polysialylated ncam expression during motor axon outgrowth and myogenesis in the fetal rat
TRANSCRIPT

Polysialylated NCAM Expression DuringMotor Axon Outgrowth and Myogenesis
in the Fetal Rat
DOUGLAS W. ALLAN* AND JOHN J. GREER
Department of Physiology, Division of Neuroscience, University of Alberta, Edmonton,Alberta T6G 2S2, Canada
ABSTRACTPolysialylation of the neural cell adhesion molecule (NCAM) converts it into an
anti-adhesive molecule, attenuating intercellular adhesion and repelling apposed mem-branes. Previous studies have demonstrated that interaxonal repulsion, or defasciculation,induced by polysialylated NCAM (PSA-NCAM) expressed along outgrowing chick motor axonspromotes intramuscular branching and facilitates differential guidance of segregating axonalpopulations. In the present study, we have examined the expression of PSA-NCAM in adeveloping mammalian motor system during axonal outgrowth, separation of distinct axonalpopulations, and intramuscular branching. Furthermore, we provide the first clear demonstra-tion of the spatiotemporal modulation of PSA-NCAM expression on myotubes during eachstage of myogenesis. Immunohistochemical labelling was used to compare the spatiotemporalpattern of PSA-NCAM expression with those of total-NCAM, the cell adhesion molecule L1,and growth associated protein (GAP-43) during development of the phrenic nerve anddiaphragm of fetal rats (embryonic days, E11–E19). During segregation of phrenic andbrachial axonal populations at the brachial plexus (E12.5–E13), PSA-NCAM expression wasrestricted to phrenics, being absent from brachial motoneurons. Both populations labelledequivalently for NCAM, L1, and GAP-43. We postulate that PSA-NCAM may be a componentof the molecular machinery that specifically guides phrenic motoneuron growth at thebrachial plexus. During diaphragmatic morphogenesis, PSA-NCAM expression: (i) remainedhigh within the phrenic nerve throughout intramuscular branching; (ii) was transientlyup-regulated on myotubes during myotube separation associated with primary and secondarymyogenesis; (iii) was restricted to those regions of primary and secondary myotube mem-branes, which were juxtaposed and about to separate. These data suggest a role forPSA-NCAM in the guidance of specific subsets of mammalian motoneurons and in intramus-cular branching, and demonstrate an intimate correlation between PSA-NCAM expressionand myotube separation. J. Comp. Neurol. 391:275–292, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: phrenic; diaphragm; L1; GAP-43; axon guidance; development
Modulation of cellular adhesion and repulsion is animportant mechanism by which developing axons andmuscle cells regulate cellular interactions during motoraxon pathfinding and muscle morphogenesis. During seg-regation of motor nerve trunks, adherent axons mustdefasciculate to allow for differential growth into muscle-specific nerve fascicles. Global and preferential adhesiveand repulsive factors have been demonstrated to influenceaxonal migration at such decision points (reviewed inTessier-Lavigne and Goodman, 1996). Once migratingmotor axons reach the target musculature, a reduction ininteraxonal adhesion promotes intramuscular axonal de-fasciculation and preferential growth over developingmuscle fibres (reviewed in Rutishauser and Landmesser,
1996). Similarly, during muscle morphogenesis, the fusionof myoblasts forms tightly adherent pools of myotubes,which subsequently must separate during two discrete
Grant sponsor: Medical Research Council of Canada; Grant sponsor:Alberta Lung Association; Grant sponsor: Toronto Sick Children’s HospitalFoundation; Grant sponsor: Alberta Heritage Foundation for MedicalResearch; Grant sponsor: Izaak Walton Killam Memorial ScholarshipFund.
*Correspondence to: Douglas W. Allan, Division of Neuroscience, 513HMRC, University of Alberta Edmonton, Alberta T6G 2S2, Canada.E-mail: [email protected]
Received 8 May 1997; Revised 16 September 1997; Accepted 17 Septem-ber 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 391:275–292 (1998)
r 1998 WILEY-LISS, INC.

phases of muscle formation associated with primary andsecondary myogenesis (Kelly, 1983).
Cell adhesion molecules (CAMs), postulated to act pri-marily via homophilic binding, are integral to intercellularadhesion. These CAMs include members of the immuno-globulin family, such as the neural cell adhesion molecule(NCAM) and L1, and the cadherin family. NCAM isperhaps the most ubiquitously expressed adhesion mol-ecule in nerve and muscle within the developing verte-brate embryo. It has been implicated in multiple aspects ofneuromuscular development, including promotion of axo-nal adhesion, fasciculation and outgrowth (Rutishauser etal., 1982; Landmesser et al., 1988; reviewed in Hall et al.,1996), nerve-muscle adhesion (Rutishauser et al., 1983),and adhesion and fusion of myoblasts and myotubes(reviewed in McDonald et al., 1995).
The balance between intercellular adhesion and repul-sion can be modulated by simply altering the ratio inexpression of molecules that promote either adhesion orrepulsion. For example, modulation of the expressionlevels of invertebrate NCAM homologues, Drosophila fas-ciclin II and Aplysia apCAM, correlates with the degree ofinterneuronal and neuromuscular adhesion (Lin et al.,1994; Lin and Goodman, 1994; Zhu et al., 1995; Schuster etal., 1996a, 1996b). An alternative mechanism available tovertebrates is to modulate the adhesivity of NCAM itself,via modification of its molecular structure. Thus, althoughNCAM is expressed by the majority of growing vertebrateaxons and developing muscle, developmentally regulatedfunctional variations may be generated by the multiplicityof distinct isoforms, which result from both alternatemRNA splicing and differential post-translational process-ing (reviewed in Barthels et al., 1992; Daniloff et al., 1994;Doherty et al., 1995; Rutishauser and Landmesser, 1996).A particularly striking modification of NCAM adhesive-ness occurs via the post-translational addition of longpolymers of sialyl residues to the extracellular domain ofNCAM (Hoffman and Edelman, 1983; reviewed in Sekiand Arai, 1993a; Landmesser and Rutishauser, 1996). Indirect contrast to NCAM, polysialylated NCAM (PSA-NCAM) is postulated to sterically and electrostaticallyattenuate intercellular interactions, resulting in the ‘‘spac-ing’’ of apposed membranes (Yang et al., 1992, 1994). Afunctional role for PSA-NCAM-induced attenuation ofmembrane adhesion has been demonstrated in the develop-ing chick hindlimb where its promotion of interaxonaldefasciculation is required for axonal branching and thedifferential guidance of axonal populations from a commonnerve trunk. Enzymatic removal of PSA from NCAM shiftsthe balance of adhesion and repulsion to primarily L1-mediated adhesion, causing excessive axonal fascicula-tion, thereby inhibiting branching and differential guid-ance (Landmesser et al., 1990; Zhang et al., 1992; Tang etal., 1992, 1994).
Previous studies of PSA-NCAM in developing mamma-lian motor systems have been limited to the determinationthat PSA-NCAM is expressed within the embryonic spinalcord and by spinal motoneurons (Boisseau et al., 1991;Seki and Arai, 1993b). In the present study, we expandupon these observations by examining PSA-NCAM expres-sion in developing rat motoneurons and how it correlateswith the segregation of distinct mammalian motor axonpopulations and nerve branching. Furthermore, we pro-vide the first clear demonstration of the spatiotemporalcorrelation between myotube separation and PSA-NCAM
expression during primary and secondary myogenesis.These data are necessary to clarify the proposed role ofPSA-NCAM in the repulsion of myotube membranes neces-sary for normal myogenesis (Fredette et al., 1993).
Toward these objectives, we have examined the spatio-temporal expression of PSA-NCAM throughout motoneu-ron and myotube development (embryonic days, E11–E19)within a simple mammalian neuromuscular system, thephrenic nerve, and diaphragm of the fetal rat. The phrenicnerve-diaphragm axis of the fetal rat proved to be particu-larly amenable for addressing these issue. Due to theanatomical isolation of the phrenic nerve and diaphragmfrom other motor systems and the thin, essentially two-dimensional nature of the fetal diaphragm, a clear delinea-tion of axonal growth and myotube formation was possibleat all developmental stages studied. Furthermore, thephrenic nerve and diaphragm are primarily recruited for asingle motor task that commences at a well-defined time(Jansen and Chernick, 1991; Greer et al., 1992), therebysimplifying the correlation between the expression ofdevelopmentally regulated molecules and the onset offunctional recruitment.
MATERIALS AND METHODS
Tissue preparation
The timing of dam pregnancy was determined from theappearance of sperm plugs in the breeding cages, desig-nated embryonic day 0 (E0). Fetal rats (E11–E19) wereremoved individually after Caesarian section from timed-pregnant rats anaesthetised with halothane (1.2–1.5%delivered in 95% 02 and 5% CO2) and maintained at ,37oCby radiant heat as approved by the Animal WelfareCommittee at the University of Alberta. After removal offetuses, dams were killed by deepening anaesthesia (5%halothane) for 10 to 15 minutes, followed by cervicaldislocation. Once removed, fetal age was confirmed bycomparison of crown-rump length measurements withthose published by Angulo y Gonzalez (1932). Embryoswere decapitated, and the thoracic and abdominal cavitieswere exposed while immersed in 4% paraformaldehyde ina 0.2 M sodium phosphate buffer (PBS, pH 7.4). Tissueswere then post-fixed at 4oC for 4 to 20 hours. The followingnumbers of embryos (E) taken from a specified number ofdams (D) were used for this study: (i) whole embryosections—E11.5 (10E, 4D), E12.5 (19E, 9D), E13 (38E,15D), E14 (12E, 7D); (ii) diaphragm whole mounts—E14.5–E15 (22E, 8D), E15.5–E16 (39E, 14D), E16.5 (34E, 14D),E17–E17.5 (29E, 9D), E18–E18.5 (16E, 4D), E19 (11E, 1D);(iii) diaphragm cross-sections—E15.5–E16 (16E, 4D), E16.5(6E, 2D), E17–E17.5 (8E, 2D), E18–E18.5 (6E, 2D), E19(6E, 2D).
Whole embryo sections. After fixation, fetuses werewashed in PBS and embedded in a 20% gelatin/glycerolsolution, which was hardened by cooling. A small blockcontaining the fetus was excised and fixed overnight. Theblock was appropriately oriented and Vibratome sectionedat 40 µm (for transverse sectioning) and 70 µm (for sagittalsectioning). Due to the low density of axons within thephrenic nerve during E12.5 and E13, cutting sectionsmuch thinner typically did not provide for a clear visualisa-tion of the respective immunolabelled axonal populations.The best resolution for determining immunolabelling pat-terns within brachial and phrenic axonal populations wasobtained by comparing the respective labelling for total-
276 D.W. ALLAN AND J.J. GREER

