production of monoclonal antibodies against thermostable direct hemolysin of vibrio parahaemolyticus...
TRANSCRIPT

Med Microbiol Immunol (1989) 178:245-253
�9 Springer-Verlag 1989
Production of monoclonal antibodies against thermo- stable direct hemolysin of Vibrio parahaemolyticus and application of the antibodies for enzyme-linked immunosorbent assay*
T. Honda, Y. Ni, M. Yoh, and T. Miwatani
Research Institute for Microbial Diseases, Osaka University, Yamadaoka, Suita, Osaka 565, Japan
Abstract. A total nine hybridoma cell lines that produced monoclonal antibodies against thermostable direct hemolysin (Vp-TDH), a possible pathogenic toxin, of Kanagawa phenomenon-positive Vibrio parahaemolyti- cus was isolated and characterized. These monoclonal antibodies (mAbs) were divided into a minimum of five different specificity groups, including mAbs specific to Vp-TDH and common to Vp-TDH and Vp-TRH, a Vp-TDH- related hemolysin produced by Kanagawa phenomenon-negative V. paraha- emolyticus. An enzyme-linked immunosorbent assay (ELISA) using mAb-1- D, a mAb specific for Vp-TDH, was developed for specific detection of Vp- TDH. On the other hand, the ELISA using mAb-9-D, and mAb common to both Vp-TDH and Vp-TRH, could be used for detection of both Vp-TDH and Vp-TRH. Thus, by combining these two ELISAs differential detection of Vp- TDH and Vp-TRH can be performed. Hence, the two ELISAs were applied for various strains of V. parahaemolyticus and it was found that most Kanagawa phenomenon-positive and -negative clinical isolates produced Vp- TDH and Vp-TRH, respectively, but all environmental strains, that were Kanagawa phenomenon-negative, produced neither toxin.
Introduction
Vibrioparahaemolyticus is a major cause of acute gastroenteritis worldwide [12, 13, 15, 19]. For example, this organism has been found to account for about 40% to 60 % of all food poisoning cases in Japan [ 15]. V. parahaemolyticus is classified into
* Supported by a Grant-in-Aid for Scientific Research and a Grant-in-Aid for Developmental Scientific Research from the Ministry of Education, Science, and Culture of Japan
Offprint requests to." T. Honda

246 T. Honda et al.
two types on the basis of the Kanagawa phenomenon [17, 22], a test of whether organisms cause hemolysis by means of thermostable direct hemolysin (Vp-TDH) on a special blood agar medium (Wagatsuma medium). Various studies including epidemiological analysis [5-9, 12, 13, 15, 17-20, 22-26] suggest that a positive Kanagawa phenomenon is closely associated with the pathogenicity of this organism in humans; for example most clinical isolates (96.5% of 2,270 strains) have been found to be Kanagawa positive, whereas 99.0% strains of 650 cultures isolated from the environment have been found to be Kanagawa negative [22]. Thus, various studies on Vp-TDH have been performed, some of which described enterotoxicity [18], cardiotoxicity [5] or cytotoxicity [23] as biological activities of Vp-TDH. Amino acid sequences of Vp-TDH have been deduced by both protein [26] and DNA sequencing [20]. It has also been reported that Vp-TDH takes a dimer form composed of two identical subunits [24].
Despite the accumulated evidence, the detailed antigenic structures of Vp-TDH are not yet well known. In this study, we report, for the first time, on the isolation and characterization of hybridomas which produce monoclonal antibodies (mAbs) against Vp-TDH, and their application in an enzyme-linked immunosor- bent assay (ELISA) to detect Vp-TDH. We also discuss the immunological cross- reactivity with Vp-TDH of a newly described hemolysin (Vp-TDH-related hemolysin; Vp-TRH) produced by Kanagawa phenomenon-negative clinical isolates of V. parahaemolyticus [10, 11].
Materials and methods
Bacterial strains and culture. All strains, including Kanagawa-positive clinical isolates, Kanagawa-negative clinical isolates and Kanagawa-negative environmen- tal (from sea water and fish) isolates, used in these experiments were from the Laboratory for Culture Collections, Research Institute for Microbial Diseases, Osaka University, Osaka, Japan. V. parahaemolyticus strains were cultured at 37~ under rotation (20rpm) for 20h in 1 ml of SPP broth [10, 11], consisting of 5g NaCI, 10g peptone (Difco Laboratories, Detroit, Mich.), 2g glucose, and 5g dibasic sodium phosphate per liter, in 5-ml test tubes.
