prenatal cocaine exposure and postnatal hypoxia independently decrease carotid body dopamine in...
TRANSCRIPT

ELSEVIER
Neurotoxicology and Teratology, Vol. 18, No. 3, pp. 283-287,1996 Copyright 0 1996 Elsevier Science Inc. Printed in the USA. All rights reserved
0892-0362/96 $15.00 + .OO
PI1 SO892-0362(96)00043-S
Prenatal Cocaine Exposure and Postnatal Hypoxia Independently Decrease Carotid Body
Dopamine in Neonatal Rats
JACK W. LIPTON,“? ANCHALEE YUENGSRIGUL,_F ZAO DUNG LING,* DEBRA E. WEESE-MAYER? AND PAUL M. CARVEY*
Departments of *Neurological Sciences and tpediatrics, Rush Medical College of Rush University, Rush Children’s Hospital, Rush-Presbyterian-St. Luke’s Medical Center, Chicago, IL 60612
Received 25 April 1995; Accepted 11 January 1996
LIPTON, J. W., A. YUENGSRIGUL, Z. D. LING, D. E. WEESE-MAYER AND P. M. CARVEY. Prenatal cocaine expo- sure and postnatal hypoxia independently decrease carotid body dopamine in neonatal rats. NEUROTOXICOL TERATOL B(3) 283-287,1996.-The effects of prenatal cocaine exposure on the levels of carotid body dopamine (DA) and its metabo- lites 3,4_dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA) were investigated in 5-day-old rat pups ex- posed to normoxic and hypoxic conditions. Timed-pregnant Sprague-Dawley rats were injected b.i.d. with either cocaine HCl (30 mg/kg) or isotonic saline (1 ml/kg) from gestational days 7-21. On the fifth postnatal day, pups were subjected to either 20 min of 0.21 or 0.08 fractional inspired oxygen (F102). Under a strictly timed protocol, both carotid bodies were removed from each pup, placed in an antioxidant solution to prevent DA breakdown, and subsequently analyzed via HPLC with electro- chemical detection to determine carotid body DA and DOPAC content. Two-way ANOVA revealed decreases in DA in co- caine-exposed pups. No HVA was detectable in any of the samples. The 0.08 F,Oz condition decreased DA compared to 0.21 F102. The additive consequences of DA depletion resulting from the combination of prenatal cocaine and postnatal hypoxia decreased carotid body DA to 14% of control levels, with several animals exhibiting DA content below detection limits. Con- sidering the role of the carotid body in the ventilatory response to hypoxia, these data suggest that prenatal cocaine exposure may adversely affect the normal chemoreceptive function of the carotid body.
In utero cocaine Ventilation DOPAC Teratogenic effects
ABNORMAL ventilatory patterns (7,42,48) and altered ven- tilatory and arousal responses to hypoxia and hypercapnia (33,47) have been reported in the human infant after prenatal substance abuse that included cocaine. Smaller decrements in carbon dioxide tension during hypoxia in cocaine-exposed newborns have also been observed, suggesting that such in- fants exhibit a blunted ventilatory response to hypoxia (48). Taken together, these studies suggest that infants of sub- stance-abusing mothers not only exhibit altered respiratory patterns, they also have an impaired repertoire of protective responses to hypoxia and hypercapnia during sleep.
Structures associated with respiratory function and control are present in both the central (brain stem) and the peripheral nervous system. The brain stem dorsal respiratory group in-
cludes the nucleus tractus solitarius, which is active during in- spiration (38,44,45). The brain stem ventral respiratory group consists of the nucleus ambiguus and nucleus retroambiguus, which are active during both inspiration and expiration (35, 38,44). Although other neurotransmitters are involved, DA is one of the main neurotransmitters in these ponto-medullary areas (4,8,10). The carotid body, located at the bifurcation of the common carotid artery, is a peripheral chemoreceptor that is sensitive to changes in arterial oxygen pressure (PaOJ (14,46). It contains catecholaminergic glomus cells, sheath cells, and a complicated network of thin-walled capillaries (34). The carotid body is innervated by the carotid sinus nerve, an afferent sensory projection from the petrosal gan- glion, which includes catecholaminergic neurons. DA release
Requests for reprints should be addressed to Debra E. Weese-Mayer, M.D., Professor of Pediatrics, Chief, Pediatric Respiratory Medicine, Rush Children’s Hospital, 1653 W. Congress Parkway, Chicago, IL 60612.
