polypeptide composition of urea- and heat-resistant
TRANSCRIPT

JOURNAL OF VIROLOGY, Oct. 1974, p. 821-833Copyright 0 1974 American Society for Microbiology
Vol. 14, No. 4Printed in U.S.A.
Polypeptide Composition of Urea- and Heat-ResistantMutants of Poliovirus Types 1 and 2
ROBERT FENNELL' AND BRUCE A. PHILLIPS
Department of Microbiology, University of Pittsburgh, School of Medicine, Pittsburgh, Pennsylvania 15261
Received for publication 16 May 1974
Five urea-resistant and two heat-resistant mutants of poliovirus types 1 and2 were isolated and their structural and nonstructural polypeptides compared tothose of their wild-type, parental strains in an attempt to correlate mutantphenotypes with alterations in specific capsid polypeptides. Four of the sevenmutants were found to contain polypeptides which differed in molecular weightfrom their respective parental viruses. However, resistance of virions to heat- or
urea-inactivation could not be attributed to changes in particular capsidpolypeptides because alterations were detected in all but one of the capsidcomponents. For two of the urea-resistant mutants and one heat-resistantmutant, no differences were found in the molecular weights of the capsid andnoncapsid polypeptides. These results, and the fact that at least 12 selectivetreatments were required to obtain stable mutants, indicate that: (i) suchphenotypes probably can be expressed by mutations affecting one or more of thelarger capsid polypeptides, and (ii) such phenotypes reflect multiplemutational steps.
Urea-resistant and heat-resistant mutants ofpoliovirus can be isolated by the repeatedinactivation (with heat or urea) and propaga-tion of the surviving virus population (6, 16).The resistance to inactivation is a genotypiccharacter of virus (2, 3, 6, 10, 12, 17). Acomparison of the inactivation kinetics of theRNA obtained from the resistant mutants andthe sensitive parental viruses showed that theresistance to inactivation could be attributed toan alteration in the capsid structure and not toan increased stability of the nucleic acid (3, 6).This study was undertaken to investigate (i)whether the capsid polypeptides from urea-resistant and heat-resistant mutants could bedifferentiated from their parental viruses bypolyacrylamide gel electrophoresis; and (ii)whether alterations in specific viral polypep-tide could be correlated with resistance to ureaor heat inactivation.
MATERIALS AND METHODSCell culture. S-3 HeLa cells were used exclusively.
Suspension cultures were maintained in Eagle mini-mal essential medium (MEM) made with spinner salts(Grand Island Biological Co., Grand Island, N.Y.)supplemented with 5% calf serum (Flow Laboratories,Rockville, Md.) and 2.0 mM glutamine. AA/50 MEMis minimal essential medium (Eagle) with spinner salts
I Present address: Microbiological Associates, Inc., Walk-ersville, Md. 21793.
containing l/50th the normal amount of each aminoacid.
Viruses. The wild-type poliovirus strains used inthese experiments were type 1, LSc; type 1, Mahoney(Mah); type 2, MEF,; and type 2, Sabin (2-Sab). Thetype 1 viruses were laboratory stocks. The type 1, LScstrain and its urea-resistant mutant, LSc-UR (seebelow), were obtained from J. Hallum and J. S.Youngner. The type 2, Sabin vaccine strain wasobtained from Lederle Laboratories, Pearl River, N.Y.In all cases, uncloned virus stocks were used. Urea-resistant (UR) and heat-resistant (HR) mutants ofthese virus stocks were isolated as described below.
Propagation of viruses. HeLa cells in suspensionculture (4 x 101 per ml) were infected with virus withan input multiplicity of 10 PFU per cell. Thirtyminutes after infection, 5% calf serum was added. Sixand one-half hours after infection, the cells werecentrifuged and the cell pellet was resuspended in asmall volume of 0.02 M phosphate buffer, pH 7.0(PB). After repeated (three to four times) freezing andthawing to rupture the cells, the cell debris wasremoved by centrifugation and the supernatant fluidwas collected. The virus was pelleted (70,000 x g for 3h), sonically treated in a Raytheon sonic oscillator for2 min, resuspended in PB, and assayed for infectivity.
Isolation of urea- and heat-resistant mutants.Urea-resistant and heat-resistant mutants of the fourwild-type (nonresistant) strains of poliovirus wereobtained by the method described by Hallum andYoungner (6). The wild-type viruses were propagatedto titers of approximately 109 PFU per ml. Prior tourea-inactivation, the virus stock was diluted 10-1 inPB and sonically treated for 60 s in a Raytheon sonic
821
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

FENNELL AND PHILLIPS
oscillator. The virus then was added to an equalvolume of 6 M urea in PB and the mixture wasincubated for 2 h at 37 C with occasional mixing. Thevirus was diluted 10- 2 in 20 ml of MEM and added tomonolayer cultures. The procedure was repeated andafter each treatment and passage, the virus popula-tion was tested for urea sensitivity. Approximately 12passages were required to obtain a stable populationof virus which showed no significant drop in titerwhen tested for urea sensitivity.The same experimental design was used for the
isolation of the heat-resistant mutants. Virus stockswere sonically treated, diluted 10- 2 in PB, andexposed to 50 C for 2 h. The virus then was diluted10-fold in 20 ml of MEM and passaged in cellmonolayers as above. The isolation of heat-resistantviruses likewise required about 12 passages of treatedvirus populations.
