pre- and post-harvest ecology of fungi causing spoilage of foods and other stored products
TRANSCRIPT

Journal of Applied Bacteriology Symposium Supplement 1989, 11 S-25s
Pre- and post-harvest ecology of fungi causing spoilage of foods and other stored products
J . LACEY Agriculture and Food Research Council Institute of Arable Crops Research, Rothamsted Experimental Station, Harpenden, Hertfordshire AL5 2JQ, U K
1. Introduction, 11s 2. Expression of water availability, 12s 3. The field environment, 13s
3.1 The role of water, 13s 3.2 Temperature, 13s 3.3 Other factors, 13s
4. The storage environment, 14s 4.1 Water availability, 14s 4.2 Temperature, 14s
4.2.1 Temperature and fungal growth, 14s 4.2.2 Temperature resistance of spores, 15s
4.3 Hydrogen ion concentration, 15s 4.4 Gaseous composition, 15s 4.5 Microbial interactions, 15s 4.6 Interactions between fungi and arthropods, 16s 4.7 Chemical preservatives, 16s 4.8 Irradiation, 16s 4.9 Other factors, 16s
5.1 Phytophthora infestans (Mont.) de Bary, 17s 5.2 Rhizopus stolonyer (Ehren: Fr.) Lindner, 17s 5.3 Botrytis cinerea Persoon, 17s 5.4 Cladosporium species 18s 5.5 Alternaria alternata (Fr.) Keissler, 18s 5.6 Fusarium species, 19s 5.7 Penicillium species, 20s 5.8 Aspergillus flavus Link, A. parasiticus Speare, 21s 5.9 Other Aspergillus species, 22s 5.10 Eurotium species, 23s 5.11 Wallemia sebi (Fries) von Arx, 23s 5.12 Byssochlamys species, 23s 5.13 Monascus (Xeromyces) bisporus (Fraser) von Arx, 24s
5. Ecological determinants of spoilage fungi, 17s
6. Conclusion, 24s 7. References, 24s
1. Introduction
The spoilage of food and stored products pre- and post-harvest involves a wide range of fungi that differ greatly in their ecological determinants. They range from plant pathogens, weak pathogens and superficial saprophytes that disfigure and damage foods before harvest to moulds that cause losses of dry matter and quality in store. Some of these may produce toxic metabolites of greater concern to the consumer than any effect on the appearance of the product.
The colonization of aerial plant parts by micro-organisms starts almost as soon as leaves or inflo- rescences are exposed to the air. Bacteria usually colonize first but they are soon followed by yeasts and then by pathogenic and saprophytic filamentous fungi. Filamentous fungi continue to develop throughout the plant’s growth but especially as the plant senesces and seed ripens. Harvest pro- foundly disturbs the ecosystem and marks the transition from the extremes of the field environment

12s J . Lacey to the relatively stable conditions of storage. This is accompanied by a profound change in the composition of the microflora.
Fungi present on plants before harvest have traditionally been referred to as field fungi. Typically these include species of Cladosporium, Alternaria, Epicoccum, Verticillium, Fusarium and phytopatho- genic fungi. The boundary between pathogens and saprophytes is blurred since there is a gradation from leaf surface saprophytes through fungi that can grow endophytically and are sometimes weakly pathogenic, e.g. Alternaria, to strongly necrotrophic pathogens. Field fungi may persist on fresh foods after harvest and may be the cause of disfiguring lesions on fruit and vegetables. In stored products, such as cereal grains, however, they persist only if the grain is dry enough to prevent growth of typical storage fungi. These usually occur in only small numbers before harvest and are chiefly species with Aspergillus and Penicillium anamorphs. As water content increases, fungal growth becomes more vigorous, leading to spontaneous heating of the substrate and the growth of more thermotolerant fungi, including species of Absidia, Rhizomucor and Humicola often accompanied by thermophilic actinomycetes.
However, the concept of field and storage fungi should not be pressed too far. The recognition that field species could somethimes grow in storage led to the designation of an intermediate group, and more recently it has been recognized that storage species, especially Aspergillus Jlaous, often occur in the field in humid tropical areas. Organisms should therefore be classified according to their ecologi- cal requirements, rather than whether they occur in field or store.
Both before harvest and in store the growth of filamentous fungi is determined by the environment, especially by water availability, temperature and gaseous composition; by interactions with other micro-organisms, both pro- and eukaryotes, and with arthropods; and by measures taken for their control. In this paper, the role of these factors will be reviewed and illustrated by reference to a range of fungi that can cause spoilage to foods and stored products. Information has been gleaned from a variety of sources and further information and references may be obtained from Hirst et al. (1965), Lacey (1967, 1980, 1986a, b), Panasenko (1967), Lacey et al. (1983), Hill & Lacey (1983a, b, 1984), Magan & Lacey (1984a, b, c, d, 1985), Cuero et al. (1987a, b, 1988) and Pitt & Hocking (1985).
2. Expression of water availability
Three measures of water availability have been used in microbiology: equilibrium relative humidity (ERH), water activity (aw) and water potential ($). Equilibrium relative humidity (ERH) is the relative humidity of the intergranular air in equilibrium with the water content of the product; a, is the ratio of the vapour pressure over the product to that over pure water at the same temperature and pres- sure; and $ is the free energy of water in a system relative to the free energy of a reference pool of pure, free water of specified mass or volume which is considered to have zero water potential. Equi- librium relative humidity (ERH) and a, are numerically the same but ERH is expressed as a percent- age and a, as a decimal fraction of one. Water potential ($) is measured in units of kilo- or megapascals (MPa) and it can generally be subdivided into osmotic, turgor, matric, and gravitational components. Water activity (a,) has been widely used in systems where osmotic potential is the major component and is related to t,b by the following expression:
I(, = RT/V, log, a,
where R is the ideal gas constant (8.314 J k- ' mol-I), T is absolute temperature (K) and V, is the partial molar volume of water. Water activity (a,) can also account for the matric component but it does not account for turgor or gravitational components. Thus, water potential in a system at equi- librium is everywhere equal but a, is not necessarily so, since it only contributes to two components. The relationship between water content and water availability changes with temperature, whichever measure is used, but $ changes more slowly at high a , than at low. Thus at 20°C, a change from 1.00 to 0.99 a, represents a t,b change of 1.36 MPa while from 0.50 to 0.49 a , is represented by a change of 2.73 MPa. A full explanation of the use of water potential is given by Papendick & Mulla (1986). In much of microbiology, I++ has replaced a, and there is pressure for food and storage microbiology to conform with the recommendations of the Royal Society (1976). In this paper, $ will be used but,

Pre- and post-harvest ecology of fungi 13s for convenience, the equivalent a , at 25°C or, in outdoor situations, relative humidity will also be given.
3. The field environment
Fungi growing on plants are subject to extremes of conditions depending on the nature of the crop and its microclimatic characteristics. Diurnal temperature fluctuations are greatest at the height within the crop at which leaf area index is greatest while the uppermost parts of the plant, e.g. ears of cereal plants, are exposed to the highest wind speeds, lowest vapour pressures and most radiation.