NCAM, PSA-NCAM, growth-associated protein (GAP-43),and L1 between comparable thick sections taken from thesame region of different fetuses of the same age (often fromthe same litter). However, for more direct comparison ofPSA-NCAM and total-NCAM expression on cervical axonswithin a given animal, serial sections from E12.5 fetuseswere alternately processed for the two immunolabels. Allserial sections were washed in PBS, treated with theimmunoreagents while free floating, and mounted ontochrome alum gelatin-subbed slides, dried, coverslipped,and examined by standard light microscopy.
Diaphragm whole-mounts. Whole diaphragms wereisolated immediately from fetal rats, pinned flat, andpostfixed at 4oC for 4 to 20 hours. They were then washedin PBS, treated with the immunoreagents while freefloating, and mounted on subbed slides.
Diaphragm cross-sections. The following two proce-dures were followed: (1) Subsequent to fixation, dia-phragms were embedded in a 20% gelatin/glycerol solu-tion. Blocks were hardened and fixed as above and orientedfor either mediolateral or dorsoventral Vibratome section-ing at 30 to 40 µm. This produced sections that demon-strated the longitudinal and transverse axes of myotubes,respectively. Serial sections were washed in PBS andalternately treated with the immunoreagents for PSA-NCAM and total-NCAM, mounted, and coverslipped sothat the sections were kept moist until examination.PSA-NCAM immunolabelled sections only are presented.(2) Whole diaphragms were treated with the immunore-agents while free floating, and embedded in paraffin wax.Dorsoventral (transverse) sections were cut on a micro-tome (8 µm). These were mounted onto non-subbed slides,dried, and coverslipped. All diaphragm cross-sections fromboth methods were examined with Nomarski optics.
Antibodies
All primary antibodies used were diluted in PBS with1% goat serum and 0.1% NaN3. Detection of GAP-43 wasperformed by using the mouse anti-GAP-43 IgG monoclo-nal antibody (MAb; Sigma, St. Louis, MO) at a dilution of1:500. This antibody recognises an epitope on both phos-phorylated and dephosphorylated forms of GAP-43 (Meiriet al., 1991). Polysialylated NCAM was detected by usingthe mouse anti-PSA IgM MAb, termed 12E3 (a generousgift from T. Seki; Juntendo University, Tokyo, Japan).12E3-containing ascities fluid was diluted to 1:5,000 to1:10,000. This MAb detects PSA-NCAM when a polymer of6 or more a2,8-linked sialic acid residues are present anddoes not cross-react with other sialylated species, such asgangliosides (Seki and Arai, 1991; Sato et al., 1995).Total-NCAM was detected by using a rabbit polyclonalantisera (a generous gift from E. Bock; University ofCopenhagen) at a dilution of 1:2,000 to 1:3,000. Theantibody detects the three major isoforms of rat NCAM(120, 140, 180 kilodalton; Andersson et al., 1993). L1 wasdetected by using a rabbit polyclonal antisera to rat L1 (agenerous gift from V. Lemmon; Case Western University,Cleveland, OH) at a dilution of 1:1,000. All secondaryantibodies were diluted to 1:200 in PBS containing 1% goatserum. Secondary antibodies used for GAP-43 and PSA-NCAM labelling were peroxidase-conjugated goat anti-mouse IgG (Sigma; whole molecule) and biotinylated goatanti-mouse IgM (Sigma; µ-chain specific) respectively. Forboth total-NCAM and L1, peroxidase-conjugated goat anti-rabbit IgG (Sigma; whole molecule) was used.
Immunohistochemistry
Tissues were immersed in methanol containing 0.3%hydrogen peroxide for 20 to 45 minutes, followed byincubation in 1:20 goat serum (Sigma) in PBS for 1 hour.After washing in PBS, tissues were incubated in dilutedprimary antibody over 1 to 2 nights at 4oC. After washingin PBS, tissues were incubated in the appropriate second-ary antibody for 1–2 hours at room temperature. Tissuestreated with 12E3 anti-PSA MAb were further incubatedin a 1% avidin biotinylated-peroxidase complex (ABC;Vectastain, Vector Laboratories, Burlington, ON) for 1 to 2hours at room temperature. After thorough PBS washing,antibody labelling was revealed by a DAB (3,3-diaminoben-zidine tetrahydrochloride) product intensified with nickel(0.1 M Tris buffered solution, pH 8, containing 0.04% DABwith 0.04% H2O2 and 0.6% nickel ammonium sulphate) for5 to 15 minutes at room temperature. This protocolproduced an intense purple-black precipitate. Controlswere provided by primary antibody omission or inappropri-ate secondary antibodies. Photographs were digitallyscanned (Hewlett Packard ScanJet) into a computer, la-belled by using Corel Draw software (Microsoft) andprinted with a dye sublimation printer (Codonics NP600,Mitsubishi).
RESULTS
GAP-43 immunolabelling was used to provide a clearvisualisation of the full extent of axonal outgrowth amongstcervical motoneurons at various stages of developmentbeyond E12 (GAP-43 labelling was weak at earlier ages).To determine the spatiotemporal pattern of NCAM sialyla-tion on axonal membranes, immunolabelling for total-NCAM and PSA-NCAM were compared. Immunolabellingfor the functionally interrelated adhesion molecule L1 wasalso performed during times of brachial and phrenic axonseparation. Finally, immunolabelling of the developingdiaphragm for PSA-NCAM provided information regard-ing PSA-NCAM expression on phrenic axons during thetime of intramuscular branching and on myoblasts andmyotubes during primary and secondary myogenesis. Thepatterns of phrenic axon outgrowth, diaphragmatic myo-tube formation and immunolabelling for PSA-NCAM, total-NCAM, GAP-43, and L1 were remarkably consistentamongst embryonic rats of a given age, and thus theresults are presented as a series of representative figuresat several important developmental stages.
Formation of cervical ventral roots (E11.5)
We began our studies at age E11.5, immediately aftercholine acetyltransferase- and neurofilament-positive mo-toneurons are first observed within the cervical spinal cord(Phelps et al., 1988; Chen and Chiu, 1992). Figure 1illustrates the comparison between total-NCAM and PSA-NCAM immunolabelling at cervical segment C5 from twodifferent fetuses of the same age. At this age, cervicalventral roots were first emerging from the spinal cord andhad extended as far as the myotome. Small numbers ofaxons had penetrated the myotome, whereas others hadbegun to turn ventrally. Total-NCAM labelling (Fig. 1A )was primarily detected within the ventricular zone, com-missural somata and their axons, ventral motoneuron
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 277