283

284 LIPTON ET AL.
in the carotid body can be produced in response to decreased ~0, or pH, or increased pCOz (40). Although the chemo- transduction mechanism in the carotid body remains incom- pletely defined, available data suggest that DA, the principal neurotransmitter of the glomus cell, plays an important func- tional role in this process (1,27).
Prenatal cocaine exposure has been shown to influence DA levels in the striatum (5152) and nucleus accumbens (36) by blocking the DA transporter (24). In addition, dopaminer- gic structures are implicated in the regulation of respiratory drive (n. tractus solitarius, carotid body) (5,10,11,14,15,19, 20,32,46). If these well-documented actions of cocaine glo- bally affect the development of the DA system, then one would expect to observe manifestations of this drug exposure in the DA-containing structures of the respiratory control sys- tem. Thus, it seemed logical to evaluate the consequences of prenatal cocaine on one of these DA-containing respiratory structures. The following study examined the consequences of in utero cocaine administration on the carotid body. If prena- tal cocaine administration hinders carotid body development, one would expect to see decreases in carotid body DA con- tent and/or metabolism. In an effort to determine possible mechanisms whereby cocaine induces changes in the ventila- tory response to hypoxia in the neonate, the impact of prena- tal cocaine administration and/or hypoxia on DA levels in the carotid body of the 5-day-old rat pup was examined.
METHOD
Drug Administration and Subjects
Eight timed-pregnant Sprague-Dawley rats (Zivic-Miller) were housed individually in a 12-h light/dark cycled room, given free access to food and water, and weighed daily. Each dam was injected SC, twice per day with 30 mg/kg (0.089 mmol/kg) cocaine HCl or 1 ml/kg isotonic saline from gesta- tional days 7-21 [embryonic (E); E7-E21]. The injection sites were rotated daily and monitored for tissue necrosis. The pups born to these dams were randomly selected for study at 5 days postnatal age (P.5).
Rationale for Drug Administration
Dosages less than or equal to the 30 mg/kg used in this study have been shown to produce increased stereotypic be- haviors and sensitization in rats (21,22) as well as ventilatory alterations in rabbits (SO) without producing seizures or overt teratogenic malformations (9). Because abusers generally self-administer cocaine more than once per day (16) we ad- ministered cocaine at 30 mgikg twice per day, as opposed to a single loading dose of 60 mg/kg. This enabled us to extend co- caine’s action and provide a drug exposure paradigm that bet- ter approximated human abuse patterns than would a single dose. To avoid interference with ovum implantation (gesta- tional day S), drug administration was initiated on E7 and continued through E21 (the final full day of gestation) to al- low for exposure throughout gestation.
Physical Measures, Atmospheric Exposure, and Carotid Body Removal
Body weight and crown-rump length were measured for each pup. A strictly timed protocol for the processing of ca- rotid bodies was devised because previous attempts at quanti- fying carotid body DA indicated that neurotransmitter degra- dation was time-dependent. Groups of pups exposed to either
cocaine (COC) or saline (SAL) during gestation were each di- vided into subgroups for normoxic [21% fractional inspired oxygen (0.21 FiO,)] and hypoxic (0.08 FiOz) exposures.
Pups were placed into clear cylindrical chambers with a single inlet attached to a “Y” connector that blended the O2 and N2 sources. F,O, was controlled by manipulating the flow meters on each tank (total flow rate of 6 limin) and monitored using an FIOz analyzer (Teledyne, La Puente, CA). Soda lime was placed in the chamber to reduce atmospheric COz. Pups were exposed to the normoxic or hypoxic condition for 14 min followed by a 2-min infusion of 4% halothane (at the appro- priate F,Oz). The animals were then removed from the cham- ber while still being maintained on the halothane mixture via a neonatal gas mask that was connected to the inlet line from the chamber. After a vertical incision in the mid-sternal line, the underlying SC tissue was removed to expose the bifurca- tion of the common carotid arteries. Under a dissecting mi- croscope, the superior cervical ganglion was dissected clear of the carotid bodies. At 5 min after the onset of anesthesia, the left carotid body (still attached to the carotid bifurcation) was removed, followed 2 min later by the removal of the right ca- rotid body. This resulted in a normoxic or hypoxic exposure, averaging 20 min (a range of 19-21 min), before carotid body removal.
