to salivary a-amylase;
It i s generally regarded that the mode,of attack of various a-
amylases on substrate is random (2,10), and the enzyme-product complex

108
dissociates i t s e l f after single catalytic events. It is also assumed
that a-amylases have equal preference for a l l a-1, 4 bonds, except those
near the branch point and adjacent to the two ends, which are known to
be more resistant to enzymatic attack. Such an observation is supported
by a rapid decrease in iodine colour and viscosity (6), but with the
availability of chromatographic results as reported above i t was apparent
that such a group of highly characteristic malto-oligosaccharides and
malto-dextrins could not be produced through random attack (1). This
problem has been resolved (5). The i n i t i a l attack of a-amylase with the
substrate was described as cleaving of one bond or a multiple of bonds
in the proximity of the f i r s t . The specific nature of this multiple
attack on substrate could be determined by enzymes with the production of
characteristic series of malto-saccharides (1,3). The action pattern of
_. amylophilus a-amylase isoenzyme 1 is in line with this hypothesis,
and the concept of multiple attack at the site of encounter explains the
data presented for this a-amylase.
Kung et a l . (9) reported that variation in the curves relating
blue value to the increase in the reducing values were due to the d i f f e r
ent a-amylases degrading amylose in different chain lengths. But chroma^
tographic results have shown that.different a-amylases produced different
types of digestion products and these maltodextrins are characteristic
of individual enzymes, as mentioned above. Robyt and French (15) reported
that porcine pancreatic a-amylase and human salivary a-amylase digest
produced very identical results but their blue value-reducing value
curves were very different. These observations are not supported by the

109
explanation offered by Kung et a l . (9). . Robyt and French (15) explained
these differences by multiple attack mechanism and also calculated the
degree of multiple attack for different a-amylases by determining the
ratio of the reducing value of the oligosaccharide fraction to that of
the polysaccharide fraction. In the case of JB. amylophilus a-amylase
isoenzyme 1 the,degree of multiple attack as calculated by the method of
Robyt and French (15) is 2 at optimum pH 6.7 and temperature 44°C
(Table XI). Under the.optimal condition of pH and temperature, porcine
pancreatic a-amylase had a degree of multiple attack of 6, three times
that of human salivary and Aspergillus oryzae a-amylase (15).
r

110
TABLE XI
ESTIMATION OF THE DEGREE OF MULTIPLE ATTACK BY B. AMYLOPHILUS ISOENZYME 1*
%BV1 3 5 r
90.0 38 12.0 115.75 3.1
81.0 78.2 26.0 66.70 3.0
72.0 111.0 36.0 59.50 3.08
63.0 123.0 40.0 49.50 . 3.07
3.06**
From the ratio of the total reducing value to the reducing value of 67% ethanol polysaccharide precipitate.
" * , * • • • • Average r determination.
Degree of multiple attack was determined by the method of Robyt and French (15). The definitions used are the same as those used by these authors.
1. BV = (At/AO) x 100 where Ao and At are the absorbancies (620 mu) of the iodine complex of the digest at zerotime and at t times of hydrolysis.
2. RV_ = Total reducing value is expressed as mg of apparent maltose/ml digest.
3. RVp = Reducing value of 67% ethanol precipitate in terms of mg of apparent maltose/ml of digest.
4. Average degree of polymerization of 67% ethanol precipitate determined by the quotient: [Total carbohydrate (mg/ml)/apparent maltose (mg/ml)] x 2.:
5. Quotient of the total reducing value divided by the reducing value of the 67% ethanol precipitate (RV^RVp) .
6. Degree of multiple attack (r-1) = (3.06-1) = 2.06.

