po2, and pco2 changes in the mantle cavity of scrobicularia plana (bivalvia) under normal and stress...
TRANSCRIPT

Estuaritw and Coast& Marine Science (1979) 9, 499-507
p0, andpC0, Changes in the Mantle Cavity of Scrobicularia plana (Bivalvia) under Normal and Stress Conditions
H. B. Akberali and E. R. Trueman Department of Zoology, University of Manchester, Oxford Road, Manchester Mr3 9PL., U.K., England
Received 14 September 1978 and in revisedform 7 December 1978
Keywords: bivalves ; clams ; oxygen ; carbon dioxide ; hearts; respiration
Changes of heart rate, $&IS and pC01 in the mantle cavity water of Scrobicu- Zariu plana have been followed during activity cycles and quiescence whilst continually immersed in seawater and under certain experimental stress conditions.
During quiescence of 3 h duration, the oxygen content of the mantle cavity fell to about 50% of the active condition and carbon dioxide increased by about zoo%.
When subjected either to osmotic stress or aerial exposure, oxygen in the mantle cavity fell steadily until about 3 h had elapsed, when a sudden brief increase occurred. This is probably due to the slight opening of the valve margins allowing diffusion.
The level of CO* in the mantle cavity becomes stabilised about 4 h after valve closure. Although this may in part be due to outward diffusion, it appears likely that other mechanisms are also involved.
Introduction
When the valves of most bivalves are closed the heart-rate is reduced, to be followed by a rapid increase in rate when the shell opens. An essentially similar basic response commonly occurs in respect of various forms of respiratory stress (Trueman & Lowe, 1971; Coleman & Trueman, 1971; Bayne, 1971a; Taylor, 1976a; Brand, 1976). Shell closure has also been shown to result in a rapid decline in the oxygen content of the mantle cavity water, e.g. in MytiZus moon & Pritchard, 1970)~ Arctica (Taylor, 1976b) and Pecten (Brand & Roberts, ‘973).
Scrobicularia plana (da Costa), an estuarine bivalve often found abundantly in intertidal muds in seawater of reduced salinity (Green, 1968), exhibits an unusual pattern of short-term variation in heart rate closely related to the rate of pumping of water through the mantle cavity (Earll, 1975; Akberali, 1978). This is in contrast to small variations in heart activity observed in Ostrea edulis (Walne, 1972) or their total absence in Mytilus edulis (Thompson & Bayne, 1972). Only in species such as S. plana, which show increased sensitivity to pumping, is a direct relationship between heart rate and water flow through the mantle cavity likely to occur. The onset of pumping by Scrobiculariu marks the commencement of an active period with a rapid increase of both heart rate and amplitude whilst at the termination of pumping heart rate falls gradually as the bivalve enters a period of ventilatory pause. An active period
499 0302--3524/79/100499+09 802.00/o @ 1979 Academic Press Inc. (London) Ltd.

500 H. B. Akberali &3 E. R. Trueman
I I Minutes
Figure I. Recordings of the heartbeat of Scrobiculuria plum during A, activity; B, ventilatory pause; and C, quiescence. Arrows indicate withdrawal of sample of mantle cavity water by insertion of a hypodermic needle through a rubber dia- phragm.
followed by a ventilatory pause has been termed an activity cycle (Akberali, 1978). The active period is commonly of 15 min-1 h duration, the ventilatory pause 15-45 min. (Akberali, 1978). The ventilatory pause may, however, be extended for up to 4--12 h, into what has been termed a period of quiescence, when the valves close and all movements cease. Bradycardia then occurs, occasionally resulting in complete cessation of the heart beat (Earll, 1975 ;
Akberali, 1978). All these changes may occur during constant immersion of the bivalve in sea water.
Recent studies of Scrobicularia under various stress situations indicate some of its beha- vioural and physiological responses (Akberali, 1977; Akberali et al., 1977). In the present study pOs and $0, levels in the mantle cavity water have been monitored under normal and short term experimental stress situations and an attempt is made to relate these observa- tions to the behaviour of Scrobicularia.
