polyamine metabolism and function in brain
TRANSCRIPT

Neuro{hemistr) International. Vol. 3. No. 2, pp. 95 I I 0 1981. 0197-0186,'81,'020095-16$02.00,0 Printed in Great Britain, © 1981 Pergamon Press Ltd.
REVIEW
P O L Y A M I N E METABOLISM A N D F U N C T I O N IN BRAIN
N . SELLER
Centre de Recherche Merrell International, 16 rue d'Ankara, 67084 Strasbourg Cedex, France
CRITIQUE
by GRAHAM G. StrAw
Abstract -Putrescine, spermidine and spermine are simple alipathic polycations of ubiquitous occurrence. The pathways of biosynthesis and catabolism, and changes of the concentrations of these compounds in brain under various conditions are discussed.
The pharmacological properties of the polyamines have not yet revealed functions which are characteristic only for the CNS, but preliminary evidence suggests structural roles in membranes and a modulatory function in certain neuronal systems.
Putrescine (1,4-diaminobutane) and its derivatives spermidine and spermine occur in all living organ- isms, and share this ubiquitous occurrence with amino acids, proteins and nucleic acids. They possess structures (Fig. 1) in which positive charges are distri- buted along a flexible chain. These charges allow them to interact with a variety of negatively charged molecules nucleic acids, proteins, anionic sites of membranes and they can exert a clamp-like action, for instance between the two strands of the DNA double helix and within the helical parts of single stranded RNAs Cohen (1971, 1978).
In this function they cannot be entirely substituted by Mg 2+, with which they otherwise share many effects on enzymatic reactions. Based on a large number of observations one can state now that the polyamines affect practically all reactions in vitro, in which nucleic acids are involved Cohen (1971); Bach- rach (1973); J~inne, P6s6 and Raina (1978). These reactions comprise all steps in the transcription and translation of genetic information. This suggests a multifunctional role of the polyamines within the machinery that is involved in cell growth and replica- tion. With the development of specific inhibitors of polyamine biosynthesis Heby and J~inne (1981) it could indeed be shown that depletion of putrescine
Abbreviations: CNS: central nervous system; PNS: peripheral nervous system; CSF: cerebrospinal fluid; GABA: 4-aminobutyrate; GABA-T: 4-aminobutyrate: 2-oxoglutarate aminotransferase (EC 2.6.1.19): GAD: L-glutamate l-carboxy-lyase (EC 4.1.1.15); MAO: mono- amine oxidase (EC 1.4.3.4); DAO: diamine oxidase (EC 1.4.3.6t.
95
and spermidine in eukaryotic cells leads to a prolon- gation of the cell cycle and finally to an arrest of cell proliferation in the G1 phase Morris and Harada (1980); Heby and Andersson (1980); Mamont, Duchesne, Joder-Ohlenbusch and Grove (1978).
The ever increasing attention which the polyamines are now attracting derives from the fact that their metabolism was found to be enhanced in fast growing tumours Russell (1973); Raina, Eloranta, Pajula, M~ntyj~irvi and Tuomi (1980) and the expectation that inhibitors of polyamine synthesis might be useful in the therapy of proliferative diseases Seller, Danzin, Prakash and Koch-Weser (1978).
The interest in potential functions of the polyamines within the nervous system has always been relatively low, although persistent. Various bio-
H H \ 1
H- ~ A ~ N . H / \
H H PUTRESCINE
H H H H \ 1 \ 1
H. ~ N ~ N - H I H H
SPERMIDINE
H H H H
l \ / \ H H H H
SPERMINE Fig. 1. Structural formula of putrescine, spermidine and
spermine.
N e t 3 2 ,~

96 N. SEILER
chemical, methodical and pharmacological aspects have been occasionally summarized Kremzner (1973); Seiler (1973; 1980) Seiler and Deckardt (1978a, b); Snyder, Shaskan and Harik (1973); Ingoglia and Stur- man (1978); Shaw (1979a); Slotkin (1979).
It is the purpose of this paper to summarize those aspects of polyamine biochemistry and pharmacology which may be useful as starting points for future work that aims at the elucidation of functions of the polyamines, which are characteristic for the CNS.
POLYAMINE BIOSYNTHESIS AND CATABOLISM
Cellular Mg 2+ and other metal cations derive from the circulation. Their levels are dependent on their diffusion or transport through membranes. Ion-ion
interactions of metallic cations can be interfered with either by displacement by other cations or by neutral- ization of the negative charge of the binding sites by metabolic reactions. In contrast, the organic polyca- tions can be formed, conjugated and degraded within the cell, and their formation and degradation is regu- lated. This permits subtle adjustment of intracellular concentrations according to physiological needs. Binding can in addition be terminated by reactions, which neutralize the positive charges of the polyca- tions, by acetylation or other conjugation reactions.
Formation of spermidine and spermine from intra- ventricularly injected labelled putrescine has been re- peatedly shown in various species Snyder. Kreuz, Medina and Russell (1970, 1973); Fischer, Korr, Seiler and Werner (1972a); Seiler (1973); Nakajima (1973):
SPERMINE N8-(2-Carboxyethyl)-sper~nldlne ; Spermic scld
CO 2
SAM 2
51d;~eo:h~isthi°- " ~ ~ AcetylCoA
Decarboxyl. SAM Acetate ~ ~ ~/ 7 N l-Acetyl sper/alne
NS-Acetyl spermldlne 4
5 '-Methylthio- • ~ CO2 ~ adenosine I 3
SAM J ." D e c a r b o x y l . Jk 2 S ~
Putreanlne ' SPERMIOIN£ ~ Isoputreanlne
,/ co 2 ~'
ORNITHINE ~ 9 z4
Ni-Acetylputresclne
c0k ./ Glutamic semlaldehyde
L Glutamate
Succ ln lc s e m l a l d e h y d e
Fig. 2. Biosynthetic and catabolic reactions of the polyamines in vertebrate tissue. 1: Ornithine decarboxylase; 2: S-adenosylmethionine decarboxylase; 3 : spermidine synthase; 4: spermine synthase: 5: AcetylCoA: polyamine N l-acetyltransferase (cytoplasmic); 6: AcetylCoA: spermidine Na-acetyltrans- ferase (nuclear); 7: polyamine oxidase; 8: monoamine oxidase; aldehyde dehydrogenase: 9: diamine oxidase; aldehyde dehydrogenase; 10: ornithine: 2-oxoacid aminotransferase; 11: GABA-T; 12: GAD.

