proteolytic activity of rumen microorganisms of cattle and buffalo
TRANSCRIPT

Proteolytic Activity of Rumen Microorganisms of Cattle and Buffalo
U. R. MEHRA, U, CHETAL, B. P. SINGH, and Y. R. SAXENA 1
Division of Animal Nutrition Indian Veterinary Research Institute
Izatnagar -- 243122 (U.P.), India
ABSTRACT
Microbial proteolytic activity of ru- men of large farm animals, buffalo and cattle, were studied. Enzymic proteolytic activity of microbes of buffalo rumen was much less than that of cattle; however, the profile was reversed for chemical constituents. The proteolysis proceeded progressively with time, but at all inter- vals rumen liquor of buffalo was much less proteolytic than that of cattle. The pattern was similar for different sub- strates. The significance of the "sparing effect" of rumen microorganisms of buffalo on degradation of dietary protein is discussed.
INTRODUCTION
As early as 1938, Sym (37) reported in the bovine rumen a highly active proteinase (pro- ---h,tieLc~. s _.~ enzy:r~e) attributable to microbial origin. Hunt and Moore (14) observed Flavo- bacterium sp., isolated from cow's tureen were proteolytic. Systematic studies of micro- bial proteolytic activity in ruminants have been confined essentially to sheep( l , 2, 3, 4, 5, 6, 7, 19, 38), and information is scanty on large farm animals. Furthermore, there are no reports of proteolytic activity of the rumen microbes of water buffalo (Bos bubalis), a native of Asia.
We investigated the enzymic proteolytic activity (proteinases) of the rumen micro- organisms of cattle (Bos indicus) and buffalo (Bos bubalis) under similar nutritional regime and studied the relative quantitative differ-
Received October 17, 1977. 1For inquiries: Indian Institute of Sugarcane
Research, Lucknow (U.P.) -- 226002, India. 2 Composition of mineral mixture (on dry matter
basis): calcium 28%, phosphorus 12%, iron .75%, iodine (as KI) .13%, copper .13%, manganese .15%, cobalt .026%, fluorine .04% (15).
ence(s) in the pattern of their proteolytic activity. Also, some of the chemical constitu- ents of the rumen microbes of these animals were compared.
MATERIALS AND METHODS
Studies on chemical constituents (Table 1), proteolytic activity (Table 2, Group A), and typical properties of proteolytic enzyme(s) of the rumen microorganisms were of one each of adult male cattle (Bos indicus) and Murrah buffalo (Bos bubalis) weighing 390 kg and 466 kg. The findings of the proteolytic activity, however, were substantiated further by ex- tending these studies to three male cattle weighing 280, 305, and 330 kg and to three male Murrah buffaloes weighing 248, 254, and 316 kg (Table 2, Group B). The animals fitted with rumen fistulas were under similar dietary and environmental conditions and were fed a concentrate mixture according to Sen's bulletin (34). The concentrate mixture consisted of groundnut cake, crushed maize, and wheat bran in equal parts, 2% mineral m i x t u r J , and 1% sodium chloride. Vitamin A (20,000 IU each animal) also was provided daily. Wheat straw (bboosa) was fed ad libitum as the sole source of roughage.
Processing of Rumen Liquor
Wheat straw and water were removed 16 h prior to rumen liquor. The rumen liquor was collected at 1030 h in a sterilized container, in ice, .5 h prior to feeding the animals. Maxi- mum care was taken to obtain a representative sample by withdrawing the rumen liquor from different places in the rumen. The rumen liquor was strained through eight-fold of muslin cloth and subsequently centrifuged to sediment microorganisms in a Sorvall refrigerated cen- trifuge (Model RC2-V, USA) at 4 C at 34,858 x g (17,000 rpm) for 20 rain. The supernatant was discarded and the pellets were washed