Test of urea sensitivity. Virus in PB (37 C) wasadded to an equal volume of 6 M urea in PB, pH 7.0,at 37 C. At designated times, 0.1-ml samples werediluted 10-2 in cold MEM and assayed for infectivity.The results of virus inactivations were expressed asthe log,0 of the surviving infectivity, PIPo, where Pequals the titer of survivors at time, t, and Po is thetiter of virus at zero time. The infectivity was mea-sured by the agar cell-suspension technique usingHeLa cells (1).
Test of heat sensitivity. Virus, which was preincu-bated at 22 C, was diluted 10-2 in 9.9 ml ofPB at 50 C(± 0.25 C). At the designated times, 0.1-ml sampleswere diluted 10- 2 in cold MEM and assayed forinfectivity. The results of heat-inactivation experi-ments were expressed also as the log,0 of the survivinginfectivity.
Preparation of cytoplasmic extracts containingradioactively labeled virus-specific proteins. Cellsin suspension culture (4 x 106/ml) containing 4 jg ofactinomycin D per ml and 3 mM guanidine wereinfected with an input multiplicity of 50 PFU per cell.Two hours after infection the cells were washed free ofguanidine (reversal) and resuspended in amino acid-deficient medium (AA/50 MEM). Radioactive aminoacids were added 90 to 120 min after guanidinereversal. Seven hours after infection the cells werecentrifuged into a pellet (800 x g for 10 min),resuspended in reticulocyte standard buffer (RSB;0.01 M NaCl, 0.0015 M MgCl2, 0.01 M Tris, pH 7.4) toa concentration of 107 cells per ml, and disrupted in aglass Dounce homogenizer. Nuclei and intact cellswere removed by centrifugation (1,000 x g for 4 min).The supernatant fraction was used as a cytoplasmicextract.
Preparation of purified virus labeled with radio-active amino acids. Suspension cultures of cells(4 x 107 per ml) were infected at a multiplicity of 50PFU per cell. Calf serum was added 45 min afterinfection. Four hours after infection, the cells werecentrifuged and resuspended to 8 x 106 cells per ml inAA/50 MEM containing 4 mM glutamine, 5% calfserum, and labeled amino acid mixture (10 uCi/ml).Six and one-half hours after infection the cells werecentrifuged into a pellet and resuspended in PB to aconcentration of 4 x 107 cells per ml. The cells were
disrupted by freeze-thawing. Cellular debris was re-moved by centrifugation and the supernatant fluidwas layered on an 11 ml, 10 to 30% sucrose gradientcontaining PB and 10 mM EDTA. The gradient wascentrifuged in an SW-41 rotor, 52,000 x g for 18 h.After centrifugation the gradient was continuouslyscanned at 260 nm by pumping the gradient through acontinuous flow cell in a Gilford recording spectropho-tometer. Usually, about 20 fractions were collected. Aportion of each fraction was assayed for radioactivity.The virus and empty capsid bands were located aspeaks of radioactivity, optical density, and, in thecase of virions, infectivity.
The virus or empty capsid fractions were pooledand dialyzed overnight against 1,000 volumes of PB.The dialyzed material was brought up to 4.0 ml withPB. Two grams of CsCl was added and the mixturewas centrifuged to equilibrium in a SW 39 rotor(37,000 RPM for 18-24 hours at 5 C). Five dropfractions were collected from the bottom of the tube.A portion of each fraction was assayed for radioactiv-ity. The virus or noninfectious empty capsids werelocated as peaks of infectivity and radiactivity, re-spectively.
Polyacrylamide gel electrophoresis of viralpolypeptides. Purified virus, empty capsids, or cyto-plasmic extracts were dissociated by the addition ofone-tenth volumes of glacial acetic acid, 5 M urea,and 10% sodium dodecyl sulfate (SDS). After 30 minof incubation at 37 C, the dissociated polypeptideswere dialyzed overnight against 6,000 volumes of 0.01M phosphate buffer, pH 7.0, containing 0.1% SDS, 0.5M urea, and 0.01 M 2-mercaptoethanol (14). Immedi-ately prior to electrophoresing, the samples wereplaced in boiling water for approximately 5 min.
Samples containing the dissociated radioactiveviral polypeptides in 5% sucrose were layered on 10%acrylamide gels (20 cm by 5 nm) containing 0.5 Murea, 0.1% SDS, and 0.1 M phosphate buffer, pH 7.0.After electrophoresis, the gels were fractionated andcollected automatically into scintillation vials by aSavant Autogeldivider and fraction collector. Approx-imately 100 fractions were collected for each gel.The gel fractions were dried and 0.3 ml of hydrogen
peroxide (30%) was added to each vial. All vials wereincubated at 37 C overnight. Three drops of Soluene-100 (Packard Instrument Co, Downers Grove, Ill.) orNCS reagent (Amersham-Searle, Arlington Heights,Ill.) were added and the sealed vials incubated over-night at 37 C. One drop of saturated ascorbic acid and12.5 ml of Aquasol scintillation fluid (New EnglandNuclear Corp., Boston, Mass.) were added and thefractions were counted in a Packard model 3375 scin-tillation spectrometer. Discrimination settings wereadjusted to give 10% spillover of ["4C]CPM into thetritium channel and 0.1% spillover of [3H]CPM intothe "4C-channel. With these discrimination settingsthe counting efficiency was approximatey 45 and65% for 3H and '4C, respectively, when compared tothe radioactivity determined when the channels wereset for optimal, nondiscriminating counting. The re-covery of radioactivity from the fractionated gelsvaried between 70 and 95%.