Materials and methods
Scrobicularia plana, shell length 4 cm, were collected from Morecambe Bay and maintained in Manchester University in tanks of well aerated seawater (100~/~=31%,,S) at IO%, which was also the temperature used in all experiments. The clams were immediately, and subse-

~0, and pCOa in mantle cavity of Scrobicularia 501
* . * +
Figure 2. Mantle cavitypO,, pCo, and heart rate (H.R.) of individual Scrobicularia plana during active periods (0) and ventilatory pauses (0) when constantly immersed in sea water. Each point for the 14 animals during active periods is the mean of 6 determinations and each point for the 5 animals during ventilator? pauses is the mean of 4 determinations. Bars indicate S.E., points for the same ammal are joined by broken lines.
quently regularly fed with Phaeoductylum cultures. Heart rate was monitored by use of the impedance technique (Trueman et al., 1973).
For the study of oxygen and carbon dioxide levels during spontaneous activity cycles whilst continually immersed in seawater, one valve of each experimental animal was fixed to the base of a I 1 crystallising dish while a hole of 1-2 mm diameter was carefully drilled in the upper valve and covered with a thin rubber diaphragm. This allowed samples of 0.1~0.2 ml to be withdrawn from the mantle cavity, by means of a hypodermic needle, for determination of ~0, and $0,. The normal pattern of behaviour was little affected by this procedure as ohservcd from recordings of the heart rate (Figure I) and examination of the tissues aftcl completion of the experiment confirmed that the fluid withdrawn was from the mantle cavity without damage to gills or foot. Only one sample was taken from each individual at any one time during each active period or ventilatory pause so as to minimise the change in volume in the mantle cavity.
In a further series of experiments mantle cavity water was withdrawn by the insertion of a hypodermic needle between the ventral valve margins. Each animal was then discarded from the experiment. Employing this technique a large number of animals was used in the following cxperimcntal regimes: transference from 100% seawater, (i) to well aerated 20’):~

502 H. B. Akberali &Y E. R. Trueman
seawater; (ii) to 20”/~ seawater for 24 h and then transference back to 100’):) seawater; (iii) to air with free valves and with valves forcibly closed with rubber bands; (iv) to an atmosphere of nitrogen.
pC0, and ~0, determinations were made on mantle cavity water using a Radiometer PHM 73 PH/Blood Gas Monitor with GMA z Precision Gas Supply coupled to BMS3 MK2 Blood Microsystem, with a thermostatic water bath temperature of 37°C. Samples were injected into the Blood Gas Monitor immediately following withdrawal from the animal. It would have been appropriate to have used microelectrodes implanted in the mantle cavity of Scrobicularia to monitor oxygen and carbon dioxide levels continuously but sufficiently small sized electrodes were not available.
