plant-derived chemicals as tick repellents - diva portal172830/fulltext01.pdf · plant-derived...
TRANSCRIPT

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2008
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 574
Plant-Derived Chemicals as TickRepellents
SAMIRA SADEK GARBOUI
ISSN 1651-6214ISBN 978-91-554-7337-2urn:nbn:se:uu:diva-9368

���������� �������� �� ������ �������� � �� �������� ������� � ���������������������� ������� � �������� �������� !� "�#$%& '!� ������� (�������������� %� &))* �� )+,)) �� �-� ������ � ���� � .-����-�/ 0-� �������� 1��� ��������� � �����-/
��������
"���2 3������ "/ &))*/ .���#������� 4-������� �� 0��2 5��������/ 6��� ����������� ���������/ ������� ��� � ���� ����� � � ������� ���� ������� �� �� ������� ���� �� ��� � ������ %$7/ ! ��/ ������/ 8"9� +$*#+ #%%7#$''$#&/
��� � ������� �� �-� ��� ����� � :��� ��������� �� 0��2#��� ����-������ � �����/5�������� ������ � ��������� ���� � ������� ������ ���2 ����� �� �� �-������ ��������������� � ���2#��� ��������/8 �������� ������ ������ � ���- ������� 1��- ����6 �������; �� ��������� �����
��������� � 1��- �-� �������� ��� � ������ �������� �')<�� ��������� ����������� � �� ')<�� � ������� ���� � �� � �� ')<�� ����� ���� � �� � )<�� ��������� ������ �������� ���������� !���� ��� �� ��� � �&)<� �� ���-�� =������ ��>� �)/%<� /)<� &/)< �� %/)<� �-1�� ���� �������� �������� ������ �" ������� ���-�/8 � ���2#������� 1���� � ����#������ "1���� 1� ������ �� �����?��� �����������
���-���� �-� ������� ���#���2 �������� �������� � ����6 �������; ����#�� �1��������� � ����6 �������; ������ 59*! �� ����������� ��������� ����� ���������� -������ �-� �������� �� � �" �������� �� �-��� ��������� � �>/ ���- ��������1�� �������� � ����� �� ������� ���������� � �& ���-� 1-��- 1��� �-� ������ �����������/ ����-�� ���2 ������ �-� ���-� 1��� ������� �� )#� �������� �� ������������������� ���1�� ������� ���-� �� �-� �������� ���� �� ��� ���1�� ��������/@������ ������ ��� ����- �� ����� ������ � " ���� � �� �� �-� �������� ��� �
" ���� � �� ���� :�� �� 3���� 9������ �� �" � ������ �" ��������� �� !" �� ��� ����� :����� 1��� �������� �� ���� �-��� ����#�������� �".��� �� �-� ���������1��� ��������� �� ��� �-��������-� # ���� ����������� �34#�"�/ 01 ������A�������� -�������� � �-� " ���� � �� �� 1��� B#�����-����� �� -������/0-��� 1��� ������� �� ������ �� ����-����� � ���� ��������� ���2 �������������� 1��- �1�� ���������/ 8 ��� �-��� ��� ��� :���� ����� �-� ��� �������-�������� 1��� C#����� ������� B#����� ������� C#�-��������� 7#������B#�-�������� �� D#�������/0-� �������� ���� ������� ������ ������ �������� 21 � -��� ������������
����������� ��E� ����� �������� ���������/
# �$���% 8���� ������� ������ �������� ���� ���2 ��������� "1���
����� ��� & '�����( � ���� �� � )������( ' ���� ��� ���� �����( ���� ����*���( +����" ,-�( ������� ���� �����( �)./0123 �������( �$ � �
F "����� "���2 3����� &))*
8""� !% #!& 78"9� +$*#+ #%%7#$''$#&��,�,��,��,����#+'!* �-���,EE��/2�/��E������G��H��,�,��,��,����#+'!*�

Glory be to Allah (God) we have knowledge except what Allah has taught us. Verily, Allah is the All-Knower and the All-Wise. The Nobel Quraan (Surah 2, versus 32)
In memory of my Father Elsadek Garboui To my mother Fathia Ahmed To my Family Gumaa, Maram, Rouaa and Mossa