Purified hemoiysins. Vp-TDH and Vp-TRH were purified from cultures of clinical isolates of Kanagawa phenomenon-positive (T4750) and -negative (TH4037) V. parahaemolyticus, respectively, by published techniques [6, 10, 11].
Preparation of mAbs. For mouse immunization, a Vp-TDH suspension in phosphate-buffered saline (PBS, 0.01 M, pH 7.2) was mixed with an equal amount of complete Freund adjuvant, and then 0.25 ml of the mixture, containing 5 lag or 15 lag of the toxin, was injected intraperitoneally into BALB/c mice at 2-week intervals. After 4 weeks, another booster dose of 20 lag of toxin with incomplete Freund's adjuvant was given intraperitoneally. A final booster of 15 lag of toxin in 0.3 ml of PBS was intravenously injected via the tail vein. Three days after the last booster injection, spleen cells from the immunized mice were fused with X6-Ag 8.653 myeloma cells using polyethylene glycol (PEG 1500, Boehringer Mannheim,

Monoclonal antibody against hemolysin of V. parahaemolyticus 247
FRG) according to published methods [3, 14]. The resulting hybrid cells were selected by treatment with Celgrosser-H medium (Sumitomo Seiyaku Co., Osaka, Japan) supplemented with 2.5 % fetal bovine serum and 1 X 10 -4 M hypoxanthine (H), 4 X 10 -7 M aminopterin (A) and 1.6 X 10 -5 M thymidine (T) (HAT).
Hybridomas developed were screened according to their ability to produce mAbs recognizing Vp-TDH bound to vinyl microtiter wells using the ELISA protocol (see below). Positive hybridomas were recloned at least twice by limiting dilution. The positive hybridomas in ELISA were grown in Celgrosser-H medium supplemented with HT and the culture supernatants, obtained by centrifugation at 2,500g for 15 min, were used in the following experiments.
ELISA. ELISA for detecting mAbs against Vp-TDH was carried out essentially as reported previously [9]. A flat-bottom microplate precoated with 100 gl of Vp- TDH (4 gg/ml) or bovine serum albumin (4 gg/ml; negative control) was reacted with culture supernatants (70 gl each) of hybridomas.
The procedure for detection of Vp-TDH or/and Vp-TRH by a sandwich ELISA has also been described previously [9]. After precoating a vinyl microplate with polyclonal rabbit anti-Vp-TDH immunoglobulin, test samples (100 gl), mAbs (100 gl of 1:4,000 diluted ascites) and then alkaline phosphatase-conjugated goat anti-mouse immunoglobulins (100 lal) were reacted in turn.
Immunoblot (Western blot) analysis. Western blot analysis using Vp-TDH or Vp- TRH and mAbs was performed essentially as described previously [16]. The nitrocellulose strip-transferred toxins were reacted in 5 ml of culture supernatant of the hybridomas overnight at 4~ with gentle shaking. After reacting with peroxidase-conjugated rabbit anti-mouse immunoglobulin (1:1,000 dilution; Zymed Laboratories, San Francisco, Calif.), enzyme activity was detected by exposing the strips to 10 mM Tris-HCl buffer (pH 7.0) containing 0.05 % (wt/vol) 4 chloro-1-naphthol and 0.015 % (vol/vol) hydrogen peroxidase at room temperatu- re for about 40 min.
Isotype determination of mAbs. Isotype determination of immunoglobulin in culture supernatants of each monoclone was performed using a MonoAb-ID EIA kit (Zymed Laboratories,) according to the procedure recommended by the manufacturer.
Determination of mAb concentration. To determinate mAb concentrations in culture supernatants from hybridoma cell culture, radial immunodiffusion assays were performed as described previously [4]. Goat anti-mouse IgG (Cappel, Organon Teknika Corporation, West Chester, Pa) was diluted to 1:300 in 1.0% Noble agar (Difco). A purified mouse IgG (Zymed Laboratories) was used as a standard.