Polyamine metabolism and function in brain 97
Shaskan and Snyder (1973); Halliday and Shaw (1976); Seiler, AI-Therib, Fischer and Erdmann (197%); Shaw (1979b), but very little work has been done on the mechanism of polyamine biosynthesis, the regulation of the activities of the enzymes involved or the regulation of brain polyamine levels. There is little doubt, however, that the biosynthetic and catabolic reactions are the same in brain as in other organs. Figure 2 gives an overview of these reactions.
Spermidine and spermine can be regarded as meta- bolic derivatives of L-ornithine and L-methionine. In the mammalian organism putrescine is formed exclus- ively from ornithine by decarboxylation. In order to form spermidine a 3-aminopropyl moiety is trans- ferred from decarboxylated S-adenosylmethionine to putrescine by spermidine synthase. An analogous reaction takes place to form spermine from sper- midine.
The two decarboxylases, ornithine decarboxylase (EC 4.1.1.17) and S-adenosylmethionine decarboxy-
lase (EC 4.1.1.50), possess certain extraordinary features. They have apparent biological half-lives of the order of 10-40 rain; they are inducible by a great variety of factors, and they are regulated indirectly by their respective reaction products. Figure 3 illustrates some of the presumed interrelationships, and shows that putrescine, the long neglected simple aliphatic diamine, is at the center of these complex regulatory mechanisms. A similar situation is expected to hold for decarboxylated S-adenosylmethionine Mamont, Joder-Ohlenbusch, Nussli and Grove (1981).
Ornithine decarboxylase is a pyridoxalphosphate dependent enzyme, while S-adenosylmethionine decarboxylase contains pyruvate as prosthetic group. The two aminopropyltransferases, spermidine syn- thase and spermine synthase are normally present in excess and are not dependent on any co-factors. Brain is an especially rich source of these enzymes Raina, Pajula and Eloranta (1976) (Table 1). They are influenced in their activity by spermine and putres- cine. respectively, but seem not to be regulated via de
_ ~ 1 1 1 1 1 ~
- I " nov° I l ' . , ' . m . / -- I synth. I [ formation
S A M _ ~ Orn J I" - - - + -- t-
I -- - - 4" -- I"
X i" I ~ c o = - I+ - I I - , - _ + - - i : _ _ ) . . - I,*. . . . . . . . . . . -- _ uecarooxylated;-~,- - i i ~ I ~ 1 " . PUTRESCINE I - - ' ' - - i - s A M -I--I--I--l-.l--I-,l--I--I- I _ I "1--t--I-.t.1-1- - - I / i l I I I I i l l i t I I i l I I l i I l l I I I iJ I - I [+
~. T ISPERMIDINE L ,~- ] I+ - I $ . , T . . $ E I '.- $ . . I +
~' I I I I I I I I I I I I I SPERI~ IDINE !--- - ~ - - C O 2
[ ',- O.oorb .,o,. (-- I I I I '~- S A M I--
%1 I I J SPERMINE I ~ - - - J I J_-
- r - - ' i 1 . . . . . I: - , ~ -_ ~ U . . . . . S P E R M I N E ' ,1--
I I I I I I I I I I I I I I I I I I I I I I I I ~
Fig. 3. Regulatory interrelations between the polyamines and their synthetic enzymes. + + + + indi- cates activation or enhancement; IIll indicates inhibition or suppression. (The scheme is partially hypothetical. It serves to illustrate the very complex regulatory systems controlling the two decarboxy- lases which furnish the immediate precursors for spermidine and spermine formation). SAM: S-adeno- syl-L-methionine, SAM-DC: S-adenosyl-L-methionine carboxy-lyase (EC 4.1.1.50). Orn: ornithine.
Orn-DC: L-ornithine l-carboxy-lyase (EC 4.1.1.17),
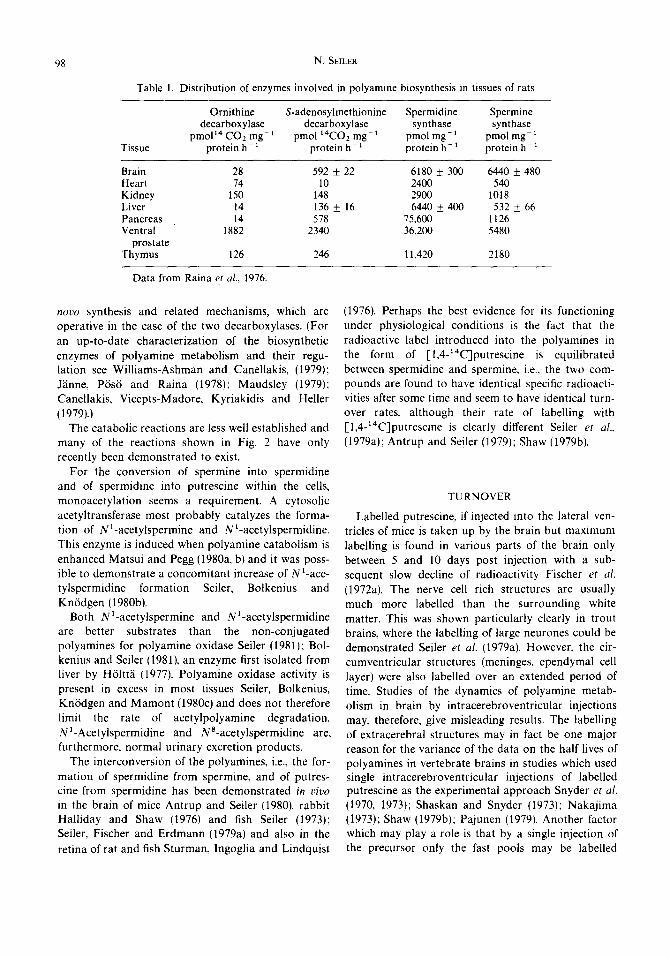
98 N. SELLER
Table 1. Distribution of enzymes involved in polyamine biosynthesis in tissues of rats
Tissue
Ornithine S-adenosylmethionine Spermidine Spermine decarboxylase decarboxylase synthase synthase
pmoP4CO2mg -1 pmo114CO2mg-1 pmolmg -l pmolmg -1 protein h -x protein h -1 protein h -~ protein h-1
Brain Heart Kidney Liver Pancreas Ventral
prostate Thymus
28 592 ± 22 6180 ± 300 6440 ± 480 74 10 2400 540
150 148 2900 1018 14 136 ± 16 6440 ± 400 532 ± 66 14 578 75,600 1126
1882 2340 36,200 5480
126 246 11,420 2180
Data from Raina et al., 1976.
novo synthesis and related mechanisms, which are operative in the case of the two decarboxylases. (For an up-to-date characterization of the biosynthetic enzymes of polyamine metabolism and their regu- lation see Williams-Ashman and Canellakis, (1979); Jiinne, P6s6 and Raina (1978); Maudsley (1979); Canellakis, Vicepts-Madore, Kyriakidis and Heller (1979).)
The catabolic reactions are less well established and many of the reactions shown in Fig. 2 have only recently been demonstrated to exist.
For the conversion of spermine into spermidine and of spermidine into putrescine within the cells, monoacetylation seems a requirement. A cytosolic acetyltransferase most probably catalyzes the forma- tion of N~-acetylspermine and N~-acetylspermidine. This enzyme is induced when polyamine catabolism is enhanced Matsui and Pegg (1980a, b) and it was poss- ible to demonstrate a concomitant increase of N~-ace - tylspermidine formation Seiler, Bolkenius and Kn6dgen (1980b).
Both Na-acetylspermine and Na-acetylspermidine are better substrates than the non-conjugated polyamines for polyamine oxidase Seiler (1981); Bol- kenius and Seiler (1981), an enzyme first isolated from liver by H61tta (1977). Polyamine oxidase activity is present in excess in most tissues Seiler, Bolkenius, Kn6dgen and Mamont (1980c) and does not therefore limit the rate of acetylpolyamine degradation. Nl-Acetylspermidine and Na-acetylspermidine are, furthermore, normal urinary excretion products.
The interconversion of the polyamines, i.e., the for- mation of spermidine from spermine, and of putres- cine from spermidine has been demonstrated in v ivo
in the brain of mice Antrup and Seiler (1980), rabbit Halliday and Shaw (1976) and fish Seiler (1973); Seiler, Fischer and Erdmann (1979a) and also in the retina of rat and fish Sturman, Ingoglia and Lindquist
(1976). Perhaps the best evidence for its functioning under physiological conditions is the fact that the radioactive label introduced into the polyamines in the form of [1,4-~4C]putrescine is equilibrated between spermidine and spermine, i.e., the two com- pounds are found to have identical specific radioacti- vities after some time and seem to have identical turn- over rates, although their rate of labelling with [1,4-x4C]putrescine is clearly different Seiler et al.,
(1979a); Antrup and Seiler (1979); Shaw (1979b).
TURNOVER
Labelled putrescine, if injected into the lateral ven- tricles of mice is taken up by the brain but maximum labelling is found in various parts of the brain only between 5 and 10 days post injection with a sub- sequent slow decline of radioactivity Fischer et al.
(1972a). The nerve cell rich structures are usually much more labelled than the surrounding white matter. This was shown particularly clearly in trout brains, where the labelling of large neurones could be demonstrated Seiler et al. (197%). However, the cir- cumventricular structures (meninges, ependymal cell layer) were also labelled over an extended period of time. Studies of the dynamics of polyamine metab- olism in brain by intracerebroventricular injections may, therefore, give misleading results. The labelling of extracerebral structures may in fact be one major reason for the variance of the data on the half lives of polyamines in vertebrate brains in studies which used single intracerebroventricular injections of labelled putrescine as the experimental approach Snyder et al.
(1970, 1973); Shaskan and Snyder (1973): Nakajima (1973); Shaw (1979b); Pajunen (1979). Another factor which may play a role is that by a single injection of the precursor only the fast pools may be labelled

Polyamine metabolism and function in brain 99
whereas the slow pools may be labelled incompletely; i.e., the semilog plotting of the specific radioactivities of the polyamines with time will give different slopes, depending on the extent of labelling of the various polyamine pools.
In a recent experiment, repeated intraperitoneal injections of trace amounts of [1,4-14C]putrescine, [2-14C]methionine or of [2-3H]methionine were used to label fast and slow polyamine pools Antrup and Seiler 11980). Although the existence of polyamine pools with different turnover rates is not known at present, their occurrence in brain with its cellular heterogeneity has to be postulated.
The most pertinent results of this work were as follows: Spermidine and spermine had apparently the same biological half-lives of about 42 days, if [1,4-~4C]putrescine was used as the precursor. In other tissues, the half lives of the polyamines seemed considerably shorter; in liver for instance 12-13 days for spermidine and 16-18 days for spermine. The ob- served half-life of spermidine and spermine in brain was. however, significantly lower, i.e,, in the same range as that observed in liver and other visceral organs, if labelled methionine was the precursor.
These results can be explained if we assume that putrescine and spermidine, which are formed by degradation of spermidine and spermine respectively, are reutilized for polyamine biosynthesis, whereas the C3 moiety originating from methionine, is irreversibly eliminated from the polyamine cycle during the step of oxidative degradation by polyamine oxidase, as is formulated in Fig. 2. The more complete putrescine reutilization is, the greater is the difference between the observable biological half lives of the polyamines with putrescine and methionine as precursors. Brain certainly has an exceptional position in this regard, as compared with other organs Antrup and Seiler (1980) but putrescine reutilization could be a general phenomenon, active to different degrees in different cells and during different physiological states of the same cell.
In a system with a low equilibrium concentration of putrescine, a low activity of ornithine decarboxylase but high activities of S-adenosylmethionine decarboxy- lase and spermidine and spermine synthases, as is the case in the mature rat brain (Table 1) a certain rate of polyamine turnover can be maintained even at a very low ornithine decarboxylase activity. This enzyme has the function of replacing the putrescine moiety which is irreversibly lost by elimination of the acetylderiva- rives or by their oxidative degradation, and by the conversion of spermidine and spermine into other derivatives, such as putreanine, spermic acid, etc.
(Fig. 2). S-adenosylmethionine will only be partially activated due to the low equilibrium levels of putres- cine. However, the local induction of ornithine decar- boxylase with a concomitant increase of the putres- cine concentration will significantly increase the syn- thesis rate of spermidine and spermine due to the activation of S-adenosylmethionine decarboxylase by putrescine (Fig. 3). In contrast, a system, such as a neuroblast or a tumour cell with a high ornithine decarboxylase activity and a concomitantly high putrescine concentration presumably has a fully acti- vated S-adenosylmethionine decarboxylase activity. A further increase of the polyamine synthesis rate can in such systems be achieved by a further stoichiometric increase of the putrescine concentration, and this will mainly affect spermidine formation.
For any given compound which regulates cellular function, concentration changes near the site of its action are essential. These can be achieved by various mechanisms, among which the change of relative rates of biosynthesis and degradation is one possibility. The higher the basal turnover rate is, the faster can equi- librium concentrations of important compounds be adjusted to new physiological needs. One can assume that a high degree of putrescine reutilization is the mechanistic basis for the adaptation of local needs of spermidine and spermine in the brain.
TRANSFORMATION OF PUTRESCINE INTO GABA
Disregarding the decarboxylation of t.-glutamate by bacteria, GABA formation was believed for over two decades to be restricted to the CNS. However, after systemic injection of trace amounts of [l,4-~4C]pu- trescine into mice or rats, putrescine-carbon was found incorporated into GABA within the tissues, in- cluding the brain Seiler, Wiechmann, Fischer and Werner (1971). These findings have been confirmed and extended Seiler and Kn6dgen (1971); Seiler, AI- Therib and Kataoka (1973); Seiler et al., (1979a); Seiler and A1-Therib (1974a); Seiler and Eichentopf (1975); de Mello, Bachrach and Nirenberg (1976); Konishi, Nakajima and Sano (1977); Tsuji and Naka- jima (1978); Sobue and Nakajima (1978) and GABA formation from putrescine was also reported to occur in cultured neuroblastoma and glioma cells Kremzner, Hiller and Simon (1975); Sobue and Naka- jima (1977).
GABA is formed continuously from putrescine as a result of polyamine degradation. This is demonstrated by the fact that radioactivity is found in GABA after a single intracerebral injection of labelled putrescine in