Cover photo: Biopix.dk

List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals:
I Jaenson, T. G. T., Garboui, S. and Pålsson, K. 2006. Repellency
of oils of lemon eucalyptus, geranium, and lavender and the mosquito repellent MyggA Natural to Ixodes ricinus (Acari: Ixodidae) in the laboratory and field. J. Med. Entomol. 43:731-736.
II Garboui, S. S., Jaenson, T. G. T. and Pålsson, K. 2006. Repel-
lency of MyggA® Natural spray (para-menthane 3, 8-diol) and RB86 (neem oil) against the tick Ixodes ricinus (Acari: Ixodidae) in the field in east-central Sweden. Exp. Appl. Acarol. 40:271-277.
III Garboui, S. S., Jaenson, T. G. T., Borg-Karlson, A-K., and Påls-
son, K. 2007. Repellency of methyl jasmonate to Ixodes ricinus nymphs (Acari: Ixodidae). Exp. Appl. Acarol.42:209-215.
IV Ashitani, T., Garboui, S. S., Pålsson, K., Vongsombath, Ch.,
Jaenson, T. G. T., and Borg-Karlson, A-K. 2008. Hyptis suaveolens a source for arthropod repellent compounds. Repel-lency of sesquiterpene oxides and sulfides to Ixodes ricinus. Submitted.
V Garboui, S. S., Borg-Karlson, A-K., and Pålsson, K. 2008. Tick
repellent properties of three Libyan plants. Submitted. Papers I, II and III are reprinted with the kind permission of the publisher.


Contents
Introduction.....................................................................................................9 Ticks (Acari: Ixodidae) ..............................................................................9 Tick Control .............................................................................................11
Biological control ................................................................................11 Chemical Control.................................................................................12 Personal Protection ..............................................................................16
Plants used in this study ...........................................................................17 Aims .........................................................................................................20
Materials and Methods..................................................................................21 Study areas ...............................................................................................21 Sampling method......................................................................................21 Plant oils and arthropod repellents ...........................................................22 Laboratory bioassays................................................................................23
Repellency test on I. ricinus nymphs...................................................23 Steam distillation (IV and V)...............................................................24 Collection of volatile substances from the leaves and oils (IV and V)..........24 Identification of compounds (IV and V)..............................................25
Field experiments .....................................................................................26 Statistical analyses....................................................................................27
Results and Discussion .................................................................................28 Laboratory bioassays................................................................................28
Repellency test on I. ricinus nymphs...................................................28 Field experiments .....................................................................................41 Concluding remarks .................................................................................48
Sammanfattning (Swedish summary) ...........................................................49
Acknowledgements.......................................................................................52
References.....................................................................................................54

Abbreviations
AI3-37220 1-(3-cyclohexen-1-yl-carbonyl)-2-methylpiperidine
DEET N, N-diethyl-3-methylbenzamide GC-MS Gas Chromatography-Mass
Spectrometry IR3535 Insect Repellent (3-(N-n-butyl-N-
acetyl)-aminopropionic acid ethyl ester).
KBR3023
LB
1-piperidinecarboxylic acid, 2-(2-hydroxyethyl)-, 1-ethylpropylester Lyme borreliosis
LD Lyme disease MJ Methyljasmonate MS Mass Spectrometry NIST National Institute of Standards and
Technology PMD Para-menthane-3, 8-diol RB86 Neem oil 70% SPME Solid Phase Micro Extraction TBE Tick-borne encephalitis