Neutralization. Neutralization of the hemolytic activity of hemolysins by the isolated mAbs was determined as follows. A mixture of 25 gl of toxin containing 2 hemolytic units (HU) [10] diluted with 10 mM Tris-HC1 (pH 7.0) buffer containing 0.85 % NaCI (Tris~ and 75 lal of concentrated or diluted (to adjust 10 or 80 gg/ml

248 T. Honda et al.
of immunoglobulin concentration) culture supernatants of each hybridoma were preincubated at 37~ for 30min, and then 100gl of a 2% suspension of washed human erythrocytes suspended in Tris-S was added and incubated at 37~ for 30 min. After incubation, supernatants obtained by centrifugation at 1,500g for 2 rain were transferred to a flat-bottom microtiter plate, and the A540 was measured in a Titertek Multiskan MC apparatus. As negative controls, the culture medium (Celgrosser-H) was used instead of culture supernatants of hybridomas. When the culture supernatants of given hybridoma showed neutralizing activity with an immunoglobulin concentration of 10tag/ml, it was considered as positive for neutralization.
ModifiedElek test. To determine the Kanagawa phenomenon, a modified Elek test developed by ourselves was used as described previously [7].
Hemolysin assay. To detect Vp-TDH and Vp-TRH differentially, the gel after conventional slab polyacrylamide gel electrophoresis (PAGE) [2] of crude concentrated (tenfold) culture supernatants of test organisms was overlaid on an agar plate in Tris-S containing 5 % human erythrocytes and the plate incubated at 37~ for 2-3 h before reading the results [11].
Results
Establishment of hybridomas producing antibodies against Vp-TDH
A fusion experiment using three immunized mice was performed and a total of 469 wells containing hybridomas was obtained. Of these, 32 hybridomas were found to produce antibodies against Vp-TDH. Nine stable monoclones (Table 1) producing anti-Vp-TDH antibody were finally established. All mAbs isolated belonged to the IgG class (Table 1).
Characterization of mAbs
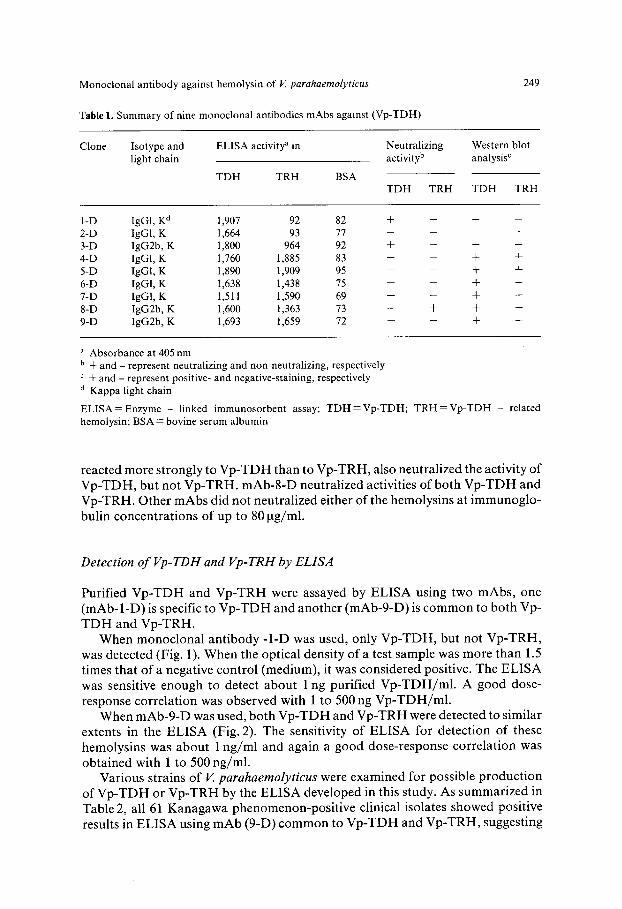
Cross-reactivity of isolated mAbs was analyzed by ELISA using Vp-TDH-, Vp- TRH- or BSA-coated plates. As summarized in Table 1, mAbs-l-D and -2-D recogniszed Vp-TDH but not Vp-TRH. mAbs-3-D and -8-D reacted strongly to Vp-TDH and weakly to Vp-TRH. Other mAbs recognized both Vp-TDH and Vp- TRH to a similar extent in the ELISA.