populations and their axons, the floorplate, and myotome.In contrast, PSA-NCAM labelling (Fig. 1B) was restrictedto the myotome and a subpopulation of motoneuronswithin the ventrolateral spinal cord and their axons withinthe ventral root.
Formation of the brachial plexus and axonaldivergence (E12.5–E13)
E12.5: Initiation of axonal segregation and differen-
tial growth at the brachial plexus. The maturationalchanges in the cervical spinal cord and surrounding tis-sues during the next day were visualised via immunolabel-ling for GAP-43, L1, total-NCAM, and PSA-NCAM (Fig. 2).The motoneuron population within the ventral horn hadincreased in size, dorsal root ganglia (DRG) had formed,and commissural axons had begun to cross the floorplate.Phrenic and brachial motor axons now extended toward anarea lateral of the cardinal vein at the base of the forelimbbuds. There, the brachial plexi had formed, and theearliest projections of the phrenic (continuing ventrally)and brachial nerves (turning laterally) could be observed.PSA-NCAM labelling (Fig. 2D) had increased in a numberof spinal cord regions, including the floor plate and commis-sural somata and axons. PSA-NCAM labelling had alsoincreased among motoneurons, although it was restrictedto a subpopulation of motoneurons within the ventral hornand a subpopulation of axons extending within the cervicalventral roots. In particular, only regions within the ventro-medial motoneuron populations (i.e., region of the phrenic
and dorsal ramus motoneuron nuclei) and the axons whichextended within the phrenic nerve and dorsal ramusconsistently showed high levels of PSA-NCAM labelling.In contrast, PSA-NCAM labelling was barely detectable ortotally absent along brachial motor axons. Recently gener-ated dorsal root ganglion (DRG) neurons were also largelyPSA-NCAM negative, whereas they labelled strongly fortotal-NCAM, GAP-43, and L1. Interestingly, the paucity ofPSA labelling within the DRG was maintained throughoutembryonic development and, once formed, sensory projec-tions within the dermis were absent of PSA-NCAM immu-noreactivity (Allan and Greer, unpublished observations).
Further demonstration of the differential labelling be-tween total-NCAM and PSA-NCAM is provided in Figure3, which shows three consecutive serial sections takenfrom the same fetus that were alternately labelled fortotal-NCAM and PSA-NCAM. It is clear that PSA-NCAMlabelling was localised to phrenic axons at the brachialplexus and to ventromedial motoneuron populations withinthe ventral horn.
E13: Growth of the phrenic nerve from the brachial
plexus to the primordial diaphragm. Maturationalchanges achieved by E13 were also visualised via immuno-labelling for total-NCAM, GAP-43, L1, and PSA-NCAM(Fig. 4). By this age, dorsal and ventral brachial branchescould be clearly defined and a greater number of phrenicaxons had grown beyond the brachial plexus. Total-NCAM, GAP-43, and L1 similarly labelled all identifiableneuronal structures at equivalent intensities. PSA-NCAM
Fig. 1. Early formation of the cervical ventral root. Photos showtotal-neural cell adhesion molecule (NCAM; A) and polysialylatedneural cell adhesion molecule (PSA-NCAM; B) immunolabelling intransverse sections (40 µm) from the same level of the cervical spinalcord isolated from two fetal rats aged E11.5. Both total-NCAM andPSA-NCAM label motoneurons and myotome. However, in contrast to
total-NCAM, PSA-NCAM labelling does not appear in the floorplate,ventricular zone, or commissural somata and their short axons.Furthermore, PSA-NCAM labelling is restricted to a subpopulation ofsomata within the ventral motoneuron populations and axons withinthe ventral root. Com, commissural somata; FP, floor plate; My,myotome; VR, ventral root. Scale bar 5 100 µm in B, also applies to A.
278 D.W. ALLAN AND J.J. GREER

labelling, however, was again more localised. The somataand axons of commissural, dorsal ramus, and phrenicneurons labelled strongly for PSA-NCAM. However, ven-tral motoneuronal somata outside the ventromedial pool,axons projecting into the forelimb bud, and the dorsal rootganglion (DRG) were specifically absent or labelled weaklyfor PSA-NCAM. Brachial axons did not begin to expressconsistently high levels of PSA-NCAM until E14, when theemerging forelimb musculature was beginning to form(Allan and Greer, unpublished observations).
Figure 5A shows GAP-43 labelling within a sagittalsection of a whole decapitated fetal rat aged E13. Phrenicaxons clearly can be seen to extend beyond the brachialplexus toward the diaphragmatic primordium, the pleuro-peritoneal fold (Allan and Greer, 1997). A comparisonbetween GAP-43 (Fig. 5B) and PSA-NCAM (Fig. 5C)labelling of axons within the region of the plexus furtherdemonstrates that PSA-NCAM was preferentially ex-pressed by a subset of axons within the brachial plexus,presumably phrenic. This figure also demonstratesthat phrenic axons do not travel through the plexus as atightly fasciculated group, but rather seem to be inter-mingled with brachial axons. It is also clear from Figure5C that PSA-NCAM expression within ventral roots C4–C6(C7 also visible) is greater than that which can be ac-counted for by phrenic axons. However, as can be observedin Figures 3 and 4, axons within the dorsal ramus arealso PSA-NCAM-positive. In summary, although phrenicaxons are not the only cervical axons to express PSA-NCAM, they are the only ones to do so within the brachialplexus during the period of phrenic and brachial axonseparation.
Development of intramuscular branches andmyogenesis (E14.5–E19)
E14.5: Initial stages of phrenic nerve trifurcation
within the primordial diaphragm and the onset of
primary myogenesis. Whole-mount diaphragms takenfrom fetal rats aged between E14.5–E15 and labelled forGAP-43 illustrate the site at which the phrenic nervecommenced branching within the primordial diaphragm(Fig. 6A). A close-up view of the initial branching patternon the right hemidiaphragm (Fig. 6B) shows the trifurca-tion of the nerve trunk into primary branches projectingtoward crural, dorsal, and sternal regions of the develop-ing diaphragm.At the time of initial phrenic nerve trifurca-tion, PSA-NCAM labelling was very high within all threenerve branches (Fig. 6C,D). It was at this stage that astriated pattern of PSA-NCAM labelling was first observedrunning transversely across the dorsal and sternalbranches of the phrenic nerve. The striated pattern corre-sponded to PSA-NCAM expression on recently fused pri-mary myotubes. Primary myotube formation, as detectedby PSA-NCAM labelling, was limited to regions immedi-ately surrounding the dorsal and sternal branches, extend-ing no more than ,350 µm in a mediolateral direction fromthe primary branches. These data are consistent withprevious studies demonstrating that diaphragmatic myo-tubes form sequentially in correlation with the progressionof phrenic nerve intramuscular branching (Bennett andPettigrew, 1974; Allan and Greer, 1997). Myotube forma-tion, however, was not initiated in the vicinity of the cruralbranch as it traversed the dorsal muscle region before thedorsal branch did so. Rather, it was not until the cruralbranch actually reached its appropriate muscular target
Fig. 2. Restricted polysialylated neural cell adhesion molecule(PSA-NCAM) expression during initiation of axonal segregation at thebrachial plexus. Transverse sections (40 µm) from four fetal rats agedembryonic day 12.5, comparing immunolabelling for growth associ-ated protein (GAP-43; A), the cell adhesion molecule L1 (B), total-NCAM (C), and PSA-NCAM (D) at the same segmental level. Phrenicand brachial axons have begun to segregate at the brachial plexus.Phrenics continue ventrally as brachials turn laterally into the limb
bud. GAP-43 delineates axonal processes of the dorsal ramus, commis-surals, dorsal root ganglion (DRG), phrenics (Ph), and brachials (Br).L1 and total-NCAM label these axons to approximately the sameextent as GAP-43, although there appears to be less L1 withincommissural axons. PSA-NCAM labelling is specifically absent frombrachial axons and is concentrated within the ventromedial region ofthe ventral horn (arrow in D). Com, commissural somata; FP, floorplate; My, myotome; V, ventral horn. Scale bars 5 100 µm.
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 279

