plant molecular systematics spring 2011. “problems” with morphological data… convergence and...
TRANSCRIPT

Plant Molecular Systematics
Spring 2011

“Problems” with morphologicaldata…
• Convergence and parallelisms
• Reduction and character loss
• Phenotypic vs. genotypic differences
• Evaluation of homology
• Misinterpretation of change or polarity
• Limitation on number of characters
• Phenotypic plasticity

Always searching for new types of characters…
Is molecular data intrinsically better than morphological data?

Central Dogma
Lipid pigments: chlorophyll lycopenes xanthophylls carotene
Phenolics: flavonols flavones tannins anthocyanins
Iridoid compounds
Alkaloids(N-containing)e.g. nicotine caffeine morphine betalains
Secondary Metabolites
Terpenes

Development of Molecular (Chemical) Systematic Methods
“Chemosystematics”
• Early methods relied on chromatography to separate complex mixtures of secondary metabolites, detect them, and then compare between taxa “spot botanists” – very phenetic
• Better separation and identification methods developed (HPLC, MS) – used pathway stages as cladistic characters - phytochemistry
• Move away from secondary metabolites to proteins• Early protein studies used immunological reactions• Development of improved electrophoretic methods – permitted
direct protein comparisons between taxa• Comparison of seed storage proteins• Development of allozyme techniques – direct estimates of genetic
relationships based on allele frequency

Molecular (DNA) Systematics
• Next step was to examine DNA directly through examination and comparison of restriction fragments (RFLP bands)
• Technology evolved to make it feasible to sequence DNA directly
• Initially limited to single genes or non-coding regions
• Now feasible to sequence large numbers of genes or regions or even whole genomes relatively quickly

Molecular Systematics- Can obtain phylogenetically informative characters from any genome of the organism- Assumes that genomes accumulate molecular changes by lineage, as morphological characters do- Possibly greater assurance of homology with molecular data (less likely to misinterpret characters) but homoplasy happens!- Principal advantages are the much greater number of molecular characters available & greater comparability across lineages

Genomes of the Plant Cell
Nuclear
Plastid
Mitochondrial

Table 5.1 from the text

Selection of DNA region to compare:
• Should be present in all taxa to be compared
• Must have some knowledge of the gene or other genomic region to develop primers, etc.
• Evolutionary rate of sequence changes must be appropriate to the taxonomic level(s) being investigated; “slow” genes versus “fast” genes
• It is desirable that sequences can be readily aligned
• The biology of the gene (or other DNA sequence) must be understood to assure homology

Genes frequently used for phylogenetic studies of plants:
• Mitochondrial genome – uniparentally inherited, but genes evolve very slowly and structural rearrangements happen very frequently, so generally not useful in studying relationships
• Plastid genome – uniparentally inherited- rbcL – ribulose-bisphosphate carboxylase large subunit- ndhF – NADH dehydrogenase subunit F- atpB – ATP synthetase subunit B- matK – maturase subunit K- rpl16 intron – ribosomal protein L16 intron
• Nuclear genome – biparentally inherited- ITS region – internal transcribed spacers ITS1 and ITS2- 18S, 26S ribosomal nuclear DNA repeat- adh – alcohol dehydrogenase

Plastid Genome
-Circular, derived from endosymbio- sis of cyanobacteria
-Three zones: LSC (large single copy region) SSC (small single copy region) IR (inverted repeats)
- Genes related to PS and protein synthesis Marchantia polymorpha - liverwort

Figure 5.1 from the text

The Polymerase Chain Reaction (PCR)
Figure 5.2 from the text

Automated Sequencing
Scanning of gel to detect fluorescently-labeled DNAs; data fed directly to computer.