The reactivity of these mAbs to the toxins was also examined by Western blot analysis. The results of Western blot analysis with all mAbs are also summarized in Table 1. The two clones (1-D and 2-D), which recognized only Vp-TDH in ELISA, showed no binding activity to Vp-TDH in Western blot analysis. Clone 3-D was non-reactive in blot analysis, although it was reactive with both Vp-TRH and Vp- TDH in ELISA. The results of the neutralization of hemolytic activities by mAbs are also summarized in Table 1. Of the clones which recognized only Vp-TDH in the ELISA, mAb-l -D could neutralize the activity of Vp-TDH. mAb-3-D, which
Monoclonal antibody against hemolysin of V. parahaemolyticus
Table 1. Summary of nine monoclonal antibodies mAbs against (Vp-TDH)
249
Clone Isotype and ELISA activity a in Neutralizing light chain activity b
TDH TRH BSA
Western blot analysis c
TDH TRH TDH TRH
1-D IgGl, K d 1,907 92 82 + -- -- -- 2-D IgGl, K 1,664 93 77 . . . . 3-D IgG2b, K 1,800 964 92 + -- - -- 4-D IgG1, K 1,760 1,885 83 -- -- + + 5-D IgG1, K 1,890 1,909 95 -- -- + + 6-D IgGl, K 1,638 1,438 75 -- -- + + 7-D IgGl, K 1,511 1,590 69 -- -- + + 8-D IgG2b, K 1,600 1,363 73 + + + + 9-D IgG2b, K 1,693 1,659 72 -- -- § +
a Absorbance at 405 nm b + and - represent neutralizing and non-neutralizing, respectively
+ and - represent positive- and negative-staining, respectively a Kappa light chain
ELISA = Enzyme - linked immunosorbent assay; TDH=Vp-TDH; TRH =Vp-TDH - related hemolysin; BSA = bovine serum albumin
reacted more s t rongly to V p - T D H than to V p - T R H , also neutra l ized the act ivi ty of V p - T D H , but not V p - T R H . m A b - 8 - D neutra l ized activities o f bo th V p - T D H and V p - T R H . Other m A b s did not neutra l ized ei ther o f the hemolysins at i m m u n o g l o -
bulin concent ra t ions of up to 80 tag/ml.
Detection of Vp-TDH and Vp-TRH by ELISA
Purif ied V p - T D H and V p - T R H were assayed by E L I S A using two mAbs , one ( m A b - l - D ) is specific to V p - T D H and ano ther (mAb-9-D) is c o m m o n to bo th Vp-
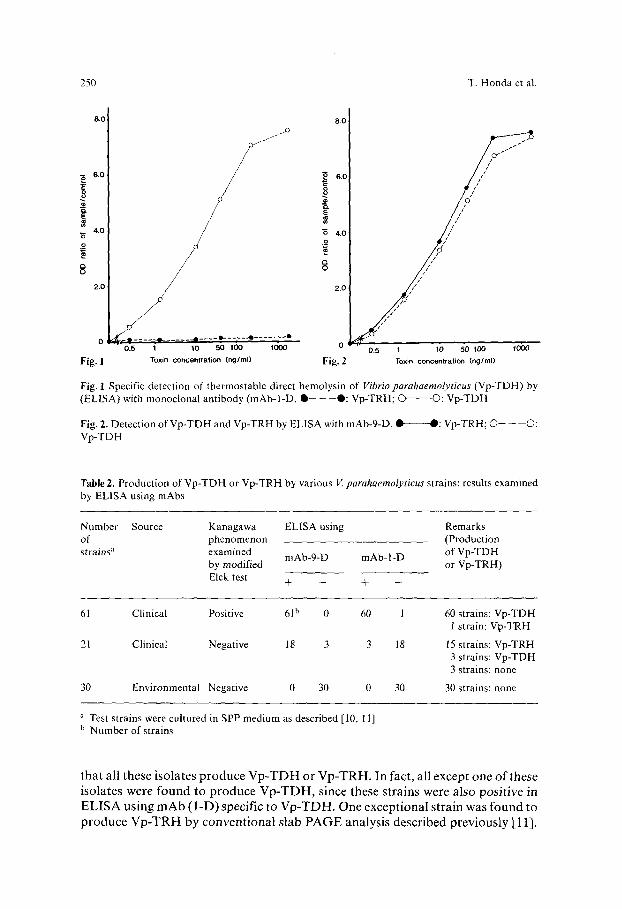
T D H and V p - T R H . W h e n m o n o c l o n a l an t ibody -1-D was used, only V p - T D H , but not V p - T R H ,
was detected (Fig. 1). When the opt ical density of a test sample was more than 1.5 t imes that o f a negat ive cont ro l (medium), it was considered positive. The E L I S A
was sensitive enough to detect about 1 ng purif ied V p - T D H / m l . A g o o d dose-
response cor re la t ion was observed with 1 to 500 ng V p - T D H / m l . When m A b - 9 - D was used, bo th V p - T D H and V p - T R H were detected to s imilar