within the crural region of the diaphragm that myotubeformation was initiated in its vicinity (ZE15). Thus, itwould appear that the arrival of axonal endings coincideswith the initial formation of primary myotubes and theirsubsequent expression of PSA-NCAM at the appropriatetarget region. Immunolabelling for total-NCAM at this ageresulted in excessively heavy staining in the vicinity of thenerve and developing muscle (labelling axons, myotubes,and myoblasts), which prevented a clear discrimination ofaxonal and myotube structure (not shown).
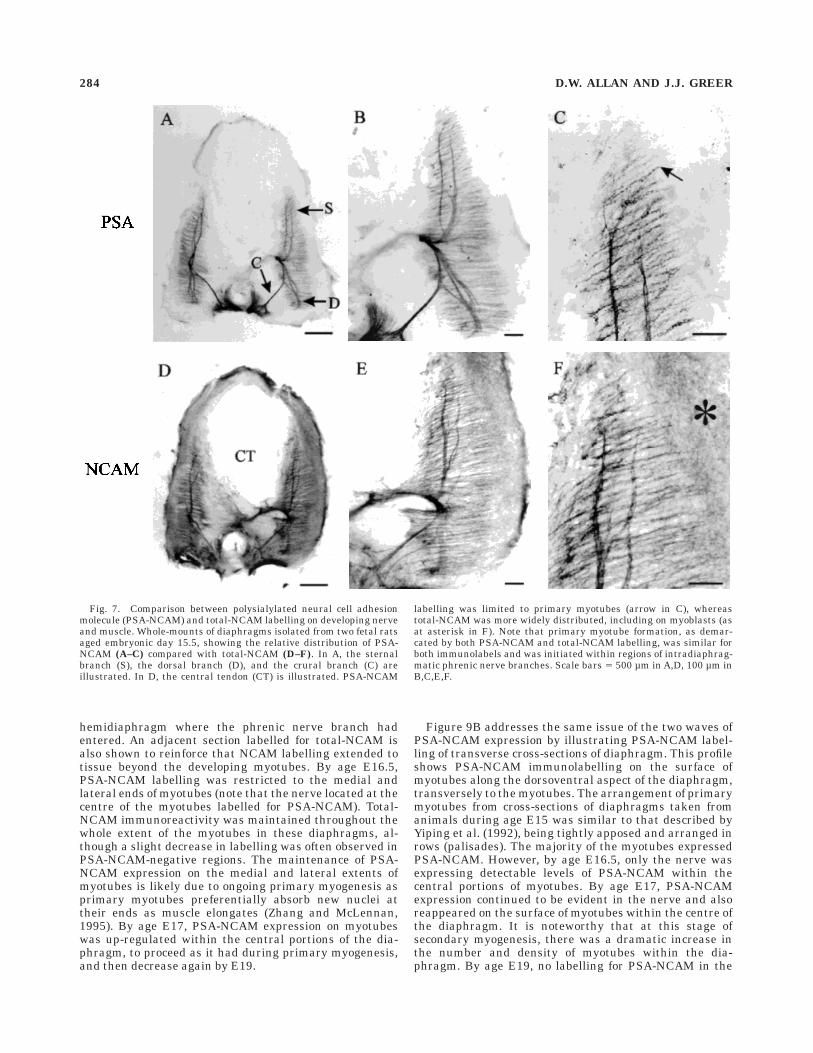
E15–E15.5: Elaboration of phrenic nerve intramuscu-
lar branching and primary myogenesis. Figure 7 illus-trates the extent of phrenic nerve innervation within thedeveloping diaphragm at age E15.5. The three majorbranches were well defined, and the crural branch hadreached its target area. PSA-NCAM labelling remainedhigh within the phrenic axons as higher order branchingprogressed (Fig. 7A–C). The formation, and subsequentPSA-NCAM labelling, of primary myotubes had pro-gressed dorsoventrally in conjunction with the intramuscu-lar growth of the primary phrenic intramuscular branchesand mediolaterally from the site of initial axon-myotubecontact as myotube elongation proceeded. A similar rela-tionship between the extent of intramuscular branching ofthe phrenic nerve and myotube formation has been ob-served via combining immunolabelling for GAP-43 anddesmin to delineate phrenic axons and myotubes, respec-tively (Allan and Greer, 1997). Figure 7C demonstratesthat primary myotubes began to form and express PSA-NCAM up to 150–200 µm distal to the tip of the growingphrenic axons. This finding suggests the possibility that
myotube formation and subsequent PSA-NCAM expres-sion in vivo may be regulated by neuronal-derived diffus-ible substances or the transmission of an electrical signalvia gap junctions within adjacent primary myotubes.
A comparison between PSA-NCAM and total-NCAM(Fig. 7D–F) labelling showed that immunolabelling withthese two antibodies overlapped. Moreover, it also becameevident from comparing these two immunolabels, thattotal-NCAM was being expressed on cells beyond thesubset that were expressing PSA-NCAM. Comparison ofFigures 7C and 7F, in particular, show that total-NCAMlabelling was present between and beyond the boundariesof the striated PSA-NCAM immunoreactivity of primarymyotubes. This pattern of total-NCAM labelling coincideswell with the fact that unfused primary myoblasts withindeveloping muscle express NCAM (Covault and Sanes,1986). Furthermore, the distribution (close proximity tomyotubes) of the total-NCAM labelling that surroundsmyotubes was similar to that of the p75 low-affinity nervegrowth factor receptor (Allan and Greer, 1997), which wehave postulated to label myoblasts in developing dia-phragms. For these reasons we believe that myoblastsrevealed by total-NCAM and p75 immunolabelling arePSA-NCAM-negative. In serially sectioned and alter-nately labelled diaphragm sections (as in Figs. 9A vs. B),there were no discernible differences between myotubelength as determined by PSA-NCAM vs. total-NCAMlabelling. This finding indicates that PSA-NCAM expres-sion commences shortly after, or coincidentally with, thetime when myoblasts fuse to form and/or elongate primarymyotubes.
Fig. 3. Comparison of total-neural cell adhesion molecule (NCAM)and polysialylated (PSA)-NCAM labelling during axonal segregationat the brachial plexus. Serial sections (40 µm, A most caudal) takenfrom a fetal rat aged embryonic day 12.5 and alternately immunola-belled for total-NCAM (A,C) and PSA-NCAM (B). In comparison to
total-NCAM, PSA-NCAM is absent from brachial axons, dorsal rootganglia, and ventral horn somata outside the ventromedial region. Br,brachial axons; DRG, dorsal root ganglion; FL, forelimb bud; My,myotome; Ph, phrenic axons. Scale bar 5 100 µm in C (applies to A–C).
280 D.W. ALLAN AND J.J. GREER

E16–E19: Maturation of axonal branching pattern
and biphasic PSA-NCAM expression along myotubes
Intramuscular branching. Intramuscular branches ofthe phrenic nerve continued to elaborate from ages E16(Fig. 8A) through to E18 (Fig. 8D), by which time the fullextent of the diaphragm had become innervated. Duringlate E17, labelling for PSA-NCAM within the nerve startedto decrease in a proximal-to-distal manner to a pointwhere it was barely detectable by age E18.5–E19, exceptwithin those diaphragm regions innervated at the lateststage (e.g., the sternal region, Fig. 8D). This is demon-strated by an absence of phrenic branch PSA-NCAMimmunolabelling in central diaphragmatic regions by E18(Fig. 8D), which proceeds distally up to E19. The reductionin PSA-NCAM labelling consistently correlated with de-creased expression of GAP-43 within phrenic axons,whereas total-NCAM labelling was maintained up to thelatest age studied (E19) (data not shown).
Myogenesis: Whole-mount diaphragms. Heavy label-ling for PSA-NCAM on developing primary myotubespersisted into age E16 (Fig. 8A). As with earlier ages, thespread of PSA-NCAM labelling in the dorsoventral direc-tion coincided with the growth of phrenic axons, and in themediolateral direction as a function of time since thecentre of a myotube had formed and been initially inner-vated. However, it became apparent from immunolabel-ling for PSA-NCAM in diaphragms between ages E16 and
E19 (Fig. 8A–D) that there was a striking bimodal expres-sion of PSA-NCAM within the diaphragmatic muscula-ture. At approximately age E16.5, there was a consistentdown-regulation of PSA-NCAM expression within themajority of the diaphragmatic musculature (Fig. 8B).Down-regulation occurred in central muscle regions whereprimary myogenesis was essentially complete and primarymyotubes had separated (Harris, 1981; Harris et al., 1989;Yiping et al., 1992). PSA-NCAM labelling had becomelimited to regions of most recent innervation (the moredistal regions of sternal and dorsal muscle zones) and morerecent myoblast addition (the more lateral extents ofmyotubes). Thus, PSA-NCAM expression along regions ofmyotubes formed during E14.5 had been maintained forapproximately 1.5–2 days, and the labelling observedduring E16.5 was limited to regions that had only becomePSA-NCAM-positive within the previous 1.5 days.
Asecond wave of PSA-NCAM expression spread through-out the diaphragm starting during early E17 from theinitial insertion point of the phrenic nerve into the dia-phragm (Fig. 8C). This corresponded with the onset of themajority of secondary myogenesis (Harris et al., 1989;Yiping et al., 1992). As during primary myogenesis, PSA-NCAM expression first appeared within the middle ofmyotubes, around the point of innervation, and progressedmediolaterally and dorsoventrally as secondary myogen-esis proceeded into E18 (Fig. 8D). PSA-NCAM expression
Fig. 4. Restricted polysialylated neural cell adhesion molecule(PSA-NCAM) expression as axonal segregation at the brachial plexusproceeds. Transverse sections (40 µm) taken from four fetal rats agedembryonic day 13 showing the relative immunolabelling for growthassociated protein (GAP-43) (A), the cell adhesion molecule, L1 (B),total-NCAM (C), and PSA-NCAM (D). Labelling for PSA-NCAM washigh on phrenic axons (Ph) but at very low levels or absent on brachial
axons (Br). Furthermore, PSA-NCAM was specifically localised to theventromedial region of the ventral horn (arrow in D). Also, there wasvery little PSA-NCAM labelling within the dorsal root ganglion (DRG)or dorsal root entry zone (DREZ). See text for further description ofPSA-NCAM expression in other regions. CV, cardinal vein; DR, dorsalramus; My, myotome. Scale bars 5 100 µm.
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 281