1978 J Dairy Sci 61:1573--i578 1573
1574 MEHRA ET AL.
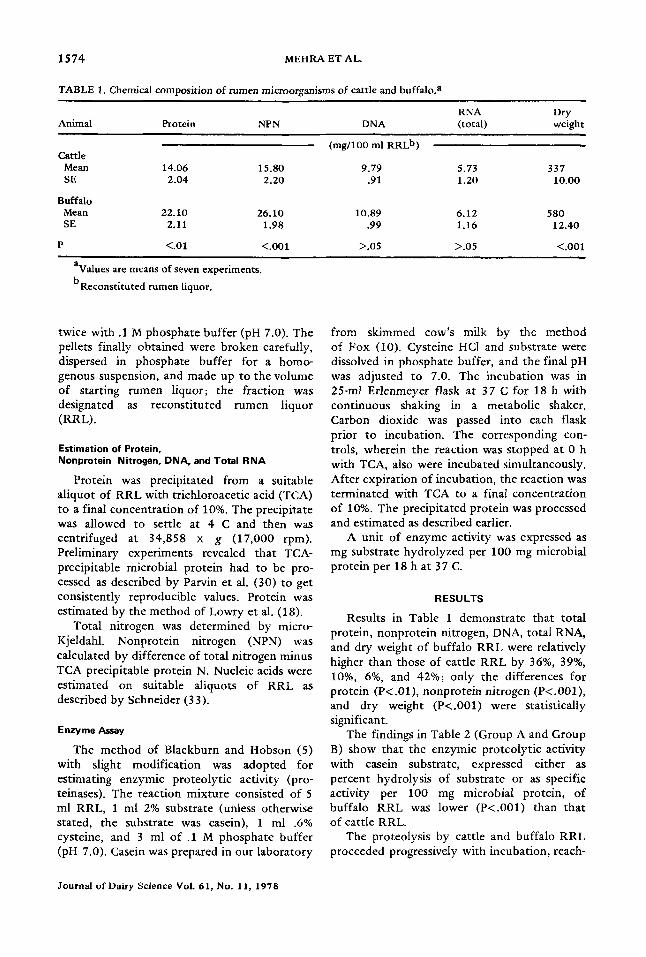
TABLE 1. Chemical composition of rumen microorganisms of cattle and buffalo, a
RNA Dry Animal Protein NPN DNA (total) weight
(mg/100 ml RRL b) Cattle
Mean 14.06 15.80 9.79 5.73 337 SE 2.04 2.20 .91 1.20 10.00
Buffalo Mean 22.10 26.10 10.89 6.12 580 SE 2.11 1.98 .99 1.16 12.40
P <.01 <.001 >.05 >.05 <.001
avalues are means of seven experiments. b Reconstituted rumen liquor.
twice with .1 M phosphate buffer (pH 7.0). The pallets finally obtained were broken carefully, dispersed in phosphate buffer for a homo- genous suspension, and made up to the volume of starting rumen liquor; the fraction was designated as reconsti tuted rumen liquor (RRL).
Estimation of Protein, Nonprotein Nitrogen, DNA, and Total RNA
Protein was precipitated from a suitable aliquot of RRL with trichloroacetic acid (TCA) to a final concentration of 10%. The precipitate was allowed to settle at 4 C and then was centrifuged at 34,858 × g (17,000 rpm). Preliminary experiments revealed that TCA- precipitable microbial protein had to be pro- cessed as described by Parvin et al. (30) to get consistently reproducible values. Protein was estimated by the method of Lowry et al. (18).
Total nitrogen was determined by micro- Kjeldahl. Nonprotein nitrogen (NPN) was calculated by difference of total nitrogen minus TCA precipitable protein N. Nucleic acids were estimated on suitable aliquots of RRL as described by Schneider (3 3).
Enzyme Assay
The method of Blackburn and Hobson (5) with slight modification was adopted for estimating enzymic proteolytic activity (pro- teinases). The reaction mixture consisted of 5 ml RRL, 1 ml 2% substrate (unless otherwise stated, the substrate was casein), 1 ml .6% cysteine, and 3 ml of .1 M phosphate buffer (pH 7.0). Casein was prepared in our laboratory
from skimmed cow's milk by the method of Fox (10). Cysteine HC1 and substrate were dissolved in phosphate buffer, and the final pH was adjusted to 7.0. The incubation was in 25-ml Erlenmeyer flask at 37 C for 18 h with continuous shaking in a metabolic shaker. Carbon dioxide was passed into each flask prior to incubation. The corresponding con- trois, wherein the reaction was stopped at 0 h with TCA, also were incubated simultaneously. After expiration of incubation, the reaction was terminated with TCA to a final concentrat ion of 10%. The precipitated protein was processed and estimated as described earlier.
A unit of enzyme activity was expressed as mg substrate hydrolyzed per 100 mg microbial protein per 18 h at 37 C.
RESULTS
Results in Table 1 demonstrate that total protein, nonprotein nitrogen, DNA, total RNA, and dry weight of buffalo RRL were relatively higher than those of cattle RRL by 36%, 39%, 10%, 6%, and 42%; only the differences for protein (P<.01), nonprotein nitrogen (P<.001), and dry weight (P<.001) were statistically significant.