Specimen Preparation
The intact carotid body/bifurcation assemblies were imme- diately placed in 300 ~1 of a sodium metabisulfite antioxidant solution (6) (4°C) and frozen with dry ice. The samples were later thawed and homogenized for 15 s while surrounded by an ice water slurry and then centrifuged at 100,000 X g for 30 min at 4°C. The supernatant was reserved and stored at -80°C for subsequent HPLC analysis.
HPLC Analysis
The samples were analyzed for DA and its metabolites 3,4- dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA) via HPLC with electrochemical detection. The mobile phase consisted of a 14% methanol solution in distilled H20 containing 21 g/l (0.1 M) citric acid, 10.65 g/l (0.075 M) NazHPO,, 176 mg/l (0.8 M) heptanesulfonic acid, and 36 mg/l (0.097 mM) EDTA at a pH of 4.1. The electrochemical detec- tor (Coulochem 5100A, ESA, Bedford, MA) oxidation elec- trode was set at +0.45 V with a flow rate of 1.0 ml/min. Sam- ples were maintained in a refrigerated (4°C) autosampler (WISP 712, Waters Associates, Marlborough, MA) prior to injection. Unknown sample neurotransmitter levels were quantified via a six-point external standard calibration curve. Values below detectable limits were considered to be zero values.
Statistical Analysis
Maternal and pup weights and pup crown-rump lengths were analyzed using Student’s t-test. All data were expressed as mean 2 SEM. The data from HPLC analysis were calcu- lated using the litter as the unit of analysis (all pups from a given litter and condition were collapsed to provide a mean value). A two-way analysis of variance (ANOVA) was used to analyze the results of the HPLC data with the first factor being prenatal drug exposure (cocaine vs. saline) and the sec- ond factor being atmospheric exposure (0.21 Fro, vs. 0.08
Fro,).
COCAINE-INDUCED CAROTID BODY DA CHANGES
RESULTS
Physical Measures
Although slight to moderate tissue necrosis was noted on all cocaine-exposed dams, sufficient undamaged skin areas were accessible for all SC administrations. There were no sig- nificant differences across drug exposures in baseline dam weights (289 t 4.2 g COC vs. 291 + 3.8 g SAL), weight gains throughout pregnancy (53 ? 4.3 g COC vs. 59 ? 3.6 g SAL), or the number of pups per litter (11 2 1.2 COC vs. 10 2 1.5 SAL). The experimental design provided for 32 pups (four pups each from eight litters). Eleven carotid bodies were lost due to procedural errors or violations of the timed protocol and were not analyzed. The remaining carotid bodies (n = 29 COC vs. II = 24 SAL) were analyzed as outlined above.