extents in the E L I S A (Fig. 2). The sensitivity o f E L I S A for de tec t ion of these hemolys ins was abou t 1 n g / m l and again a good dose-response cor re la t ion was
ob ta ined with 1 to 500 ng /ml . Various strains o f V. parahaemolyticus were examined for possible p roduc t i on
of V p - T D H or V p - T R H by the E L I S A deve loped in this study. As summar i zed in Table 2, all 61 K a n a g a w a phenomenon-pos i t i ve clinical isolates showed posi t ive
results in E L I S A using m A b (9-D) c o m m o n to V p - T D H and V p - T R H , suggest ing
250 T. Honda et al.
8,0
6.0
4.0
._o
Q O
2.0
0
Fig. 1
8.0
i 6.0
i 4.0
8
2.0
�9 � 9 _Q 0 - -4D �9 -p _ . . . . "- - - - - - - - T . . . . . . . . . . -'-- 0.5 1 10 50 1OO 1000 0
Toxin concentration (ng/ml) Fig, 2
U / /
? '
0.5 "J 10 50 tOO 1000
Toxin concentration (ng/ml)
Fig. 1 Specific detection of thermostable direct hemolysin of Vibrio parahaemolyticus (Vp-TDH) by (ELISA) with monoclonal antibody (mAb-l-D. Q - - - - - 0 : Vp-TRH; O O: Vp-TDH
Fig. 2. Detection of Vp-TDH and Vp-TRH by ELISA with mAb-9-D. 1~-----O: Vp-TRH; �9 -- --O: Vp-TDH
T a b l e 2. Production of Vp-TDH or Vp-TRH by various V. parahaemolyticus strains: results examined by ELISA using mAbs
Number Source Kanagawa ELISA using Remarks of phenomenon (Production strains a examined mAb-9-D mAb-l-D of Vp-TDH
by modified or Vp-TRH) Elek test § - - + -
61 Clinical Positive 61 b 0 60 1
21 Clinical Negative 18 3 3 18
30 Environmental Negative 0 30 0 30
60 strains: Vp-TDH 1 strain: Vp-TRH
15 strains: Vp-TRH 3 strains: Vp-TDH 3 strains: none
30 strains: none
~' Test strains were cultured in SPP medium as described [10, 11] b Number of strains
that all these isolates produce V p - T D H or Vp-TRH. In fact, all except one of these isolates were found to produce Vp-TDH, since these strains were also positive in ELISA using mAb ( l -D) specific to Vp-TDH. One exceptional strain was found to produce Vp-TRH by conventional slab PAGE analysis described previously [11].

Monoclonal antibody against hemolysin of I~ parahaemolyticus 251
Strains of Kanagawa phenomenon-negative clinical isolates were also examined by ELISA. Although most of these strains (18 out of 21) were positive in ELISA using mAb common to Vp-TDH and Vp-TRH, 15 were also negative in ELISA using mAb (l-D) specific to Vp-TDH, suggesting that they produce only Vp-TRH. This was verified by conventional PAGE analysis [11]. On the other hand, all environmental strains were found to be negative in ELISA using both mAbs. There were no strains which produced both Vp-TDH and Vp-TRH at the same time.