Reagents and isotopes. Radioactive amino acid
822 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

MUTANTS OF POLIOVIRUS TYPES 1 AND 2
tmixtures ( 14C and 3H) were obtained from NewEngland Nuclear Corp., Boston, Mass. ActinomycinD was obtained from Merck Sharp & Dohme, WestPoint, Pa. Guanidine-hydrochloride was purchasedfrom Eastman Organic Chemicals, Rochester, N.Y.The urea used for the inactivation kinetics was anultra-pure grade purchased from Schwartz-Mann,Orangeburg, N.Y.
RESULTSInactivation kinetics. The exposure of wild-
type strains of poliovirus type 1 to 3 M urea at37 C resulted in a multi-hit type of inactivationkinetics (Fig. 1). The maximum drop in titerwas obtained by 15 min of inactivation, afterwhich time the surviving fraction of virus re-mained unchanged. Type 2 strains were signifi-cantly more resistant to urea-inactivation (Fig.1), an interesting finding because most type 2strains appear to be more heat resistant thantype 1 strains (16). Their inactivation kineticsmore closely resembled single-hit kinetics. De-
+ 1.0
0
-.1.0 t
-2.0 +
0
0
9-
0
0
- 3.0
-4.0 t
-5.0 +
spite the greater initial resistance of the type2 strains, as compared to type 1 viruses, thesame number of urea exposures and passageswas required before a stable population of re-sistant mutants was obtained. It was found thatif a virus population exhibited, for example, a0.5 log10 drop in titer in 15 min, such a viruspopulation, upon further passage in the ab-sence of the selection teatment, would revertrapidly to a less resistant state.
In contrast, all of the urea-resistant mutantsexhibited no significant inactivation after 15min in 3 M urea at 37 C. The inactivationkinetics of the mutants were determined after atleast two passages of the virus without ureatreatment and reflect the kinetics of the popula-tions used in subsequent experiments.Two strains of viruses were used to obtain
heat-resistant mutants. Heat inactivation ofthese viruses appeared to follow single-hit ki-netics and again the wild-type 2 strain was moreresistant than the wild-type 1 strain (Fig. 2). As
0 5 1 0 15 20 25 30MINUTES IN 3M UREA
FIG. 1. Urea-inactivation kinetics of the urea-resistant mutants and their wild-type viruses. Virus in PB, at37 C, was added to an equal volume of 6M urea in PB and incubated at 37 C. Samples were diluted in MEMfora plaque assay of the surviving infectivity. P = PFUIml at the sample time; P0 = PFU/ml at zero time.Symbols: LSc, 0; LSc-UR, 0; MEF1, x; MEF1-UL, ®; Brun, 0; Brun-UR, U; Mah, A; Mah-UR, A; 2-Sab, O;2-Sab-UR, O.
2-BAB
\> \s " s ~~~~-L -
"B
X% _ _ O
s ~~~BRUN
823VOL. 14, 1974
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

FENNELL AND PHILLIPS
MINUTES AT 500C1 2 3
+1.0
0
-1.0
04
-2.00
-3.0 1
-4.0
- Le0
4
FIG. 2. Heat-inactivation kinetics of the heat-resistant mutants and their wild-type viruses. A 1-mlamount of virus (at 22 C) was added to 9.0 ml of PBpreincubated at 50 C. At various times, samples were
diluted in MEM (at 22 C) and assayed for thesurviving virus.
noted above, about 12 passages of virus, eachfollowing heat exposure, were necessary in orderto get stable mutants.Polyacrylamide gel electrophoresis of vi-
rion polypeptides. Representative electro-pherograms are shown in Fig. 3 and 4. Highlypurified, radioactively labeled virions were pre-pared for electrophoresis as described. Thepolypeptides from the mutant viruses were
co-electrophoresed with those of their respectiveparental strains. Controls consisted of co-elec-trophoresing 3H- and "4C-labeled preparationsof the same virus, as exemplified in Fig. 3A.Differences in the polypeptide patterns were
detected in four of the seven pairs of viruses. Asplit in the VP-2a polypeptide of the Brun-URvirus was consistently found (Fig. 3D). TheVP-1 polypeptide of the LSc_UR virus was
approximately 5,000 daltons smaller than theVP-1 of the parental LSc virus; and the VP-3polypeptide of the LSc_UR virus was approxi-mately 4,000 daltons larger than that of the LScvirus (Fig. 3E). The 2-Sab_UR virus containedboth a VP-1 and a VP-2 component smallerthan their respective components present in theparental virus, 2-Sab (Fig. 3F). The heat-resist-ant mutant of the LSc strain, LSc-H , con-
tained a BP-3 component of greater molecularweight (migrated slightly slower) than the VP-3
polypeptide of the parental, LSc virus (Fig. 4A).No differences were detected in the polypeptidecompositions of the urea-resistant mutant oftype 1, Mahoney (Mah-UR), or either of themutants derived from type 2, MEF, (MEF,-HRand MEF1_UR). No differences were ever de-tected in the VP-4 polypeptides when the elec-trophoresis was carried out for shorter times soas to retain this component on the gel (Fig. 3A).Polyacrylamide gel electrophoresis of