Pup weights at P5 did not differ across prenatal drug expo- sure conditions (COC 11.0 + 0.3 g vs. SAL 10.8 ? 0.2 g). However, cocaine-exposed pups had a significantly shorter crown-rump length compared to saline-exposed pups (46.1 ? 0.5 mm vs. 48.3 + 0.7 mm, respectively; p < 0.01, two-tailed t-test).
Atmospheric Exposure
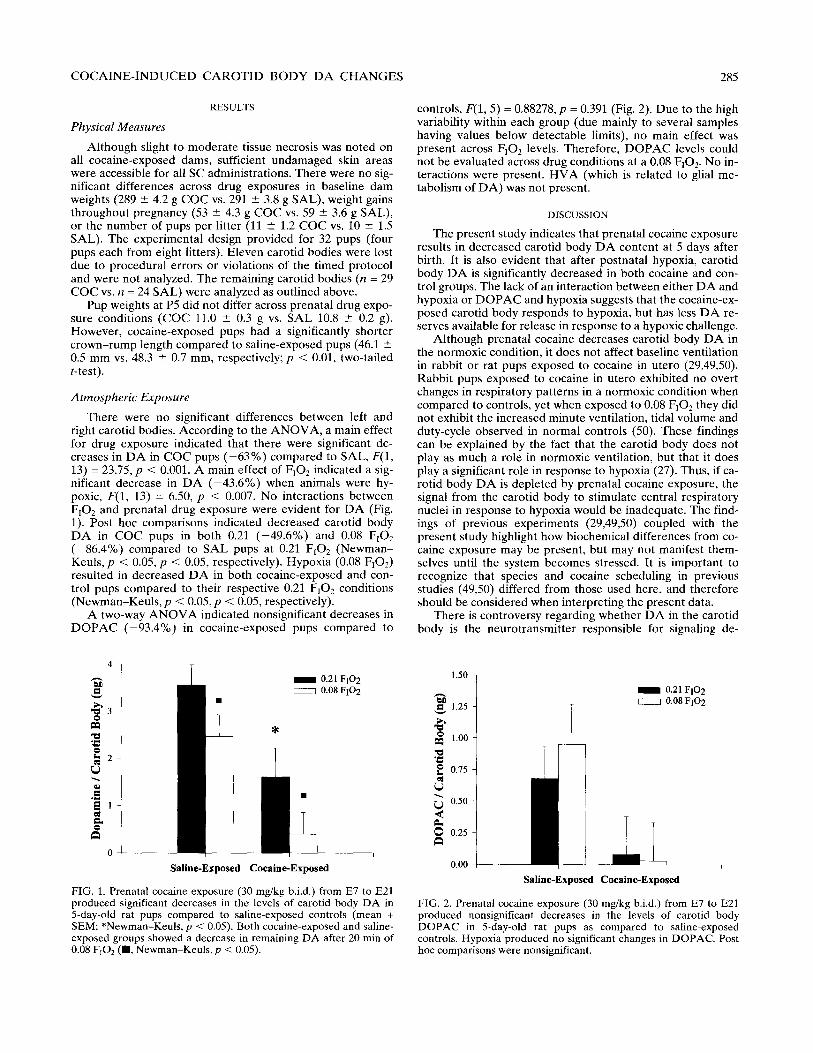
There were no significant differences between left and right carotid bodies. According to the ANOVA, a main effect for drug exposure indicated that there were significant de- creases in DA in COC pups (-63%) compared to SAL, F(1, 13) = 23.75, p < 0.001. A main effect of FiO, indicated a sig- nificant decrease in DA (-43.6%) when animals were hy- poxic, F(1, 13) = 6.50, p < 0.007. No interactions between F,Oz and prenatal drug exposure were evident for DA (Fig. 1). Post hoc comparisons indicated decreased carotid body DA in COC pups in both 0.21 (-49.6%) and 0.08 F102 (-86.4%) compared to SAL pups at 0.21 F,O, (Newman- Keuls, p < 0.05, p < 0.05, respectively). Hypoxia (0.08 FrOJ resulted in decreased DA in both cocaine-exposed and con- trol pups compared to their respective 0.21 F102 conditions (Newman-Keuls, p < 0.05, p < 0.05, respectively).
A two-way ANOVA indicated nonsignificant decreases in DOPAC (-93.4%) in cocaine-exposed pups compared to
Saline-Expo! sed
m 0.21 FIO2 / 0.08 FIO2
Cocaine-Exposed
FIG. 1. Prenatal cocaine exposure (30 mg/kg b.i.d.) from E7 to E21 produced significant decreases in the levels of carotid body DA in 5-day-old rat pups compared to saline-exposed controls (mean + SEM; *Newman-Keuls, p < 0.05). Both cocaine-exposed and saline- exposed groups showed a decrease in remaining DA after 20 min of 0.08 FIOz (m. Newman-Keuls,p < 0.05).
285
controls, F(1, 5) = 0.88278, p = 0.391 (Fig. 2). Due to the high variability within each group (due mainly to several samples having values below detectable limits), no main effect was present across F,Oz levels. Therefore, DOPAC levels could not be evaluated across drug conditions at a 0.08 FrOr. No in- teractions were present. HVA (which is related to glial me- tabolism of DA) was not present.
DISCUSSION
The present study indicates that prenatal cocaine exposure
results in decreased carotid body DA content at 5 days after birth. It is also evident that after postnatal hypoxia, carotid body DA is significantly decreased in both cocaine and con- trol groups. The lack of an interaction between either DA and hypoxia or DOPAC and hypoxia suggests that the cocaine-ex- posed carotid body responds to hypoxia, but has less DA re- serves available for release in response to a hypoxic challenge.