Results
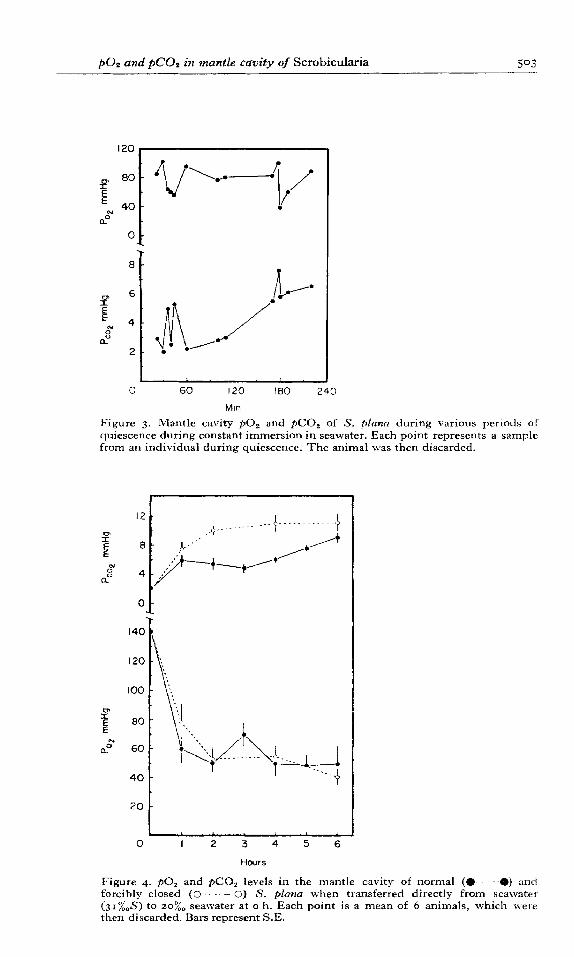
Some 14 Scrobicularia, prepared for intermittent samples to be withdrawn from the mantle cavity via a rubber diaphragm, were kept continually immersed in seawater of normal salinity (3 I%$?). During separate active periods 6 water samples were withdrawn from each clam for determination of ~0, and pC0,; and during separate ventilatory pauses 4 samples were similarly withdrawn but from only 5 of these clams. The samples were withdrawn during activity cycles at random intervals at least 5 min after commencement of an active period and 10-15 min after the onset of a ventilatory pause. Heart rate was monitored continuously and the results of this experiment are conveniently summarised in Figure 2. Although the results show individual variation particularly in respect of the heart rate, there is a significant reduction in heart rate and ~0, between active period and ventilatory pauses (16-18+ 10~ 13 beats/min; 140-160-t SO-I IO mm Hg respectively). During activity thepC0, rarely exceeded 2 mm Hg but on cessation of pumping never fell below this value. The techniques of taking samples from the mantle cavity at a given time after the commencement of an active or inactive period does not allow continuous monitoring of the ~0, and pC0, levels. With- drawal of a sample disturbs the course of an activity cycle sufficiently (Figure I) to prevent further samples being taken during that cycle. Accordingly a subsequent cycle had to be awaited before a further sample could be obtained. Continuous monitoring of the mantle cavity water by implantation of electrodes would appear to be the only feasible method of following p0 s andpC0, during these activity cycles and we have not been able to accomplish this. In its present form the experiment does not show whether thereis a threshold level of oxygen or carbon dioxide which might cause pumping to recommence in individual clams. However, it is significant that during quiescence lower levels ofp0 s and higher values of&O, are attained than during temporary cessation of pumping and that these levels may be maintained for more than IOO min (Figure 3). This suggests that the absolute level of oxygen and carbon dioxide are not critical factors in the control of pumping activity. During various periods of quiescence when the valves are closed and the siphons withdrawn, the ~0, is about 88 mm Hg and does not gradually decrease, whilst $0, tends to increase with the duration of quiescence. However, both of these curves exhibit some rather characteristic fluctuations which are conveniently discussed below.
Transference of Scrobicularia from 100% to 207~ seawater normally results in valve closure for some days and marked bradycardia, although some valve movement has been detected after several hours (Akberali, 1978). During a period of 6 h after transference, ~0, decreases and $0, gradually increases (Figure 4). However, after about 3 h there is an increase in oxygen tension and a decrease in carbon dioxide. This may be interpreted as the result of slight gaping of the valves, possibly to test the media, since in clams which have
pOa and pCOr in mantle cavity of Scrobicularia 503 ___
0 60 120 180 240
Mln
Figure 3. Mantle cavity ~0, and pC0, of S. plum during various periods of quiescence during constant immersion in seawater. Each point represents a sample from an individual during quiescence. The animal was then discarded.
8
12 i
_.....__.._.. 0 .-.......... .~ +
,&.:
,.‘1--+,,,*’
*’ /+
4 / ,/
0
60
I 0 I 2 3 4 5 6
Hours
Figure 4. ~0, and pC0, levels in the mantle cavity of normal (e-----O) and forcibly closed (0 - - - 0) S. plana when transferred directly from seawater (31”/wS) to 20%~ seawater at o h. Each point is a mean of 5 animals, which were then discarded. Bars represent S.E.