was again down-regulated coincident with the completionof the majority of secondary myogenesis (E19 for the majorityof the diaphragm), following a comparable time course asduring primary myogenesis. This pattern of PSA-NCAMexpression is consistent with the notion that secondary myo-tubes extend longitudinally along the primary myotubes radi-ating from the point of nerve innervation (Duxson and Sheard,1995). An exception to the bimodal pattern of PSA-NCAMexpression in the developing diaphragm was observed in thecrural region (Fig. 9). Both primary and secondary myogenesiswere likely prolonged within this region as the crural areathickened considerably in relation to other regions of thediaphragm. PSA-NCAM expression in the crural region wassubsequently maintained throughout to the last develop-mental stage that we systematically studied (E19).
Myogenesis: Diaphragm cross-sections. In addition toimmunolabelling of diaphragm whole mounts, a series of
immunoreactions were performed on longitudinal (30–40µm) and transverse (8–30 µm) cross-sections of embryonicdiaphragms to illustrate the following two points: (i) thewaves of PSA-NCAM expression observed in the whole-mounts were not merely an artifact due to variability inthe efficacy of immunolabelling associated with age-related increased thickening of the developing diaphragm;(ii) the two waves of PSA-NCAM expression correlatedprecisely with the two waves of myotube formation charac-teristic of muscle formation (primary and secondary myo-genesis).
Figure 9A shows examples of longitudinal sections fromsimilar regions of immunolabelled diaphragm sectionstaken from fetuses of various ages. This profile shows thelevel of PSA-NCAM immunolabelling along the mediolat-eral aspect of the diaphragm and myotubes. At age E15,PSA-NCAM labelling was restricted to the centre of the
Fig. 5. Migration of phrenic axons toward the primordial dia-phragm. A: Growth associated protein (GAP-43) labelling of a sagittalsection (60 µm) of whole fetal rat illustrates the extent of phrenicnerve (Ph) development at age embryonic day 13. At this age, phrenicaxons are approaching the primordial diaphragm, the pleuroperito-neal fold, at the dorsal extent of the liver (Li). H, heart; ST, septumtransversum; S, stomach. B,C: Close-up of the brachial plexus and the
phrenic nerve (Ph) shows GAP-43 (B) and polysialylated neural celladhesion molecule (PSA-NCAM) (C) labelling of axons within thebrachial plexus and within the growing phrenic nerve. PSA-NCAMwas specifically expressed by phrenic axons within and beyond thebrachial plexus. Asterisk in C, cardinal vein. Scale bars 5 500 µm in A,100 µm in B,C.
282 D.W. ALLAN AND J.J. GREER

Fig. 6. Polysialylated neural cell adhesion molecule (PSA-NCAM)labelling is high on phrenic axons and myotubes during the onset ofinitial innervation. Whole-mounts of diaphragms isolated from twofetal rats aged late embryonic day 14. A,B: Initial nerve branching asdetected by growth associated protein (GAP-43) immunolabelling. InB, the sternal branch (S), the dorsal branch (D), and the crural branch
(C) are illustrated. C,D: Corresponding labelling for PSA-NCAM inthe phrenic nerve and on the first myotubes (arrow in D) to formwithin the diaphragm. In C, the openings for the passage of the aorta(A), eosophagus (E), and inferior vena cava (VC) are illustrated. Scalebars 5 500 µm in A,C, 100 µm in B,D.
hemidiaphragm where the phrenic nerve branch hadentered. An adjacent section labelled for total-NCAM isalso shown to reinforce that NCAM labelling extended totissue beyond the developing myotubes. By age E16.5,PSA-NCAM labelling was restricted to the medial andlateral ends of myotubes (note that the nerve located at thecentre of the myotubes labelled for PSA-NCAM). Total-NCAM immunoreactivity was maintained throughout thewhole extent of the myotubes in these diaphragms, al-though a slight decrease in labelling was often observed inPSA-NCAM-negative regions. The maintenance of PSA-NCAM expression on the medial and lateral extents ofmyotubes is likely due to ongoing primary myogenesis asprimary myotubes preferentially absorb new nuclei attheir ends as muscle elongates (Zhang and McLennan,1995). By age E17, PSA-NCAM expression on myotubeswas up-regulated within the central portions of the dia-phragm, to proceed as it had during primary myogenesis,and then decrease again by E19.
Figure 9B addresses the same issue of the two waves ofPSA-NCAM expression by illustrating PSA-NCAM label-ling of transverse cross-sections of diaphragm. This profileshows PSA-NCAM immunolabelling on the surface ofmyotubes along the dorsoventral aspect of the diaphragm,transversely to the myotubes. The arrangement of primarymyotubes from cross-sections of diaphragms taken fromanimals during age E15 was similar to that described byYiping et al. (1992), being tightly apposed and arranged inrows (palisades). The majority of the myotubes expressedPSA-NCAM. However, by age E16.5, only the nerve wasexpressing detectable levels of PSA-NCAM within thecentral portions of myotubes. By age E17, PSA-NCAMexpression continued to be evident in the nerve and alsoreappeared on the surface of myotubes within the centre ofthe diaphragm. It is noteworthy that at this stage ofsecondary myogenesis, there was a dramatic increase inthe number and density of myotubes within the dia-phragm. By age E19, no labelling for PSA-NCAM in the
Fig. 7. Comparison between polysialylated neural cell adhesionmolecule (PSA-NCAM) and total-NCAM labelling on developing nerveand muscle. Whole-mounts of diaphragms isolated from two fetal ratsaged embryonic day 15.5, showing the relative distribution of PSA-NCAM (A–C) compared with total-NCAM (D–F). In A, the sternalbranch (S), the dorsal branch (D), and the crural branch (C) areillustrated. In D, the central tendon (CT) is illustrated. PSA-NCAM
labelling was limited to primary myotubes (arrow in C), whereastotal-NCAM was more widely distributed, including on myoblasts (asat asterisk in F). Note that primary myotube formation, as demar-cated by both PSA-NCAM and total-NCAM labelling, was similar forboth immunolabels and was initiated within regions of intradiaphrag-matic phrenic nerve branches. Scale bars 5 500 µm in A,D, 100 µm inB,C,E,F.
284 D.W. ALLAN AND J.J. GREER

Fig. 8. Two waves of polysialylated neural cell adhesion molecule(PSA-NCAM) labelling in developing diaphragm myotubes. Whole-mounts of diaphragms isolated from fetal rats aged embryonic day (E)16 (A), E16.5 (B), E17.5 (C), and E18 (D), showing the relativedistribution of PSA-NCAM in the nerve and muscle at various ages. InD, the central tendon (CT) is illustrated. PSA-NCAM labelling in the
nerve decreased as the mature branching pattern was attained.PSA-NCAM labelling along myotubes appeared in two waves, sepa-rated by down-regulation during E16.5. The crural musculature(asterisk in C), however, maintained high levels of PSA-NCAMexpression throughout all times examined. See text for further details.Scale bars 5 500 µm.
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 285

Fig. 9. Immunolabelling of thin sections of diaphragm, illustratingthe biphasic expression of polysialylated neural cell adhesion molecule(PSA-NCAM) on myotubes. A: Longitudinal sections (30–40 µm) ofdiaphragm muscle taken from fetal rats aged embryonic day (E) 15,E16.5, and E17, showing the waves of PSA-NCAM labelling along themediolateral axis of the myotubes. Top two photos also illustrate therestricted localisation of PSA-NCAM labelling within the early devel-oping diaphragm in relation to total-NCAM labelling. PSA expressionwas high within the central regions of primary myotubes at E15.PSA-NCAM labelling was subsequently down-regulated in the centralportion of myotubes at age E16.5, and reappeared within the central
portion during secondary myogenesis at age E17. B: Transversecross-sections (8–30 µm) showing the distribution of myotubes andPSA-NCAM labelling in diaphragms from fetal rats aged E15 to E19.Further evidence that PSA-NCAM expression appeared in two waves,during primary (E15) and secondary (E17) myogenesis. At stagesbetween primary and secondary myogenesis (E16.5) and upon comple-tion of secondary myogenesis (E19), PSA-NCAM expression wasdown-regulated. Arrows point to phrenic axons, showing that PSA-NCAM labelling was constant on the nerve up to E17. Scale bars 5100 µm.