3.1 T H E ROLE OF W A T E R
Water affects the colonization of plants in a variety of ways. Rainwater can disperse spores by splash- ing them from leaf surfaces and soil, by washing leaves and carrying spores to the soil, and so to underground organs, and by providing water for the explosive discharge of ascospores. Dew may also stimulate ascospore discharge and provide conditions for infection while high humidity and, for some fungi at least, the underlying plant cells provide water for continued growth although, with age and senescence, rain and humidity increasingly determine water availability. Drying or alternating high and low humidities may also stimulate active spore release mechanisms in some fungi. Water or its absence may affect the susceptibility of the plant to infection, perhaps through effects on phytoalexins or through premature senescence leading to immaturity of organs at harvest.
Fungi growing on plants generally have a high requirement for water. For germination, superficial saprophytes have minimum $ of -22.4 to -14.5 MPa (85-90% RH), for growth of -17.6 to -11.5 MPa (88-92% RH) and for sporulation of - 14.5 to - 11.5 MPa (9@92% RH) . For pathogens, the lower limits for sporulation range from -14.5 to -2.8 MPa (9&98%RH) with maximum sporu- lation at -4.2 to 0 MPa (97-100% RH). Some phycomycetes may require a film of water for zoo- spore germination and subsequent infection to be successful.
3.2 T E M P E R A T U R E
Most fungi growing on plants before harvest grow well in the temperature range 0 to 30°C although some can grow to 35°C and above and others below freezing. Different parts of the life cycle may respond differently to temperature. For instance in the Peronosporales, there may be different tem- perature optima for sporangial production, germination by zoospores and for germination by germ tubes. Tissue temperatures, through radiation heat loss, may differ markedly from those of the ambient air, the greatest extremes occurring at the height of maximum leaf area index.
3.3 OTHER FACTORS
Organisms growing on plant surfaces and the sporophores of internal fungi are exposed to a variety of other environmental factors affecting their growth and dispersal. They may be affected by fungi- cides or deposits of pollutants. These may have a selective effect, inhibiting some elements of the microflora while allowing others to develop preferentially. Where saprophytes are antagonizing potential pathogens or competing with them for extraneous nutrients from pollen grains or aphid honeydew, their removal could enhance disease. Many examples of fungal interactions on leaf sur- faces have been reported, for instance, spatial competition between Alternaria, Cochliobolus and Acre- monium and between Cladosporium and Epicoccum, also antagonism between each of these groups and between Alternaria and Fusarium. The combined drying effects of lower vapour pressures and greater wind speeds at the tops of plants means that lower parts remain at $ favourable to fungal growth for longer than the tops. This effect is emphasized if the crop lodges, leading to increased incidence of Fusarium and Penicillium spp. Solar radiation may be lethal to spores and perhaps accounts for the frequency of dark-spored fungi in these environments. Pollution and the so-called open air factor may also affect the viability of airborne spores. A knowledge of the aetiology and

14s J . Lacey disease, especially of conditions favouring spore dispersal and infection, may aid the design of suit- able control measures.
4. The storage environment
Conditions during storage are more equable than those in the field and, for some foods, temperature, humidity and/or gaseous composition may be closely controlled. Chemicals may also be used to modify the environment and prevent arthropod or fungal development.
4.1 WATER AVAILABILITY
Fungal activity in stored products is closely controlled by the availability of water. Monascus (Xeromyces) bisporus is the most xerophilic fungus known, growing at JI down to -68.0 MPa (0.6 a,). As $ is increased above this level, an increasing number of fungi can grow and metabolic activity increases until it becomes sufficient to cause spontaneous heating. Greatest tolerance of low $ usually occurs close to the optimum temperature for growth but the minimum $ tolerated may be modified by pH and solute in experimental systems. Spore germination often occurs to a slightly lower minimum $ than mycelial growth, and sporulation to a slightly higher minimum $, although both differences may reflect the very slow growth that occurs at marginal $. A greater $ is usually required for teleomorph formation than for anamorph development. Thus mature teleomorphs of Emericella nidulans are produced only down to -7.1 MPa (0.95 a,) but the anamorphs to -22.4 MPa (0.85
Often the conditions under which fungi form in greatest numbers are those at which they survive or compete best, not at which they grow best. For instance, Eurotiwn spp. (members of the Asper- gillus glaucus group) may predominate in grain at -49.1 MPa (0.70 a,) although their optimum is about -14.5 MPa (0.90 a,). Nevertheless, the predominant species is often a good indicator of conditions in which products have been stored. Safe storage of cereal grains for up to 3 months can be obtained at -45.2 MPa (0.72 a,) but drying to -59.3 MPa (0.65 a,,.) is required for storage for 2-3 years. Commercial trading often takes place at higher $ and can lead to serious losses during shipment. The safe storage period may be decreased if the lot contains many immature green grains or if damp, deteriorated grains are mixed with dry grains to achieve a marketable standard.
Moisture migration, through the movement of water vapour from a warmer to a cooler area where it condenses, is an important cause of deterioration during storage and shipment. It may result from solar heating of one side of the store, from temperature differences between the grain and ambient air, or from arthropod or microbial heating giving hot spots. Thus, grain which had been uniformly dry can develop areas with sumcient $ for microbial development, producing further water by respir- ation, giving rise to further moisture migration.
a,).
4.2 T E M P E R A T U R E
In food and storage microbiology, two aspects of temperature are important: its role in determining microbial growth rates and its use in pasteurization and sterilization of heat-preserved products.
4.2.1 Temperature and fungal growth
Fungi differ widely in the ranges of temperature that allow their growth and in that giving best growth. For instance Penicillium aurantiogriseum can grow over the range -4 to + 35°C while Humi- cola lanuginosa requires 30-60°C. Aspergillus fumigatus has an exceptionally wide range, from 1& 55°C. Most fungi found in stored products, however, thrive within the range 10-40"C and have optima of 2S35"C. Lowering temperature decreases the rate of metabolism and is often used to slow deterioration. Even freezing is no guarantee of safety unless to below -10°C. Low temperature storage gwes fungi a selective advantage over bacteria in stored meat, allowing the development of black colonies of Cladosporium spp. and other species in sub-zero temperatures. Chilling to 1-2°C has

Pre- and post-harvest ecology of fungi 15s
also been used to slow deterioration of moist grain but this can still allow the development of Penicil- lium and Fusarium spp. and the production of their mycotoxins.
Metabolic activity in moist stored products releases heat and, if this is not removed by ventilation, it can give rise to spontaneous heating. Maximum temperatures attained are related to $ and whether heating is due to arthropods or micro-organisms. Insect infestation is independent of $ but requires a minimum of 17°C. Mites require at least -95.4 MPa (0.50 a,) and temperatures above 3°C. Both insects and mites can initiate heating to 40°C. By contrast, fungi can initiate heating at temperatures as low as -6°C and give maxima of up to 6S70"C when thermophilic fungi and actinomycetes become involved.
4.2.2 Temperature resistance of spores
Fungal spores are less resistant to heat than the endospores of bacteria. However, they are a problem in acid foods preserved by pasteurization in which bacteria are unable to grow. The most resistant fungi are ascomycetes, in which the ascospores are far more resistant than the conidia, with Byssoch- lamys spp. and Neosartoryafischeri able to survive 10 min heating at 90°C.
4.3 HYDROGEN ION CONCENTRATION
Most fungi grow well over a pH range of 3 to 8 and have an optimum at about pH 5. They compete poorly with bacteria at pH 7 or above when $ is close to 0 MPa (1.0 a,), but below pH 5, the likelihood of bacterial growth diminishes and fungi gradually predominate. Changing pH may alter the fungal response to other limiting factors, such as $. For instance, decreasing pH from 6.5 to 4.0 decreased the tolerance to low $ by a range of Aspergillus and Penicillium species, especially close to the limits for their growth.