504 H. B. Akberali C9 E. R. Trueman
been forcibly closed no such changes have been observed (Figure 4). Indeed the ~0, of the latter is maintained at between 40 and 55 mm Hg whilst the pC0, increases rapidly until it appears to stabilise after 4 h. After Scrobiculuria was maintained in 20% seawater for q. h the course of this last experiment was effectively reversed by transfer of Scrobiculariu into normal seawater (Figure 5). For the first IO min there was little response with the heart rate remaining at 5-7 beats/min and $0, at about 12 mm Hg. The ~0, somewhat surprisingl! fell sharply initially but attained more stable, near normal, levels within an hour. These
12
2
120
f 100
2 80
6C
f\ d t’ , -,A-t’i v \ t
0 40 80 120
Mtn
Figure 5. PO,, @CO, and heart rate of S. plana transferred from seawater to 201~~
seawater for 24 h when they were replaced in xoo’/J seawater at o min. Each point for ~0, and pC0, is mean of 7 animals, which were then discarded. Bars represent S.E. Heart rate (H.R.) presented from one typical animal.
observations suggest an initial shock reaction to the osmotic change, the animal immediately
closing its valves and after IO min slowly gaping as evidenced by the increase in heart rate. Recovery from exposure to low salinity is thus relatively rapid.
A further experiment exposed Scrobiculariu to air both with the valves free and forcibly closed, and to an atmosphere of nitrogen, with the valves free, all at a constant temperature of IOT (Figure 6). Under all these conditions, the mantle cavity ~0, declined and the&O,

pea and pCOe in mantle cavity of Scrobicularia 50.5
rose sharply during the first hour. The oxygen content of clams both forcibly closed and in nitrogen is essentially similar. They differ from unrestrained clams in air which exhibit a peak of ~0, after 3 hours and anincreasing oxygen content at the end of the experiment. This peak may be brought about by Scrobiculuriu opening its valve margins slightly allowing inward diffusion of oxygen. The peak in ~0, is remarkably similar to that shown after 3 h when Scrobiculuriu are transferred from seawater to a lower salinity. This suggests that the clams may test and react to their habitat 2 or 3 h after application of stress. The carbon. dioxide content of clams in all three conditions of emersion is similar, although higher values for those forcibly closed may indicate some outward diffusion both in air and in an atmosphere of nitrogen. The heartbeat of Scrobiculuriu is very irregular in air, with much individual variation (Earll, 1975).
0 123456
Hours
Figure 6. p02 and pCOe of mantle cavity water of S. plana when placed (at o h) in air (+--@), in atmosphere of nitrogen (m---w) or in air with valves forcibly closed ( O- - - 0). Each point a mean of 7 animals. Bars represent S.E.
Discussion
In Scrobiculuria plana the heart rate has been shown (Earll, 1975) to be closely related to the pumping of water through the mantle cavity during activity cycles. During ventilatory pauses, the heart rate and oxygen content of the contained water fell, whilst the amount of carbon dioxide increased. A partial pressure of oxygen of not less than 140 mm Hg is normal during pumping but this fell to about IOO mm Hg during inactivity over a period of IO-IS
min.