nerve or myotubes could be detected within diaphragmaticcross-sections.
PSA-NCAM expression between developing myotubes.Figure 10B–C shows highly magnified views of PSA-NCAM on primary myotubes during age E15. PSA-NCAMlabelling was discretely limited to regions of the primarymyotube membranes, which were in very close contactwith their neighbours. This finding is in contrast tototal-NCAM expression, which is present on the full extentof the myotube membrane surface through to birth (Covaultand Sanes, 1986; Fredette et al., 1993).
Figure 10D–F shows a highly magnified view of PSA-NCAM on primary and secondary myotubes at age E17.5.The density of myotubes had increased and two distinctsizes of myotubes were obvious. PSA-NCAM labelling wasdiscretely localised on apposed membranes between adja-cent secondary myotubes and between adjacent primaryand secondary myotubes. Labelling often appeared as twoapposed lines, which were frequently intermittent, indica-tive of PSA-NCAM expression by both apposed myotubes.
Figure 11 is an illustration that summarises our interpre-tation of the timing and role of PSA-NCAM expression onprimary and secondary myotubes. Specifically, it shows theutilisation of nonpolysialylated NCAM in initial myoblastadhesion but the transient up-regulation of PSA-NCAMupon myoblast fusion to enable separation of myotubes(see Discussion section below).
DISCUSSION
PSA-NCAM expression and phrenicnerve guidance
Guidance of phrenic axon growth and branching is likelycontrolled by the combined influences of a number ofdevelopmentally regulated molecules expressed by thenerve, muscle, and surrounding tissue. In the currentstudy, we examined the expression of one such candidate,PSA-NCAM, which induces membrane separation andthereby, axonal defasciculation. We demonstrate that spe-cific expression of PSA-NCAM by phrenic axons correlateswell with a proposed role in phrenic axon guidance at thebrachial plexus and intramuscular branching. Further-more, our data suggest that the segregation of two motoraxon populations may be achieved by PSA-NCAM expres-sion of only one population.
PSA-NCAM and phrenic-brachial axon separation.
During E12–E13, phrenic and brachial axons from mul-tiple lower cervical segments merge at the brachial plexi.Subsequently, the two populations diverge as phrenicaxons continue to grow ventrally toward the diaphragmprimordium and brachial axons turn laterally to grow intothe limb bud. From the onset of ventral root formationthrough to the completion of axonal segregation at thebrachial plexi, PSA-NCAM expression was high withinphrenic and dorsal ramus populations but extremely lowor absent in brachial axonal populations. Brachial axonsdid not start to express regularly high levels of PSA-NCAM until approximately E14, approaching the onset ofinnervation of the forelimb musculature. Such spatiotem-porally restricted PSA-NCAM expression is in contrast tothe expression of total-NCAM, L1, and GAP-43, becauseimmunolabelling for these molecules was equivalent foraxons in all motor populations at each stage of outgrowth.During motor axon growth into the chick hindlimb, NCAMsialylation occurs coincidentally, and is perhaps triggered
by, axonal growth into the plexus region (Tang et al., 1992).However, in the present study, PSA-NCAM expression wasrestricted to a subset of motoneurons from the veryinitiation of motor axonal outgrowth, before interactionwith either the plexus region or the target musculature.Thus, we postulate that during the initial stages of motoraxon outgrowth in the rat, levels of NCAM sialylation maybe directly governed by groups of genes, possibly related topopulation identity (reviewed in Tanabe and Jessell, 1996).Differences in the expression of the LIM homeodomainfamily of transcription factors have been demonstratedbetween different populations of spinal motoneurons (Tsu-chida et al., 1994; Appel et al., 1995). Such putativedifferences in the combination of genes transcribed arepostulated to drive the expression of unique combinationsof molecules by each motoneuron population, which contrib-ute to differential axonal pathfinding. Further studiescharacterising the promoter region of the sialyltransfer-ase(s) responsible for polysialylation of NCAM may shedlight on the regulatory mechanism underlying differentiallevels of PSA-NCAM expression between motoneuron popu-lations (see Yoshida et al., 1996).
At later stages of axonal outgrowth, there were correla-tions between changes in the polysialylation levels ofcervical motor axons and the timing of functional recruit-ment and target interactions. This finding agrees with thedata illustrating that motoneuron polysialyltranferaseactivity and concomitant levels of NCAM polysialylationcan be modulated by target interaction, synaptogenesis,and neuromuscular electrical activity (Bruses et al., 1995).However, the mechanism by which any of these factorsmight modify polysialylation levels within axons remainsundetermined.
From a functional perspective, the distinct expression ofPSA-NCAM by phrenic axons at the brachial plexus raisesa number of questions with regard to the potential role ofPSA-NCAM in promoting the separation of brachial andphrenic axonal populations. The most insightful work todate regarding the actions of PSA-NCAM on the segrega-tion of axonal populations comes from the studies of thedeveloping chick hindlimb by Tang et al. (1992, 1994). Inthose series of experiments, a combination of PSA removalfrom NCAM with the enzyme a2,8-linked sialic acid endo-neuraminidase (endoN) and the use of function blockingantibodies demonstrated that PSA-NCAM decreased inter-axonal adhesion primarily by attenuating L1-mediatedadhesion. Specifically, PSA-NCAM acted to interfere withthe adhesive interactions of L1 and thus allowed for thenecessary axonal defasciculation required for segregationand accurate pathfinding into dorsal and ventral nervetrunks. The data provided in the present study raises thepossibility that PSA-NCAM could act similarly in the fetalrat by means of interference with L1-mediated adhesionbetween phrenic and brachial axonal populations, both ofwhich express similarly high levels of L1. It is interestingin the case of phrenic and brachial axons that only onepopulation expressed high levels of PSA-NCAM. In thechick hindlimb, although the absolute levels of PSA-NCAM differs amongst the separating population of axons,there is not the extreme situation where only one axonalpopulation expresses regularly detectable levels of PSA-NCAM. Thus, the current data provide evidence suggest-ing that functionally relevant defasciculation may occurwhen only one population of axons expresses PSA-NCAM.Furthermore, PSA-NCAM may be playing a more selective
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 287

Fig. 10. Discrete expression of PSA-NCAM on membranes ofadjacent primary and secondary myotubes. A–C: PSA-NCAM label-ling of transverse cross-sections (8 µm) of diaphragm (taken duringembryonic day [E] 15), showing the relative localisation of PSA-NCAMexpression on developing primary myotubes. A is a low magnificationview of the muscle cross-section showing the groupings of developingmyotubes into distinct clusters throughout the belly of the muscle. B,C
are higher magnifications, illustrating that PSA-NCAM labelling wasdiscretely located on portions of primary myotube membranes whichare juxtaposed and presumably ready to separate. D–F: Low (D) andhigh magnification views (E and F) of diaphragm transverse sections(30 µm) from fetal rats aged E17.5. PSA-NCAM labelling was locatedon apposing membranes of primary and secondary myotubes. Scalebars 5 50 µm in A, 5 µm in B,C, 200 µm in D, 7 µm in E,F.

role at the rat brachial plexus than at the chick cruralplexus, providing a specific defasciculatory ability tophrenic axons which may be necessary for differentialguidance at the plexus.
Alternatively to the passive role of hindering L1-mediated adhesion, PSA-NCAM may be playing a moreinstructive role in selective phrenic axon guidance. In thisregard, it is interesting that the effect of PSA-NCAMexpression on the adhesion of growing axons depends uponthe substrate tested (Acheson et al., 1991; Boisseau et al.,1991; Zhang et al., 1992). PSA-NCAM on phrenic axonstherefore may be acting to prevent interaction with aputative cue utilised by brachial axons for guidance, oralter the response of phrenic axons to cues common to bothphrenics and brachials. Furthermore, the spatial separa-tion of surfaces induced by PSA may promote certainmolecular interactions not favoured across closely apposedsurfaces, as postulated by Yang et al. (1992). Currently, weare examining the effects of removing PSA from NCAM onphrenic axons to determine which of the above-mentionedproposed functions PSA-NCAM is serving during phrenicand brachial nerve separation and outgrowth.
Intramuscular branching. At approximately ageE14.5, phrenic axons defasciculate to form three character-istic intramuscular branches. The axons that make up thethree branches are not compartmentalised into branch-specific fascicles before this time (Laskowski and Sanes,1987). Thus, at the initial point of trifurcation, individualaxons must separate from their neighbouring axons andreorganise so as to become associated with other axonsdestined for the same branch (Laskowski and Owens,1994). The high levels of PSA-NCAM immunoreactivityassociated with the primary branching site is consistentwith the idea that it is involved in the axonal defascicula-tion necessary for this re-ordering process (Landmesser etal., 1990). PSA-NCAM expression remained high in pri-mary and higher order branches of intradiaphragmaticphrenic axons through to age E17.5, at which time themature pattern of innervation was approximated, themajority of secondary myotubes had formed, and respirato-ry-related rhythmic electrical activity of the phrenic nervehad commenced. Subsequently, from age E17.5 through toE19, PSA-NCAM expression decreased within the nerve ina spatiotemporal manner that paralleled the regional
Fig. 11. Summary of the expression of polysialylated neural celladhesion molecule (PSA-NCAM) during primary and secondary myo-genesis within the developing diaphragm. Primary myogenesis com-mences with the fusion of myoblasts. NCAM is likely one of theadhesion molecules involved in this process. Primary myotubes areinitially aligned and attached with neighbouring membranes, in part,via NCAM-mediated adhesion. Upon fusion, up-regulation of PSA-NCAM on myotube membranes facilitates the separation of primary
myotubes. Subsequently, PSA-NCAM is down-regulated along regionswhere myotubes have separated. Secondary myogenesis proceeds fromthe central, innervated portion of separated primary myotubes. In-creased expression of PSA-NCAM on the membranes of primary andsecondary myotubes facilitates separation of primary and secondarymyotubes, followed rapidly by down-regulation upon separated re-gions of myotubes. See Discussion section for further details.
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 289