Discussion
In this report, we have demonstrated for the first time the production of mAbs developed against Vp-TDH, which is considered to be closely associated with the enteropathogenicity of Kanagawa phenomenon-positive V. parahaemolyticus [ 13, 15, 17, 22], a common cause of acute gastroenteritis. The mAbs developed against Vp-TDH in this study can be classified into five groups: (1) antibody (l-D) recognizing only Vp-TDH in ELISA and neutralizing only the activity of Vp- TDH, (2) antibody (2-D) recognizing only Vp-TDH in ELISA but not neutralizing the activity, (3) antibody (3-D) strongly recognizing Vp-TDH and weakly recognizing Vp-TRH and neutralizing the activity of Vp-TDH, (4) antibody (8-D) neutralizing and recognizing both Vp-TDH and Vp-TRH, and (5) antibodies (4-D, 5-D, 6-D, 7-D and 9-D) recognizing both Vp-TDH and Vp-TRH in ELISA, but not neutralizing their activities. These results indicate the presence of at least five epitopes in the Vp-TDH molecule. The present data also clearly demonstrates the presence of epitopes in Vp-TDH and Vp-TRH unique and common to these two toxins.
In this study, we also described ELISA systems using mAbs developed against Vp-TDH. The ELISA was found to be sensitive enough for detection and identification of production of Vp-TDH and Vp-TRH. When an mAb (9-D) common to Vp-TDH and Vp-TRH was used in ELISA, strains which produce either Vp-TDH or Vp-TRH were detected. On the other hand, when an mAb (l-D) specific to Vp-TDH was used in ELISA, only strains producing Vp-TDH were detected, and thus, strains producing Vp-TRH could also be identified. Although several ELISA systems for detection of Vp-TDH have been reported [9, 21], all used polyclonal antibodies and thus could not differentiate Vp-TDH- and Vp- TRH-producing strains. In our ELISA using mAbs-l-D and -9-D, however, strains which produce Vp-TDH or Vp-TRH can be differentiated. Although several methods to improve detection of Kanagawa phenomenon have been developed [1, 7-9, 17, 21], the present system should provide an advantage in various types of research, especially in terms of epidemiological analysis on the distribution of Vp-TDH and Vp-TRH production among strains V. parahaemolyti- cus from various sources. Results obtained by the ELISA in this study show that most Kanagawa phenomenon-negative clinical isolates produce Vp-TRH, whereas environmental strains produce neither Vp-TDH nor Vp-TRH. The data in this study also confirm that most Kanagawa phenomenon-positive clinical isolates produce Vp-TDH [17, 22]. These data strongly suggest the importance of Vp-TRH or Vp-TDH in the pathogenesis of V. parahaemolyticus.

252 T. Honda et al.
References
1. Chan D, Chung JK, Tak Y, Seol SY (1975) Nature of the Kanagawa phenomenon of Vibrio parahaemolyticus. Infect Immun 12:81-87
2. Davis BJ (1964) Disc electrophoresis. II. Method and application to human serum proteins. Ann NY Acad Sci 121:404-427
3. Galfre G, Howe SC, Milstein C, Butcher GW, Howard JC (1977) Antibodies to major histocompatibility antigens produced by hybrid cell lines. Nature 266:530-552
4. Holmes R, Twiddy EM (1983) Characterization of monoclonal antibodies that react with unique and cross-reacting determinants of cholera enterotoxin and its subunits. Infect Immun 42:914-923
5. Honda T, Goshima K, Takeda Y, Sugino Y, Miwatani T (1976) Demonstration of the cardiotoxicity of the thermostable direct hemolysin (lethal toxin) produced by V. parahaemolyticus. Infect Immun 13:163-171
6. Honda T, Taga S, Takeda T, Hashibunan MA, Takeda Y, Miwatani T (1976) Identification of lethal toxin with the thermostable direct hemolysin produced by Vibrio parahaemolyticus and some physicochemical properties of the purified toxin. Infect immun 13:133-139