empty capsid polypeptides. The 73S emptycapsids of the mutants and their respectiveparental viruses were also examined for differ-ences in polypeptide composition. The electro-pherograms showed the same alterations of thepolypeptides as seen in the virion polypeptides(Fig. 5). The LSc-UR empty capsid polypep-tides, VP-1 and VP-3, were, respectively,smaller and larger than the corresponding poly-peptides of the parental LSc strain (Fig. 5C).There was some difficulty in the identificationof certain polypeptides (see arrow Fig. 5C).When such aberrant polypeptides arose, it wasdifficult to adhere to the standard definition ofpoliovirus polypeptides described by Maizel etal. (8). So the question remained as to whether agiven polypeptide could be defined as one of theVP-2 or VP-3 polypeptides.The VP-1 and VP-2 polypeptides of the 2-
Sab-UR empty capsids were smaller than thoseof the parental 2-Sab virus (Fig. 5E). The2-Sab-UR empty capsids, unlike the virions,contained several components migrating theregion of its VP-1; one of these polypeptidesmigrated coincidently with the VP-1 of itsparental type (see bracketer region, Fig. 5E).The type 1, Brunhilde empty capsids containedvery little VP-2 component; therefore, any pos-sible differences in the VP-2 polypeptides couldnot be detected (Fig. 5D). The empty capsids ofall the strains examined, except type 1, Brun-hilde, contained a major VP-2 component(s).The electrophoresis of the polypeptides from thetype 2, MEF1 strain repeatedly showed a singlemajor VP-2 polypeptide (Fig. 5B). None of theempty capsids from the resistant mutantsshowed any difference from their respectiveparental types in the mobility of their NCVP-6polypeptide.
It proved difficult to obtain partially purifiedempty capsids from the heat-resistant mutants.No significant differences between parental andmutant viruses could be detected; however, theamount of radioactivity in the empty capsidpolypeptides was very small (Fig. 6).Polyacrylamide gel electrophoresis of cyto-
plasmic polypeptides. Radioactively labeled,virus-specific polypeptides from infected cells
IftO~0MIE F1 HR_
K LSO-HR
\ No"I-
N NN %MP
N\.
I ^° _ X EF-
e~~~~~loW,MnP\~~~~1..----
N.. LS
J. VIROL.824
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

800
6001
10 20 30 40 50 60 70 80FRACTION
90
O 10 20 30 40 50 60 70 80 90FRACTION
EO LSC NL
LSc-UR ,. > >
_LS
1400-
200
1000
IL 8oo
600-
400-
200
100
0.UL
aILu
0 10 20 30 40 50 60 70 80 90 1O0 0 10 20 30 40 50 60 70 80 90 100
FRACTION FRACTION
FIG. 3. Polyacrylamide gel electrophoresis of the dissociated virion polypeptides from the urea-resistantmutants and their wild-type viruses. The radioactively labeled polypeptides from dissociated virions were lay-ered on 10%o acrylamide-SDS gels (10 cm by 6 mm), electrophoresed, fractionated into scintillation vials, andcounted for radioactivity. Panel A. Co-electrophoresis of two preparations of type 1. Mahoney virionpolypeptides. Electrophoresis was for 15 h at 80 V, 8 mA. Symbols: [3H]Mah, 0; [14C]Mah, 0. Panel B.Co-electrophoresis of type 1, Mahoney, and Mah-U' virion polypeptides. Electrophoresis was for 22 h at 80 V, 8mA. Symbols: ["4C]Mah, 0; [SH]Mah-UR, 0. Panel C. Co-electrophoresis of type 2, MEF1 and MEF1-LR virionpolypeptides. Electrophoresis was for 22 h at 80 V, 8 mA. Symbols: ['4C]MEF,, 0; [sH]MEF,-UR, 0. Panel D.Co-electrophoresis of type 1, Brunhilde and Brun-U' virion polypeptides. Electrophoresis was for 16 h at 80 V, 7mA. Symbols: [14C]Brun, 0; [SH]Brun-UR, 0. Panel E. Co-electrophoresis of type 1, LSc and LSc-UR virionpolypeptides. Electrophoresis was for 21 h at 80 V, 8 mA. Symbols: [14C]LSc, 0; [3H]LSc-UR, 0. Panel F.Co-electrophoresis of type 2, Sabin and 2-Sab-UI virion polypeptides. Electrophoresis was for 15 h at 80 V, 7mA. Symbols: ["4C]2-Sab, 0; [3H]2-Sab-UR, 0.
825
a.
A
o Moh* Mah
0.
I
ItB
o Moh* Moh-UR
0.
10 20 30 40 50 60 70 80 90 1IFRACTION
C0 MEF,* MEF,- UR
ra.
b-
Ig_\\_~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~sw
400
2001
400
3001
a2u 200
100 F
300
200a1
1001
FRACTION
7CL
q
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

FENNELL AND PHILLIPS
600
400
20
200
F RACTJON
U
100
F R A CT IO N
FIG. 4. Polyacrylamide gel electrophoresis of the dissociated virion polypeptides from the heat-resistantmutants and their wild-type virions. Preparation and electrophoresis of virion polypeptides was the same as inFig. 6. Panel A. Co-electrophoresis of type 1, LSc and LSc-HR virion polypeptides. Electrophoresis was for 22 hat 80 V, 8 mA. Symbols: [3H]LSc, 0; [14C]LSc-HR, *. Panel B. Co-electrophoresis of type 2, MEF1 andMEF1-HR virion polypeptides. Electrophoresis was for 15 h at 80 V, 7 mA. Symbols: [14C]MEF,, 0;[3H]MEF,-HR, *.