9
Introduction
Ticks have recently attracted great attention and are currently considered to be second only to mosquitoes as vectors of human infectious diseases throughout the world. Each vector tick species prefers particular optimal environmental conditions and biotopes: these preferences determine the geo-graphic distribution of ticks and, consequently, the risk areas for tick-borne diseases. In non-tropical areas the main vector-borne disease is Lyme disease (LD) caused by Borrelia burgdorferi (Johnson) sensu lato and transmitted by Ixodes spp. Lyme disease is the most commonly reported vector-borne hu-man illness in the northern temperate hemisphere. Personal protection with repellents against blood-sucking arthropods is one strategy to prevent infec-tion with vector-borne diseases. Increasingly efforts have been made to de-velop alternatives to replace synthetic chemical repellents including the ex-traction and evaluation for repellent activity of plant products. It is well known that plants produce a wide range of compounds that act as a defense against herbivory. The development and use of locally available plant prod-ucts showing repellent activity is thus an alternative strategy for prevention of tick-borne diseases. Several studies have been carried out to evaluate the repellent action of compounds from various parts of plants. Some have pro-vided good protection against many blood-sucking arthropods (Davis 1985, Gupta and Rutledge 1989, Malonza et al. 1992, Ansari and Razdan 1994, Barnard 1999, Barnard 2000, Jaenson et al. 2003). Repellents are relatively inexpensive and a practical means of protection against nuisance and disease vector arthropods, especially when other control measures are not feasible.
Ticks (Acari: Ixodidae) Ixodidae, or hard ticks, are large blood-sucking Acari. The family Ixodidae is comprised of 13 genera of which 7 contain species of medical importance (Kettle 1995). The largest genus is Ixodes, which is estimated to have more than 200 species world wide (Kettle 1995). The developmental cycle of ticks includes four stages: egg, larva, nymph and adult. There are no sexually differentiating morphological characters until the adult stage is reached. Ixodes are usually present in humid environments, quest freely on the vege-tation or in the host’s nest or burrows and feed only once in each stage. Ticks are obligate ectoparasites of terrestrial vertebrates and are well known

10
as important vectors transmitting pathogenic micro-organisms to animals and humans in many parts of the world (Sonenshine 1993). Ticks affect their hosts by transmitting disease organisms, causing paralysis, reducing their growth rates and milk production, and by injuries leading to secondary infec-tions (Kettle 1995). In Sweden, >10 species of Ixodidae (hard ticks) occur on a regular basis (Jaenson et al. 1994). The most common and the medically most important tick in Europe is Ixodes ricinus L. This tick species has a very wide geographical distribution throughout most of Europe, including the British Isles. Stable populations are also present in northern Africa (Gray 1991). I. ricinus is involved in the transmission of spirochetes of B. burgdorferi s. l. which causes Lyme borreliosis (LB) or Lyme disease (Sonenshine 1993), the most common tick-borne human disease in Europe and the United States. The highest reported frequencies of the disease are in middle Europe, particularly in Germany, Austria, Slovenia, and Sweden (Dennis et al. 2002). In Sweden alone, each year an estimated 10,000 people become infected with Lyme borreliosis spirochetes (SIIDC 2007). B. burgdorferi is a com-plex of >10 genospecies of spirochetal bacteria. Five Borrelia genospecies have been found associated with I. ricinus (Piesman and Gern 2004). The genospecies responsible for human LB are B. burgdorferi sensu stricto (s. s.), B. garinii (Baranton) and B. afzelii (Canica). Each of these is associated with different tick/host systems and different clinical manifestations of the disease (Baranton et al. 1992, Gern and Falco 2000). LB is a multi-systemic syndrome, affecting mainly the skin (erythema migrans, rash), joints, nerv-ous system, and heart (Baranton et al. 1992, Gern and Falco 2000). Ery-thema migrans is the cardinal sign of the disease caused by migrating spiro-chetes and appears on the site of a tick bite some days or weeks after infec-tion (Gern and Falco 2000). Some strains of Borrelia do not have any affect on the skin: flu-like symptoms and excessive fatigue are often the first no-ticeable indications of infection (ILADS 2006). In Sweden, infection is typi-cally caused by nymphal ticks wich is the most abundant of the infective stages since adult femal ticks are less abundant and also probably more eas-ily detected and removed (Jaenson 1991).
Several species of small and medium-sized mammals and ground-
feeding birds serve as reservoirs for the bacteria and are also the main hosts for I. ricinus larvae and nymph (Baranton et al. 1992, Gern and Falco 2000).
The viral disease tick-borne encephalitis (TBE) is widely distributed in Europe. In Sweden there are about 150-200 pepole annually become seri-ously ill due to infection with the virus causing TBE (SIIDC 2007) . The main reservoirs for the TBE virus are small mammals and the distribution is restricted to certain areas around Lake Mälaren and to some islands in the