REFERENCES IV:B
1. Abdullah, M. , D. French and J. F. Robyt. 1966. Multiple attack by a-amylase. " Arch. Biochem. Biophys. 114:595.
2. Bird, R. and R. H. Hopkins. 1952. The action of some a-amylases on amylose. Biochem. J. 56:80.
3. Dube, S. R. and P. Nordin. 1962. The action pattern of sorghum a-amylase. Arch. Biochem. Biophys. 99:105.
4. French, D. and G. M. Wild. Correlation of carbohydrate structure with papergram mobility. J. Amer. Chem. Soc. 75:2612.
5. . 1957. Recent developments and theoretical aspects of amylase action. Bakers Digest. 31:24.
6. Greenwood, C. T., A. W. Macgregor and E. Milne. 1965. a-Amylosis of starch. Staerke. 17:219.
7. Hanrahan, V. M. and M. L. Caldwell. 1953. A study of the action of Taka-amylase. J. Amer. Chem. Soc. 75:2191.
8. Huber, C. N., H. D. Scobell and E. E. Fisher. 1968. Thin layer chromatography of malto-oligasaccharides and megalasaccharides with mixed support and multiple irrigation. Anal. Chem. 40:207.
9. Kung, J. T., V. M. Hanrahan and M. L. Caldwell. 1953. A comparison of the action of several alpha amylases upon a linear fraction from corn starch. J. Amer. Chem. Soc.. 75:5548.
10. Myrback, K. 1948. Products of the enzymic degradation of starch and glycogen. Advance in Carbohydrate Chem. 3:251.
11. Rahman, Sh. Saif-^ur-, C. R. Krishnamurti and W. D. Kitts. 1968. Separation of Cello-saccharides by thin layer chromatography. J. Chromat. 38:400.
12. Robyt, J. and D. French. .1963. Action pattern and specificity of an amylase from Bacillus s u b t i l i s . Arch. Biochem. Biophys. 100:451.

112
13. Robyt, J. and D. French. 1964. Purification and action pattern of an amylase from Bacillus polymyxa. Arch. Biochem. Biophys. 104:338.
14. . 1962. Action pattern of some alpha-type amylases. Ph.D. thesis. Iowa State University.
15. . and D. French. 1967. Multiple attack hypothesis of ct-amylase action. Action of porcine, pancreatic, human salivary and Aspergillus oryzae. Arch. Biochem. Biophys. 122:8.
16. Swanson, M. A. 1948. Studies on the structure of polysaccharides IV. Relation of iodine colour to structure. J. Biol. Chem. 172:825.

113
C. Immunochemical Studies on a-Amylase•Isoenzyme 1
1. Inhibition of Enzymic Activity by Antibody
The results of enzymic inhibition by antibody are essentially
similar to those of bacterial and mold a-amylases (5,13). Normal control
serum did not inhibit the enzymic activity unlike Nomura and Wada (5)
who obtained slight inhibition of the enzymic activity. The time course
study of the inhibition of the a-amylase activity by antibody disclosed
that neutralisation was complete in 1 hour.
Various concentrations of antibody were added to the constant
amount of a-amylase and incubated at 37°C. After 1 hour, the enzymic
activity was determined and percentage of remaining activity was calcu
lated. The results are presented in Figure 19. The curve was linear
u n t i l 84 per cent of the enzymatic activity was neutralized, that i s , the
quantity of enzyme neutralized was proportional to that antibody added
in the region of antigen excess. A small amount of residual activity (16
per cent) remained in the presence of excess antibody.
This might be explained either by reversible dissociation of anti
body-enzyme complex or that the complex exhibits amylase activity (13).
Cinader (3) has also reported.that enzyme-antibody complexes themselves
have some residual activity.~" Cinader and Lafferty (2) have.established
the presence of three types of antibodies to biologically active antigen.
These antibodies can affect the activity of enzyme in three different
ways: antibody may combine and inhibit; antibody may combine with anti
gen but not inhibit i t s activity; and antibody may combine, not inhibit,
and may interfere in the combination with inhibiting antibody.

114
Figure 19 Neutralisation curve of a-amylase isoenzyme -
1 with antiserum.
One ml of enzyme solution (8.0 units) was
mixed with 1.0 ml of antiserum containing
various amounts of original antiserum,
a-amylase activity was determined after the
incubation of 1 hour at 37°C.


116
2. Ouchterlony Double-Diffusion Analysis
Results of double diffusion in agar gel are shown in Figure 20.
Antisera against f_. amylophilus a-amylase isoenzyme 1 were found to be
monospecific, indicating that i t was composed of a single antigenic
component. There was no precipitation line observed between B_. amylo
philus a-amylase isoenzyme 1 and antisera to amylases of various origins
(Figure 20). Similarly the reaction between an antiserum to _. amylo
philus a-amylase isoenzyme 1 and amylases of various origins (B. su b t i l i s ,
A. oryzae and hog pancreas) was negative. This experiment demonstrated
that _. amylophilus a-amylase isoenzyme carried antigenic determinants
which were distinct from those present on the a-amylase of hog pancreas,
_B. subtilis and A. oryzae.
Antibodies were also successfully obtained against hog pancreatic
a-amylase. Although the amylases formed precipitates with their respec
tive antisera, none of these reacted against each other, indicating that
these proteins are antigenically distinct. These results are in agree
ment with the findings of.Nomura and Wada (5).
3. Immunoelectrophoretic Analysis
Immunoelectrophoretic analysis, with antisera to _B. amylophilus
a-amylase isoenzyme 1, revealed the presence of only a single antigenic
component (Figure 21). It was also noted that _. subtilis a-amylase
moved to the cathode side and A. oryzae to the anode side, confirming
the results of Wada (13).