100 N. SEILER
trout brain, even after more than 2 months. The specific activities of these two compounds decline in parallel their decline being related to the rate of polyamine metabolism Seiler et al (1979a). The con- tinuous formation of putrescine from spermidine was shown in the same experiment.
In an organ with low diamine oxidase activity such as the adult mammalian brain Burckard, Gey and Pletscher (1963) acetylation to monoacetylputrescine seems a prerequisite to the transformation of putres- cine into GABA. This is shown by the fact, that ad- dition of acetylCoA to brain homogenates or isolated neurones and glial cells enhances dramatically the for- mation of GABA from putrescine Seiler, Schmidt- Glenewinkel and Sarhan (1979c).
Monoacetylputrescine is a substrate of MAO and is transformed into GABA in a 3 step reaction Seiler and AI-Therib (1974a) (Fig. 2). Thus there is a close analogy between the degradation of the polyamines spermine and spermidine, and of putrescine.
The acetylases responsible for the formation of monoacetylputrescine have not been definitively identified. The nuclear enzyme, which was found first, Seller and A1-Therib (1974b) is capable of forming NS-acetylspermidine from spermidine Blankenship and Walle (1978) and it acetylates histones Libby (1978). Microsomes also contain a putrescine acetylat- ing activity, and it is not impossible that a cytosolic putrescine acetylating enzyme exists as well. Its assay is presently hampered by the presence of an acetylpo- lyamine deacetylase, which splits NS-acetylspermidine and monoacetylputrescine hydrolytically Blankenship (1978).
In tissues rich in DAO, putrescine can of course be directly oxidized to 4-aminobutyraldehyde, which may undergo further transformations ; a considerable part of intraperitoneally injected labelled putrescine is degraded to COz via GABA Seiler and Eichentopf (1975); Sourkes and Missala (1978).
Compared to the formation of GABA by decar- boxylation of glutamate the transformation of putres- cine into GABA is of minor significance from a quantitative point of view, as is demonstrated by the dilution of the radioactivity in GABA, when com- pared to that of putrescine Seiler et al. (1979a); Antrup and Seiler (1980). But it is known that GAD activity is low at early stages of brain development and increases in parallel with synaptogenesis van den Berg, van Kempen, Schade and Veldstra (1965). This is also true for growing nerve cells in culture Seiler, Sarhan and Roth-Schechter (1980d). Nonetheless GABA is found in the embryonal brain before signifi- cant GAD activities are detectable van den Berg et al.,
(1965); Roberts and Kuriyama (1968); Seiler, Bink and Grove (1980a). The finding that GABA formation from putrescine is independent of glutamate decar- boxylase activity prompted studies of GABA forma- tion from labelled glutamate and putrescine in the developing chick embryo retina de Mello et al. (1976) and brain Sobue and Nakajima (1978). As expected, a significant transformation of glutamate into GABA was only observed after day 8 of embryonal develop- ment, whereas the maximal rate of transformation of putrescine into GABA was between 6 and 8 days, with slow rates after day 12. Since the time of maxi- mal putrescine degradation corresponds perhaps best with the time of early differentiation of the neuro- blasts to neuronal cells, one may speculate that for- mation of GABA is essential for cellular development. The high rate of transformation of putrescine into GABA during the stationary phase of growth in neuroblastoma cells was interpreted in the same way Sobue and Nakajima (1977). However, increased cata- bolic activities during stationary growth as compared with the logarithmic phase of growth are typical McCormick (1978) and may indicate the necessity of the cells to dispose of excessive polyamines which have accumulated during rapid growth. GABA for- mation from putrescine might nevertheless have a specific functional significance at a certain stage of brain development and deserves further attention.
POLYAMINES IN MATURE VERTEBRATE BRAINS
Brain levels of the polyamines are of the same order of magnitude as in other tissues. One cannot, there- fore, draw any conclusions as to specific functional roles based upon the average amine levels in the brain.
Whole brain putrescine concentrations are around 10-15 nmol g ~ in a variety of small species. Sper- midine varies between 300 and 600nmolg 1 and spermine between 280 and 410 nmol g-I wet tissue weight Seiler and Lamberty (1975a, b); Seiler and Schmidt-Glenewinkel (1975); Marton, Heby, Wilson and Lee (1974). Guinea pig is exceptional among mammals with regard to its high putrescine concen- tration (90 _+ 3nmolg-1) Seiler and Lamberty (1975b). In fish brain, however, putrescine concen- trations exceed those of spermidine and spermine Seiler (1973); Seiler and Lamberty (1973).
Regional patterns of putrescine and the polyamines are available only for rat, cat and human brains Harik and Snyder (1974); Seiler and Schmidt-Glene- winkel (1975); Kremzner (1973) but spermidine and

Polyamine metabolism and function in brain 101
spermine were studied in rabbit Halliday and Shaw (1976), sheep, dog, human Shaw and Pateman (1973) and monkey brains Michaelson and Coffman (1968); Russell, Gfeller, Marton and Legendre (1974).
As a general feature it appears that spermine con- centrations are relatively higher in structures rich in nerve cells, whereas white matter, spinal cord and peripheral nerves are extremely rich in spermidine. Putrescine concentrations usually follow the pattern of spermidine distribution. Figure 4 shows the distri- bution of the polyamines and of protein and RNA based on the DNA content, i.e., 'average" cellular con- tents of these cell constituents are compared. It appears from this figure that spinal cord and peri- pheral nerves contain much more spermidine and putrescine, as one would expect from the correspond- ing RNA and protein concentrations. This finding caused Mugnaini (1978) to suggest spermidine as a potential candidate for a short range molecular bridge between adjacent myelin membranes. In support of this interesting hypothesis is the autoradiographic
localization of spermidine in the myelin sheath of peripheral nerves Fischer, Schr6der and Seiler (1972b), Furthermore it was shown that spermidine has a slower turnover rate in myelin than in other structures of the nervous system Giorgi (1978). This finding was also interpreted in terms of a specific role of spermidine within the myelin structure. In myelin deficient mutants (quaking and jimpy mutant mice) deficiencies of spermidine were seen in the midbrain and spinal cord Russell and Meier (1975). This could also be taken in support of the above hypothesis.
In striking contrast to spermidine, the average cellular concentration of spermine is nearly constant in all brain regions, as appears from Fig. 4. This ob- servation is in strong support for different functions of spermine and spermidine. The preferential localiz- ation of spermine within the neuronal cell layers, together with the extraordinarily high spermine syn- thase activity in brain, as compared to other tissues suggest for spermine a specific role within the nerve cells.
~ 2
E I c
~nJ Pr°tein E
iO~ t Sperm/ne
Fig. 4. Distribution of putrescine, spermidine~ spermine RNA and proteins in the brain and peripheral nerves of adult rats. The values are expressed as molar ratios with DNA-phosphate. The hatched areas
indicate S.D. (From Seiler and Schmidt-Glenewinkel (1975)).

102 N. SELLER
~o.
36. o.
3z. Z O 28"
2/,.
E
lZ-
8 -
4 -
O"
2 6 -
°
o. ~ -
~ z2.
|,BI 16
1.¢..~
~0.16 o C~
1 0.12
ON.-
O- O05-
~ O03-
E
002.
ooi.
-7 ¥
e~ -a
0(~rth) 10 21 Days ~0[AdutO f t
Ptote~n ~
. . . . . . .~._.4 . . . . .
i
/ "
/ t " /
Specmidine , ~
°oi l° . . . " . .
. e e •
e..e"
j l
1,1,
o ~o ~o s~
A A -7 0 (Birth)
I "'~"-,,, .,& I='utr~sci~
i i I i ! i i i 900 I100 1300 1500 1700 1900 rng
Brom Weight
A A A 10 21 Days ~O(Adult }
Fig. 5. Developmental changes of polyamines, RNA and proteins in the brain of rats. The values are expressed as molar ratios with DNA-phosphate. The vertical bars indicate S.D. (The curves were
constructed using the experimental data of Seiler and Lamberty (1975)).