11
Baltic Sea. However, the recent changes in the climate of Sweden may affect this distribution (Lindgren et al. 2000).
Other bacterial diseases transmitted by I. ricinus are known from Sweden.
A spotted fever group Rickettsia was reported as being indigenous to Swe-den (Nilsson et al. 1997) and has been isolated from I. ricinus in Sweden (Nilsson et al. 1999). Moreover I. ricinus in Sweden is a vector of anaplas-mosis (Anaplasma phagocytophilum Foggie.), Q-fever (Coxiella burnetii Derrick.), tularaemia (Francisella tularensis Olsufjev.) and babesiosis (Babesia divergens (M'Fadyean & Stockman) and B. microti Brown) to man and farm animals (Jaenson 1999).
Tick Control Several strategies, including the use of biological control and chemical con-trol have been shown to reduce the number of vector-ticks in areas where Lyme disease and other tick-borne diseases are endemic. The cooperative engagement of public health information on personal protection against tick bites and Borrelia infection will make it possible to reduce the incidence of Lyme disease and other diseases.
Biological control
Many natural enemies of ticks including predators, parasites, parasitoids and pathogens have been artificially employed to lower and maintain pest popu-lations at a non-harmful level. Biological control studies have shown that many organisms can reduce the survival, growth, or reproduction success of pest or nuisance species, including those that cause human or animal dis-eases. Many pathogens and predators of blood-sucking arthropods have been known for a long time but few of them have been developed for biocontrol.
Predators and pathogens of ticks Predators: An old review states that predators or natural enemies are not efficient and there is little potential for biological tick control because the populations of ticks are so huge (Cole 1965). More recently, a review found that pathogens and predators contribute significantly towards limiting the population of ticks: spiders, ants, beetles, rodents, shrews, birds and reptiles predating on different genera of ixodid and argasid ticks have potential for biological control (Samish and Rehacek 1999). Another study confirmed that predators are efficient in biocontrol of ticks in different habitats and may reach up to 100% control in some cases (Fleetwood et al. 1984). Predation rates by ants and the house mouse (Mus musculus L.) of engorged females of the cattle tick Boophilus microplus (Canestrini)in pastures were higher in

12
long grass than in medium or short grass (Mwangi et al. 1991b, Sutherst et al. 2000), and two to eight times more effective in open areas than in a pas-ture areas (Fleetwood et al. 1984). Predation of engorged females of the cat-tle tick B. microplus in southern Queensland indicated that ants and the house mouse (M. musculus) were the only significant predators (Sutherst et al. 2000). Nine genera of spiders were identified as predators of five genera of hard ticks and two of soft ticks (Carroll 1995). Some studies have re-ported that the encyrtid wasp Ixodiphagus hookeri is an important parasitoid of Ixodes spp. including I. ricinus (Mwangi et al. 1991a). It has also been reported that I. scapularis is parasitized by the same wasp species (Stafford et al. 1996).
Pathogens: These may enter arthropods through contamination of food; however, this is not an efficient way to introduce pathogens into blood-sucking arthropods such as ticks. Ticks have become adapted as vectors, reservoirs, or propagation sites of rickettsia (Raoult and Roux 1997) and even though they are serving as vectors of Borrelia spirochaetes ticks may be affected by these bacteria (Lefcort and Durden 1996). In some cases, de-pending on the degree of infection and other circumstances, such an infec-tion may lead to death of the tick. Other bacteria are non-specific to ticks and some of them are also pathogenic to domestic animals, and even to humans (Samish and Rehacek 1999). Tick-borne viruses can cause damage to ticks, e.g. to the salivary glands, but they do not play an important role in reducing tick populations (Samish and Rehacek 1999), although recent studies found that baculoviruses may act as biological acaricides against ticks (Assenga et al. 2006)
Fungi have the ability to enter through the cuticle and to disperse in the tick body: they have been reported as major pathogens of ticks (Samish and Rehacek 1999). A study of different habitats indicated that the viability of I. ricinus eggs is lower in a forest biotope, where fungi are present, than in other environments, where the fungi are absent (Honzakova et al. 1975). Spores of the entomopathogenic fungus Metarhizium anisopliae (Metschn.) caused 96% mortality in the laboratory compared to 53% mortality in the field to unfed adult Ixodes scapularis Say. (Benjamin et al. 2002). Entomo-pathogenic fungi seem to have a significant impact on the size of I. ricinus population, with adult females being the most frequently infected stage (Kalsbeek et al. 1995).
Chemical Control
Personal protection has generally focused on the use of chemical repellents and acaricides. A repellent is “a chemical that, acting in the vapour phase, prevents an insect (or other arthropod) from reaching a target to which it would otherwise be attracted” (Browne 1977).