Although prenatal cocaine decreases carotid body DA in the normoxic condition, it does not affect baseline ventilation in rabbit or rat pups exposed to cocaine in utero (29,49,50). Rabbit pups exposed to cocaine in utero exhibited no overt changes in respiratory patterns in a normoxic condition when compared to controls, yet when exposed to 0.08 F102 they did not exhibit the increased minute ventilation, tidal volume and duty-cycle observed in normal controls (50). These findings can be explained by the fact that the carotid body does not play as much a role in normoxic ventilation, but that it does play a significant role in response to hypoxia (27). Thus, if ca- rotid body DA is depleted by prenatal cocaine exposure, the signal from the carotid body to stimulate central respiratory nuclei in response to hypoxia would be inadequate. The find- ings of previous experiments (29,49,50) coupled with the present study highlight how biochemical differences from co- caine exposure may be present, but may not manifest them- selves until the system becomes stressed. It is important to recognize that species and cocaine scheduling in previous studies (49,50) differed from those used here, and therefore should be considered when interpreting the present data.
There is controversy regarding whether DA in the carotid body is the neurotransmitter responsible for signaling de-
1.50
% 3 1.25
% g 1.00
s
g 0.75
4
2 0.50
2
8 o.25
0.00 c
‘I i
I I
Saline-Exposed CocaineExposed
m 0.21 FIO2
i
/ 0.08 FIO2
FIG. 2. Prenatal cocaine exposure (30 mg/kg b.i.d.) from E7 to E21 produced nonsignificant decreases in the levels of carotid body DOPAC in 5-day-old rat pups as compared to saline-exposed controls. Hypoxia produced no significant changes in DOPAC. Post hoc comparisons were nonsignificant.

286
creases in arterial PO2 and whether it is responsible for the signaling of the supposed chemoreceptor, the glomus cell. Retrograde tracing and immunohistochemistry in the guinea pig carotid body found tyrosine hydroxylase-like (TH-LI) cells that did not react with antibodies to other enzymes in- volved in catecholamine synthesis, such as aromatic amino acid decarboxylase, dopamine+hydroxylase, and phenyleth- anolamine-N-methyl-transferase (PNMT) (25), suggesting that the TH-LI neurons lack DA altogether. Yet, exposure of ca- rotid body glomus cells to DA or anoxia revealed increases in their electrical activity (23). Almitrine (which increases venti- lation by stimulating the carotid body arterial chemorecep- tors) increased the resting release of 3H-catecholamines from carotid bodies (2) and further augmented the release of 3H-cate- cholamines from carotid bodies when incubated in a hypoxic solution. These studies as well as many others (1,3,26,27,41) offer strong evidence that the release of DA from carotid bodies is involved in the augmentation of the respiratory re- sponse to hypoxia and that this response is mediated through the action of glomus cells.
Additionally, DA is thought to have an inhibitory effect on respiratory drive. It has been postulated that the tonic inhibi- tory influence of DA could be involved in antagonizing venti- latory drive in chronically hypoxic conditions (i.e., high alti- tudes) to protect against hypocapnia (30). Direct intra-arterial DA infusion diminishes electroneural activity of the hypoglo- ssal, phrenic, and recurrent laryngeal nerves in cats (43). Stud- ies in newborn kittens (aged O-3 days) indicate that IV injec- tions of DA inhibited chemosensory discharge in normoxia (31). Yet it should be noted that DA infusion in the same study had an excitatory effect upon hypoxic kittens. These studies indicate that the inhibitory/excitatory actions of DA in the carotid body are dependent upon species, timing, and dos- age of exogenous DA infusion, as well as the developmental status of the animal studied.
This study’s quantification of carotid body DA using a timed protocol has provided values appreciably higher than those seen in other studies. Values for neonatal rat carotid body DA have previously been reported at about 1.13 ng (about one-third of the values shown here) (18). Quantifica-
LIPTON ET AL.
tion of adult carotid body has ranged from 1.359 to 1.611 ng/ carotid body (13,17,18,37,39). These disparities highlight the need for rapid, timed removal of the carotid body when used for the quantification of neurotransmitter levels to prevent re- duced or inconsistent values. In addition, it must be noted that several DOPAC samples in this study were below detectable limits, especially in the cocaine groups. This was most likely not due to a lack of sensitivity in HPLC-EC detection (our de- tection threshold was 0.25 ngiml for DOPAC and 0.10 ngiml for DA), but is rather an indication that very little DA is me- tabolized into DOPAC in the cocaine-exposed animals, sug- gesting lower DA turnover. Although the levels of DOPAC did not reach significance, the general trend towards de- creased DOPAC in cocaine-exposed animals supports our hy- pothesis of lower DA turnover.