826 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

VOL. 14, 1974 MUTANTS OF POLIOVIRUS TYPES 1 AND 2
JV\J T
A If&OMOh Q ~CL >L
* Mah-U"o9
200
CL
0 10 20 30 40 50 60 70 80 94FRACTION
0 10 20 30 40 50FRACTION
60 70 80 90
0~~~ ~~
-~ ~~~0
0 10 20 30 40 50 60 70 80 90FRACTION
FIG. 5. Polyacrylamide gel electrophoresis of the dissociated empty capsid polypeptides from the urea-resist-ant mutants and their wild-type viruses. The purified empty capsids were dissociated and prepared forelectrophoresis on 10%,o acrylamide-SDS gels as described in Materials and Methods. Panel A. Co-electrophore-sis of type 1, Mahoney and Mah-U" empty capsid polypeptides. Electrophoresis was for 20 h at 80 V, 7 mA.Symbols: [3H]Mah, 0; [14C]Mah-UR, 0. Panel B. Co-electrophoresis of type 2, MEF1 and MEF1-UR emptycapsid polypeptides. Electrophoresis was for 15.5 h at 80 V, 8 mA. Symbols: [3H]MEF-UIR, 0; [14C]MEF,, 0.
Panel C. Co-electrophoresis of type 1, LSc and LSc-UR empty capsid polypeptides. Electrophoresis was for 20 hat 80 V, 8 mA. Symbols: [3H]LSc, 0; ['4C]LSc-UL, 0. Panel D. Co-electrophoresis of type 1, Brunhilde andBrun-UR empty capsid polypeptides. Electrophoresis was for 20 h at 80 V, 8 mA. Symbols: [14C]Brun, 0;[3H]Brun-UL, 0. Panel E. Co-electrophoresis of type 2, Sabin and 2-Sab-UR empty capsid polypeptides.Electrophoresis was for 20 h at 80 V, 8 mA. Symbols: ['4C]2-Sab, 0; [3H]2-Sab-UR, 0.
827
I I
CO LSC* LSC -IR
A~~~~~~~~~~~~~~~~~~~1100
a.C-) 0.
C)
FRACTION
FRACTION
.9ac
0
50
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

FENNELL AND PHILLIPS
80
60
A.
UL
40
20
0
100
80
60
A.
VL 40
20
0
0 10 20 30 40 50 60 70 80F RACTION
0 10 20 30 40 50 60 70 80F RACTION
FIG. 6. Polyacrylamide gel electrophoresis of the dissociated empty capsid polypeptides from the heat-resist-ant mutants and their wild-type viruses. The purified empty capsids were dissociated and prepared forelectrophoresis on 10% acrylamide-SDS gels as described in Materials and Methods. Panel A. Co-electrophore-sis of type 2, MEF, and MEF-HR empty capsid polypeptides. Electrophoresis was for 22 h at 80 V, 8 mA. Sym-bols: [3HIMEF1, Q; [14C]MEF1HR, *. Panel B. Co-electrophoresis of type 1, LSc and LSc-ULP empty capsidpolypeptides. Electrophoresis was for 22 h at 80 V, 8 mA. Symbols: [14C]LSc, 0; [3H]LSc-HR, *.
were prepared and electrophoresed as described Mah-UR-infected cells repeatedly migratedabove. Unlike the VP-3 polypeptide seen in the slightly faster than the VP-3 polypeptide of theelectropherograms of the virion and empty cap- type 1, Mahoney-infected cells (Fig. 7A). Asid polypeptides, the VP-3 component from the similar disparity was detected in the electro-
828 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

MUTANTS OF POLIOVIRUS TYPES 1 AND 2
1750
10000
500
° IL
a
7
6
a.
2
40 50 60 70 80 90 100
FRACTION
0 10 20 30 40 50 60 70 80 90FRACTION
600
400IL
200
1200
1000
aSoo
600I
400
200
0 10 20 30 40 50 60 70 80 90 100FRACTION
FIG. 7. Polyacrylamide gel electrophoresis of radioactively labeled, viral-specific, cytoplasmic polypeptidesfrom cells infected with the urea-resistant mutants and their wild-type viruses. Cytoplasmic extracts containingradioactively labeled, viral-specific polypeptides were prepared for electrophoresis on 10% acrylamide-SDS gelsas described in Materials and Methods. Panel A. Co-electrophoresis of type 1, Mahoney and Mah-URcytoplasmic polypeptides. Electrophoresis was for 15 h at 80 V, 8 mA. Symbols: [l4C]Mah-UR, Q; [3H]Mah, *.Panel B. Co-electrophoresis of type 1, Brunhilde and Brun-U" cytoplasmic polypeptides. Electrophoresis wasfor 20 h at 80 V, 8 mA. Symbols: [14C]Brun-U5, 0; [3H]Brun, *. Panel C. Co-electrophoresis of type 2, MEF,and MEF,-UR cytoplasmic polypeptides. Electrophoresis was for 16 h at 80 V, 8 mA. Symbols: [14CJMEF1, 0;
['H]MEF,-U", 0. Panel D. Co-electrophoresis of type 1, LSc and LSc-U'1 cytoplasmic polypeptides.Electrophoresis was for 24 h at 80 V, 8 mA. Symbols: [3H]LSc, 0; [14C]LSc-UR, 0. Panel E. Co-electrophoresisof type 2, Sabin and 2-Sab-U' cytoplasmic polypeptides. Electrophoresis was for 15 h at 80 V, 8 mA. Symbols:[3H]2-Sab, 0; [l4C]2-Sab-UR, Q.