13
Chemical repellents include synthetic and natural repellents for use on the skin and clothes of humans as well as on the fur of domestic animals. Toxi-cants, such as permethrin and other pyrethroids can be used on e.g., clothes, bed nets, sleeping bags and tents. Repellents do not all share a single mode of action, and surprisingly little is known about how repellents act on their target arthropods (Davis 1985). Moreover, the responses to these compounds vary; e.g., different species of mosquitoes may react differently to the same repellent (Rutledge et al. 1983, Curtis et al. 1987). The most desirable char-acteristics of repellents applied on skin include: distance activity and long-lasting effect, if possible against many arthropods: not causing adaptation or resistance; and non-irritating to skin. They should also be almost without smell or colour, and of low cost (Barnard 2000). Repellents to be used for the impregnation of clothes should be resistant to washing, harsh weather or other rough conditions and should be persistent (resistant to chemical dena-turing) (Barnard 2000). It is important to note that repellents on skin usually last for only a few hours whereas pyrethroids on fabric may last for between 6 months and several years (Barnard 2000).
Synthetic repellents Numerous synthetic chemical insect repellents have been discovered in the last half of the 20th century. DEET (N, N-diethyl-3-methylbenzamide) super-seded all of the repellents tested in the 1950s and still remains the principal effective repellent in use (Gupta and Rutledge 1994). DEET is an effective broad-spectrum repellent and widely available. The majority of commercial arthropod repellents for human protection contain DEET (Gupta and Rutledge 1989), which is primarily a mosquito repellent, but can also act as a tick repellent and shows effect against I. scapularis (Carroll et al. 2004). DEET together with plant oils occurs, in a commercially available repellent, MyggA in Sweden and has significant repellent activity against nymphs of I. ricinus (Jaenson et al. 2003). The behavioural response of mosquitoes to DEET varies within and between species in relation to the geographical ori-gin of the test strains (Rutledge et al. 1978). Some anopheline vectors of malaria, such as Anopheles pulcherrimus Theobald and An. Albimanus Lin-naeus, are more tolerant to DEET than Aedes mosquitoes, particularly Ae. aegypti (Curtis et al. 1987, Badolo et al. 2004).
An active repellent recently developed using molecular modelling tech-niques is 1-piperidinecarboxylic acid, 2-(2-hydroxyethyl)-, 1- methylpro-pylester also known as KBR 3023, Icaridin or Picaridin, marketed by Bayer AG under the trade name Bayrepel® (Nentwig et al. 2002). KBR 3023 was at least as effective as DEET against the malaria vector An. gambiae and a West African urban strain of Ae. aegypti in a laboratory study (Badolo et al. 2004). A study of topical repellents has evaluated DEET and KBR 3023 against Amblyomma hebraeum: KBR 3023 was less effective at 1hr than DEET in the laboratory (Pretorius et al. 2003).