Because of its importance in respiratory circuitry, the ob- served alterations in carotid body DA of cocaine-exposed rat pups likely contribute to the altered compensatory response to hypoxia (29,49,50). When these findings are examined in the context of previous observations of striatal DA deficien- cies in cocaine-exposed pups exposed to hypoxia (51,.52), it appears as though cocaine may be inducing global reductions in DA. If this supposition is true, then one could expect to see several other behavioral and physiological manifestations of cocaine’s teratogenic effects in the developing neonate. Fur- ther study is necessary to determine whether the changes in carotid body DA impact upon the biochemistry of respiratory nuclei in the brain stem. In addition, the exact duration, tim- ing and dosage of prenatal cocaine that are needed to induce such changes should be elucidated along with the specific mechanisms by which cocaine alters the functioning of this pe- ripheral chemoreceptive organ.
ACKNOWLEDGEMENTS
This research was funded in part by grants from the United Par- kinson’s Foundation (P.M.C.) and the National Institute on Drug Abuse (DA 0.5730, J.W.L.). The authors would like to thank Colleen M. Buhrfiend, B.S., for her technical assistance and the National In- stitute on Drug Abuse Research Drug Supply System for supplying the cocaine HCl used in these experiments.
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
Almaraz, L.; Perez-Garcia, M. T.; Gonzalez, C. Presence of Dl receptors in the rabbit carotid body. Neurosci. Lett. 132:259-262; 1991. Almaraz, L.; Rigual. R.; Obeso, A.; Evrard, Y.; Gonzalez, C. Effects of almitrine on the release of catecholamines from the rabbit carotid body in vitro. Br. J. Pharmacol. 106697-702; 1992. Almaraz, L.; Wang, 2. Z.; Stensaas, L. J.; Fidone, S. J. Release of dopamine from carotid sinus nerve fibers innervating type I cells in the cat carotid body. Biol. Signals 2:1&26; 1993. Bjorklund, A.; Falck, B.; Stenevi, U. Classification of monoamine neurones in the rat mesencephalon: Distribution of a new monoamine neurone system. Brain Res. 32:269-285; 1971. Cardenas, H.; Zapata, P. Dopamine-induced ventilatory depres- sion in the rat, mediated by carotid nerve afferents. Neurosci. Lett. 24:29-33; 1981. Carvey, P. M.; Kao, L. C.; Zhang, T. J.; Amdur, R. L.; Lin, D. H.; Singh, R.; Klawans, H. L. Dopaminergic alterations in cotreat- ments attenuating haloperidol-induced hypersensitivity. Pharma- col. Biochem. Behav. 35291-300; 1990. Chasnoff, I. J.; Hunt, C. E.; Kletter, R.; Kaplan, D. Prenatal cocaine exposure is associated with respiratory pattern abnormal- ities [see comments]. Am. J. Dis. Child 143583-587; 1989. Chiba, T.; Doba, N. Catecholaminergic axo-axonic synapses in the nucleus of the tractus solitarius (pars commissuralis) of the
9.
10.
11.
cat: Possible relation to presynaptic regulation of baroreceptor reflexes. Brain Res. 102:255-265; 1976. Church, M. W.; Overbeck, G. W.; Andrzejczak, A. L. Prenatal cocaine exposure in the Long-Evans rat: I. Dose-dependent effects on gestation, mortality, and postnatal maturation. Neuro- toxicol. Teratol. 12:327-334; 1990. Dahlstrom. A.; Fuxe. K. Evidence for the existence of monoam- ine-containing neurons in the central nervous system I. Demon- stration of monoamines in the cell bodies of brain stem neurons. Acta. Physiol. Stand. Suppl. 232:1-55; 1964. Docherty, R. J.; McQueen, D. S. Inhibitory action of dopamine on cat carotid chemoreceptors. J. Physiol. (Lond.) 279:425436; 1978.
13.
14.
15.
Favre, R.; de Haut, M.; Dalmaz, Y.; Pequignot, J. M.; Peyrin, L. Peripheral distribution of free dopamine and its metabolites in the rat. J. Neural Transm. 66:135-149; 1986. Fidone, S. J.; Gonzalez, C. Initiation and control of chemorecep- tor activity in the carotid body. In: Cherniak, N.S., Widdicombe, J.G., eds. Handbook of physiology III; the respiratory system. Baltimore: American Physiological Society; 1986:247-312. Fidone, S. J.; Zapata, P.; Stensaas, L. J. Axonal transport of labeled material into sensory nerve ending of cat carotid body. Brain Res. 124:9-28: 1977.