117
Figure 20 A diagramatic representation of immunodiffusion precipitation reaction between _. amylophilus a-amylase isoenzyme 1 and anti-amylase and anti-amylase antiserum of various origins.
Well 1 - _. amylophilus a-amylase isoenzyme 1.
Well 2 - Anti-_. amylophilus a-amylase isoenzyme 1 antiserum.
Well 3 - Anti-B. subtilis a-amylase antiserum.
Well 4 - Anti-A. aspergillus a-amylase antiserum.
Well 5 - Anti-hog pancreatic a-amylase antiserum.

118

119
Figure 21 A diagramatic representation of Immunoelectrophoresis of 13. amylophilus a-amylase isoenzyme 1.
Medium: 2 per cent agar in pH 8.2, 1=0.033 Veronal buffer.
Antigen: 5 mg/ml added in central well.
Electrophoresis was carried out at 5 mA per slide for 2 hours. Bromophenol blue was used as tracking dye. After electrophoresis antiserum was added in troughs 1 and 2. Precipitation line between a-amylase. isoenzyme 1 and i t s antiserum was observed.

120
4 -
I

121
4. Quantitative Precipitation Analysis
The precipitation reaction was studied quantitatively and a typi
cal curve with one equivalence point was obtained. These results are
presented in Table XII and Figure 22. The molar ratio range of antibody
to antigen in precipitates at equivalence and in the antibody excess re
gion were found to be between 1.8 and 2.31. Other reported values re
garding the molar ratio between 13. subtilis a-amylase and i t s antibody
are 2.16 (7); 1.9 (12) and for Taka-amylase A 2.58 (8), 2.6, 2.8, 4.0
(10). At the point of equivalence tests of the supernatant solution indi
cated well defined zones of antibody excess, equivalence and antigen
excess (Table XII). The quantity of precipitate decreased in the anti
gen excess zone.
During Immunoelectrophoresis at pH 8.2 and agar gel diffusion, a
single precipitate line formed between I3_. amylophilus a-amylase isoenzyme
1 and i t s antibody. It is suggested from the above reported results
that the a-amylase isoenzyme 1 preparation contained a single antigenic
component.
5. Effect of N -Bromosuccinimide and Urea on Antigenicity
Treatment of a-amylase with N-bromosuccinimide completely des
troyed - enzymatic activity (Table XIII). This result was probably due to
the oxidation of tryptophan residues at the catalytic site of the enzyme
molecule (6). Immunochemical analysis was performed to detect molecular
configuration differences between native and NBS-treated enzyme. The
quantitative precipitation curve with the NBS-modified enzyme preparation
was found to be comparable to that given by native enzyme in the area of
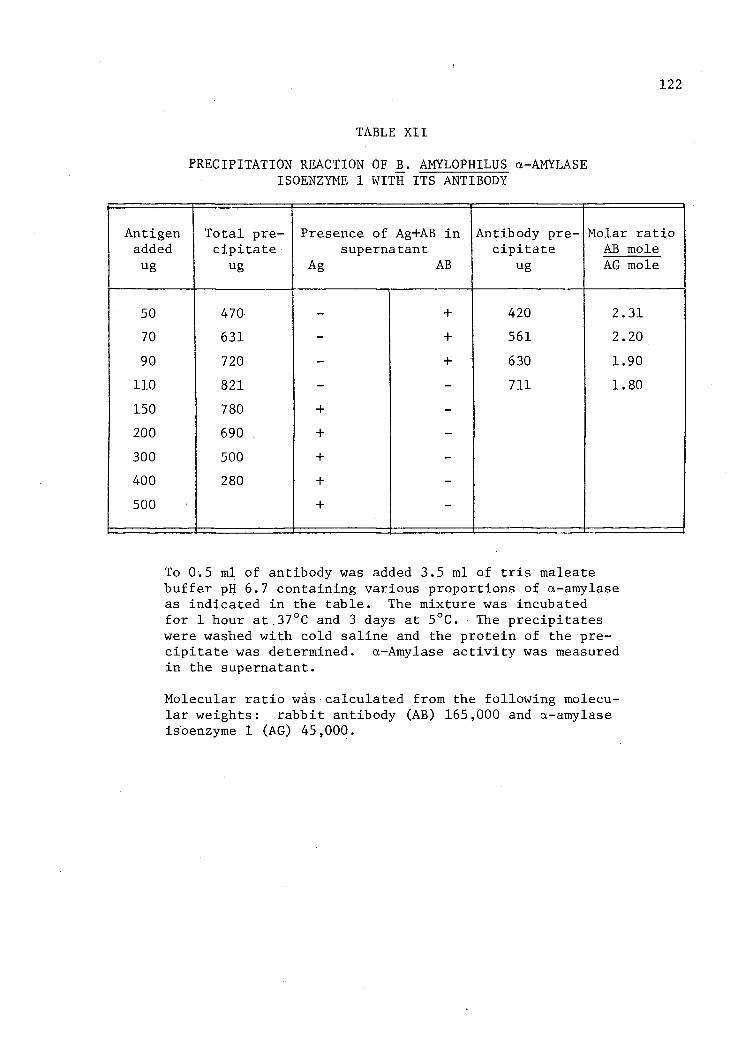
122
TABLE XII
PRECIPITATION REACTION OF B. AMYLOPHILUS a-AMYLASE ISOENZYME 1 WITH ITS ANTIBODY
Antigen added ug
Total precipitate
ug
Presence of Ag+AB in supernatant
Ag AB
Antibody precipitate
ug
Molar ratio AB mole AG mole
50 470 - + 420 2.31 70 631 - + 561 2.20 90 720 - + 630 1.90
110 821 - - 711 1.80 150 780 + -200 690 + -300 500 + -400 280 + -500 + -
To 0.5 ml of antibody was added 3.5 ml of t r i s maleate buffer pH 6.7 containing various proportions of a-amylase as indicated in the table. The mixture was incubated for 1 hour at 37°C and 3 days at 5°C. The precipitates were washed with cold saline and the protein of the precipitate was determined. a-Amylase activity was measured in the supernatant.
Molecular ratio was calculated from the following molecular weights: rabbit antibody (AB) 165,000 and a-amylase isoenzyme 1 (AG) 45,000.