506 H. B. Akberali & E. R. Trueman
The ~0, does not fall much below IOO mm Hg during activity cycles but, during more prolonged quiescence, short periods of much lower oxygen levels occur in the mantle cavity, although about 80 mmHg is maintained for most of the quiescent period (Figure 3). Possibly at about this level the animal changes to anaerobic metabolism even although the mantle cavity water is not completery anoxic as occurs in some other invertebrates (Mangum & Van Winkle, 1973; Moon & Pritchard, 1970). It is only when the valves are held tightly closed either forcibly or by the shock of transfer to low salinities that substantially lower levels of oxygen are obtained (Figure 4). This suggests that in all other conditions some inward diffusion of oxygen may occur. In contrast to the levels of oxygen the amount of carbon dioxide gradually increases during quiescence. Sharp peaks of increased carbon dioxide also occur and correspond to low oxygen levels; note that one peak occurs 3 h after the commencement of quiescence (Figure 3). The restoration of the previous levels aftel these peaks suggest that the animals may gain oxygen and lose carbon dioxide by diflusion, probably between the ventral mantle margins. This is probably also the explanation of the peak in oxygen content of the mantle cavity 3 h after the transference from IOO(~~ to 20”,,
seawater (Figure 4) or after emersion (Figure 5). When the valves are forcibly closed this feature does not occur in either experiment. In these experiments we have not monitored valve movements, but previous observations (Akberali, 1978) indicate that Scrobicularia C;U~ gape very slightly and may allow some gaseous diffusion even although no gaping movement of the valves can be detected. Slight gaping of the valves for respiration in air is a feature which occurs in a number of bivalves, e.g. Modiolus demissus (Lent, 1968), Isognomon ahztus (Truc- man& Lowe, 1971) and Cardium edule (Boyden, 1972).
The features occurring in Scrobicularia immediately following quiescence can be nwst
readily reproduced experimentally by imposing stress and subsequently returning to normal conditions (Figure 5). In most bivalves, including S. plarza (Akberali, 1978) a common feature of recovery is an overshoot in respect of heart rate and amplitude (Bayne, 197Ia, b; Taylor, r976a, 6) which is considered due to the higher demands placed upon circulator! and respiratory systems (Brand & Roberts, 1973). Another feature observed was the repeated adduction and relaxation of the valves probably to effect hyperventilation of the mantle cavity. Although observed in Scrobicularia it does not show as a series of abrupt changes in the curves for ~0, and $0, (Figure 5) since these curves are from a number of animals, each discarded after sampling, so reflecting a net effect. Nevertheless 2 steps on the oxygen curve suggest a somewhat intermittent, rather than gradual, recovery compatible with hyperventilation of the mantle cavity. Valve closure of ScrobiczzZaria during short term stress conditions or natural quiescence results in a drop in the pH of the mantle cavity (Akberali PC Hodgson, in prep.). With the onset of activity, rapid valve adductions occur and are associated with a drop in pH after a short time lag suggesting a release of acid metabolites from the tissues into the mantle cavity. With the opening of the valves and commencement of ciliary pumping the pH of the mantle cavity water returns to normal levels.
In all experimental situations involving valve closure the carbon dioxide content of the mantle cavity shows an initial rapid increase and after about 3 h stabilises. Thus carbon dioxide either must diffuse out over a certain threshold level or it is being buffered. Calcium is mobilised from the shell during anaerobic respiration, presumably to buffer the by- products of this metabolism and to prevent a pH drop (Akberali et al., 1977) but in the production of free calcium ions from the shell carbon dioxide is also liberated (Wijsman, 1975) which in turn would decrease the pH unless the animal could deal with it in another manner. Mobilisation of calcium does not apparently show an increase of carbon dioxide in the mantle cavity in the experiments described here. The greater amount of carbon dioxide in

~0% and pCOz in mantle cavity of Scrobicularia 507
the mantle cavity of animals which have been forcibly closed for G h (Figures 4 and 5) suggests that some diffusion may normally occur. But even in the latter animals a stable condition obtains after 3 h. This suggests that further attention should be given to the problems of stabilisation of the carbon dioxide level during anaerobiosis.
Acknowledgements
This work was carried out with the support of N.E.R.C. Research Grant GR3/3436.
References
Akberali, H. B. 1977 Responses of Scrobicularia plana to varying salinities. Ph.D. Thesis, University of Manchester, 151 pp.