4.4 GASEOUS COMPOSITION
Fungi causing deterioration of stored products are usually considered obligate aerobes but the partial pressure of 0, necessary for their growth is often overestimated. With many fungi, the concentration of 0, in the atmosphere must be decreased to <0.14% before linear growth is decreased by 50%. Increases in CO, concentration to 5-10% can stimulate growth of some species, especially when $ is high, and >15% CO, may be required to halve growth rates. Indeed Penicillium roquefortii may grow with up to 80% CO, providing there is at least 4.2% 0,. As well as differing in their tolerance of low 0, and high CO, concentrations, species may be more greatly affected when both components are altered rather than either alone. Thus, they may respond more to high CO, when 0, concentra- tion is low than when it is close to ambient conditions. The tolerance of P. roquefortii to high CO, levels decreases with decreasing temperature and $ as well with decreasing 0, concentration. Modi- fied atmospheres have been used for the control of both insects and fungi in stored grains but nitro- gen must be oxygen free while CO,/O, regimes suitable for controlling insects are insufficient to control fungi.
4.5 MI C ROB1 AL INTER ACTIONS
Fungi rarely occur in a monospecific culture in stored products but more often as a group of inter- acting species of fungi and bacteria. The species concerned and the nature of their interactions may alter with temperature, $, gas composition and other factors to change the predominant species. Interactions on agar range from a mutual intermingling of the hyphae with no effect on growth rate, through mutual inhibition on contact or at a distance, to inhibition of one organism, also on contact or at a distance, while the other continues to grow through the colony at the same or a slower rate. By giving these interactions numerical values, indices of dominance (ID) can be derived for groups of organisms interacting in pairs. The largest ID (most competitive or antagonistic species) were found with P. brevicompactum, P . hordei, P . roquefortii, A. fumigatus and A. nidulans. Decreasing $ from

16s J . Lacey
-2.8 MPa (0.98 a,) to -7.1 MPa (0.95 a,) increased the I, of P. breoicompactum but decreased that of most other species, while a further decrease to - 14.5 MPa (0.90 a,) decreased the I , of all fungi. All Penicillium spp. tested and A. nidulans gave a high I, against 'field' fungi. Only F . culmorum could compete well with storage species. However, a stored product is more complex since it may be composed of a series of microniches with little or no diffusion of metabolites between them. Conse- quently the effects of adding mixed inocula to grain may not relate well to I,.
4.6 I N T E R A C T I O N S B E T W E E N F U N G I A N D A R T H R O P O D S
Insects are important agents of stored product deterioration in their own right but they also interact closely with fungi. Insects can enhance fungal growth through the damage they cause, e.g. feeding damage by Sitophilus; through metabolic activity increasing $ and temperature; and through dis- semination of spores. They can inhibit fungal growth by the secretion of quinones (e.g. by Tribolium castaneum) or by feeding on fungi. Conversely, fungi can promote insect growth by acting as feed- stuffs supporting the complete life-cycle, e.g., P. citrinum and Ahasoerus aduena or they can inhibit growth through their metabolites, e.g. aflatoxin retards larval growth of Corcyra cephalonica. However, although T-2 toxin inhibits egg hatch and retards larval growth in Tribolium confusum, it increases fecundity. Fungi, e.g. Penicillium spp., may attract insects while others, e.g. Aspergillus spp., may repel them, although the same species are still attractive to mites (Dunkel 1988).
4.7 C H E M I C A L PRESERVATIVES
Infection of fruit and vegetables during storage may be prevented or decreased by the use of fungi- cides before harvest or as dips before storage, and growth of moulds in foods and stored products may be controlled by using weak acids such as benzoic, sorbic, propionic, acetic, nitrous or sul- phurous acids. However, the small concentrations permitted in foods may be effective only to one pH unit above their pK,. At higher pH, there is too little undissociated acid for any effect on mould growth. The quantity of propionic acid added to stored products must be related to their water content if moulding is to be prevented. In hay this quantity is 1.3 g propionic acid per 100 g water and a similar level will be required in other products, unless the pH differs markedly from that of hay (about pH 6). The acid must also be distributed uniformly through the bulk if fungi tolerant of propionic or other acids are to be controlled. Otherwise these may grow in un- or under-treated pockets, may metabolize the acid and allow growth of less tolerant species, e.g. Eurotium spp., Paecil- omyces uariotii, A . fraous and P. roquefortii. Sorbic acid has even been suggested for the control of both insects and fungi. Besides fungicides and fungistats, protection against post-harvest fungal dete- rioration may also be given by chemicals that inhibit sprout growth, as on potatoes, carrots and onions. Fumigants against insects may also control fungi.
4.8 I R R A D I A T I O N
Foods may be preserved against post-harvest fungal deterioration by irradiation but species differ in their susceptibility to irradiation so that undesirably large doses may be required. For instance, doses of 0.3 KGy are required to eliminate Penicillium spp. from maize seeds, 1.2 KGy for Aspergillus spp., 6.0 KGy for Bacillus spp. and 12 KGy for yeasts and Fusarium spp. (Cuero et al. 1986). Inadequate irradiation could lead to the growth of fungi able to produce mycotoxins and their production could be enhanced by interaction with other fungi, yeasts or bacteria that also survive. Reports differ however about whether more or less aflatoxin is produced when grain is irradiated rather than autoclaved perhaps as a consequence of such interactions or of different doses of irradiation.
4.9 OTHER F A C T O R S
Knowing when products become contaminated with spores and of the ways in which those spores are dispersed may identify the time and place where control measures can be employed most effectively.

Pre- and post-harvest ecology of fungi 17s Fungi may be brought into store on plant surfaces or they may be deposited from the air during post-harvest handling, perhaps from spores from a previous crop; they may be present in the harvest- ing machinery, e.g. combine harvesters for grain; or they may contaminate the water used to wash fruit or vegetables but their dispersal may be restricted by packing materials which may also prevent infection by decreasing bruising or as a result of incorporating chemical control agents.
5. Ecological determinants of spoilage fungi
5.1 P H Y T O P H T H O R A INFESTANS (MONT.) DE BARY
Temperature ("C) i,h (for growth, MPa)
Range Optimum Minimum Optimum
Phytophthora infestans 4 to 26 20 - 22.4 0 (Mont.) de Bary (85% RH) (100% RH)
Phytophthora infestans causes late blight of potatoes, a necrotic disease that can infect all parts of the plant before harvest but which can also cause further spoilage in store. Initially, the leaves are infect- ed from sporangia formed on shoots that arise from infected seed tubers. Sporangia require a minimum II/ of -7.1 MPa (95% RH) to form and are dispersed through the air, or more rapidly, in rain splash. Rain also washes sporangia from the leaves into the soil to infect tubers and provides suitable conditions for their germination by zoospores and for infection of leaves and tubers. Germi- nation of zoospores occurs between 6 and 15°C but sporangia can also germinate by germ tubes at higher temperatures, up to 30°C. Tuber infection can occur following only 5 mm rain and after 5% of the haulm has been destroyed, growth of tubers can be counterbalanced by increased losses from blight. Fungicide treatment may increase tuber disease by prolonging the life of the haulm and allow- ing more occasions when rain can wash spores down to the tubers. Soil can remain infective for at least 3 weeks after the haulm is destroyed so that wound infection can occur if tubers are harvested too soon. A proportion of infections at harvest may be restricted to arrested lesions a few mm long that may develop further in store, perhaps in response to changes in tuber physiology. High humidity in store may allow infected tubers to sporulate through lenticels to infect neighbouring tubers and secondary soft rotting by bacteria may also develop.