Polyamine metabolism and function in brain 103
POLYAMINES AND BRAIN DEVELOPMENT High putrescine levels are characteristic of imma-
ture brains, whereas the ratios of spermidine, spermine, to either RNA or DNA are usually not markedly different from those in the adult brains, although the brain weight at birth may only be a fraction of that of the adult animal Seiler and Lam- betty (1975b).
In agreement with the presumed role of putrescine in rapid cell proliferation J~inne et al. (1978); Wil- liams-Ashman and Canellakis (1979) maximal ornith- ine decarboxylase activity and putrescence levels co- incide in brain with the time of maximal nerve cell proliferation, and decreasing putrescine levels are the common characteristic of the developing brain in all
species studied so far Anderson and Schanberg (1972); Gilad and Kopin (1978); Seiler and Lamberty (1975a); Sturman and Gaull (1975); McAnulty, Yusuf, Dicker- son, Hey and Waterlow (1977); Heinrich-Hirsch, Ahlers and Peter (1977); Sobue and Nakajima (1977). This pattern is also typical for the developing retina de Mello er al. (1976); Macaione and Calatroni (1978). In contrast, S-adenosylmethionine decarboxy- lase activity is low at early stages of embryonal maturation Sturman and Gaull (1975); Gilad and Kopin (1978); Schmidt and Cantoni (1973). In rat brain this enzyme starts to increase its activity only from about day 10 of postnatal life. At this time, cellu- lar spermine levels have already reached adult values, and only spermidine concentrations tend to increase further at a later stage of development (Fig. 5). In other words, spermidine and spermine concentrations are not directly related to the S-adenosylmethionine decarboxylase activity, but are obviously regulated by other factors as well.
The situation is similar, although not completely identical in pure neuronal cultures from 8 day-old chick embryo hemispheres. In this system the cellular ornithine decarboxylase activity increased slightly when the neuroblasts started to differentiate. After three days in culture, however, ornithine decarboxy- lase activity and putrescine and spermine levels remained constant, whereas spermidine levels con- tinued to increase during the period of maturation, which was characterized by the excessive formation and elongation of neurites and dendrites, and the for- mation of synaptic contacts. S-adenosylmethionine decarboxylase activity doubled during the period of excessive cell maturation Seiler et al. (1980d),
The mammalian brain grows rapidly during the early phases of development but the growth rate de- creases dramatically after functional maturation. This can be seen in Fig. 5 for the rat brain. Its weight is
increased by a factor of about 7 during the first 21 days of postnatal life but during the following year the net weight gain is less than during the first three weeks.
In contrast functionally mature fish brains grow at a considerable rate in parallel with the overall growth of the body. In this case, putrescine, spermidine and spermine concentrations increase with the brain weight; both cellular hypertrophy and hyperplasia is observed, and it is most remarkable, that the increase of spermidine and putrescine parallel that of RNA and total protein, whereas spermine parallels that of DNA Seiler and Lamberty (1973). This is again an indication of differences in the functional roles of spermidine and spermine, and is perhaps the best example of the existence of direct quantitative re- lationships between the polyamines and nucleic acids in a growing physiologically complex system.
It has been pointed out that brain putrescine levels normally parallel the ornithine decarboxylase activity. Therefore it could be assumed that putrescine levels are exclusively regulated by the activity of this enzyme. During rat brain development, however, the total activity of the nuclear putrescine acetylating enzyme is reciprocal with the total putrescine level. Furthermore, the increase of the environmental tem- perature of trout is followed by a decrease of the brain putrescine pool, and an increase of the acetylase activity Seiler, Lamberty and AI-Therib {1975). In view of the role of acetylation as a catabolic step in polyamine degradation it is not impossible that the rate of putrescine acetylation may influence cerebral putrescine levels as well and may thus participate in the regulation of the rate of polyamine biosynthesis.
PHARMACOLOGICAL PROPERTIES
The pharmacological properties of the amine neurotransmitters contributed a great deal to our knowledge of their function. In the case of the amino acid neurotransmitters, however, much less could be derived from their overall actions and it was necess- ary to apply them locally and follow their actions by electrophysiological methods, in order to obtain use- ful information. The situation seems similar as far as the pharmacology of the polyamines is concerned,
The pharmacological properties of spermidine and spermine are qualitatively the same except that spermine usually acts at lower concentrations. Putres- cine also produces some of the effects of the polyamines, however, at very high doses. Central effects of the polyamines can be observed with high systemic doses, but all the neuropharmacological

104 N. SELLER
effects can also be produced by intracerebroventricu- lar administration of a few ,ug of the polyamines.
Putrescine injected in 100/~g amounts into the third cerebral ventricle of chicks produced 30 min later a significant depletion of GABA in the diencephalon, but not in the cerebral hemispheres. This effect appeared to be dependent on the inhibition of GAD Nistico, lentile~ Rotiroti and di Giorgi (1980). In con- trast, the putrescine effects in mice Anderson, Cross- land and Shaw (1975) have been suggested to be due to the increased formation of spermidine and spermine in the brain, and were described to occur with a latency of 24 h. It is well established that intra- ventricularly administered spermine and spermidine produce convulsions, which develop over a period of several hours Anderson et al. (1975). Convulsions and electrocortical epileptogenic discharges were rapidly seen in the chicken however, following a 5 10min period of sedation. Since these are known to occur after depletion of GABA Meldrum (1978) it was sug- gested that intraventricular putrescine impairs GABA neurones.
In agreement with this finding it was shown by Pajunen (1979) that intraventricular injections in mice of 10 nmol of spermine caused an increase of ornith- ine decarboxylase, a fluctuation of GAD activities and a decrease of S-adenosylmethionine decarboxylase ac- tivity within 2 h. It led to putrescine accumulation, with a maximum at 30 h and a significant decrease of brain GABA levels. Similar although less marked results were obtained with spermidine and putrescine.
Although speculative at present, it may nevertheless be of interest in this connection to summarize briefly a few biochemical and pharmacological findings, which seem to suggest interrelationships between the polyamines and GABA other than the metabolic con- version of putrescine into GABA Seiler (1980}.
If rats are exposed to hyperbaric oxygen, so that they are convulsing, they show a decrease of brain polyamine levels by 403,, within less than 4 h Tsvet- nenko and Shugalev (1978). The interrelationships between brain GABA and exposure to hyperbaric oxygen are well established Wood (19751. Further ex- ploration of this model could therefore produce insights into the relationships between polyamines and convulsive states as well as into GABA- polyamine interrelations. The finding, that allylglycine administration produces an increase of ornithine decarboxylase activity with a concomitant elevation of brain putrescine levels, and a decrease of S-adeno- sylmethionine decarboxylase activity, although with no significant effect on spermidine or spermine Paju- nen, Hietala, Baruch-Virransalo and Piha (1978b!
belongs to the same category of findings. However, it should be emphasized, that these observations are dif- ficult to interpret: the administration of a specific in- hibitor of GABA-T which increased brain GABA levels, also produced an increase of ornithine decar- boxylase activity and of putrescine concentrations, and a decrease of S-adenosylmethionine decarboxy- lase activity, although the sequence of the events was somewhat different, compared to that after allylgly- cine administration Seiler, Bink and Grove (1979b, 1980a). In other words depletion and elevation of GABA by inhibition of two different enzymes led to comparable changes of brain polyamine metabolism. It appears from these obviously conflicting results that only well defined models and very specifically acting inhibitors of GABA and polyamine metab- olism will be useful in the future elucidation of the presumed interactions between these systems. The effects of compounds with ill-defined action mechan- isms such as fl-(p-chlorophenyl)-7-aminobutyric acid Pajunen, Virransalo, Hietala and Piha (1978c) or models of very complex physiological events, such as electroconvulsions, Pajunen, Virransalo and Piha 1978a) are of restricted value in this regard. In future work in this area, general metabolic changes which are concomitant with changes in brain GABA levels, have also to be emphasized. It is known, for instance, that in anoxia, ornithine decarboxylase activity is enhanced Kleihues, Hossmann, Pegg, Kobayashi and Zimmermann (1975). Spermidine and spermine are in
vitro reversible, non-competitive inhibitors of succinic semialdehyde dehydrogenase (EC 1.2.1.16) Lapinjoki Pajunen, Hietala and Piha (1980). This finding sug- gests a potential target enzyme for polyamines as regulators of GABA degradation.
With regards to the presumed role of GABAergic neurones in aggressive behavior Mandel, Cisielski, Maitre, Simler, Mack and Kempf (19791 some behav- ioral studies are of interest. Mice made aggressive by isolation showed an increase of brain spermidine con- centration which paralleled the development of aggressivity. It returned to normal values when the aggressive behavior was reversed by grouped housing Tadano (19741; Tadano, Onoki and Kissara (1974).
There are several lines of evidence that polyamines might interact with cholinergic neuronal systems. Large doses (>0.1 mmolkg 1) of spermine given intraperitoneally produce a curare-like paralysis of the musculature and the animals die from respirat- ory arrest Shaw (1972); Buss (1963). In the isolated guinea pig ileum polyamines selectively antagonize the contractions produced by nicotine Shaw (1972). The inhibition of acetylcholinesterase (EC 3.1.1.7.) in