123
Figure 22 Precipitation curve of a-amylase isoenzyme 1
with i t s antibody. The condition of the
reaction as described in Table XII.


125
TABLE XIII
EFFECT OF N-BROMOSUCCINIMIDE ON ACTIVITY OF ct-AMYLASE ISOENZYME 1
Reagent Treatment Length of treatment Residual activity Molar ratio minutes % NBS/enzyme
NBS 6 15 8
NBS 7 15 0
a-Amylase (1 mg/ml) in 0.05 M sodium acetate buffer
(pH 6.0) was treated with NBS. The reaction was termi
nated with Na2S0^ solution and enzymic activity was
determined.

126
antibody excess and in the equivalence zone (Figure 23). It is suggested
that catalytic and antigen sites are distinct. Similar are the results
with B. subtilis a-amylase (6), A. oryzae a-amylase (11) and bovine ribo-
nuclease (1).
Treatment of a-amylase with urea (8 M) completely inactivated the
catalytic a b i l i t y of the enzyme (Table XIV) . Urea treated (8 M) ct-
amylase did not form any precipitates with the antibody. Therefore both
enzymic activity and antigenic.characters were destroyed completely. The
effect of urea is due to an unfolding of protein molecules bringing
about the loss of biological activity (9). Comparable immunochemical re
sults have been reported with urea treated phage lysozyme (4). It i s
interesting to note that both phage lysozyme (4) and .B. amylophilus a-
amylase isoenzyme 1 do not have disulphide linkage, whether this com
plete loss of antigenic character is related to the absence of disulphide
bonds is not known (10). It is interesting that Aspergillus a-amylase,
which contains disulphide linkages, has been shown to retain part of i t s
immunochemical reactivity following urea treatment (10). The retention
of a certain proportion of i t s antigenic reactivity may be related to
disulphide linkages which probably stabilize certain sections of the
enzyme molecule making i t resistant to urea denaturation (10).
6. The Neutralisation of Amylase-Antiamylase System by Starch '
a-Amylase was incubated with antibody for 50 minutes at 37°C.
After this, 5 per cent starch solution was added in the incubation mix
ture and amylase activity was determined by the iodine reaction. The

127
Figure 23 Precipitation curve of N-bromosuccinimide
treated and native a-amylase with i t s anti
body. Reaction condition as reported in
Table XII.
x—x. NBS treated enzyme.
Native enzyme.