termination of further axonal growth and branching.Total-NCAM expression remained high on phrenic axonsup to birth, suggesting that PSA down-regulation isachieved by either specific removal of PSA from NCAM ornonspecific turn-over of NCAM coincident with specificdown-regulation of NCAM sialylation. The regulatorymechanism controlling PSA-NCAM expression in the nerveis unknown, but there have been suggestions that in-creased neural activity, the interaction between nerve andtarget, or both, are playing a key role in the down-regulation, likely through the modification of polysialyl-transferase activity (Landmesser et al., 1990; Bruses et al.,1995). Our results are in general agreement with, andprovide a functional correlate for, these proposals, becausePSA-NCAM expression within axons of the phrenic nervestarts to decrease when respiratory drive transmissioncommences (Greer et al., 1992) and when axonal branch-ing has ceased and synaptic maturation is under waywithin the region innervated by the axon.
PSA-NCAM expression ondeveloping myotubes
A number of cell adhesion molecules, including severalforms of NCAM, integrins, and cadherins are developmen-tally regulated during myogenesis and are believed tomediate phases of cell adhesion and separation (Covault etal., 1986; Rosen et al., 1992; Fredette et al., 1993; reviewedin McDonald et al., 1995). Because PSA-NCAM functionsas an anti-adhesive molecule and is expressed by develop-ing myotubes, it is a prime candidate for involvement inmyotube separation. A previous study reported that PSA-NCAM levels within developing chick musculature in-creased progressively throughout myogenesis, peakingduring secondary myogenesis (Fredette et al., 1993). How-ever, it was unclear as to whether PSA-NCAM was associ-ated with separation of primary myotubes, and limitationof its immunoreactivity to free surfaces of myotubes dur-ing secondary myogenesis would appear counter-intuitiveto a role in separation.
Past studies have shown that NCAM is expressed onmyoblasts and upon all surfaces of developing myotubesduring myogenesis (Covault and Sanes, 1986). Our datademonstrates that the NCAM expressed between veryclosely apposed regions of myotube membranes is highlypolysialylated during both primary and secondary myogen-esis. Upon myotube separation, more distantly apposedmembranes were no longer immunoreactive for PSA-NCAM. Functionally, such localised distribution is consis-tent with the proposed role for PSA-NCAM as attenuatingadhesion at points of cell separation. Furthermore, theabsence of PSA-NCAM from free surfaces would allownonpolysialylated NCAM to mediate axon-myotube adhe-sion. During corticospinal tract development in the spinalcord, a similar cellular restriction of PSA-NCAM overtotal-NCAM has been observed (Daston et al., 1996),which correlates to the localised ability of those axons tobranch. However, the mechanisms(s) responsible for pre-cisely targeting PSA-NCAM over total-NCAM to specificregions of the cell are currently unknown.
It was interesting to note that between myotubes, PSAimmunoreactivity seemed to appear on both apposed myo-tubes transiently during primary and secondary myogen-esis. Therefore, primary myotubes may undergo two dis-crete periods of NCAM sialylation. The intrinsic cellular
mechanisms regulating the transient expression of PSA-NCAM on developing myotube membranes are not known,but likely result from precise temporal regulation of eitherpolysialyltransferase activity or alternate splicing of NCAMin isoforms that differ in their ability to be polysialylated.The levels of PSA-NCAM expression by ciliary ganglionneurons is closely regulated by polysialyltransferase activ-ity (Bruses et al., 1995). Furthermore, a temporally regu-lated switch of NCAM isoform expression was observed tocoincide with polysialylation of NCAM in chick myotubes(Fredette et al., 1993) which may alter affinity for PSAaddition (see Small et al., 1988; Nelson et al., 1995).
The underlying regulator of temporally regulatedchanges in PSA-NCAM expression on myotubes is not wellunderstood. However, within the diaphragm, PSA-NCAMexpression commenced from the onset of primary myotubeformation at E14.5, very soon after initial myoblast fusion.The close proximity of axons to newly forming myotubes inthe diaphragm implicates axon-myotube communicationin triggering myotube formation and PSA-NCAM expres-sion. At this early stage, phrenic axons are electricallyexcitable and can induce contraction of diaphragmaticmyotubes (Greer et al., 1992; Laskowski and Owens,1994). The idea of phrenic nerve-induced PSA expressionwithin the diaphragm is further supported by the demon-stration that neuromuscular electrical activity and myo-tube contractility positively regulate myotube expressionof PSA-NCAM in a mechanism involving an influx ofcalcium and activation of protein kinase C (Fredette et al.,1993; Rafuse and Landmesser, 1996). Alternatively, physi-cal contact between developing myotubes was also inti-mately correlated with PSA-NCAM labelling, with theexpression on primary and secondary myotubes commenc-ing upon initial contact and declining after separation.This putative contact-related control may be mediated byany of a combination of inter-myotube gap junctionalcommunication, contact-stimulated myotube surface recep-tor activity and/or nerve-induced contractile activity.
SUMMARY
The aim of the present study was to examine PSA-NCAM expression during various developmental stages ofa model mammalian neuromuscular system, the phrenicnerve-diaphragm axis in fetal rats. PSA-NCAM was highlyexpressed by phrenic axons throughout nerve outgrowthup to its down-regulation once the mature intramuscularbranching pattern had been attained, secondary myogen-esis was complete and functional recruitment had com-menced. Its pattern of expression suggests a specific rolefor PSA-NCAM in phrenic axon guidance at the brachialplexus and during intramuscular branching. In relation tomuscle development, PSA-NCAM expression was dis-cretely limited, spatially and temporally, to myotubes thatwere undergoing separation during both primary andsecondary myogenesis.
ACKNOWLEDGMENTS
J.J.G. was awarded an Alberta Heritage Foundation forMedical Research (AHFMR) Scholarship. D.W.A. wasawarded an AHFMR Studentship and an Izaak WaltonKillam Memorial Scholarship. The authors thank J. Carterfor technical assistance and Drs. V. Rafuse, K. Bagnall,and S. Fu for valuable comments.
290 D.W. ALLAN AND J.J. GREER