The findings in Table 2 (Group A and Group B) show that the enzymic proteolyt ic activity with casein substrate, expressed either as percent hydrolysis of substrate or as specific activity per 100 mg microbial protein, of buffalo RRL was lower (P<.001) than that of cattle RRL.
The proteolysis by cattle and buffalo RRL proceeded progressively with incubation, reach-
Journal of Dairy Science Vol. 61, No. 11, 1978

MICROBIAL ACTIVITY IN RUMINANTS 1575
0
e~
0
2
0
~4
<
0
U o . ~
~.~
m.N ,.,
rs
o
~-~
e~
~ 0
< ' .~
t~ e~ U
ing a maximum of 20 h when about 75% of casein was hydrolyzed by cattle RRL whereas for buffalo RRL the hydrolysis was only about 40%. At all intervals, the buffalo RRL was always less proteolyt ic than cattle RRL (Fig. 1).
Data in Table 3 point out that, both with cattle and buffalo RRL, the degree of proteoly- sis was highest with casein as a substrate and lowest with bovine serum albumin. However, irrespective of substrate, the proteolyt ic activ- i ty of buffalo RRL was always lower than that of cattle RRL.
Furthermore, results in Table 4 show that anaerobic incubation stimulated proteolysis of cattle and buffalo RRL by 40% and 29%.
DISCUSSION
The biological value of dietary protein in ruminants depends precisely upon the following three factors: a) the dietary protein that evades degradation in the rumen, b) the quan- t i ty and quality of microbial protein synthe- sized from dietary and endogenous nitrogen, and c) the amount of net protein flowing into abomasum. Conclusions of several reviewers indicate that as little as 40% or as much as 80% of the dietary protein might be degraded in the rumen (13, 16, 17, 32, 35, 36). In addition, approximately 30% of the microbial protein produced in the rumen also is degraded therein (16, 22, 23, 24). Bacterial proteinases are constitutive enzymes which do not appear to be subject to metabolic and dietary control. Addit ional dietary urea, for exampJe, had no sparing effect on ruminal degradation of dietary
9 0
7O u )
°s0 £3 > - "1-
I-- ~3o
Io
BUFFALO
4 8 12 16 TIME (HOURS)
FIG. 1. Effect of time on proteolysis.
2 0
Journal of Dairy Science Vol. 61, No. 11, 1978

1576 MEHRA ET AL.
TABLE 3. Effect of substrate on proteolytic activity of cattle and buffalo RRL.a, b
Cattle Sp activ- Sp activ- ity/lO0 mg Percent ity/100 mg
Substrate protein hydrolysis protein
Buffalo
Percent hydrolysis
Casein 366 76 214 50 Hemoglobin 88 25 26 12 Bovine serum albumin 62 13 20 6
aReconstituted rumen liquor. bAverage of two experiments.
protein (28). Published information on manipu- lation of ruminal protease and deaminase activity is scanty. Ramirez (31) has reported diminished deaminase activity in the rumen microbes of animals fed low protein, molasses- urea diets. At tempts of Hogan and Weston (12) to control protease and/or deaminas6 activity with antibiotics did not produce encouraging results. Yet, another way to circumvent rumen microbial degradation of dietary protein has been suggested wherein the food passes directly through the esophageal groove into abomasum thus bypassing the rumen altogether (25, 26, 27).
The findings of our investigation, although confined to in vitro, reveal that buffalo RRL is far less proteolytic than that of cattle; a similar situation might prevail in vivo as well. The proteolysis proceeded progressively in both animals, but at each interval the buffalo RRL was much less proteolyt ic than cattle (Fig. 1). These findings are substantiated further by experiments with different substrates (casein, hemoglobin, and bovine serum albumin). Although the degree of proteolysis varied with the substrate, casein was hydrolyzed to the maximum whereas bovine serum albumin was to the lowest degree (Table 3). The solubility of all three substrates in .1 M phosphate buffer (pH 7.0) was about the same, yet their suscep- tibili ty to enzymatic degradation varied. In this regard, our findings appear to be at variance with the view that the solubility of dietary proteins in rumen liquor determines the criter- ion for the degree of proteolysis (8, 19, 20, 21). It appears plausible that substrate specificity, as well as substrate solubility, also affects the
degree of proteolysis in the rumen. Furthermore, buffalo RRL appears to have
more microbial population than cattle as evidenced by more total protein, nonprotein nitrogen, and dry weight. Pant and Roy (29) and Ganiev and Kafarov (11) also have reported that buffalo rumen liquor has higher total bacterial count than cattle. Anaerobic condi- tion potentiates proteolysis both in buffalo and cattle. In this regard, these findings are in conformity with those on sheep rumen liquor (5).