14
Another chemical repellent namely 3-(N-n-butyl-N-acetyl)-aminopropionic acid ethyl ester (Insect-Repellent 3535 or IR3535) has been used in Europe for more than 20 years and was approved for use in the United States in 1999. The repellency of IR 3535 is equal to DEET against Ae. aegypt, Culex quinquefasciatus (Say) and Cx. Tritaeniorhynchus Giles (Thavara et al. 2001). Another study of a commercially available repellent spray that con-tains DEET and IR3535 has shown that the repellency of an insect repellent spray is moderately effective in reducing tick bites and the risk of tick-borne diseases (Staub et al. 2002).
The repellent 1-(3-cyclohexen-1-yl-carbonyl)-2-methylpiperidine (AI3-37220) has shown to protect against the ticks I. scapularis and A. ameri-canum. It showed more repellent efficacy than DEET against A. americanum (Solberg et al. 1995, Carroll et al. 2004). Laboratory studies on mosquitoes indicated that AI3-37220 offers as much protection as DEET against An. albimanus and greater protection against Ae. aegypti (Klun et al. 2004).
In the development of new repellents, their safety, both for the environ-ment in general and for the user, is of great importance; this encourages the exploration of natural products.
Natural repellents Plants produce a huge range of so-called secondary compounds, which may have an ecological function as pollinator attractants (color, smell), chemicals for adaptation to physical stresses, attractants of natural enemies (predators or parasitoids); or as repellents, or antifeedants against herbivorous insects. Many of these compounds are also effective repellents against mosquitoes and other biting arthropods, against microorganisms (phytoalexins), and other plants. Such allelochemicals play a role in “chemical warfare” between plants (allelopathic interactions) and include natural herbicides, which act in “weed aggressiveness”, competition, and in regulation of plant density (Balandrin et al. 1985). Extracts and essential oils from numerous plants species repel or kill insects. Insecticidal plants are used mainly in the “de-veloping” countries, where plants may be grown locally or harvested from natural populations. They are less expensive than synthetic chemicals, which are often petrochemical derivatives (Balandrin et al. 1985).
Plant-based repellents against ticks (Acari: Ixodoidea) Plant-derived chemicals that kill or repel ticks are extracted from various parts of the plant, including the flowers, fruits, leaves, and wood. During the early 1990s, research to find alternatives to DEET focused on the oil of Corymbia citriodora (Hook.) K. D. Hill & L. A. S. Johnson, because it was known to be a natural insect repellent (Schreck and Leonhardt 1991). Para-menthane-3, 8-diol (PMD) from C. citriodora essential oil is the active in-gredient in MyggA Natural, a commercially available repellent against mos-quitoes in Sweden. It has been shown to repel nymphs of I. ricinus in the

15
laboratory and field (Jaenson et al. 2005b). Para-menthane-3, 8-diol (PMD) has broad-spectrum arthropod repellent properties. PMD has been evaluated in the laboratory and field against I. ricinus nymphs: tick attachment and feeding were reduced on rabbits treated with PMD (Trigg 1996) and on hu-mans (Gardulf et al. 2004).
Swedish plants whose leaf extracts or oils have been studied for arthropod repellency include Myrica gale L., Rhododendron tomentosum (Stokes) H. Harmaja (formerly Ledum palustre L.) and Artemisia absinthium L. (Jaenson et al. 2005a). In another study in Sweden, various plants or plant-derived substances were tested on I. ricinus. It was shown that citronella (Cymbopo-gon nardus (L.) Rendle), cloves (Allium sativum L.) and lily of the valley (Convallaria majalis L.) had a long lasting repellent effect (up to 8 hr), while Achillea millefolium L. and volatile oils of lavender (Lavandula spp.), gera-nium (Pelargonium spp.), eucalyptus (Eucalyptus spp.), birch (Betula spp.), peppermint (Mentha spp.) and/or pine (Pinus spp.) tar showed some repel-lency initially and just for a short duration (Thorsell et al. 2006). Cleome monophylla L. oil exhibits tick repellency; at the highest dose, its effect was comparable to that of DEET (Ndungu et al. 1995). A study of Gynandropsis gynandra (L.) Birq. showed that it has repellent and acaricidal properties to Rhipicephalus appendiculatus and Amblyomma variegatum ticks of all ac-tive stages (Malonza et al. 1992, Ndungu et al. 1995). Another study showed that G. gynandra essential oil repels R. appendiculatus (Lwande et al. 1998). In research conducted with six grass species namely Cynodon dactylon (L.) Pers., Brachiaria decumbens (Stapf.), Melinis minutiflora (P.) Beauv., Pen-nisetum clandestinum (Hochst.) ex Chiov., Hyparrhenia rufa (Stapf.) and Andropogon gayanus Kunth. it was found that while they all exhibit repel-lency, the Andropogon and Melinis species exhibit the greatest anti-tick re-pellent property against larvae of Boophilus microplus (Thompson et al. 1978). Extract from various African plants, including Margaritaria dis-coidea (Bail.) G. L. Webster. , Ocimum suave (Willd.) (oil extracted from leaves) and G. gynandra have strong acaricidal and/or repellent activity to R. appendiculatus (Kayaa 2000). The tropical plants Stylosanthes humilis (Kunth.) and S. hamata (L.) Taub repel B. microplus larvae; the repellent activity was found in all parts of the plants (Francisco et al. 2003).
Neem oil is widely used in its raw form as an agricultural insecticide and more recently has been employed against medically important insects and ticks. Extensive research has now been carried out to determine the potential effects of neem products against arthropods of medical and veterinary im-portance. It has a range of modes of action against insects including growth regulation, fecundity suppression, oviposition repellency and antifeedancy (Mulla and Su 1999). A study on ticks showed that neem oil induced signifi-cant mortality rates of newly hatched larvae, unfed larvae, and unfed adults of Hyalomma anatolicum excavatum Koch (Abdel-Shafy and Zayed 2002). Undiluted neem oil (100%) has been found to produce 100% mortality in