LITERATURE CITED
Acheson, A., J.L. Sunshine, and U. Rutishauser (1991) NCAM polysialicacid can regulate both cell-cell and cell-substrate interactions. J. CellBiol. 114:143–153.
Allan, D.W., and J.J. Greer (1997) Embryogenesis of the phrenic nerve anddiaphragm in the fetal rat. J. Comp. Neurol. 381:in press
Andersson, A., M. Olsen, D. Zhernosekov, H. Gaardsvoll, L. Krog, D.Linnemann, and E. Bock (1993) Age-related changes in expression ofthe neural cell adhesion molecule in skeletal muscle: A comparativestudy of newborn, adult and aged rats. Biochem. J. 290:641–648.
Angulo y Gonzalez, A.W. (1932) The prenatal growth of the albino rat. Anat.Rec. 52:117–138.
Appel, B., V. Korzh, E. Glasgow, S. Thor, T. Edlund, I.B. Dawid, and J.S.Eisen (1995) Motoneuron fate specification revealed by patterned LIMhomeobox gene expression in embryonic zebrafish. Development 121:4117–4125.
Barthels, D., G. Vopper, A. Boned, H. Cremer, and H. Wille (1992) Highdegree of NCAM diversity generated by alternative RNA splicing inbrain and muscle. Eur. J. Neurosci. 4:327–337.
Bennett, M.R., and A.G. Pettigrew (1974) The formation of synapses instriated muscle during development. J. Physiol. (Lond) 241:515–545.
Boisseau, S., J. Nedelec, V. Poirier, G. Rougon, and M. Simonneau (1991)Analysis of high PSA N-CAM expression during mammalian spinal cordand peripheral nervous system development. Development 112:69–82.
Bruses, J.L., S. Oka, and U. Rutishauser (1995) NCAM-associated polysi-alic acid on ciliary ganglion neurons is regulated by polysialyltransfer-ase levels and interaction with muscle. J. Neurosci. 15:8310–8319.
Chen, E.W., and A.Y. Chiu (1992) Early stages in the development of spinalmotor neurons. J. Comp. Neurol. 320:291–303.
Covault, J., and J.R. Sanes (1986) Distribution of NCAM in synaptic andextrasynaptic portions of developing and adult skeletal muscle. J. CellBiol. 102:716–730.
Covault, J., J.P. Merlie, C. Goridis, and J.R. Sanes (1986) Molecular formsof N-CAM and its RNA in developing and denervated skeletal muscle. J.Cell Biol. 102:731–739.
Daniloff, J.K., V. Moore, and E.H. Oliver (1994) Activity of neural celladhesion molecule (NCAM) components: A review. Cytobios 79:97–106.
Daston, M.M., M. Bastmeyer, U. Rutishauser, and D.D. O’Leary (1996)Spatially restricted increase in polysialic acid enhances corticospinalaxon branching related to target recognition and innervation. J.Neurosci. 16:5488–5497.
Doherty, P., M.S. Fazeli, and F.S. Walsh (1995) The neural cell adhesionmolecule and synaptic plasticity. J. Neurobiol. 26:437–446.
Duxson, M.J., and P.W. Sheard (1995) Formation of new myotubes occursexclusively at the multiple innervation zones of an embryonic largemuscle. Dev. Dyn. 204:391–405.
Fredette, B.J., U. Rutishauser, and L. Landmesser (1993) Regulation andactivity-dependence of N-cadherin, NCAM isoforms and polysialic acidon chick myotubes during development. J. Cell Biol. 6:1867–1888.
Greer, J.J., J.C. Smith, and J.L. Feldman (1992) Respiratory and locomotorpatterns generated in the fetal rat brain-stem spinal cord in vitro. J.Neurophysiol. 67:996–999.
Hall, H., F.S. Walsh, and P. Doherty (1996) Review: A role for the FGFreceptor in the axonal growth response stimulated by cell adhesionmolecules? Cell Adhes. Commun. 3:441–450.
Harris, A.J. (1981) Embryonic growth and innervation of rat skeletalmuscles: I. Neural regulation of muscle fiber numbers. Phil. Trans. R.Soc. Lond. B 293:257–277.
Harris, A.J., R.B. Fitzsimons, and J.C. McEwan (1989) Neural control of thesequence of expression of myosin heavy chain isoforms in foetalmammalian muscles. Development 107:751–769.
Hoffman, S., and G.M. Edelman (1983) Kinetics of homophilic binding byembryonic and adult forms of the neural cell adhesion molecule. Proc.Natl. Acad. Sci. USA 80:5761–5766.
Jansen, A.H., and V. Chernick (1991) Fetal breathing and development ofcontrol of breathing. J. Appl. Physiol. 70:1431–1446.
Kelly, A.M. (1983) Emergence of specialization of skeletal muscle. In L.D.Peachey (ed): Handbook of Physiology, Section 10, Baltimore: WaverlyPress, pp. 507–537.
Landmesser, L., L. Dahm, K. Schultz, and U. Rutishauser (1988) Distinctroles for adhesion molecules during innervation of embryonic chickmuscle. Dev. Biol. 130:645–670.
Landmesser, L., L. Dahm, J. Tang, and U. Rutishauser (1990) Polysialicacid as a regulator of intramuscular nerve branching during embryonicdevelopment. Neuron 4:655–667.
Laskowski, M.B., and J.L. Owens (1994) Embryonic expression of motoneu-ron topography in the rat diaphragm muscle. Dev. Biol. 166:502–508.
Laskowski, M.B., and J.R. Sanes (1987) Topographic mapping of motorpools onto skeletal muscles. J. Neurosci. 7:252–260.
Lin, D.M., and C.S. Goodman (1994) Ectopic and increased expression offasciclin II alters motoneuron growth cone guidance. Neuron 13:507–523.
Lin, D.M., R.D. Fetter, C. Kopczynski, G. Grenninloh, and C.S. Goodman(1994) Genetic analysis of Fasciclin II in Drosophila: Defasciculation,refasciculation, and altered fasciculation. Neuron 13:1055–1069.
McDonald, K.A., A.F. Horwitz, and K.A. Knudsen (1995) Adhesion mol-ecules and skeletal myogenesis. Semin. Dev. Biol. 6:105–116.
Meiri, K.F., L.E. Bickerstaff, and J.E. Schwob (1991) Monoclonal antibodiesshow that kinase C phosphorylation of GAP-43 during axonogenesis isboth spatially and temporally restricted in vivo. J. Cell Biol. 112:991–1007.
Nelson, R.W., P.A. Bates, and U. Rutishauser (1995) Protein determinantsfor specific polysialylation of the neural cell adhesion molecule. J. Biol.Chem. 270:17171–17179.
Phelps, P.E., R.P. Barber, and J.E. Vaughn (1988) Generation patterns offour groups of cholinergic neurons in rat cervical spinal cord: Acombined tritiated thymidine autoradiographic and choline acetyltrans-ferase immunocytochemical study. J. Comp. Neurol. 229:459–472.
Rafuse, V.F., and L. Landmesser (1996) Contractile activity regulatesisoform expression and polysialylation of NCAM in cultured myotubes:Involvement of Ca21 and protein kinase C. J. Cell Biol. 132:969–983.
Rosen, G.D., J.R. Sanes, R. LaChance, J.M. Cunningham, J. Roman, andD.C. Dean (1992) Role for the integrin VLA-4 and its counter receptorVCAM-1 in myogenesis. Cell 69:1107–1119.
Rutishauser, U., and L. Landmesser (1996) Polysialic acid in the vertebratenervous system: A promoter of plasticity in cell-cell interactions. TINS19:422–427.
Rutishauser, U., S. Hoffman, and G.M. Edelman, (1982) Binding propertiesof a cell adhesion molecule from neural tissue. Proc. Natl. Acad. Sci.USA 79:685–689.
Rutishauser, U., M. Grumet, and G. Edelman (1983) Neural cell adhesionmolecule mediates initial interactions between spinal cord neurons andmuscle cells in culture. J. Cell Biol. 97:145–150.
Sato, C., K. Kitajima, S. Inoue, T. Seki, F.A. Troy, and Y. Inoue (1995)Characterization of the antigenic specificity of four different anti-(a28-linked polysialic acid) antibodies by using lipid-conjugated oligo/polysialic acids. J. Biol. Chem. 270:18923–18928.
Schuster, C.M., G.W. Davis, R.D. Fetter, and C.S. Goodman (1996a) Geneticdissection of structural and functional components of synaptic plastic-ity: I. Fasciclin II controls synaptic stabilization and growth. Neuron17:641–654.
Schuster, C.M., G.W. Davis, R.D. Fetter, and C.S. Goodman (1996b) Geneticdissection of structural and functional components of synaptic plastic-ity: II. Fasciclin II controls presynaptic structural plasticity. Neuron17:655–667.
Seki, T., and Y. Arai (1991) Expression of highly polysialylated NCAM in theneocortex and piriform cortex of the developing and adult rat. Anat.Embryol. 184:395–401.
Seki, T., and Y. Arai (1993a) Distribution and possible roles of the highlypolysialylated neural cell adhesion molecule (NCAM-H) in the develop-ing and adult central nervous system. Neurosci. Res. 17:265–290.
Seki, T., and Y. Arai (1993b) Highly polysialylated NCAM expression in thedeveloping and adult rat spinal cord. Dev. Brain Res. 73:141–145.
Small, S.J., S.L. Haines, and R.A. Akeson (1988) Polypeptide variation inan N-CAM extracellular immunoglobulin-like fold is developmentallyregulated through alternative splicing. Neuron 1:1007–1017.
Tanabe, Y., and T.M. Jessell (1996) Diversity and pattern in the developingspinal cord. Science 274:1115–1123.
Tang, J., L. Landmesser, and U. Rutishauser (1992) Polysialic acid influ-ences specific pathfinding by avian motoneurons. Neuron 8:1031–1044.
Tang, J., U. Rutishauser, and L. Landmesser (1994) Polysialic acid regu-lates growth cone behaviour during the sorting of motor axons in theplexus region. Neuron 13:405–414.
Tessier-Lavigne, M., and C.S. Goodman (1996) The molecular biology ofaxon guidance. TINS 274:1123–1133.
Tsuchida, T., M. Ensini, S.B. Morton, M. Baldassare, T. Edlund, T.M.Jessell, and S.L. Pfaff (1994) Topographic organization of embryonic
PSA-NCAM EXPRESSION IN FETAL RAT NERVE AND MUSCLE 291

motor neurons defined by expression of LIM homeobox genes. Cell79:957–970.
Yang, P., X. Yin, and U. Rutishauser (1992) Intercellular space is affected bythe polysialic acid content of NCAM. J. Cell Biol. 116:1487–1496.
Yang, P., D. Major, and U. Rutishauser (1994) Role of charge and hydrationin effects of polysialic acid on molecular interactions on and between cellmembranes. J. Biol. Chem. 269:23039–23044.
Yiping, L., D. Appelt, A.M. Kelly, and C. Franzini-Armstrong (1992)Differences in the rat histogenesis of EDL and diaphragm in rat. Dev.Dyn. 193:359–369.
Yoshida, Y., N. Kurosawa, T. Kanematsu, N. Kojima, and S. Tsuji (1996)
Genomic structure and promoter activity of the mouse polysialic acidsynthase gene (mST8Sia II). J. Biol. Chem. 271:30167–30173.
Zhang, H., R.H. Miller, and U. Rutishauser (1992) Polysialic acid isrequired for optimal growth of axons on a neuronal substrate. J.Neurosci. 12:3107–3114.
Zhang, M., and I.S. McLennan (1995) During secondary myotube forma-tion, primary myotubes preferentially absorb new nuclei at their ends.Dev. Dyn. 204:168–177.
Zhu, H., F. Wu, and S. Sketcher (1995) Changes in expression anddistribution of Aplysia cell adhesion molecules can influence synapseformation and elimination in vitro. J. Neurosci. 15:4173–4183.
292 D.W. ALLAN AND J.J. GREER

MATERIALSAND METHODSRESULTSDISCUSSIONSUMMARYACKNOWLEDGMENTSLITERATURECITED
Uncomposed text in Xyvision Standard Format Page 1