5.2 R H I Z O P U S STOLONIFER (EHREN: FR.) LINDNER
Temperature ("C) i,h (for growth, MPa)
Range Optimum Minimum Optimum
Rhizopus stolonifer 5 to 34 26 to 29 -10.0 0 (Ehren : Fr.) Lindner (0.93 a,) (1.00 a,)
Rhizopus stolonifer is important as a destructive rot of many vegetables and fruits and as a food mould. Its pectinases are heat stable, resisting pasteurization and causing the softening and disinte- gration of heat-sterilized fruits. Like other Mucorales, it is reported to grow slowly in anaerobic conditions but it requires >0.5% 0, for sporulation. The fungus is ubiquitous and infection of fruit occurs through wounds. Disease may thus be controlled by avoiding injury and bruising, by cooling rapidly to < 10°C and by treatment with moist air at 44°C for 40-60 min. At low temperatures it may be replaced by Sclerotinia spp.
5.3 B O T R Y T I S C I N E R E A P E R S O O N
Temperature ("C) i,h (for growth, MPa)
Range Optimum Minimum Optimum
Botrytis cinerea Persoon -2 to 35 22 to 35 -10.0 to -7.1 0 (0.93 to 0.95 a,) (1.00 a,)

18s J . Lacey
Botrytis cinerea is an ubiquitous saprophyte that often causes rots of soft fruit before and after harvest. Growth can occur at pH 2-8 and with only 1 % 0,. It may be controlled in the same way as R. stolonij'er, by decreasing humidity and temperature and by decreasing 0, to >0.5%, although off flavours may develop with 0.25% 0, or less.
5.4 C L A D O S P O R I U M SPECIES
Temperature ('C) $ (for growth, MPa)
Range Optimum Minimum Optimum
Cladosporium cladosporioides - 5 to 32 24 to 25 - 17.6 0 (Fres.) de Vries (0.88 a,) (1.00 a,)
(Pers.) Link (0.85 a,) (0.95 a,) Cladosporium herbarum -10 to 32 25 - 22.4 -7.1
Cladosporium species are among the most abundant components of the daytime summer air spora. They are widespread on the ears of cereals at harvest and reported from many other substrates. Their ability to grow at low temperatures allows growth on refrigerated and frozen products such as cheese and meat. Growth of C . herbarum is halved when 0, is decreased to 0.7 or CO, increased to 13% and that of C . cludosporioides is halved at 1.3% and > 15% CO, . Cladosporium herbarum is reported to sporulate with only 0.25% 0,.
5.5 A L T E R N A R I A A L T E R N A T A ( F R ) K E I S S L E R
Temperature ("C) IJ (for growth, MPa)
Range Optimum Minimum Optimum
Alternnria alternata (Fr.) 0 to 35 20 to 25 - 17.6 0 Keissler (0.88 a,) (1.00 a,)
Alternaria alternata is, after Cladosporium spp., perhaps the most common airborne fungus spore over much of the world. It is also a common component of the phylloplane mycoflora of most plants. In cereal crops, leaves and grains become colonized soon after emergence and sub-epidermal penetra- tion may soon occur so that the fungus is able to tolerate fungicidal applications and can be isolated from most grain at harvest. Alternaria alternata often occurs with other Alternaria spp. It may be weakly pathogenic and has been implicated in the conspicuous black or brown discoloration of cereal grains known as black point, causing subsequent discoloration of flour. It competes spatially with other fungi on the plant surface and antagonistically with Cladosporium, Epicoccum and Fusarium, perhaps explaining its decreased incidence when Fusarium culmorum is abundant, although hyphae intermingle freely in agar culture. After harvest, A. alternata survives in cereal grains as long as these are stored well (<59.3 MPa, 0.65 a,) but in wetter grain, when fungi typical of storage can grow, Alternaria soon dies.
Alternaria alternata is of concern because it produces several mycotoxins, including tenuazonic acid, alternariol, alternariol monomethyl ether, altenuene and altertoxins, of which the first is prob- ably the most toxic while others may be mutagenic. In sorghum in the USA, the occurrence of Alternaria toxins correlated with the degree of grain discoloration by fungi and the number of rain- days in September and October (Seitz et al. 1975). The toxins have also been found in pecan nuts but tomatoes are the most important source. Fruit infection by A. alternata can lead to severe losses of tomato fruits. Infection is favoured by free moisture from rain or dew and at suboptimal tem- peratures ( < 15OC) was stimulated by water-soluble nutrient exudates. Infection of ripe fruit results in large sunken lesions with heavy sporulation but that of green fruits gives quiescent lesions that fail to enlarge with ripening. Sporulation is optimal at 27°C but is inhibited below 15°C or above 33°C (Pearson & Hall 1975). Growth is halved with 2.8% 0, or > 15% CO,.

Pre- and post-harvest ecology of fungi 5.6 F U S A R I U M SPECIES
Temperature ("C)
19s
$ (for growth, MPa)
Range Optimum Minimum Optimum
Fusarium avenaceum
Fusarium culmorum
Fusarium graminearum
Fusarium moniliforme
Fusarium poae
Fusarium sporotrichioides
Fusarium subglutinans
(Corda ex Fr.) Sacc.
(Smith) Sacc.
Schwabe
Sheld.
(Peck) Wollenw.
Sherbakoff
(Wollenw. & Reinking) Nelson et al.
(Corda) Sacc. Fusarium tricinctum
- 3 to 31
< O to 31
nd
2 to 37
2 to 39
-2 to 35
2 to 38
<o to 35
25
25
24 to 26
22 to 28
22 to 28
22 to 28
22 to 28
25
- 16.0 (0.89 a,)
(0.89 a,)
(0.89 a,)
(0.87 a,)
(0.89 a,)
(0.88 a,)
- 16.0
- 16.0
- 19.2
- 16.0
- 17.6
nd
- 16.0 (049 a,)
0 (1.0 0,) - 1.4
(0.99 a,)
(0.99 to 0.98 a,) - 1.4 to -2.8
nd
0 (1.0 a d
nd
nd
0 (1.0 a,)
Fusarium spp. are important pathogens of cereals, causing root and stalk rots and ear infections variously referred to as scab, ear rot or head blight. They may also form such mycotoxins as zearale- none and various trichothecenes in the grain either before or after harvest. The species found on wheat are chiefly F. culmorum, F. graminearum and F. avenaceum and on maize F. graminearum, F. monilijiorme and F. subglutinans. Fusarium sporotrichioides may produce lesions on wheat glumes but does not infect kernels (Vargo & Baumer 1986). The epidemiology of F. graminearum on wheat has been studied in greatest detail, chiefly in Canada (Sutton 1982). The fungus persists in host debris in the soil from which it is dispersed as ascospores and rain-splashed macroconidia. Ascospore dispersal is mainly at night and requires 95-100% RH and temperatures of 11-23°C. Insects and birds may assist the dispersal of conidia. Wheat spike infection may be favoured by the presence of anthers or pollen. These provide nutrients and may be intensely colonized before infection occurs. The spikes are thus most susceptible to infection from anthesis to soft dough stage. Infection is favoured by temperatures of 2&30°C and surface wetness that persists for 48-60 h. Temperatures < 1 5 T or surface wetness for <24 h prevent most infection. Macroconidia can be produced within 3 d of inoculation at 2530°C and in 5 d at 20°C. Fields infected by either F. culmorum, F. graminearum or by Microdochium (Fusarium) niuale may occur in clusters. With centre pivot irrigation systems, most sporulation occurred close to the pivot where conditions were probably wettest (Strausbaugh & Maloy 1986). Infection of wheat in Britain, as in Canada, is favoured by wet weather at anthesis but the chief species is F. culmorum. This is also favoured by lodging of the crop. F. avenaceum, F. poae and F . tricinctum may also be isolated from ripening wheat and barley grain and F. avenaceum from lupin pods and seeds.