Polyamine metabolism and function in brain
E ,4j
~o
3C
20
10
I I f
o 1o -~ lo -3 [ocetylchohne ] [iv/]
Fig. 6. The influence of spermine on the activity of membrane bound to acetylcholinesterase (EC 3.1.1.17) as a function of acetylcholine concentration. The following spermine concentrations were used V0mM; A0.001 mM; O0.005 raM; O0.01 mM; A0.02 raM: []0.04 mM; O0.1 mM; 0.2 raM; B0.5 mM
(Data from Kossorotow, Wolf and Seiler (1974).)
105
the presence of low acetylcholine concentrations by micromolar spermidine and spermine concentrations, and the activation of this enzyme under the same con- ditions, however, at higher acetylcholine concen- trations (Fig. 6) Kossorotow, Wolf and Seiler (1974); Anand, Gore and Kerkut (1976); Peter, Gies, Neu- meier, Sch~idler and Wegener (1979) would fit into a modulatory role of the polyamines. However, doses of spermidine and spermine which produce central effects did not affect brain acetylcholine content (or the contents of catecholamines, serotonin and GABA) Shaw (1977).
It has previously been shown that electrophoreti- cally applied polyamines depress spontaneous firing of neurones in the brain stem of the rat and the cat Wedgwood and Wollstencroft (1977). This suggested that polyamines are somehow involved in the control of neuronal activity. However perfusion of the iso- lated head ganglia of the snail with up to 0.1 mM putrescine, spermidine and spermine had no effect on the spontaneous activity of the neurones Gould and Cottrell (1976).
The question has been raised by several investiga- tors whether the polyamines are acting as neurotrans- mitters. At present there seems to be no evidence for such role, although spermidine and spermine have a higher affinity to membranes of the synaptic region, than to any other membrane fraction isolated from brain cortex homogenates Seiler and Deckardt (1976, 1978a). The fact that there is no energy dependent uptake of putrescine and the polyamines by synapto- somes, in contrast with ornithine Seiler and Deckardt (1976, 1978b) must not necessarily be interpreted as an argument against a transmitter role. Low affinity
uptake systems have been described for brain cortex slices Lajtha and Sershen (1974); Pateman and Shaw (1975). These or conjugation reactions, such as acetyl- ation, could play a role in physiological inactivation mechanisms. The stimulated release of polyamines from brain cortex, reported for monkey brain Russell et al. 11974) could, however, not be observed in guinea pigs Seller and Deckardt (1976).
Some evidence is available for the binding of spermine to synaptic vesicles Seiler and Deckardt (1976). Among the many compounds tested, sper- midine and spermine were the most effective in inhi- biting binding of acetylcholine to synaptic vesicles Kuriyama, Roberts and Vos (1968). These findings may be taken as a further indication for a potential modulatory role of the polyamines in the cholinergic system.
A general influence of the polyamines on ion trans- ports is suggested by the interaction of the polyamines with ATPases Peter et al. (1973, 1979); Tashima, Hasegawa and Mizunuma (1978): Nag and Ghosh (1973).
MODULATION OF POLYAMINE METABOLISM
Several eukaryotic cell types respond to a variety of hormones, growth promoting agents and drugs by a rapid induction of ornithine decarboxylase. It is there- fore not surprising, that similar findings were also reported for the vertebrate brain. Thus, nerve growth factor Lewis, Lakshmanan, Nagaiah, McDonnel and Guroff (1978), electrical stimulation Pajunen et al.
(1978a), reserpine Deckardt, Pujol, Belin, Seiler and Jouvet (1978), Dopa Harik (1979), and some other drugs Belin and Pujol (1977); Pajunen et al. (1978c);

106 N. SEILER
Slotkin (1979} increase the activity of this enzyme. Most interesting in this regard, however, are the find- ings that hormones such as thyroxine accelerate and corticosteroids prolong the pattern of post-natal rat brain development, and the developmental pattern of ornithine decarboxylase is changed accordingly Anderson and Schanberg (1975). Even the interrup- tion of the normal mother-infant interaction in rats caused an immediate decrease of brain ornithine decarboxylase activity, which was accompanied by declines of putrescine levels Butler and Schanberg (1977); Butler, Suskind and Schanberg (1978). Thus ornithine decarboxylase turned out to be a sensitive tool for the study of normal and perturbed ontogeny of the CNS and PNS, Slotkin (1979).
The biochemical consequences which are directly related to the induction of ornithine decarboxylase are, however, poorly understood at present. Thus it was postulated from the decrease of ornithine decar- boxylase activity which was induced by prenatal re- serpine administration, and the subsequent deficits in tyrosine hydroxylase, that a cause-and-effect relation- ship exists between the two events, and the existence of a prenatal period was defined, during which these effects of reserpine can be produced Bartolome, Seidler, Anderson and Slotkin (1976); Lau, Barto- lome, Seidler and Slotkin (1977). From the fact that inhibition of the nerve growth factor-induced ornith- ine decarboxylase in a clonal line of phaechromocy- toma cells did not prevent the differentiation of these cells, it was concluded, on the other hand, that the induction of ornithine decarboxylase in this system was not a step within the cascade of events, which finally lead to cell differentiation, but was presumably a 'dead end' or a terminal consequence of other events related to the interaction of nerve growth factor with its target cell Greene and McGuire (1978).
It is probably premature to attempt to draw far reaching conclusions about the significance of induced ornithine decarboxylase. But of course it is not unlikely, that many of the events which lead to a temporary increase of the activity of this enzyme may indeed be without any long lasting physiological con- sequence, and that a specific cellular environment is a prerequisite for the trigger function of this enzyme.
AXONAL TRANSPORT
It has been demonstrated that synaptosomes con- tain only a very low, if any, ornithine decarboxylase activity Seiler and Sarhan (1980). On the other hand the presence of putrescine in this neuronal compart- ment is most likely Seiler and Deckardt (1976; 1978a).
Disregarding the possibility that even very low ornithine decarboxylase activities may be able to satisfy the local putrescine needs, it was speculated that transport of the polyamines through the axon and their interconversion may be a possible source of putrescine Ingoglia et al. (1978). However, it appears that axonal polyamine transport occurs only in grow- ing axons, both during embryonic development and regeneration Fischer and Schmatolla (1972): Ingoglia and Sturman (1978), but it is absent in the visual sys- tem of mature rats Siegel and McClure (1975~ goldfish Ingoglia and Sturman (1978) and the R 2 neuron of Aplysia californica Kremzner and Abron (1979).
CONCLUSIONS
Polyamines are intimately involved in growth pro- cesses. This fact has been evidenced from their increased concentrations in proliferating systems. Examples related to the nervous system are the dege- nerating peripheral nerve Seiler and Schr6der (1970) experimental tumors Seller et al. (1975); Marton and Heby (1974) and human brain tumors Harik and Sutton (1979); Mirzoyan and Promyslov (19791; Kremzner (1973) and the scrapie affected mouse brain Giorgi, Field and Joyce (1972). The enhanced formation of putrescine and spermidine by gliomas has been used for the diagnosis of brain tumors by polyamine assay in the CSF, and especially for the monitoring effects of therapy Marton, Heby, Levin, Lubich, Crafts and Wilson (1976); Marton, Edwards, Levin, Lubich and Wilson (1979): Seidenfeld and Marton (1979).
Direct evidence for an essential role of polyamines in cell growth and proliferation came from the devel- opment of specific inhibitors of ornithine decarboxy- lase. By depletion of cellular putrescine and sper- midine concentrations, cultured cells of various types are arrested in growth, mostly in the G l phase Mamont et al. (1978); Heby and J~inne (1981).
It is not surprising that many of the in vitro effects of the polyamines exerted on enzymes involved in the transcription and translation of the genetic infor- mation, which were originally found in bacterial sys- tems or in liver, have also been detected in subcellular preparations of brain tissue. The stimulation of nuclear RNA polymerase II, suggests that the forma- tion of DNA-like RNA may be modulated by polyamines Singh and Sung (1972). The stimulation of the binding of template RNA and aminoacyl RNA to ribosomes Goertz (1979) suggests an important role of the polyamines in protein biosynthesis, and the stimu- lation of acetylation and phosphorylation of histories