Akberali. H. B.. Marriot. K. R. M. & Trueman. E. R. 1977 Calcium utilisation during anaerobiosis ind&ed by’osmotic shock in a bivalve moll&c. Noturk; &z&m 266,852-853. -
Akberali, H. B. 1978 Behaviour of Scrobiculuriupluna (da Costa) in water of various salinities..rozr~na( OJ Experimental Marine Biology and Ecology 34,s 1-63.
Bayne, B. L. Ig7IU Oxygen consumption by three species of lamellibranch mollusc in declining ambient oxygen tension. Comparative Biochemistry and Physiology BOA, 955-970.
Bayne, B. L. 1971b Ventilation, the heart rate and oxygen uptake by Mytilus edulis (I,.) in declining oxygen tension. Comparative Biochemistry and Physiology qoA, 1065-1085.
Boyden, C. R. 1972 The behaviour, survival and respiration of the cockles Cerastoderma (Cardium) edule and C. glaucum in air. Journal qf the Marine Biological Association oj the United Kingdom 52, 661-680.
Brand, A. R. 1976 Heart action of the freshwater bivalve Anodonta nnatina during activity. rozrrnnl of Experimental Biology 65,685-698.
Brand, A. R. & Robert, D. 1973 The cardiac responses of the scallop Pecten mnximus (L.) to respiratory stress.yournal of Experimental Marine Biology and Ecology 13,29--43,
Coleman, N. & Trueman, E. R. 1971 The effect of aerial exposure on the activity of the mussels Mytilus edulis (L.) and Modiolus modiolus (L.). J ournal of Experimental Marine Biology and Ecology 7, 295-304.
Earll, R. 1975 Temporal variation in the heart activity of Scrobicularia plana (da Costa) in constant and tidal conditions. Journal of Experimental Marine Biology and Ecology 19, 257-274.
(ireen, J. 1968 The Biology of Estuarine Animals. Sidgwick & Jackson, London, 401 pp. Lent, C. M. 1968 Air-gaping by the ribbed mussel, Modiolus demissus: (Dillwyn): effects and adaptivr
significance. Biological Bulletin of Marine Biology Laboratory, Woods Hole 134,60-73. AIangum, C. & Van Winkle, W. 1973 Responses of aquatic invertebrates to declining oxygen conditions.
American Zoologist 13,529-541. >Ioon, T. W. & Pritchard, A. W. 1970 Metabolic adaptations in vertically-separated populations of
Mytilus californianus Conrad.Journal of Experimental Marine Biology and Ecology 5, 35-46. Taylor, A. C. 1976~ The cardiac responses to shell opening and closure in the bivalve Arctica isl~~ndic~~
(L.).Journal of Experimental Biology 64,751-759. Taylor, A. C. 19766 Burrowing behaviour and anaerobiosis in the bivalve Arctica islandica (I>.). j’ournnl
of the Marine Biological Association of the United Kingdom 56, 95-109. ‘Thompson, R. J. & Bayne, B. L. 1972 Active metabolism associated with feeding in the mussel 1vJtti/zc.?
edulis L.Journal of Experimental Marine Biology and Ecology 9,111-124. Trueman, E. R. & Lowe, G. A. 1971 The effect of temperature and littoral exposure on the heart rate
of a bivalve mollusc, Isognomon alatus, in tropical conditions. Comparative Biochemistry rrnti Physiology 38A, 555-564.
l’rueman, E. R., Blatchford, J. G., Jones, H. D. & Lowe, G. 1973 Recordings of heart rate and activit! of molluscs in their natural habitat. Mulucologia 14,377-383.
1Valne, P. R. 1972 The influence of current speed, body size and water temperature on the filtration rate of five species of bivalves. journal of the Marine Biological Association of the United Kirqdom 5% 345-374.
Wijsman, T. C. M. 1975 pH fluctuations in Mytilzts edulis L. in relation to shell movements undel aerobic and anaerobic conditions. Proceedings of the Ninth European Marine Biolozy Symposiltnr (Barnes, H., ed.). 139-149.