Maize is most susceptible shortly after silk emergence. Infection is again closely related to rainfall with most, in a Canadian study, when 70-80 mm fell on 6-9 d during silking than when < 60 mm fell on 1&16 d. Insect and bird damage can enhance infection. Colonization continues for 8 weeks or more beneath the husk and during storage of husked ears in cribs until it is limited by water content falling to 20-22% ($ ca - 14.5 MPa, 0.90 a d . There may be a relationship between the occurrence of wheat head blight and maize ear rot epidemics. Fungal propagules and ergosterol and deoxy- nivalenol (DON) concentrations all increased similarly for six weeks, following inoculation of maize with F. graminearum, but propagules and DON then declined while ergosterol remained constant. Zearalenone appeared 9 weeks after inoculation (Miller et al. 1983). Zearalenone may sometimes be translocated from stem lesions to the ear.
Fusarium moniliforme and F. subglutinans are also commonly associated with maize but have differ- ent climatic preferences. In the South African Transvaal, F . moniliforme predominates in subtropical
20s J . Lacey areas and F. subghtinans in temperate areas. In the intermediate area, F. graminearurn was the pre- dominant Fusarium sp. Fusarium sporotrichioides was an important cause of alimentary toxic aleukia in the USSR after it had extensively colonized millet overwintered under snow in mild years with a heavy snowfall and slow thaw with repeated thawingjfreezing cycles. Severe winters with less snow and a rapid thaw allowed little infection.
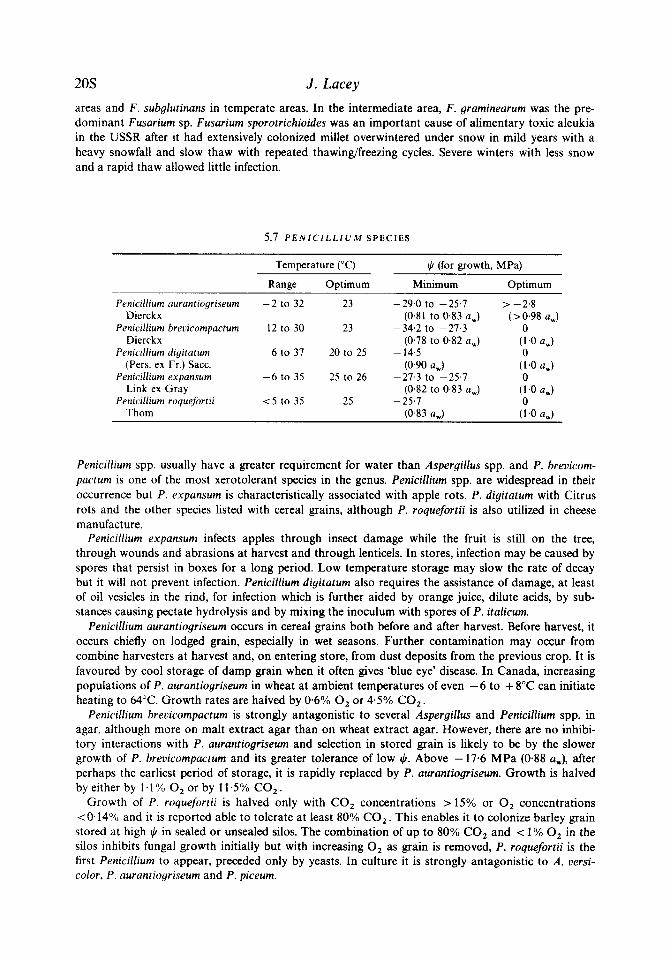
5.7 P E N I C I L L I U M SPECIES
Penicillium aurantiogriseum Dierckx
Penicillium brevicompactum Dierckx
Penicillium digitarum (Pers. ex Fr.) Sacc.
Penicillium expansum Link ex Gray
Penicillium roquefortii Thom
Temperature ("C) i,h (for growth, MPa)
Range Optimum
-2 to 32 23
12 to 30 23
6 to 31 20 to 25
-6 to 35 25 to 26
< 5 to 35 25
Minimum
- 29.0 to - 25.1 (0.81 to 0.83 a,)
(0.78 to 0.82 a,)
(0.90 a,)
-34.2 to -21.3
- 14.5
-21.3 to -25.7 (0.82 to 0.83 a,)
- 25.7 (0.83 a,)
Optimum
> -2.8 (> 0.98 a,)
0 (1.0 a,) 0
(1.0 a,) 0
Penicillium spp. usually have a greater requirement for water than Aspergillus spp. and P. brevicom- pactum is one of the most xerotolerant species in the genus. Penicillium spp. are widespread in their occurrence but P. expansum is characteristically associated with apple rots. P . digitatum with Citrus rots and the other species listed with cereal grains, although P. roquefortii is also utilized in cheese manufacture.
Penicillium expansum infects apples through insect damage while the fruit is still on the tree, through wounds and abrasions at harvest and through lenticels. In stores, infection may be caused by spores that persist in boxes for a long period. Low temperature storage may slow the rate of decay but it will not prevent infection. Penicillium digitatum also requires the assistance of damage, at least of oil vesicles in the rind, for infection which is further aided by orange juice, dilute acids, by sub- stances causing pectate hydrolysis and by mixing the inoculum with spores of P . italicum.
Penicillium aurantiogriseum occurs in cereal grains both before and after harvest. Before harvest, it occurs chiefly on lodged grain, especially in wet seasons. Further contamination may occur from combine harvesters at harvest and, on entering store, from dust deposits from the previous crop. It is favoured by cool storage of damp grain when it often gives 'blue eye' disease. In Canada, increasing populations of P. aurantiogriseum in wheat at ambient temperatures of even - 6 to + 8°C can initiate heating to 64°C. Growth rates are halved by 0.6% 0, or 4.5% CO,.
Penicillium brevicompactum is strongly antagonistic to several Aspergillus and Penicillium spp. in agar, although more on malt extract agar than on wheat extract agar. However, there are no inhibi- tory interactions with P. aurantiogriseum and selection in stored grain is likely to be by the slower growth of P. brevicompactum and its greater tolerance of low G. Above - 17.6 MPa (0.88 aw), after perhaps the earliest period of storage, it is rapidly replaced by P . aurantiogriseum. Growth is halved by either by 1.1% 0, or by 11.5% CO,.