7. Honda T, Chearkul S, Takeda Y, Miwatani T (1980) Immunological methods for detection of Kanagawa phenomenon of Vibrio parahaemolyticus. J Clin Microbiol 11:600-603
8. Honda T, Sornchai C, Takeda Y, Miwatani T (1982) Immunological detection of the Kanagawa phenomenon of Vibrioparahaemolyticus on modified selective media. J Clin Microbiol 16:734-736
9. Honda T, Yoh M, Kongmuang U, Miwatani T (1985) Enzyme-linked immunosorbent assays for detection of thermostable direct hemolysin of Vibrioparahaemolyticus. J Clin Microbio122:383-386
10. Honda T, Ni Y, Miwatani T (1988) Prification and characterization of a hemolysin produced by a clinical isolate of Kanagawa phenomenon-negative Vibrio parahaemolyticus and related to the thermostable direct hemolysin. Infect Immun 56:961-965
11. Honda T, Ni Y, Miwatani T (1989) Purification of a TDH-related hemolysin produced by a Kanagawa phenomenon negative clinical isolate of Vibrio parahaemolyticus 06:K46. FEMS Microbiol Lett 57:241-246
12. Janda JM, Powers C, Bryant RG, Abbott SL (1988) Current perspectives on the epidemiology and pathogenesis of clinically significant Vibrio spp. Clin Microbiol Rev 1:245-267
13. Joseph SW, Colwell RP, Kaper JB (1982) Vibrio parahaemolyticus and related halophilic vibrios. Crit Rev Microbiol 10:77-124
14. McKearn TJ (1980) Fusion of cells in an adherent monolayer. In: Kennett RH, McKearn TJ, Bechtol KB (ed) Monoclonal antibodies, hybridomas: a new dimension in biological analyses. Plenum Press, New York, pp 368-369
15. Miwatani T, Takeda Y (1976) Vibrio parahaemolyticus-a causative bacterium of food poisoning. Saikon, Tokyo, pp 1-109
16. Miyake M, Honda T, Miwatani T (1989) Effects of divalent cations and saccharides on Vibrio metschnikovii cytolysin-induced hemolysis of rabbit erythrocytes. Infect Immun 57:158-163
17. Miyamoto Y, Kato T, Obara Y, Akiyama S, Takizawa K, Yamai S (1969) In vitro hemolytic characteristics of Vibrio parahaernolyticus: its close correlation with human pathogenicity. J Bacteriol 100:1147-1149
18. Miyamoto Y, Obara Y, Nikkawa T, Yamai S, Kato T, Yamada Y, Ohashi M (1980) Simplified purification and biophysicochemical characteristics of Kanagawa-associated hemolysin of V. parahaemolyticus. Infect Immun 23:567-576
19. Morris JG Jr, Blake RE (1985) Cholera and other vibrioses in the United States. N Engl J Med 312:343-350
20. Nishibuchi M, Kaper JB (1985) Nucleotide sequence of the thermostable direct hemolysin gene of Vibrioparahaemolyticus. J Bacteriol 162:558-564
21. Oku Y, Uesaka Y, Hirayama T, Takeda Y (1988) Development of a highly sensitive bead-ELISA to detect bacterial toxins. Microbiol Immunol 32:807-816
22. Sakazaki R, Tamura K, Kato T, Obara Y, Yamai S, Hobo K (1968) Studies on the enteropathogenic, facultatively halophilic bacteria, Vibrioparahaernolyticus. III. Enteropathogenicity. Jpn J Med Sci Biol 21:325-331
23. Sakurai J, Honda T, Jinguji Y, Arita M, Miwatani T (1976) Cytotoxic effect of the thermostable direct hemolysin produced by V. parahaemolyticus on FL cells. Infect Immun 13:876-883

Monoclonal antibody against hemolysin of V. parahaemolyticus 253
24. Takeda Y, Taga S, Miwatani T (1978) Evidence that thermostable direct hemolysin of V. parahaemolyticus is composed of two subunits. FEMS Microbiol Lett 4:271-274
25. Toda H, Sakiyama F, Yoh M, Honda T, Miwatani T (1985) Chemical modification of thermostable direct hemolysin of V. parahaemolyticus. Jpn J Med Sci Biol 38:264-265
26. Tsunasawa S, Sugihara A, Masaki T, Sakiyama F, Takeda Y, Miwatani T, Narita K (1987) Amino acid sequence of thermostable direct hemolysin produced by V. parahaemolyticus. J Biochem 101:11-121
Received May 10, 1989