VOL. 14, 1974
IL
829
-B | St_N - o EBrun4 > - 0 Brun-U
2> > >
0
FRACTION
4000
3000
II
C o MEF,0* MEF-UR
Ii1-F Ne>>_~~~~~~~o0 10 20 30 40 50 60 70 80 90 IC
FRACTION
00
300
200 'tL
100
8
6
*04 2
3
12
10o
I r, T
D A. o LSC4C * LSC-UV
0.0.>7 r
>.l (.0. a.u>z jz >
>
'I
II~~~~~~~~~~~~~~~~~~~~~~~~~~,III
II
-I
I
Iu
E o~~& 2-SobI I IL>> >̂f (L lL02 2-Sob-Un
I >0
Ir
I
11O 10 20 30
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.
0 10 20 30 0 5, 60 70 So 90
F RACT ION
* 1i 26 36 40 s5 To t6 90
FRAC rON83()
16W
1400
1200
100W
ILu 600
400
200
0
121400
12W
a.
4W0
200
S
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

MUTANTS OF POLIOVIRUS TYPES 1 AND 2
phoresis of the LSc-UH cytoplasmic polypep-tides. The electrophoresis of the LSc-UR virionand empty capsids polypeptides had shown thatthe VP-3 polypeptide(s) was larger than theVP-3 component of the LSc virus. No differencebetween the cytoplasmic VP-3 components wasdetected (Fig. 7D). Three of the resistant vi-ruses had shown alterations in the mobility ofthe virion VP-1 polypeptide (Brun-U', LSc_UR,2-Sab-UHl). Examination of the cytoplasmicextracts revealed that cells infected with theseviruses contained multiple components whichmigrated in the VP-1 regions (Fig. 7B, D, andE). In addition, the NCVP-lb polypeptide fromthe LSc_UR-infected cells migrated slightlyfaster than the NCVP-lb polypeptide of theparental LSc virus (Fig. 7D). No difference wasdetected in the NCVP-la components and againno difference was detected in the NCVP-6polypeptides.A similar analysis of the cytoplasmic poly-
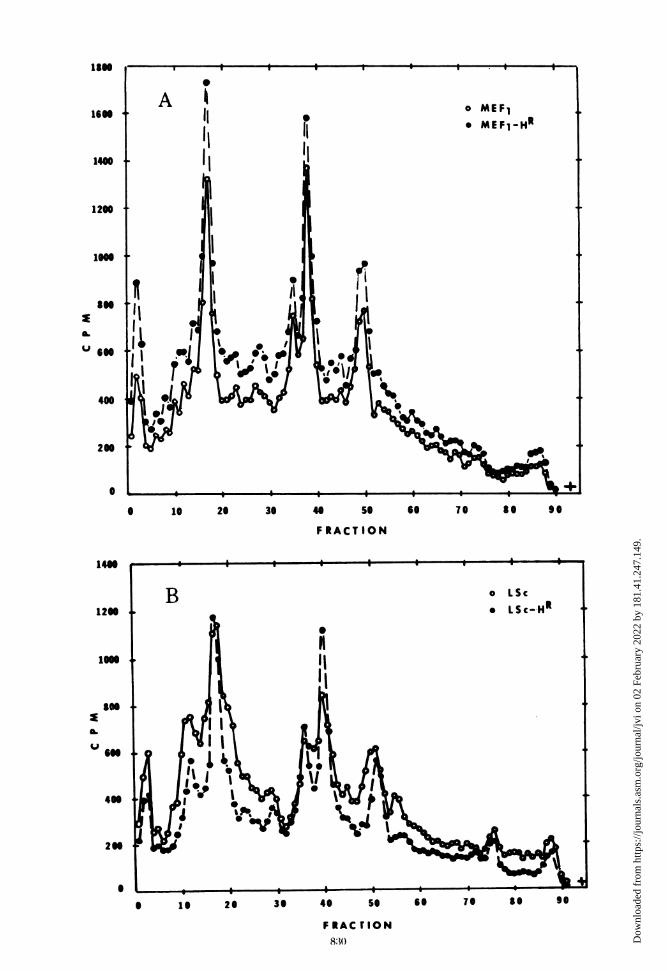
peptides synthesized in MEF1-HR- and LSc-HR-infected cells showed that there were no detect-able differences between the mutants and theirparental types (Fig. 8).