Polyamine metabolism and function in brain 107
Das and Kanungo (1979) suggests an involvement of the polyamines in the al terat ion of gene expression during brain development. These few examples may be sufficient to underline the presumed multifunctio- hal role of the polyamines. However, it is not possible at present, to pinpoint exclusive brain functions of the polyamines, But there is evidence arising in various areas, among which perhaps a role of the polyamines in membrane structures is most interesting. With regard to the nervous system, we are presumably only at the beginning of a development which might lead to new and unexpected findings related to these long neglected simple aliphatic amines.
REFERENCES
Anand, R., Gore, M. G. and Kerkut, G. A. (1976). The effect of spermine and spermidine on the hydrolysis of acetylthiocholine in the presence of rat caudate nucleus homogenate or acetylcholinesterase from electrophorus electricus. J. Neurochem. 27, 381-385.
Anderson, T. R. and Schanberg, S. M. (1972). Ornithine decarboxylase activity in developing rat brain. J. Neuro- chem. 19, 1471-1481.
Anderson, T. R. and Schanberg, S. M. (1975), Effect of thyroxine and cortisol on brain ornithine decarboxylase activity and swimming behavior in developing rat. Bio- chem. Pharmacol. 24, 495 501.
Anderson, D. J., Crossland, J. and Shaw, G. G. (1975) The actions of spermidine and spermine on the central ner- vous system, Neuropharmacology 14, 571-577.
Bachrach, U. (1973) Function of the Naturally Occurrin~j Polyamines. Academic Press, New York.
Bartolome, J., Seidler, F. J., Anderson, T. R. and Slotkin, T. A. (1976). Effects of prenatal reserpine administration on development of the rat adrenal medulla and central nervous system. J. Pharmacol. expl Therap. 197, 293 302.
Belin, M, F. and Pujol, J.-F. (1977). Effect of 6-hydroxy- dopamine on ornithine decarboxylase activity in central monoaminergic systems of rat. Biochem. Pharmacol. 26, 2473-2475.
Blankenship, J. (1978), Deacetylation of NS-acetylsper - midine by subcellular fractions of rat tissue. Archs Bio- chem. Biophys. 189, 20-27.
Blankenship, J. and Walle. T. (1978). In vitro studies of enzymatic synthesis and metabolism of N-acetylated polyamines. In: Advances in Polyamine Research, Vol. 2 Campbell, R, A.. Morris, D., Bartos, D., Daves, G.D. Jr., and Bartos F. (Eds) pp. 97 I10. Raven Press, New York.
Bolkenius, F. and Seiler, N. (1981). Acetylderivatives as intermediates in polyamine catabolism. Int. J. Biochem, 13, 287-292.
Burckard, W. P., Gey, K. F. and Pletscher, A. (1963). Diamine oxidase in the brain of vertebrates, J. Neuro- chem. 10, 183 186.
Buss, J. 11963). Die pharmakologischen Wirkungen yon Gamma-aminobutyrylcholin und yon Spermin, Sper- midin und Putrescin. M.D. Thesis, Freie Universit~it Ber- lin. Germany.
Butler, S. R. and Schanberg, S. M. (1977). Effect of mater-
nal deprivation on polyamine metabolism in prewean- ling rat-brain and heart. Life Sci. 21,877-884.
Butler. S. R., Suskind, M. R. and Schanberg, S. M. (1978). Maternal behavior as a regulator of polyamine biosyn- thesis in brain and heart of the developing rat pup. Science 199, 445-447.
Canellakis, E. S., Vicepts-Madore, D., Kyriakidis, D. A. and Heller, J. S. (1979). The regulation and function of ornithine decarboxylase and of the polyamines. Current Topics in Cell. Regul. 15, 155-200.
Cohen, S. S. (1971) Introduction to the Polyamines. Pren- tice-Hall, Engelwood Cliffs, New Jersey.
Cohen, S. S. (1978). What do polyamines do'? Nature 274, 209-210.
Das, R. and Kanungo, M. S. (1979). Effects of polyamines on in vitro phosphorylation and acetylation of histones of the cerebral cortex of rats of various ages. Biochem, Biophys. Res. Commun. 90, 708-714.
Deckardt, K., Pujol, J.-F., Belin, M. F., Seiler, N. and Jou- vet, M. (1978). Increase of ornithine decarboxylase ac- tivity elicited by reserpine in the peripheral and central monoaminergic systems of the rat. Neurochem. Res. 3, 745-753.
De Mello, F. G., Bachrach, U. and Nirenberg, M. (1976). Ornithine and glutamic acid decarboxylase activities in the developing chick retina J. Neurochem. 27, 847 851.
Fischer, H. A. and Schmatolla, E. (1972). Axonal transport of tritium labelled putrescine in the embryonic visual system of zebrafish. Science 176, 1327-1329.
Fischer, H. A., Korr, H., Seiler, N. and Werner, G. (1972a). Interrelationships between polyamines and nucleic acids I11. Metabolic and autoradiographic studies of putres- cine in mouse brain. Brain Res. 39. 197 212.
Fischer, H. A., Schr6der, J. M. and Seiler, N. (1972b) Inter- relationships between polyamines and nucleic acids IV. Autoradiographic localization of -~H-putrescine and its metabolites in normal and degenerating peripheral nerves, and spinal ganglia. Z. Zellforsch. 128, 393 405.
Gila& G. M. and Kopin, 1. J. (1979), Neurochemical aspects of neuronal ontogenesis in the developing rat cerebellum--Changes in neurotransmitter and polyamine synthesizing enzymes. J. Neurochem. 33, 1195- 1204.
GiorgL P. P. (1978). Spermidine: a constituent of the mye- lin sheath? Neurosci. Lett. 10, 335-340.
Giorgi, P. P., Field, E. J. and Joyce, G. (1972). Metabolism of polyamines in normal and scrapie affected mouse brain and spleen. J. Neurochem. 19, 255-264.
Goertz, B. (1979) Effect of polyamines on cell-free protein synthesizing systems from rat cerebral cortex, cerebellum and liver. Bruin Res. 173, 125 135.
Gould, R. M. and Cottrell, G. A. (1974). Putrescine in mol- luscs: Identification and occurrence in neurons and other tissues. Comp. Biochem. Physiol. 48B, 591 597.
Greene, L. A. and McGuire, J. C. (1978). Induction of ornithine decarboxylase by nerve growth factor dis- sociated from effects on survival and neurite outgrowth. Nature 276, 191-193.
Halliday, C. A. and Shaw, G. G. (1976). The distribution and metabolism of putrescine, spermidine and spermine injected into cerebral ventricles of rabbit. J. Neurochem. 26, 1199-1206.
Harik, S. I. (1979). Effects of L-Dopa on putrescine levels in the brain and the liver of the rat. Eur. J. Pharmacol. 54, 235 242.

108 N. SELLER
Harik. S. 1. and Snyder, S. H. ~1974). Putrescine: Regional distribution in the nervous system of the rat and the cat. Brain Res. 66, 328 331.
Harik, S. I. and Sutton, C. H. (1979). Putrescine as a bio- chemical marker of malignant brain tumors. Cancer Res. 39, 501(~5015.
Heby, O. and Andersson, G. (1980), Polyamines and the cell cycle. In: Polyamines in Biochemical Research. (Gaugas, J. M.Ed.) pp. 17 34. John Wiley, Chichester.
Heby, O. and Jfinne, J. (1981). Polyamine antimetabolites: biochemistry, specificity and biological effects of inhibi- tors of polyamine synthesis. In: Polyamines in Biology and Medicine. (Morris, D. R. and Marton, L. H. Eds.) Marcel Dekker, New York.
Heinrich-Hirsch, B., Ahlers, J. and Peter, H. W. (1977). Inhibition of Na, K-ATPase from chick brain by polyamines. Encymes 22, 235-241.
H/51ttfi. E. (1977). Oxidation of spermidine and spermine in rat liver: purification and properties of polyamine oxi- dase. Biochemistry 16, 91-100.
lngoglia, N. A. and Sturman, J. A. (1978). Axonal transport of putrescine, spermidine and spermine in the goldfish visual system: speculation on the association of axonally transported spermidine and tRNA in regenerating optic nerve. In: Adcances in Polvamine Research Vol. 2. Camp- bell, R. A.. Morris, D. R.. Bartos, D., Daves, G. D. Jr., and Bartos. F. (Eds). pp. 169 182. Raven Press, New York.
J~inne, J.. P/Ss6. H. and Raina, A. (1978). Polyamines in rapid growth and cancer Biochim. hiophys. Acta 473, 241 293.
Kleihues, K. A., Hossmann, K. A., Pegg, A. E., Kobayashi, K. and Zimmermann, V. (1975). Resuscitation of the monkey brain after one hour complete ischemia I11. Indications of metabolic recovery. Brain Res. 95, 61 75.
Konishi, H., Nakajima, T. and Sano, I. (1977). Metabolism of putrescine in the central nervous system, d. Biochem. (Tokyo) 81,355 360.
Kossorotow, A., Wolf, H. U. and Seiler, N. (1974). Regula- tory effects of polyamines on membrane-bound acetyl- cholinesterase. Biochem. J. 144, 21 27.
Kremzner. L. T. (1973~. Polyamine metabolism in normal and neoplastic neural tissue. In: Polyamines in Normal and Neoplastic Growth (Russell, D. H. Ed.) pp. 27-40. Raven press, New York.
Kremzner. L. T. and Abron. R. T. (1979). Metabolism and axonal transport of polyamines in an identified neuron. Trans. Am. Soc. Neurochem. 10, 208.
Kremzner, L. T., Hiller. J. M. and Simon. E. J. (1975). Metabolism of polyamines in mouse neuroblastoma cell culture: formation of GABA and putreanine. J. Neuro- then1. 25, 889 894.
Kuriyama, K., Roberts, E. and Vos, J. (1968~. Some charac- teristics of binding of 7-aminobutyric acid and acetylcho- line to a synaptic vesicle fraction from mouse brain. Brain Res. 9, 231 252.
Lajtha, A. and Sershen, H. (1974). Substrate specificity of uptake of diamines in mouse brain slices. Archs Biochem. Biophys. 165, 539 547.
Lapinjoki. S. P., Pajunen, A. E. I., Hietala, O. A. and Piha, R. S. (1980). The effect of potyamines on the enzymes of the N-aminobutyric acid metabolism in mouse brain in citro. FEBS Lett. 112, 289 292.
Lau, C., Bartolome, J., Seidler. F. J. and Slotkin, T. A. (1977L Critical periods for effects of prenatal reserpine
administration on development of rat brain and adrenal medulla. Neuropharmacology 16, 799-809.
Lewis, M. E., Lakshmanan, J., Nagaiah, K., McDonnell, P. and Guroff, G. (1978). Nerve growth-factor increases ac- tivity of ornithine decarboxylase in rat brain Proc. nam. Acad. Sci. U.S.A. 75, 1021- 1023.
Libby, P. R. (1978). Calf liver nuclear N-acetyltransferases. Purification and properties of two enzymes with both spermidine acetyltransferase and histone acetyltransfer- ase activities. J. Biol. Chem. 253, 233 237.
Macaione, S. and Caltroni. A. (1978). Polyamines and ornithine decarboxylase activity in the developing rat retina. Life Sci. 23, 683-690.
Mamont, P. S., Duchesne, M.-C., Joder-Ohlenbusch, A. M. and Grove, J. (1978). Effects of ornithine decarboxylase inhibitors on cultured cells. In: Enzyme-Actit~ated lrre- cersihle lnhibitors. Seller. N., Jung, M. J. and Koch- Weser, J. (Eds). pp. 43 54. Elsevier/North-Holland Bio- medical Press, Amsterdam.
Mamont, P. S., Joder-Ohlenbusch, A.-M., Nussli, M. and Grove, I. J. (1981). Indirect evidence for a strict negative control of S-adenosyl-L-methionine decarboxylase by spermidine in rat hepatoma cells. Biochem. J. 196, 411 422.
Mandel, P., Cisielski, L. Maitre, M. Simler, S., Mack, G. and Kempf, E. (1979) Involvement of central GABAergic systems in convulsions in aggressive behavior. In: GABA Biochemistry and CNS function Adv. In Exp. Med. and Biol. Vol. 123 Mandel, P. and Defeudis, F. V. (Eds). pp. 475-492. Plenum Press, New York.
Marton, L. J. and Heby, O. (1974). Polyamine metabolism in tumor, spleen and liver of tumor-bearing rats. Int. J. Cancer 13, 619-628.
Marton. L. J., Heby. O., Wilson, C. B. and Lee, P. L. Y. (1974). A method of the determination of polyamines in cerebrospinal fluid. FEBS Left. 46, 305-307.
Marton, L. J., Heby, O., Levin, V. A.. Lubich, W. P., Crafts, D. C. and Wilson, C. B. (1976). The relationship of polyamines in cerebrospinal fluid to the presence of cen- tral nervous system tumors. Cancer Res. 36, 973-977.
Marton. L. J., Edwards, M. S.. Levin, V. A.. Lubich, W. P. and Wilson, C. B. (1979) Predictive value of cerebro- spinal fluid polyamines in medulloblastoma. Cancer Res. 39, 993-997.
Matsui, I. and Pegg, A. E. (1980a). Increase in acetylation of spermidine in rat liver extracts brought about by treatment with carbontetrachloride. Biochem. Biophys. Res. Commun. 91, 1009-1015.
Matsui, I. and Pegg, A. E. (1980bl. Effect of thioacetamide, growth hormone on partial hepatectomy on spermidine acetylase activity of rat liver cytosol Biochim. hiophys. Acta 633, 87 94.
Maudsley, D. V. (1979). Regulation of polyamine biosyn- thesis. Biochem. Pharmacol. 28, 153 161.
McAnulty, P. A., Yusuf, H. K. M., Dickerson, J. W. T., Hey, E. N. and Waterlow, J. C. (1977). Polyamines of human brain during normal fetal and postnatal growth and during postnatal malnutrition. J. Neurochem. 28, 1305 1310.
McCormick, F. (1978~. Kinetics of polyamine synthesis and turnover in mouse fibroblasts. Biochem. J. 174, 427 434.
Meldrum, B. S. (1978). Gamma-Aminobutyric acid and the search for new anticonvulsant drugs. Lancet II, 304~306.
Michaelson, I. A., Coffman, P. Z. and Vedral, D. F. 11968). The regional distribution of histamine in brain in the