1000
100 200 300 400 500
Antigen added (ug)

TABLE XIV
EFFECT OF UREA ON. ACTIVITY OF a-AMYLASE ISOENZYME 1
Reagent Treatment M
Length of treatment H
Residual activity
Urea
Urea
6
8
24
24
1.0
0
a-Amylase (1 mg/ml) was treated with urea at 37°C.
After treatment the sample was dialysed against 0.02 M
tri s buffer (pH 6.7) for 24 hours and the enzymic
activity was determined.

130
percentage of starch hydrolysis was decreased due to the presence of
antibody (Figure 24, curve C), and the rate was constant for 150 minutes
at least. In the next experiment enzyme was added to 5 per cent starch
solution and after 10 minutes of incubation at 37°C, the same amount of
antibody was added. As shown in Figure 24 (curve B) the percentage of
starch hydrolysis was lower than the control curve A (Figure 24). As
the starch was hydrolysed, i t s effects as a protective agent diminished,
and the inhibitory effect of antibody appeared (Figure 24, curve B).
Therefore, i t appears that the products of starch digestion are not as
effective as starch to neutralize the amylase-antiamylase system. Wada
and Nomura (12), and Onoue et a l . (7) have also reported that starch did
interfere with amylase-antiamylase system. Antibody interference may be
due to steric hindrance or due to conformational change in the enzyme
molecule (2). In the present experiment i t is possible that both factors
may be involved, as previous experiments with NBS indicated that anti
genic and catalytic sites of enzyme molecules are different.

131
Figure 24 Inhibitory effect of starch on a-amylase isoenzyme 1 and anti-amylase system.
Curve A. 4.0 ml of 5 per cent starch + 3.0 ml of 0.02 M t r i s buffer pH 6.7 +1.0 ml of enzyme (8.0 units).
Curve B. 4.0 ml of 5 per cent starch + 2.0 ml of t r i s buffer + 1.0 ml of enzyme (8.0 units). After incubation for 10 minutes at 37°C, 1.0 ml of 20 per cent diluted immune serum was added.
Curve C. a-Amylase (8.0 units/ml) was incubated at 37°C with 1.0 ml of 20 per cent immune serum. After 50 minutes, substrate solution (4.0 ml of 5 per cent starch +2.0 ml of t r i s buffer) was added to the incubation mixture.
ml of reaction mixture was removed at i n tervals and 5 ml of 0.1 N HCl was added to stop the reaction. The remaining starch was determined by measuring the blue colour at 660 mu•in iodine reaction.

50 100 150 Time (minutes)

REFERENCES IV:C
1. Brown, Ray K. 1963. Immunological studies of bovine ribonuclease derivatives. Ann. N.Y. Acad. Sci. 103:754.
2. Cinader, B. and K. J. Lafferty. 1963. Antibody as inhibitor of ribonuclease: the role of steric hindrance, aggregate formation, and specificity. Ann. N.Y. Acad. Sci. 103:653.
3. . 1967. "Antibodies to enzymes - a discussion of the mechanism of inhibition and activation." In the proceedings of the 2nd meeting of the Federation of European Biochemical Societies, Vienna, April 21-24, 1965. Ed. by B. Cinader. Pergamon Press, Toronto.
4. Merigan, Thomas C. and William J. Dreyer.. 1963. Studies on the antigenic combining sites in Bacteriophage lysozymes. Ann. N.Y. Acad. Sci. 103:765.
5. Nomura, M. and T. Wada. 1958. Studies on amylase formation by Bacillus s u b t i l i s . V. Immunochemical studies of amylase produced by Bacillus s u b t i l i s . J. Biochem. 45:629.
6. Onoue, K., Y. Okada and Y. Yamamura. 1968. Modification of bact e r i a l a-amylase with N-Bromosuccinimide. J. Biochem. 51:443.
7. , , S. Nakashima, K. Shimada and Y. Yamamura. 1963; Studies on enzyme-antienzyme system. I. Immunochemical studies on Bacillus subtilis a-amylase; J. Biochem. 53:472.
8. Okada, Y., Y. Matsuoka, T. Yagura, T. Ikenka and Y. Yamamura. 1964. Immunochemical study of taka-amylase A and.phenylazo-benzoyl taka-amylase A. J. Biochem. 55:446.
9. Schachman, H. K. 1963. Considerations on the tertiary structure of protein., Cold Spring Harbour Symp. Quant. Biol., 28:409.
10. Sirishinha, S. and Peter Z. Allen. 1965. Immunochemical studies on a-amylase. I. Effect of denaturing agents and proteolytic enzymes on the immunochemical reactivity of a-amylase from A. oryzae.- Arch. Biochem. Biophys. 112:137.
11. 1965. Immunochemical studies on a-amylase.