Growth of P. roquefortii is halved only with CO, concentrations >15% or 0, concentrations < 0 . 1 4 O / O and it is reported able to tolerate at least 80% CO,. This enables it to colonize barley grain stored at high $ in sealed or unsealed silos. The combination of up to 80% CO, and < 1 % 0, in the silos inhibits fungal growth initially but with increasing 0, as grain is removed, P. roquefortii is the first Penicillium to appear, preceded only by yeasts. In culture it is strongly antagonistic to A. oersi- color, P. aurantiogriseum and P . piceum.

Pre- and post-harvest ecology of fungi
5.8 A S P E R G I L L U S F L A V U S L I N K , A . P A R A S I T I C U S S P E A R E
21s
Temperature ("C) (I (for growth, MPa) Range Optimum Minimum Optimum
AspmgillusJIauw Link and 6 to 45 35 to 37 -34.2 -7.1 Asperigillus parasiticus Spare (0.78 aw) (0.95 a,)
Aspergillus flauus and A. parasiticus gained prominence in food and storage mycology following the recognition of their ability to produce aflatoxins. Initially, the fungi were considered typical storage fungi and aflatoxins only a storage problem. However, colonization of maize, groundnuts and cotton- seed and aflatoxin production in the field is now proven. In the southeastern USA, maize ears are chiefly infected from late milk to dough growth stages, especially if the plants are drought and tem- perature stressed and insect damaged. Aspergillus f laws is the predominant species and can form > 90% of A. flauus group colonies isolated. Zinc is required for aflatoxin formation and infection of maize germs may be associated with increased trace element levels. However, accompanying high levels of phosphate suggest much of this may be unavailable to the fungus. Overwintering probably occurs as sclerotia that fall to the ground from the ears and subsequently produce sporophores.
Groundnuts may yield A. parasiticus more frequently than other crops but infection is again enhanced by drought and temperature stress although neither drought stress nor high temperatures alone stimulated aflatoxin production. Undamaged groundnuts, grown with - 1.5 to -2.2 MPa and geocarposphere temperatures of 26.3-29.6"C, were highly contaminated with aflatoxins but those grown at 25.7 and 31.3"C were not. Aspergillus jauus group could be isolated from many kernels, even with irrigation. Most edible grade groundnuts were infected when drought-stressed plants were grown with geocarposphere temperatures close to 30°C.
Cottonseed infection occurs in the high temperature growing areas of the USA. Infection may be enhanced by bollworm infestation but the critical period for toxin formation is a 30-45 d period following the opening of the first bolls. At this time, storms disturb the desert raising 'sand devils' that could drive spores with soil into the bolls. Six days is necessary for infection to penetrate from simulated insect damage of the carpel walls to the seed but drying is then necessary for the fungus to penetrate the seed (Lee et al. 1986, 1987).
Raisins are heated to 45-50°C during solar drying. The germinating spores of A.jauus are sensitive to these temperatures and aflatoxin production is inhibited (Hussein et al. 1986).
In store, A . flauus colonization and aflatoxin production are controlled by $, temperature, inter- granular gas composition, chemical preservatives and microbial interactions. These have usually been studied in isolation, neglecting possible interactions between different factors. Even without pre- harvest colonization, it is iikely spores enter store with the grain. Heating during drying is unlikely to affect viable inoculum levels as dormant spores can tolerate up to 110°C for 3 min but are killed by 60 min at this temperature (Kingsland 1986). Subsequently, total heat input is important in determin- ing growth, sporulation and toxin production. Growth requires at least 208 degree hours per day and sporulation, with toxin production, 270 degree hours per day. The optimum was 772 degree hours per day and between minimum and optimum, temperature cycling decreased the time before sporu- lation and toxin formation (Stutz & Krumperman 1976). Later studies (Park & Bullerman 1981, 1883) suggest that the relationship is more complex. Initiation of growth, sporulation and toxin production when temperatures alternated every 12 h between 5 and 25°C were faster than at 15°C but slower than at 18 or 25°C. With A . parasiticus, however, production of aflatoxin with cycling temperatures was greater than at 15, 18 or 25°C but with A.flauus it was greater only than at 15°C. Other studies with cheese as the substrate, suggest greater aflatoxin production with a constant 25°C than with cycling temperatures; spore germinntion, growth and toxin production being proportional to thermal input during the first 5 d incubation; and significant aflatoxin production occurring at refrigeration temperatures.

22s J . Lacey
Growth and aflatoxin production is inhibited by 40% COz at - 1.4 MPa at 15°C but aflatoxin was produced under these conditions at 25°C. At 3WC, aflatoxin was produced in small amounts with up to 60% CO, and down to 1% 0, but a combination of 5% 0, with 20% CO, inhibited almost all aflatoxin production. Treatment with 3% CO, inhibited but did not prevent fungal growth if 0, concentrations were small but no aflatoxin was produced unless there was 1.5% 0, (Wilson & Jay 1976; Buchanan et al. 1985).
Growth of A.Jauus is suppressed by interactions with Rhizopus stolonifer, Saccharomyces cerevisiae and Brevibacterium linens but stimulated by Acetobacter aceti. With Streptococcus lactis inoculation 3 d before A. parasiticus decreased growth of the mould while inoculation afterwards caused stimu- lation. Aspergillus niger did not affect sporulation of A. javus in experiments but aflatoxin production was inhibited. However in maize grain, aflatoxin was formed when the ratio of propagules of A. javus to A. niger exceeded 19:l but not when it was less than 9:l . Such interactions can be affected by $ with aflatoxin production decreased by A. niger, Penicillium viridicatum and Fusarium graminearum at -2.8 MPa but stimulated at -7.1 (0.98 a,) and - 14.5 MPa (0.95 and 0.90 a,). With a yeast, Hyphopichia burtonii, and a bacterium, Bacillus amyloliquefaciens, toxin production was increased at all $.
Substrate irradiation before inoculation may affect aflatoxin production but not growth of A.flavus although the effects are not clear. Doses of 7.5, 100, 250 and 400 KGy increased aflatoxin production in wheat and other cereals but 120 KGy decreased production in rice, compared to ethylene oxide sterilization and autoclaving.
Chemicals have been used to decrease colonization of cereal grains. Fungicides such as captan, metiram and thiram were effective at 250 ppm at pH 3-7, Topsin-M at 250 ppm and Imazalil at 5 ppm at pH 7 and sodium propionate at pH 3. Dichloran gave 60-70% inhibition at all pH values. Propionic acid is effective so long as the dose is closely related to the water content of the substrate and it is uniformly applied. Aspergillis flavus appears tolerant of propionate, colonizing inadequately treated barley and enhancing aflatoxin production.
5.9 O T H E R A S P E R G I L L U S SPECIES
Temperature ("C) $ (for growth, MPa)
Range Optimum Minimum Optimum
Aspergillus candidus Link 3 to 44 25 to 32 -39.6 -14.5 to -2.8
Aspergillus fumigatus Fres. 10 to 55 40 to 42 -22.4 -2.8 to -1.4
Aspergillus restrictus G. Smith 9 to 40 30 -47.1 - 5.6
Aspergillus versicolor (Vuill.) 4 to 39 25 to 30 -34.2 -7.1
(0.75 a,)
(0.85 a,)
(0.71 a,) (0.96 a,)
Tiraboschi (0.78 a,) (0.95 a,)
(0.90 to 0.98 a,)
(0.98 to 0.99 a,)
Aspergillus spp. are characteristic colonizers of stored products. The species differ considerably in their requirements so that the predominant species is often a good indicator of previous storage conditions. Thus, A. fumigatus is found in grain that has been stored wet and has heated while A. restrictus predominates in grain stored only slightly above a safe ~ with no heating in which its slow growth may prevent its detection before 3 months. Care is required when handling products colo- nized by A.fumigatus because it is potentially pathogenic to man. Spores of A. candidus will survive 10 min at 50°C but not at 60°C while not all isolates are able to grow at 40°C. Linear growth of A. candidus, A. fumigatus and A. versicolor is halved by, respectively > 15, > 15 and 12% CO, or by 0.45, 3.4, and 6.4% 0,.