How do we analyze molecular variation?
- DNA nucleotide sequences (point
mutations) - - Structural rearrangements
-insertions and deletions (indels)-inversions

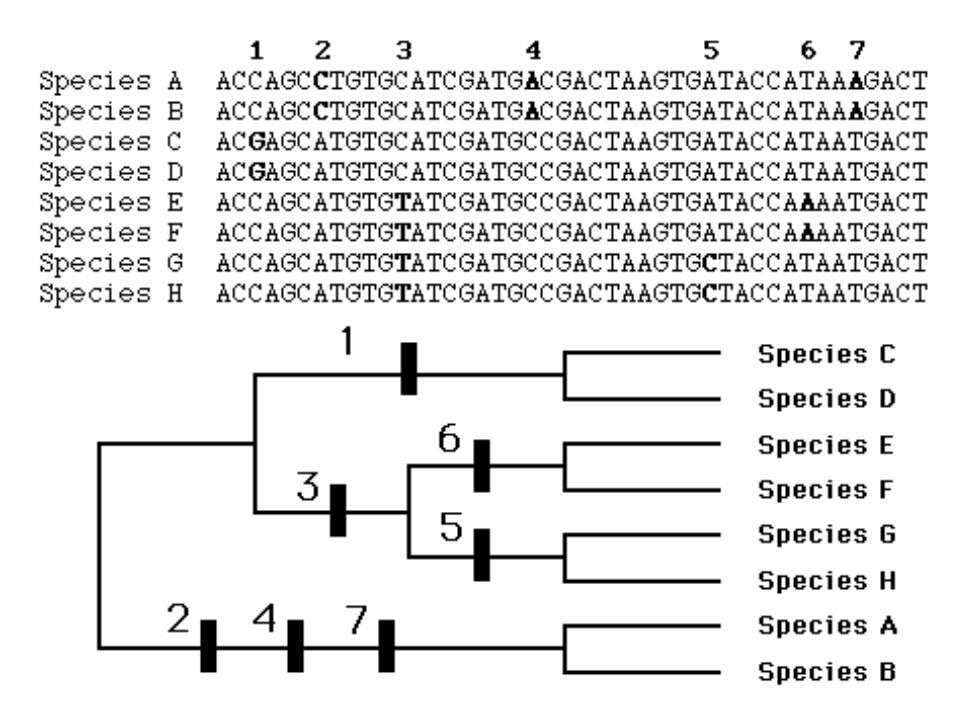
Partial sequence of rbcL (plastid gene coding for Rubisco) in Poaceae
Figure 5.3 from the text

Insertion-Deletion Events
- Can occur as single nucleotide gains or losses or as lengths of 2-many base pairs- Can also be “chunks” of DNA (i.e., losses of introns)

A molecular synapomorphy for Subfamily Cactoideae (Cactaceae) – deletion of the plastid rpoC1 intron…
(Wallace & Cota, Current Genetics, 1995)

Cactaceae: trnL Intron Deletions

North American Clades
South American Clades
Pachycereeae
Corryocactus
“Browningieae I”*
“Browningieae II”*
Cereeae
Trichocereeae
Leptocereeae
Hylocereeae
Shared Deletion 2
- 268 bp
trnL intron deletions – Columnar Cacti
(*Tribe Browningieae polyphyletic)

23 kb inversion in all Asteraceae except for members of Tribe Barnadesieae (now Subfamily Barnadesioideae)
Chloroplast DNA Inversion

Comparative DNA Sequencing
• Obtain DNA samples from representative organisms (try to represent morphological diversity) and outgroups
• Identify DNA region(s) for comparison
• Use PCR to amplify targeted region
• Carry out sequencing reactions
• Run sequencing procedures (automated)
• Align sequences
• Use aligned sequences for phylogenetic analysis (various programs using various algorithms)
• Evaluate data in context of taxonomy and morphology

Panicgrasses(Panicoideae)
Rices(Ehrhartoideae)
Bluegrasses(Pooideae)
Bamboos(Bambusoideae)
Puelioideae
PharoideaeAnomochlooideae
Needlegrasses(Aristidoideae)
Lovegrasses(Chloridoideae)
Micrairoideae
Reeds(Arundinoideae)
Oatgrasses(Danthonioideae)
Major radiationin Oligocene-Miocene epochsinto open habitats
Origin of grassesca. 70-80 myain forests
+
Stamensreduced to 3

Phylogeny ofPooideae(multipledata sets)
Figure 5.5. from the text

Figure 5.7 from the text

Figure 5.4 from the text