Polyamine metabolism and function in brain 109
Rhesus monkey (Macaca mulatta}. Biochem. Pharmacol. 17, 2435-2441.
Mirzoyan, P. A. and Promysiov, M. Z. (1979). Contents of putrescine, spermidine and spermine in tissue of the human brain glial tumors. Ukr. Biokhim. Zh. 51, 474 -476.
Morris, D. R. and Harada, J. (1980). Participation of polyamines in the proliferation of bacterial and animal cells. In: Pol)mmines in Biomedical Research. Gaugas, J. M. (Ed.) pp. 1 16. John Wiley, Chichester.
Mugnaini, E. (1978). Fine structure of myelin sheaths. Proc. Europ. Soc. Neurochem. Vol. 1. pp. 3-31. Verlag Chemie, Weinheim.
Nag, D. and Gosh, J. J. (1973). lmipramine induced changes of brain adenosine triphosphatase activity. The role of spermine in counteracting the disorganizing effect of the drug on membrane ATPase. J. Neurochem. 20, 1021 1027.
Nakajima, T. (1973). Formation of putreanine, N-(4-amino- butyl)-3-aminopropionic acid from spermidine in rat brain and liver. J. Neurochem. 20, 735-742.
Nistico, G., Ientile, R., Rotiroti, D. and di Giorgi, R. M. (1980). GABA depletion and behavioral changes pro- duced by intravenlricular putrescine in chicks. Biochem. Pharmacol. 29, 954 957.
Pajunen, A. E. I. (1979i. Polyamine metabolism in mouse brain. Acta Universitatis Ouluensis, Set. A. Sci. Rer. nat. No 79, pp. I 47.
Pajunen, A. E. 1., Hietala, O. A., Virransalo, E. L. and Piha, R. S. (1978a). Ornithine decarboxylase and adenosylmethionine decarboxylase in mouse brain- effect of electrical stimulation. J. Neurochem. 30, 281 283.
Pajunen, A. E. 1., Hietala, O. A., Baruch-Virransalo, E. L. and Piha, R. S. (1978b), The effect of DL-allylglycine on Polyamine and GABA metabolism in mouse brain, d. Neurochem. 32, 1401 1408,
Pajunen, A. E. I., Virransalo, E. L., Hietala, O. A. and Piha, R. S. I1978c). Stimulation of n-ornithine and S-adenosyl- k-methioninc decarboxylase by fl-(p-chlorophenyl)-?- aminobutyric acid in mouse tissues. Acta Chem. Scand. B32, 322 326.
Pateman, A. J. and Shaw, G. G. 119751. The uptake of spermidine and spermine by slices of mouse cerebral hemispheres. J. Neurochem. 25, 341-345.
Peter, H. W., Wolf, H. U. and Seiler, N. Influence of polyamines on two bivalent cation-activated ATPases. Hoppe-Seyler's Z. Physiol. Chem. 354, 1146-1148.
Peter, H. W., Gies, A., Neumeier, M., Sch~dler, R. and Wegener, I. 11979). Influence of the naturally occurring polyamines spermine, spermidine and putrescine on the kinetic properties of acetylcholinesterase, comparative studies with the acetylcholinesterases from the central nervous system of Manduca sexta and the synaptic plasma membrane of rat brain. Gen, Pharmacol. 10, 133-14l.
Raina, A., Pajula, R. L. and Eloranta, T. (1976). A rapid assay method for spermidine and spermine synthase, Distribution of polyamine synthesizing enzymes and methionine adenosyhransferase in rat tissues. FEBS Lett. 67, 252 256.
Raina, A., Eloranta, T., Pajula, R. L., M/intyj/irvi, R. and TuomL K, 1"1980). Polyamines in rapidly growing tissues. In: Polvamines in Biomedical Research Gaugas, J. M. {Ed.). pp. 35-50. John Wiley, Chichester.
Russell, D. H. (1973). Polyamines in Normal and Neoplastic Growth. Raven Press, New York.
Russell, D. H. and Meier, H. (1975). Alterations in the accumulation patterns of polyamines in brains of myelin deficient mice. J. Neurohiol. 6, 267-275.
Russell, D. H., Gfeller, E., Marton, L. J. and Legendre, S. M. (1974). Distribution of putrescine, spermidine and spermine in Rhesus monkey brain: decrease in sper- midine and spermine concentrations in motor cortex after electrical stimulation. J. Neurobiol. 5, 349 354.
Schmidt, G. L. and CantonL G. L. (1973). Adenosylmethio- nine decarboxylase in developing rat brain. J. Neuro- chem. 20, 1373-1385.
Seidenfeld, J. and Marton, L. J. (1979). Biochemical markers of central nervous system tumors measured in cerebrospinal fluid and their potential use in diagnosis and patient management: a review. J.N.C.I. 63, 919 931.
Seiler, N. (1973). Polyamine metabolism in the brain. In: Polyamines in Normal and Neoplastic Growth. Russell, D. H. (Ed.). pp. 137.-156. Raven Press, New York.
Seiler, N. (1980). On the role of GABA in vertebrate polyamine metabolism. Physiol. Chem. & Ph)'sics 12, 411 429.
Seller, N. (198l). Amide bond forming reactions of the polyamines. In: Polyamines in Biolo#y and Medicine. Morris, D. R. and Marton, L. H. tEds). Marcel Dekker. New York.
Seiler, N. and AI-Therib, M. J. (1974ak Putrescine catab- olism in mammalian brain Biochem. J. 144, 29 35.
Seiler, N. and AI-Therib, M. J. (1974b). Acetyl-CoA: h4-diaminobutane N-acetyl transferase. Occurrence in vertebrate organs and subcellular localization. Biochim. biophys. Acta 354, 206-212.
Seller, N. and Deckardt, K. (1976). Association of putres- cine, spermidine, spermine and GABA with structural elements of brain cells. Neurochem. Res. I, 469 499.
Seiler, N. and Deckardt, K. (1978a). Association of polyamines and GABA with structural elements of brain cells. In: Adcances m Polyamines Research Vol. 2. Camp- bell, R. A., Morris, D. R., Bartos, D., Daves, G. D. Jr., and Bartos. F. (Eds). pp. 145 159. Raven Press, New York.
Seller, N. and Deckardt, K. t1978h). Uptake of polyamines and related compounds into nerve endings. In: Adl'ances in Polvamine Research. Vol. 2 Campbell, R. A,, Morris, D. R., Bartos, D., Dares, G. D. Jr., and Bartos. F. (Edst. pp. 161 167. Raven Press. New York.
Seller, N. and Eichentopf, B. (1975). 4-Aminohutyrate in mammalian putrescine catabolism. Biochem. J. 152, 201 210.
Seiler, N. and Kn6dgen, B. (1971). Die Umwandlung yon Glutamins~ure, Putrescin, und Ornithin in die t'-Amino- buttersgure im Gehirn. Hoppe-Seyler's Z. Physiol. Chem. 352, 97-105.
Seller, N. and Lamberty, U. (1973t. Interrelationships between polyamines and nucleic acids. Changes of polyamine and nucleic acid concentrations in the grow- ing fish brain [Salmo irideus Gibb.) J. Ncurochem. 20, 709 717.
Seiler. N. and Lamberty, U. (1975a). Interrelations between polyamines and nucleic acids: changes of polyamines and nucleic acid concentrations in developing rat brain. J. Neurochem. 24, 5-13.
Seiler, N. and Lamberty, U. t1975b). Polyamines and nuc- leic acids in brains and hearts of newborn and autopha-