A diminished proteolysis, as exhibited by the rumen microorganisms of buffalo as com- pared with those of cattle, is essentially a specific phenomenon particularly when a reverse profile is projected of chemical con- stituents. In view of this situation, it is likely that relatively more of undegraded dietary protein may be passing out of buffalo rumen and is available to the animal for normal physiological functions. This possibly could be due either to elaboration of comparatively lower concentration of proteolytic enzyme(s) or to release of some "ant iproteolyt ic factors" by the rumen microbes of buffalo. In this connection, the report by Fossum (9) of a nondialyzable and ammonium sulphate- precipitable factor that inhibited bacterial proteinases is specifically noteworthy.
A C K N O W L E D G M E N T
The authors are grateful to C. M. Singh, Director, Indian Veterinary Research Institute, Izatnagar (U.P.), for interest in this investi- gation.
Journal of Dairy Science Vol. 61, No. 11, 1978

MICROBIAL A C T I V I T Y IN R U M I N A N T S 15 77
J
0
t ~
e~
0
O
0 = O
=_
a
4
M
< [...
0
<
<
, E . > . ~ =
~ . ~ ~
vl
u
<
<
~ ' ~
~ . ~ ~
, , E "d" 0
= 0
.=. ~J
o
.=.
"el- , "
d 0
Q
Q
e ~ 0
e ~
~ ~.~
~.~ • ~ , .~ ~ 0
".c,
0
REFERENCES
1 Abou Akkada, A. R., and B. H. Howard. 1962. The biochemistry of rumen protozoa. 5. The nitrogen metabolism of Entodinium. Biochem. J. 82:313.
2 Annison, E. F. 1954. Some observations on the volatile fat ty acids in the sheep's rumen. Biochem. J. 57:400.
3 Annison, E. F. 1956. Nitrogen metabolism in the sheep. Biochem. J. 64:705.
4 Appleby, J. C. 1955. The isolation and classifica- tion of proteolytic bacteria from the rumen of the sheep. J. Gen. Microbiol. 12: 526.
5 Blackburn, T. H., and P. N. Hobson. 1960. Pro- teolysis in the sheep rumen by whole and frac- t ionated rumen contents. J. Gen. Microbiol. 22:272.
6 Chalmers, M. I., D. P. Cuthbertson, and R. L. M. Synge. 1954. Ruminal ammonia formation in relation to protein requirement of sheep. I. Duo- denal administration and heat processing as factors influencing the fate o f casein supplements. J. Agr. Sci. 44:254.
7 Chalmers, M. I., and R. L. M. Synge. 1954. Ru- minal ammonia formation in relation to protein requirement of sheep. If. Comparison of casein and herring meal supplements. J. Agr. Sci. 44:263.
8 Ely, D. G., C. O. Little, P. G. Woolfolk, and G. E. Mitchell, Jr. 1967. Estimation of the extent of conversion of dietary zein to microbial protein in the rumen of lambs. J. Nutr. 91:314.
9 Fossum, R. 1971. Proteolytic enzymes and biologi- cal inhibitors. IV. Bacterial proteinase inhibitor and their effect upon enzymes of various origin. Biolog. Abstr. 52 (No. 14), 80289.
10 Fox, K. K. 1970. Casein and whey p ro t e in in by-products from milk by B. H. Wells and E. O. Whittier. The AVI Publishing Company, Inc.
11 Ganiev, M. K., and M. S. Kafrov. 1965. Changes in numbers of bacteria in the separate chambers of the forestomoches buffaloes and red cattle on different rations. Nutr. Abstr. Rev. 35:6179.
12 Hogan, J. P., and R. H. Weston. 1969. The effects o f antibiotics on ammonia accumulation and protein digestion in the rumen. Australian J. Agr. Res. 20:339. Hogan, J. P. 1975. Quantitative aspects o f nitrogen utilization in ruminants. J. Dairy Sci. 58:1164. Hunt, W. G., and R. O. Moore. 1958. The proteoly- tic system of a gram-negative bacteria isolated from the bovine rumen. AppI. Microbiol. 6:36.
15 Indian standard specifications for mineral mixture for supplementing cattle feeds. 1968. Indian Standard Institution, Manak Bhawan, New Delhi, India, page 5.
16 Leng, R. A. 1973. Salient features of the digestion of pasture by ruminants and other herbivores. Page 81 in Chemistry and biochemistry of herbage, vol. 3. G. W. Butler and R. D. Bailey, ed. Academic Press, New York.