16. Gawin. F. H. Cocaine addiction: Psychology and neurophysiology

COCAINE-INDUCED CAROTID BODY DA CHANGES 287
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
[published erratum appears in Science 1991 Aug 2;253(5019): 4941. Science 251:1580-1586; 1991. Hanbauer, I.; Hellstrom, S. The regulation of dopamine and noradrenaline in the rat carotid body and its modification by den- ervation and by hypoxia. J. Physiol. (Lond.) 282:21-34; 1978. Hellstrom, S.; Koslow, S. H. Biogenic amines in carotid body of adult and infant rats-a gas chromatographic-mass spectrometric assay. Acta Physiol. Stand. Y3:540-547; 1975. _ Hertzberg. T.: Hellstrom. S.: Lagercrantz. H.: Peauianot, J. M. Development of the arterial chemoreflex and &trove; of carotid body catecholamines in the newborn rat. J. Physiol. (Lond.) 425:211-225; 1990. Hertzberg, T.; Lagercrantz, H. Postnatal sensitivity of the periph- eral chemoreceptors in newborn infants. Arch. Dis. Child 62:1238-1241; 1987. Kalivas, P. W.; Duffy, P. Effect of acute and daily cocaine treat- ment on extracellular dopamine in the nucleus accumbens. Syn- apse 5:48-58; 1990. Kalivas, P. W.; Duffy, P. Time course of extracellular dopamine and behavioral sensitization to cocaine. I. Dopamine axon termi- nals. J. Neurosci. 13:266-275; 1993. Kholwadwala, D.; Donnelly, D. F. Maturation of carotid chemoreceptor sensitivity to hypoxia: In vitro studies in the new- born rat. J. Physiol. (Lond.) 453:461-473; 1992. Kuhar, M. J.; Ritz, M. C.; Boja, J. W. The dopamine hypothesis of the reinforcing properties of cocaine. Trends. Neurosci. 14:299- 302; 1991. Kummer, W.; Gibbins, I. L.; Stefan, P.; Kapoor, V. Catechol- amines and catecholamine-synthesizing enzymes in guinea-pig sensory ganglia. Cell Tissue Res. 261:595-606: 1990. Leitner, L. M. Dopamine metabolism in the rabbit carotid body in vitro: Effect of hypoxia and hypercapnia. Adv. Exp. Med. Biol. 337:183-190; 1993. Li, K. Y.; Ponte, J.; Sadler, C. L. Carotid body chemoreceptor response to prolonged hypoxia in the rabbit: Effects of domperi- done and prbpranoiol. J..Physiol. (Lond.) 430:1-11; 1990. _ Liuski. J.: McAllen. R. M.: Snver. K. M. The carotid chemoreceu- to; input to the respiratory neurones of the nucleus of tractus sdl- itarus. J. Physiol. (Lond.) 269:797-810; 1977. Lipton. J. W.; Davidson, T. L.; Carvey, P. M.; Weese-Mayer, D. E. Prenatal cocaine: Effect on hypoxic, ventilatory responsive- ness in neonatal rats. Respir. Physiol. (submitted). Majcherczyk, S.; Willshaw, P. The influence of hyperventilation on efferent control of peripheral chemoreceptors. Brain Res. 124:561-564; 1977. Marchal, F.; Bairam, A.; Haouzi, P.; Hascoet, J. M.; Crance, J. P.; Vert, P.; Lahiri, S. Dual responses of carotid chemosensory affer- ents to dooamine in the newborn kitten. Respir. Phvsiol. 90:173- 183; 1992.’