DISCUSSIONFive urea-resistant and two heat-resistant
mutants were isolated from five strains of polio-virus types 1 and 2. The polyacrylamide gelelectrophoresis of the polypeptides of theseviruses indicated that four of the seven mutantscontained a polypeptide(s) which differed inmolecular weight from the corresponding poly-peptide of their respective parental virions. Asummary of these findings is shown in Table 1.The urea-resistant or heat-resistant phenotypeof the virion could not be attributed to analteration in one particular polypeptide be-cause, depending on the virus strain, differenceswere seen in all but one of the capsid polypep-tides (namely, VP-4). No differences were de-tected in any of the noncapsid viral polypep-tides with the exception of the NCVP-lb compo-nent of the LSc-UR virus (Fig. 7D). Thesefindings are not surprising in that there is nocovariation with respect to the urea- and heat-resistant markers (6). It should be emphasizedthat only differences in the molecular weightsof the viral polypeptides were detectable in.these experiments. Amino acid substitutions
TABLE 1. Alterations in viral polypeptides
Alterations in viral polvpeptidesdetected in:
Comparison ofstrains Empty Cyto-
Virion capsid plasmiccasd extract
Mah-U5 vs Mah None None VP-3MEF,-U5 vs MEF, None None NoneLSc-U5 vs LSc VP-1 VP- 1 VP- 1
VP-3 VP-3Brun-UH vs Brun VP-2 Nonea None2-Sab-U 'vs 2-Sab VP-1 VP-I VP-1
VP-2 VP-2MEF,-H5vs MEF, None None NoneLSc-H' vs LSc VP-3 VP-3 VP-3
a There was insufficient amount of VP-2 in extractsto reliably detect differences between wild-type andmutant viruses.
in the polypeptides that did not affect cleavagereactions would not be detected.The electropherograms of the viral polypep-
tides from empty capsids and cytoplasmic ex-tracts generally supported the similarities ordifferences found by examination of virion poly-peptides. One should note that misleading re-sults might be obtained if only the cytoplasmicextracts from infected cells were examined. Forexample, the electrophoresis of the cytoplasmicextract of Mah-UR-infected cells often revealeda slight alteration in the VP-3 polypeptide;purified virions showed no such difference.Similarly, the electrophoresis of the cytoplas-mic extracts of LSc-UR-infected cells did notshow the altered VP-3 polypeptide repeatedlydetected in the purified virions. Therefore, theexamination of the structural polypeptides ap-pears best accomplished by using purified vi-rions and, in addition, other virus-related parti-cles. The use of crude cytoplasmic extractsalone may give incomplete or misleading re-sults.We detected in the empty capsids of most
viral strains a significant amount of the VP-2polypeptide(s). The amount of this componentvaried from being a major component in type 1,LSc and type 2, MEF1 empty capsids (Fig. 5Band C) to a very minor component in type 1,Brunhilde empty capsids (Fig. 5D). Early re-ports on the morphogenesis of type 1, Mahoneypoliovirus claimed that the empty capsids did
FIG. 8. Polyacrylamide gel electrophoresis of radioactively labeled, viral-specific cytoplasmic polypeptidesfrom cells infected with the heat-resistant mutants and their wild-type viruses. Procedure was the same as inFig. 7. Panel A. Co-electrophoresis of type 2, MEF, and MEF,-HR cytoplasmic polypeptides. Electrophoresiswas for 16 h at 80 V, 8 mA. Symbols: [14C]MEF,, 0; [3H]MEF,-HR, *. Panel B. Co-electrophoresis of type 1,LSc and LSc-HR cytoplasmic polypeptides. Electrophoresis was for 16 h at 80 V, 8 mA. Symbols: [14C]LSc, 0;[sHILSc-HR, *.
8.31VOL. 14, 1974
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

FENNELL AND PHILLIPS
not contain the VP-2 polypeptide (7). The VP-2and VP-4 polypeptides were presumably formedfrom the cleavage of the NCVP-6 polypeptideduring the final association of the empty capsidwith the viral RNA to form virions. An interme-diate structure, the provirion, was recentlycharacterized by Fernandez-Tomas and Bal-timore (5). Evidence presented here indicatesthat the empty capsids of most poliovirusstrains do contain a VP-2 polypeptide (also seeref. 15). It may be that the VP-2 polypeptidesfound in the empty capsids are not the same asthose found in the virions after encapsidation ofthe viral RNA. Of particular interest was theVP-2 polypeptide found in the empty capsidsand virions of the 2-Sab-UR virus (Figs. 3F and5E). This VP-2 polypeptide was a major compo-nent of the empty capsid particle and had thesame molecular weight as the VP-2 polypeptidefound in its parental virus. Assuming thatempty capsids are precursors to the virion, thesedata suggest that the empty capsids possess aVP-2 component which may become part of thevirion capsid. Vanden Berghe and Boeye (15)reported that purified poliovirions containedthree VP-2 polypeptides. These investigatorsalso reported the isolation of two types of emptycapsids containing different amounts of theVP-2 polypeptide. Our experiments also indi-cated multiple VP-2 components in certainstrains of poliovirus. At least one of the VP-2components may be derived from the emptycapsid.