17 Lewis, D., and H. Swan. 1971. The role of intes- tinal flora in animal nutrition. Page 149 in Micro- bes and biological productivity. Twenty-first
13
14
Journal of Dairy Science Vol. 61, No. 11, 1978

1 5 7 8 MEHRA ET AL.
Symp. Soc. Gen. Microbiol. D. E. Hughes and A. H. Rose, ed. Cambridge University Press, New York.
18 Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265.
1 9 McDonald, I. W. 1952. The role of ammonia in ruminal digestion o f protein. Biochem. J. 51:86.
20 McDonald, I. W. 1954. The extent of conversion o f food protein to microbial protein in the rumen of the sheep. Biochem. J. 56:120.
21 McDonald, I. W., and R. J. Hall. 1957. The conver- sion of casein into microbial protein in the rumen. Biochem. J. 67:400.
22 Nolan, J. V., and R. A. Leng. 1972. Dynamic aspects of ammonia and urea metabol ism in sheep. Brit. J. Nutr. 27:177.
23 Nolan, J. V., W. B. Norton, and R. A. Leng. 1972. Dynamic aspects of nitrogen metabol i sm in sheep. Page 13 in Tracer studies on non-protein nitrogen for ruminants . Int. Atomic Energy Agency, Vien- na.
24 Nolan, J. V., W. H. Norton, and R. A. Leng. 1973. Nitrogen cycling in sheep. Proc. Nutr. Soc. 32:93.
25 Orskov, E. R., and D. Benzie. 1969. Using the oessophageal groove reflex in ruminan t s as a mean of by-passing rumen fermenta t ion with high quali- ty protein and other nutrients . Proc. Nutr . Soc. 28:30A.
26 ~rskov, E. R., D. Benzie, and R. N. B. Kay. 1970. The effects of feeding procedure on closure of the oesophageal groove in young sheep. Brit. J. Nutr. 24:785.
27 ¢)rskov, E. R., C. Fraser, and E. L. Corse. 1970. The effect on protein utilization o f feeding differ- ent protein supplements via the rumen or via the abomasum in young growing sheep. Brit. J. Nutr. 24:803.
28 ~rskov, E. R., C. Fraser, I. McDonald, and R. I.
Smart. 1974. Digestion of concentrates in sheep. 5. The effect o f adding fish meal and urea together to cereal diets on protein digestion and utilization by young sheep. Brit. J. Nutr. 31:491.
29 Pant, H. C., and A. Roy. 1969. Seminar on ru- minant digestion and metabol ism. Indian Veteri- nary Research Institute, Izatnagar, U.P., India.
30 Parvin, R., S. V. Pande, and T. A. V. Subramanian. 1965. On the colorimetric buiret me thod of protein determinat ion. Anal. Biochem. 12:219.
31 Ramirez, A. 1972. Deaminative activity of tureen microflora with molasses/urea diets. Rev. Cubana Cierc. Agr. 6:35.
32 Satter, L. D., and R. E. Roffler. 1975. Nitrogen requirement and utilization in dairy cattle. J. Dairy Sci. 58:1219.
33 Schneider, W. C. 1957. Determinat ion of nucleic acids in tissues by pentose analysis. Page 680 in Methods of enzymology. Vol. 3. Sidney P. Colo- wick and Nathan O. Kaplan, ed. Academic Press, New York.
34 Sen, K. C., and S. N. Ray. 1971. Nutrit ive values o f Indian cattle feeds and the feeding of animals. Indian Council Agr. Res., New Delhi.
35 Smith, R. H. 1969. Nitrogen metabol ism and the rumen. J. Dairy Res, 36:313.
36 Smith, R. H., D. N. Salter, J. D. Sut ton, and A. B. McAllan. 1974. Synthesis and digestion of micro- bial ni trogen compounds and VFA product ion by the bovine. In Tracer studies on non-protein nitrogen for ruminants . Int. Atomic Energy Agency, Vienna.
37 Sym, E. A. 1938. Hydrolasenwirkung dis. Blind- sackinhaltes despferdes und des panseninhal tes des Rindes. I. Tell einlitung, allegemeine me thoden und proteolyt ische wirkungen. Acta Biol. Exp. Varsovie, 12:192.
38 Warner, A. C. 1. 1956. Proteolysis by rumen microorganisms. J. Gen. Microbiol. 14:733.
Journal of Dairy Science Vol. 61, No. 11, 1978