110 N. SEILER
gous and heterophagous vertebrates. Comp. Biochem. Physiol. 52B, 419-425.
Seiler, N. and Sarhan, S. (1980). On the nonoccurrence of ornithine decarboxylase in nerve endings. Neurochem. Res. 5, 97-100.
Seller, N. and Schmidt-Glenewinkel, T. (1975). Regional distribution of putrescine, spermidine and spermine in relation to the distribution of RNA and DNA in the rat nervous system. J. Neurochem. 24, 791 795.
Seiler, N. and Schr6der, J. M. (1970). Beziehungen zwis- chen Polyaminen und Nucleinsiiuren. II. Biochemische und feinstrukturelle Untersuchungen am peripheren Nerven w~ihrend der Wallerschen Degeneration. Brain Res. 22, 81-103.
Seiler, N., Wiechmann, M., Fischer, H. A. and Werner, G. (1971). The incorporation of putrescine carbon into /-aminobutyric acid in rat liver and brain in vivo. Brain Res. 28, 317-325.
Seiler, N., AI-Therib, M. J. and Kataoka, K. (1973). Forma- tion of GABA from putrescine in the brain of fish (Salmo irideus Gibb.) J. Neurochem. 20, 699 708.
Seiler, N., Lamberty, U. and AI-Therib, M. J. (1975). Ace- tylcoenzymeA: 1,4-diaminobutane N-acetyltransferase in rat brain during development in experimental brain turnouts and in brains of fish of different metabolic ac- tivity. J. Neurochem. 24, 797-800.
Seiler, N., Danzin, C., Prakash, N. J. and Koch-Weser, J. (1978). Effects of ornithine decarboxylase inhibitors in vivo. In: Enzyme-Activated Irreversible Inhibitors. Seller, N., Jung, M. J. and Koch-Weser, J. (Eds). pp. 55 72. Elsevier/North Holland Biomedical Press, Amsterdam.
Seller, N., A1-Therib, M. J., Fischer, H. A. and Erdmann, G. (1979a). Dynamic and regional aspects of polyamine metabolism in the brain of trout (Salmo irideus Gibb.) Int. J. Biochem. 10, 961-974.
Seiler, N., Bink, G. and Grove, J. (1979b). Regulatory Inter- relations between GABA levels and polyamine metab- olism. Neurochem. Res. 4, 425-435.
Seiler, N., Schmidt-Glenewinkel, T. and Sarhan, S. (1979c) On the formation of ";-aminobutyric acid from putres- cine in brain. J. Bioehem. (Tokyo) 86, 277-278.
Seiler, N., Bink, G. and Grove, J. (1980a). Relationship between GABA and polyamines in developing rat brain. Neuropharmaeology 19, 251-258.
Seiler, N., Bolkenius, F. N. and Kn6dgen, B. (1980b). Acetylation of spermidine in polyamine catabolism. Bio- chim. biophys. Acta 633, 181 190.
Seiler, N., Bolkenius, F. N., Kn6dgen, B. and Mamont, P. (1980c). Polyamine oxidase in rat tissues. Bioehim. bio- phys. Acta 615, 480-488.
Seiler, N., Sarhan, S. and Roth-Schechter, B. F. (1981). Developmental changes of the GABA and polyamine systems in isolated neurons in cell culture. Devel. Neuro- sci. 4, 181-187.
Shaskan, E. G. and Snyder, S. H. (1973) Polyamine turn- over in different regions of rat brain J. Neurochem. 20, 1453 1460.
Shaw, G. G. (1972). Some pharmacological properties of the polyamines spermine and spermidine--a re-apprai- sal. Arch. Int. Pharmacodyn. ThOr. 198, 36-48.
Shaw, G. G. (1977). Evidence against the view that the central actions of polyamines are indirectly mediated. Biochem. Pharmacol. 26, 1450-1451.
Shaw, G. G. (1979a) The polyamines in the central nervous system. Bioehem. Pharmacol. 28, 1-6.
Shaw, G. G. (1979b). The synthesis and turnover of sper- midine and spermine in mouse brain Neurochem. Res. 4, 269 275.
Shaw, G. G. and Pateman, A. J, (1973). The regional distri- bution of the polyamines spermidine and spermine in brain. J. Neurochem. 20, 1225 1230.
Siegel, L. G. and McClure, W. O. (1975). Putrescine: Effect on axoplasmic transport. Brain Res. 93, 543-547.
Singh, V. K. and Sung, S. C. (1972). Effect of spermidine on DNA-dependent RNA-polymerases from brain cell nu- clei. J. Neurochem. 19, 2885-2888.
Slotkin, T. A. (1979). Ornithine decarboxylase as a tool in developmental neurobiology. Life Sci. 24, 1623-1629.
Snyder, S. H., Kreuz, D. S., Medina, V. J. and Russell, D. H. (1970). Polyamine synthesis and turnover in rapidly growing tissue. Ann. N .Y Aead. Sci. 171, 749-772.
Snyder, S. H., Shaskan, E. G. and Harik, S. I. (1973). Polyamine disposition in the central nervous system. In: Polyamines in Normal and Neoplastic Growth. Russell, D. H. (Ed.) pp. 199 213. Raven Press, New York.
Sobue, K. and Nakajima, T. (1977). Metabolism of putres- cine in neuroblastoma and glioma cells during culture. J. Biochem. (Tokyo) 82, 1121-1126.
Sobue, K. and Nakajima, T. (1978). Changes in concen- trations of polyamines and ),-aminobutyric acid and their formation in chick embryo brain during develop- ment. J. Neurochem. 30, 277-279.
Sourkes, T. and Missala, K. (1978). Role of pyridoxine in the metabolism of putrescine in the rat. Can. J. Biochem. 56, 470-473.
Sturman, J. A. and Gaull, G. E. (1975). Polyamine metab- olism in the brain and liver of the developing monkey. J. Neurochem. 25, 267-272.
Sturman, J. A., Ingoglia, N. A. and Lindquist, T. D. (1976). Interconversion of putrescine, spermidine and spermine in goldfish and rat retina. Life Sei. 19, 719-724.
Tadano, T. (1974). Behavioral pharmacology of alkali metals. (3) Changes in polyamine levels in the brain and emotional behavior of aggressive mice originally isolated and returned to a group situation. Folia Pharmacol. Jap. 70, 457~464.
Tadano, T., Onoki, M. and Kisara, K. {1974). Behavioral pharmacology of alkali metals. 2. Changes of brain polyamine levels in mice with aggressive behavior and effects of LiCI. Folia Pharmacol. Jap. 70, 9 18.
Tashima, Y., Hasegawa, M. and Mizunuma, H. (1978). Activation of Na +, K+-adenosine triphosphatase by spermine. Biochem Biophys. Res. Commun. 82, 13-18.
Tsuji, M. and Nakajima, T. (1978). Studies on the forma- tion of v-aminobutyric acid from putrescine in rat organs and purification of its synthetic enzyme from rat intes- tine. J. Biochem. 83, 1407 1412.
Tsvetnenko, E. Z. and Shugalev, V. S. (1978). Polyamines spermine and spermidine in the brain and the liver of rats under hyperoxia. Byul. Eksp. Biol. Med. 85, 28 30.
van den Berg, C. J., van Kempen, G. M., Schade, J. P. and Veldstra, H. (1965). Levels and intracellular localization of glutamate decarboxylase and gamma-aminobutyrate transaminase and other enzymes during the development of the brain. J. Neurochem. 12, 863 869.
Wedgwood, M. A. and Wolstencroft, J. H. (1977). Effects of spermine and spermidine on single brain stem neurones. Neurapharmacology 16, 445 -446.
Williams-Ashman, H. G. and Canellakis, Z. (1979). Polyamines in mammalian biology and medicine. Per- spective in Biology and Medicine 22, 421 453.
Wood, J. (1975). The role of 7-aminobutyric acid in the mechanism of seizures Pro qress Neurobiol. 5, Part 1, 77 95.