It is clear that the cleavage of the NCVP-6polypeptide (41,000 daltons) cannot producetwo distinct VP-2 polypeptides, each with amolecular weight of 28,000 to 26,000 and a VP-4polypeptide with a molecular weight of 5,500.One possible explanation is that the two VP-2polypeptides are formed by the ambiguouscleavage of individual NCVP-6 molecules.Therefore, the VP-2 polypeptides detected inthe virions represent a composite of a mixture ofparticles; some virions contain the smaller VP-2polypeptide and others contain the larger one.Cooper et al. (4) first suggested that ambiguouscleavage of precursor polypeptides was respon-sible for the formation of altered copies of thecapsid polypeptides.Rekosh (13) determined that the gene order of
the capsid polypeptides was VP-4 - VP-2 -VP-3 -VP-1. If these capsid polypeptides require theentire length of the NCVP-la precursor, thenthe cleavage of NCVP-la occurs at two points:between NCVP-6 and VP-3, and between VP-3and VP-1. The VP-4 and VP-2 polypeptidessubsequently form from the cleavage of
NCVP-6. It was expected, therefore, that adecrease of several thousand daltons in themolecular weight of the VP-1 polypeptide (as inthe LSc-UR and 2-Sab_UR virions) would pro-duce a corresponding increase in the size of theVP-3 polypeptide. Likewise, a decrease in thesize of the VP-3 polypeptide (as in the LSc-HRvirus) should be paralleled by an increase in thesize of the VP-1 and/or NCVP-6 polypeptides.The LSc-U R virus showed both a decrease in thesize of its VP-1 polypeptide and an increase inthe size of its VP-3 polypeptide (Fig. 3E). The2-Sab-UR virus showed no increase in the size ofits VP-3 component despite the decreased sizeof its VP-1 component (Fig. 3F). Likewise, therewas no increase in the size of the VP-1 polypep-tide of the LSc-H R virus in spite of the de-creased size of its VP-3 component (Fig. 4A).Since no difference was ever detected in the sizeof the NCVP-la precursor molecules, it seemsthat there must be portions of the NCVP-laprecursor molecule that do not become thecapsid polypeptides after cleavage. It is possi-ble, therefore, that there are more cleavage sitesthan orginally hypothesized. Alternatively,there may exist a mechanism for trimming theinitial cleavage products to the proper size.Some of the polypeptides of the mutants mayexhibit an altered sensitivity to the trimmingmechanism, possibly as a consequenc of confor-mational changes due to an amino acid substi-tution, thereby producing polypeptides of al-tered size.
ACKNOWLEDGMENTSWe gratefully acknowledge the technical assistance of
Marie Urban and Navin Patel.This research was supported by a Public Health Research
grant Al 08368 from the National Institute of Allergy andInfectious Diseases.
LITERATURE CITED
1. Cooper, P. D. 1961. An improved agar cell-suspensionplaque assay for poliovirus; some factors affectingefficiency of plating. Virology 15:153-157.
2. Cooper, P. D. 1962. Studies on the structure and func-tions of the poliovirion: effect of concentrated ureasolutions. Virology 16:485-495.
3. Cooper, P. D. 1963. Effect of concentrated urea solutionon poliovirus strains adapted to different growth tem-peratures. Virology 21:322-331.
4. Cooper, P. D., D. F. Summers, and J. V. Maizel. 1970.Evidence for ambiguity in the posttranslational cleav-age of poliovirus proteins. Virology 41:408-418.
5. Fernandez-Tomas, C. B., and D. Baltimore. 1973. Mor-phogenesis of poliovirus. II. Demonstration of a newintermediate, the provirion. J. Virol. 12:1122-1130.
6. Hallum, J. V., and J. S. Youngner. 1966. Urea andthermal mutants of poliovirus. J. Bacteriol.91:2305-2308.
7. Jacobson, M., and D. Baltimore. 1968. Polypeptidecleavages in the formation of poliovirus. Proc. Nat.Acad. Sci. U.S.A. 61:77-84.
832 J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.

MUTANTS OF POLIOVIRUS TYPES 1 AND 2
8. Maizel, J. V., and D. F. Summers. 1968. Evidence fordifferences in size and composition of the poliovirus-specific polypeptides in infected HeLa cells. Virology36:48-54.
9. Maizel, J. V., B. A. Phillips, and D. F. Summers. 1967.Composition of artificially produced and naturallyoccurring empty capsids of poliovirus Type 1. Virology32:692-699.
10. Papaevangelou, G. J., and J. S. Youngner. 1961. Thermalstability of ribonucleic acid from poliovirus mutants.Virology 15:509-510.
11. Phillips, B. A., and R. H. Fennell. 1973. Polypeptidecomposition of poliovirions, naturally occurring emptycapsids, and 14S precursor particles. J. Virol.12:291-299.
12. Pohjanpelto, P. 1961. Two different variants of poliovirus.Virology 15:231-236.
13. Rekosh, D. 1972. Gene order of the poliovirus capsidproteins. J. Virol. 9:479-487.
14. Summers, D. F., J. V. Maizel, and J. E. Darnell. 1965.Evidence for virus-specific noncapsid proteins inpoliovirus infected HeLa cells. Proc. Nat. Acad. Sci.U.S.A. 54:505-513.
15. Vanden Berghe, D., and A. Boeye. 1972. New polypep-tides of poliovirus. Virology 48:604-606.
16. Youngner, J. S. 1957. Thermal inactivation studies withdifferent strains of poliovirus. J. Immunol. 78:282-290.
17. Youngner, J. S., and J. V. Hallum. 1964. Cystine-independent thermoresistant mutants of poliovirus.J. Bacteriol. 88:265-266.
VOL. 14, 1974 833
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 02
Feb
ruar
y 20
22 b
y 18
1.41
.247
.149
.