134
II. Examination of immunochemical and enzymatic activities of native and modified a-amylase from Aspergillus oryzae.• Arch. Biochem. Biophys. 112:149.
12. Wada, T. and M. Nomura. 1958. An immunochemical study of microbial amylase (1). J. Biochem. 45:639.
13. , . 1959. An immunochemical study of microbial amylase II. J. Biochem. 46:329.

CHAPTER V
CONCLUSIONS
Bacteroides amylophilus strain H-18 produces four isoenzymes of
a-amylase, as detected by disc electrophoresis and electrofocusing. Iso
electric points as determined by electrofocusing were pH 3.7, 4.5, 5.9
and 8.0. Isoenzymes were named 1, 2, 3 and 4 with respect to their i n
creasing isoelectric points. a-Amylase isoenzyme 1 was purified and
some of i t s physico-chemical properties were examined and summarized in
a subsequent section.
A. General Properties
1. The optimum pH was 6.7 and exhibited a.narrow stable range of 6.2 to
7.6.
2. Its optimum temperature was 44°C and thermal stability range was 0
to 42°C. Since optimum temperature was not within the stability
range, i t was possible that substrate (starch) protects the enzyme
from heat, denaturation during assay.
3. The thermal s t a b i l i t y of the enzyme was affected, by EDTA treatment,
due to non-availability of chelated calcium. Treatment by calcium
protected the enzyme from heat denaturation.•
4. Oxidizing agents but not reducing agents and SH-reagents inactivated
the enzymic activity. Enzyme was susceptible to urea treatment.
5. ' Amino acid analysis indicated the absence of cysteine, therefore,
disulphide linkages are not involved in maintaining the tertiary

136
structure. • Tryptophan appeared to be essential for catalytic
activity.
6. The estimated molecular weight was 45,000.
7; a-Amylase isoenzyme 1 was found to contain 3 gram atoms of calcium
per mole. Various other metals tested could not replace the calcium
in regenerating the maximum activity.
8. The isoelectric point was found to be pH 3.7.
B. Action Pattern
1. The products of enzymatic hydrolysis of starch and amylose were mal
tohexaose, maltoheptaose, maltoctaose, maltonanaose and maltodecaose
at the achroic point and sometime after i t , as revealed by thin lay
er chromatography.
2. Maltose was the smallest disaccharide detected. Since glucose was
never found in-these experiments, i t appeared that a-amylase isoen
zyme 1 does not hydrolyse maltotriose and maltose.
3. The degree of multiple attack under the optimum conditions of tem
perature and pH was 2, as calculated by the ratio of the reducing
value of the oligosaccharide fraction to that of the polysaccharide
fraction.
C. Immunochemical Properties
1. Antisera against a-amylase isoenzyme l.was found to be monospecific
and a small amount of residual activity remained in the presence of
excess of antibodies.

137
2. The inhibitory effect of starch on the amylase-antiamylase system
was demonstrated.
3. The effect of anti-amylase (isoenzyme 1) globulin on amylase of var
ious origins was studied by Ouchterlony double-diffusion, and the
results indicated that antigenic determinants of a-amylase isoenzyme
1 were distinct from those present on a-amylase of hog pancreas,
Bacillus subtilis and Aspergillus oryzae.
4. Immunoelectrophoretic analysis revealed the presence of only a
single antigenic component.
5. The precipitation reaction was studied quantitatively and a typical
curve with one equivalence point was obtained.
6. The molar ratio ranges of antibody to antigen in precipitates at
equivalence and in the antibody excess zone were found to be between
1.8 and 2.31.
7. N-Bromosuccinimide treated a-amylase.(isoenzyme 1) had no enzymic
activity, but exhibited comparable immunochemical behaviour to native
enzyme. It is possible that antigenic and catalytic sites are dis
tinct.
8. Urea treatment destroyed the a b i l i t y of the enzyme to precipitate
with i t s specific antibody.