Pre- and post-harvest ecology of fungi 5.10 E L I R O T I U M SPECIES
23s
Temperature (“C) $ (for growth, MPa)
Range Optimum Minimum Optimum Eurotium amstelodami Mangin 5 to 46 33 to 35 -47.1 to -43.3 - 10.0 to -5.6
Eurotium chevalieri Mangin 5 to 43 30 to 35 -47.1 -10.0 to -7.1 (0.71 to 0.73 a,) (0.93 to 0.96 a,)
(0.71 a,) (0.93495 a,) Eurotium repens de Barv I to 40 25 to 27 -39.6 -10.0 to -7.1
(0.75 a,)
(0.70 a,) (0.93 a,)
(0.93 to 0.95 a,) Eurotium rubrum Konig et al. 5 to 40 25 to 27 -49.1 - 10.0
Like A. restrictus, Eurotium spp. (perhaps better known as members of the Aspergillus glaucus group) are xerophilic with maximum growth rates at JI < 0 MPa (1.0 a,). They occur widely and are often numerous in hay, grain and other products with little available water. They differ in the maximum JI allowing the development of anamorph and teleomorph stages, from -39.6 to -34.2 MPa (0.75- 0.78 a,) for anamorph production to -36.0 to -20.8 MPa (0.77-0.86 a,) for teleomorph pro- duction. The ascospores, especially of E. cheualieri are also heat resistant. From 18-25% of E. cheualieri ascospores survive 10 min at 70°C compared to 0.5-3% of ascospores of other Eurotium spp. A decimal reduction time at 80°C (DsoOc) of 3.0 min has been calculated for E. chevalieri. At high JI, linear growth of E. repens is halved by > 15% CO, or by 0.6% 0,.
5.11 W A L L E M I A S E B I ( F R I E S ) VON A R X
Temperature (“C) $ (for growth, MPa) Range Optimum Minimium Optimum
Wallemia sebi (Fries) von >5 to 35 23 to 25 -51.1 -7.1 Arx (0.69 a,) (0.95 a,)
Wallemia sebi is an important cause of deterioration of dried and salted fish but it is xerophilic rather than halophilic and occurs in a wide range of other products, including cereal grains, other seeds, hay and spices. It is not often reported, however, because of its slow growth at JI close to 0 MPa. Its tolerance of low JI is dependent on solute, having a minimum JI of only -39.6 MPa (0.75 a,) when NaCl is used for control. However solute has little effect on the optimum JI for growth. No growth occurs at pH 3, pH 6-7 being optimal. A minimum JI of - 36.0 MPa (0.77 a,) has been reported for sporulation.
5.12 B Y S S O C H L A M Y S SPECIES
Temperature (“C) JI (for growth, MPa) Range ODtimum Minimum ODtimum
B.fulua Olliver 10 to 45 30 to 35 nd nd
€3. nivea Westling 10 to 40 30 to 35 -24.0 to - 16.0 nd (0.84 to 0.89 a,) nd
& Smith
Byssochlamys fulva and B. nivea are particularly important in the spoilage of heat-processed foods because of the heat resistance of their ascospores and their ability to grow with as little as 0.27% 0, to produce CO, . Decimal reduction times for B. fulua at 90°C (DgoOc) range between 1 and 12 min, depending on strain, pH, JI and the presence of preservatives or high levels of sugar. Susceptibility to heat is increased by low pH and SO, and decreased by sugar. Only ascospores of Neosartorya Jischeri (Wehmer) Malloch & Cain are more heat resistant but this species is seldom a problem.

24s J . Lacey
Byssochlarnys nivea is more often a problem in Europe than other parts of the world. It has asco- spores which are less heat resistant giving decimal reduction times at 75°C (D,5-c) of about 60 min. Ascospore germination is stimulated by a heat shock of 75°C for 5 min and is enhanced by p H 4-45 and aeration. Inoculum of Byssochlarnys spp. probably originates from soil although it has seldom been found. They have also been associated with the aerobic deterioration of silage, especially after formaldehyde treatment and may produce patulin.
5.13 W O N A S C U S ( X E R O M Y C E S ) B I S P O R U S ( F R A S E R ) V O N A R X
Temperature (-C) $ (for growth, MPa)
Range Optimum Minimum Optimum Maximum
Monascus (Xeromyces) bisporus nd ca 25 - 69.0 - 22.4 - 4.2 (Fraser) von Arx (0.61 a,) (0.85 a,) (0.97 a,)
Monascus bisporus is the most xerophilic fungus known and is important as a spoilage mould when other fungi are restricted by low t,b. It has commonly been isolated from dried prunes and a range of other foods in Australia and Britain. It is remarkable in producing a teleomorph at a t,b lower than most fungi can grow (-55.1 MPa, 0 . 6 7 ~ ~ ) . Its limited recorded distribution is more likely due to inadequate isolation methods than to its absence. Tolerance of low t,b is much affected by solute: growth is fastest on media containing glucose/fructose and least on NaC1. Spores germinate between - 19.2 and -5.6 MPa (0.87-0.96 n,) at pH 4 and not at all at pH 6.5 when NaCl is used to control $. Ascospores are heat resistant, although less so than Byssochlarnys fulua. Some survive 2 min at 90-C and give a D,,-, of 2.7-3.6 min. Monascus bisporus is able to grow in atmospheres containing 95% CO, and 1%) 0,.
6. Conclusion
The nature of spoilage, the fungi concerned and their tolerance of other fungi and environmental conditions differ widely with food or stored product. For each product, the risk of deterioration and the methods that can be used for their control depend on a knowledge of the ecological requirements of the fungus and its interactions with other micro-organisms. Before harvest, fungicides and resistant varieties can be utilized to combat spoilage but a knowledge of the effects of weather on disease epidemiology may help improve the timing of remedial measures. In stored products, environmental control is important, but often emphasis has been given to the control of only one factor, ignoring the possibility of synergistic interactions. Often the most limiting requirement of a fungus for one factor occurs when all other factors are optimal. An integrated strategy for controlling several factors could therefore give much more effective control of deterioration without requiring extreme control of any one factor. With processed foods, close attention needs to be given to the risk of contamination and rate of deterioration to determine safe shelf life while avoiding undesirable preservative levels. Pitt & Hocking (1985) also draw attention to the hazards of adding nutrients that may be utilized more readily by spoilage organisms, and to the risks from preservative-tolerant species in foods.
7. References BUCHANAN, J., SOMMER, N.F. & FORTLAGE, R.J. 1985
Aflatoxin suppression by modified atmospheres containing carbon monoxide. Journal of the Amer- ican Horticultural Society 110,638-641.