. _
Mayock, D. E.; Standaert, T. A.; Guthrie, R. D.; Woodrum, D. E. Dopamine and carotid body function in the newborn lamb. J. Appl. Physiol. 54:814-820; 1983. McCann, E. M.; Lewis, K. Control of breathing in babies of narcotic- and cocaine-abusing mothers. Early Hum. Dev. 27:175-186; 1991. McDonald, D. M. Peripheral chemoreceptors: Structure-function relationships of the carotid body. In: Hornbein, T. F., ed. Lung biology in health and disease. New York: Marcel Dekker; 1981:105-319. Merrill, E. G. The lateral respiratory neurones of the medulla: Their associations with nucleus ambiguus, nucleus retroambigua- lis, the spinal accessory nucleus and the spinal cord. Brain Res. 24:11-28; 1970.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
Minabe, Y.; Ashby, C. R., Jr.: Heyser, C.; Spear, L. P.; Wang, R. Y. The effects of prenatal cocaine exposure on spontaneously active midbrain dopamine neurons in adult male offspring: An electrophysiological study. Brain Res. 586:152-156; 1992. Mir, A. K.; Al-Neamy, K.; Pallot, D. J.; Nahorski, S. R. Cate- cholamines in the carotid body of several mammalian species: Effects of surgical and chemical sympathectomy. Brain Res. 252:335-342; 1982. Mitchell, R. A.; Berger, A. J. Neural regulation of respiration. Am. Rev. Respir. Dis. 111:206-224; 1975. Pequignot, J. M.; Cottet-Emard, J. M.; Dalmaz, Y.; De Haut de Sigy, M.; Peyrin, L. Biochemical evidence for norepinephrine stores outside the sympathetic nerves in rat carotid body. Brain Res. 3671238-243; 1986. Rigual, R.; Lopez-Lopez, J. R.; Gonzalez, C. Release of dopa- mine and chemoreceptor discharge induced by low pH and high PCOz stimulation of the cat carotid body. J. Physiol. (Lond.). 433:51Y-531; 1991. Schamel, A.; Verna, A. Localization of dopamine D2 receptor mRNA in the rabbit carotid body and petrosal ganglion by in situ hybridization. Adv. Exp. Med. Biol. 337:85-91; 1993. Silvestri, J. M.; Long, J. M.; Weese-Mayer, D. E.; Barkov, G. A. Effect of prenatal cocaine on respiration, heart rate, and sudden infant death syndrome. Pediatr. Pulmonol. 11:328-334; 1991. van Lunteren, E.; Haxhiu, M. A.; Mitra, J.; Cherniack, N. S. Effects of dopamine, isoproterenol, and lobeline on cranial and phrenic motoneurons. J. kppl. Physiol. 56:737-745; 1984. Vibert. J. F.: Bertrand. F.: Denavit-Saubie. M.: Huaelin. A. Three dimensional representation of bulbo-pontine &piratory net- works architecture from unit density maps. Brain Res. 114:227- 244; 1976. von Euler, C.; Hayward, J. N.; Marttila, I.: Wyman, R. J. Respira- tory neurones of the ventrolateral nucleus of the solitary tract of cat: Vagal input, spinal connections and morphological identifica- tion. Brain Res. 61:1-22; 1973. von Euler, U. S.; Liljestrand, G.; Zotterman, Y. The excitation mechanism of the chemoreceptors of the carotid body. Stand. Arch. Physiol. 83:132-152; 1939. Ward. S. L.: Bautista. D. B.: Woo. M. S.: Charm. M.: Schuetz. S.: Wachsman, L.; Sehgal, S.; Bean, X. Response; to hypoxia and hypercapnia in infants of substance-abusing mothers. J. Pediatr. 121:704-709; 1992. Ward, S. L.; Schuetz, S.; Kirshna, V.; Bean, X.: Wingert, W.; Wachsman, L.; Keens, T. G. Abnormal sleeping ventilatory pat- tern in infants of substance-abusing mothers. Am. J. Dis. Child 140:1015-1020; 1986. Weese-Mayer, D. E.; Barkov, G. A. Effect of cocaine in early gestation. Physiologic responses to hypoxia in newborn rabbits. km. Rev. Respir. I%. 146589-596; 1993. Weese-Maver. D. E.: Klemka-Walden. L. M.: Barkov. G. A.: Gingras, J: L. Effects of prenatal cocaine on the ventilatory response to hypoxia in newborn rabbits. Dev. Pharmacol. Ther. 18:116-124; 1992. Weese-Mayer, D. E.; Silvestri, J. M.: Lin, D.; Buhrfiend, C. M.; Lo, E. S.; Carvey, P. M. Effect of cocaine in early gestation on striatal dopamine and neurotrophic activity. Pediatr. Res. 34:389- 392; 1993. Weese-Mayer, D. E.; Silvestri, J. M.: Lin, D.; Buhrfiend, C. M.; Ptak, L. R.; Lo, E. S.; Carvey, P. M. Hypoxia after prenatal cocaine attenuates striatal dopamine and neurotrophic activity. Neurotoxicol. Teratol. 16:177-181: 1994.