CUERO, R.G., SMITH, J.E. & LACEY, J. 1986 The influ- ence of gamma irradiation and sodium hypochlorite sterilization on maize seed microflora and germi- nation. Food Microbiology 3, 107-1 13.
CUERO, R.G., SMITH, J.E. & LACEY, J. 1987a Stimu- lation by Hyphopichia burtonii and Bacillus amyloli-
quefaciens of aflatoxin production by Aspergillus fravus in irradiated maize and rice grains. Applied and Environmental Microbiology 53, 1142-1 146.
CWERO, R.G., SMITH, J.E. & LACEY, J. 1987b Inter- action of water activity, temperature and substrate on mycotoxin production by Aspergillus Jlavus, Penicillium viridicatum and Fusarium graminearum in irradiated grains. Transactions of the British Mycological Society 89,221-226.
CUERO, R., SMITH, J.E. & LACEY, J. 1988 Mycotoxin

Pre- and post-harvest ecology of fungi 25s formation by Aspergillus flavus and Fusarium gra- minearum in irradiated maize grains in the presence of other fungi. Journal of Food Protection 51, 452- 456.
DUNKEL 1988 The relationship of insects to the dete- rioration of stored grain by fungi. International Journal of Food Microbiology 7,227-244.
HILL, R.A. & LACEY, J 1983a The microflora of ripen- ing barley grain and the effects of pre-harvest fungi- cide application. Annals of Applied Biology 102, 455465.
HILL, R.A. & LACEY, J 1983b Factors determining the microflora of stored barley grain. Annals of Applied Biology 102,467483.
HILL, R.A. & LACEY, J 1984 Penicillium species associ- ated with barley grain in the U.K. Transactions of the British Mycological Society 82,297-303.
HIRST, J.M., STEDMAN, O.J., LACEY, J. & HIDE, G.A. 1965 The epidemiology of Phytophthora infestans. IV. Spraying trials 1959 to 1963 and the infection of tubers. Annals of Applied Biology 55, 373-395.
HUSSAIN, A.M., SOMMER, N.F. & FORTLAGE, R.J. 1986 Suppression of Aspergillus flavus in raisins by solar heating during sun drying. Phytopathology 76, 335- 338.
KINGSLAND, G.C. 1986 Relationship between tem- perature and survival of Aspergillus Pavus Link ex Fries on naturally contaminated maize grain. Journal of Stored Products Research 22,29-32.
LACEY, J 1967 The role of water in the spread of Phy- tophthora infestans in the potato crop. Annals of Applied Biology 59,24%255.
LACEY, J. 1980 Colonization of damp organic sub- strates and spontaneous heating. In Microbial Growth in Extreme Environments ed. Corry, J.A.S. & Board, R.G. Society of Applied Bacteriology Techni- cal Series No. 15, pp. 53-70. London: Academic Press.
LACEY, J 1986a Factors affecting fungal colonization of grain. In Spoilage and Mycotoxins of Cereals and Other Stored Products ed. Flannigan, B. Internation- al Biodeterioration 22 (Supplement), 29-34.
LACEY, J. 1986b Water availability and fungal repro- duction: patterns of spore production, liberation and dispersal. In Water, Fungi and Plants ed. Ayres P.G. & Boddy, L. B.M.S. Symposium No. 11 pp. 65-86. Cambridge: Cambridge University Press.
LACEY, J., LORD, K.A., CAYLEY, G.R., HOLDEN, G.R. & SNEATH, R.W. 1983 Problems of testing novel chemicals for the preservation of damp hay. Animal Feed Science and Technology 8,283-301.
LEE, L.S., LEE, L.V. & RUSSELL, T.E. 1986 Aflatoxin in Arizona cottonseed: field inoculation of bolls by Aspergillusflavus spores in wind driven soil. Journal of the American Oil Chemists Society 63,530-532.
LEE, L.S., LACEY, P.E. & GOYNES, W.R. 1987 Aflatoxin in Arizona cottonseed: a model study of insect- vectored entry of cotton bolls by Aspergillus flavus. Plant Disease 71,997-1001.
MAGAN, N. & LACEY, J. 1984a Effect of temperature and pH on water relations of field and storage fungi. Transactions of the British Mycological Society 82, 71-81.
MAGAN, N. & LACEY, J. 1984b Effect of water activity, temperature and substrate on interactions between
field and storage fungi. Transactions of the British Mycological Society 82,53-93.
MAGEN, N. & LACEY, J. 1984c Effects of gas composi- tion and water activity on growth of field and storage fungi and their interactions. Transactions of the British Mycological Society 82, 305-314.
MAGAN, N. & LACEY, J. 1984 Water relations of some Fusarium species from infected wheat ears and grain. Transactions of the British Mycological Society 83,281-285.
MAGAN, N. & LACEY, J. 1985 Interactions between field and storage fungi on wheat grain. Transactions of the British Mycological Society 85, 29-37.
MILLER, J.D., YOUNG, J.C. & TRENHOLM, H.L. 1983 Fusarium toxins in field corn I. Time course of fungal growth and production of deoxynivalenol and other mycotoxins. Canadian Journal of Botany 61,3080-3087.
PANASENKO, V.T. 1967 Ecology of microfungi. Botan- ical Review 33, 189-215.
PAPENDICK, R.I. & MULLA, D.J. 1985 Basic principles of cell and tissue water relationships. In Water, Fungi and Plants ed. Ayres, P.G. & Boddy, L. B.M.S. Symposium No. 11, pp. 1-25. Cambridge: Cambridge University Press.
PARK, K.Y. & BULLERMAN, L.B. 1981 Increased afla- toxin production by Aspergillus parasiticus under conditions of cycling temperatures. Journal of Food Science 46, 1147-1151.
PARK, K.Y. & BULLERMAN, L.B. 1983 Effect of cycling temperatures on aflatoxin production by Aspergillus parasiticus and Aspergillus f l a w in rice and cheddar cheese. Journal of Food Science 48, 889- 896.
PEARSON, R.C. & HALL, D.H. 1975 Factors affecting the occurrence and severity of blackmold of ripe tomato fruit caused by Alternaria alternata. Phyto- pathology 65,1352-1359.
PITT, J.I. & HOCKING, A.D. 1985 Fungi and food spoil- age. Sydney: Academic Press.
ROYAL SOCIETY 1976 Units and symbols. Philosophical Transactions of the Royal Society of London, Series B 273,433.
SEITZ, L.M., SAUER, D.B., Mom, H.E. & BURROUGHS, R. 1975 Weathered grain sorghum: natural occurrence of alternariols and storability of the grain. Phytopathology 65,1259-1263.
STRAUSBAUGH, C.A. & MALOY, O.C. 1986 Fusarium scab of irrigated wheat in central Washington. Plant Disease 70,1104-1106.
STUTZ, H.K. & KRUMPERMAN, P.H. 1976 Effect of temperature cycling on the production of aflatoxin by Aspergillus parasiticus. Applied and Environmental Microbiology 32,327-332.
SUTTON, J.C. 1982 Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Canadian Journal of Plant Pathology 4, 195-209.
VARGO, R.H. & BAUMER, J.S. 1986 Fusarium sporotri- chioides as a pathogen of spring wheat. Plant Disease 70,629-63 1.
WILSON, D.M. & JAY, E. 1976 Effect of controlled atmosphere storage on aflatoxin production in high moisture peanuts (groundnuts). Journal of Stored Products Research 12,97-100.