16
Amblyomma variegatum Santos larvae within 48 h (Ndumu et al. 1999). The repellency of neem oil to hematophagous arthropods, especially mosquito species, has been tested. Some researchers have noted that different stages of ticks may react differently to the same plant extract. One study suggested that the larval stage of I. scapularis is several times more sensitive to the acaricides than the nymphal stage (Panella et al. 1997).
Personal Protection
Many people are unaware of having been bitten by a tick – ticks are very small and their bites are usually painless. Consequently, public information on personal protection against tick bites and Borrelia infection is the best method for controlling Lyme disease or other tick-borne diseases.
When being outdoors in areas that could be tick infested clothing should be light coloured so that ticks can be easily seen; sleeves and trouser legs should be long; trouser leg bottoms should be tucked into socks or boots; long, loose hair should be covered, braided or tied. Exposed skin and cloth-ing should be sprayed with a tick repellent. When coming in from outside activities where you might have encountered ticks, it is important to do a tick check and remove any ticks as soon as possible. Attached ticks should be removed promptly and carefully to prevent disease transmission, the risk of which appears to increase with the duration of attachment (Crippa et al. 2002). Using fine pointed tweezers, the tick should be grasped as close to the skin as possible without squeezing the tick's body.

17
Plants used in this study Lemon eucalyptus or lemon-scented gum are the common names for Corymbia citriodora Hook. (family Myrtaceae). It is a perennial, aromatic tree of Australian origin that has been introduced into other parts of the world. The essential oil, extracted from freshly harvested or partially dried leaves, is widely used in the fragrance industry and for the production of synthetic menthol from citronellal, the major compound of the oil ( Rao et al. 2003). Steam-distilled C. citriodora leaf oil, is chiefly composed of citronel-lal, citronellol, 1,8-cineole, alpha-pinene, beta-pinene, caryophyllene, citronellyl-acetate, limonene, linalool, isopulegol, neo-isopulegol and eugenol (Duke 2004). The oil has a strong fresh citronella-like odor with a sweet balsamic undertone. A monoterpene of relatively low volatility, p-menthane-3,8-diol (PMD), obtained from lemon eucalyptus (C. citriodora), has been known for a long time in China for its mosquito repellent proper-ties, and has shown particular promise in this regard (Curtis et al. 1991, Trigg and Hill 1996).
Lavenders: Commercial essential oil of lavender, obtained by steam dis-
tillation from the flowers of Lavandula spp. belongs to the family La-miaceae. The common name for any plant of the genus Lavandula is laven-der. These plants grow to a height of around 50 cm. The most commonly cultivated species are: Lavandula angustifolia Mill. (English lavender) which is used in study I, L. latifolia Medik. (a Mediterranean grass-like plant), Lavandula stoechas L. (butterfly-like) and L. intermedia Emeric ex Loisel. Lavender oil is a complex of several chemical compounds. Its main contents are lavandulol, 1,8-cineole, lavandulyl acetate, linalyl acetate (3,7-dimethyl-1, 6-octadien-3-yl acetate), linalool (3,7-dimethylocta-1,6-dien-3-ol) and camphor (Duke 2004). Recently investigations were carried out to evaluate the biological activity of the various Lavandula species. Several studies have shown that both L. angustifolia oil and linalool are acaricidal (Cavanagh and Wilkinson 2002).
Geranium: Essential oil of geranium is obtained by steam distillation of
the herbaceous parts of Pelargonium graveolens L'Her. (family Gerani-aceae). Commercial geranium oil is obtained from the scented leaves of a number of Pelargonium spp. grown mainly in Egypt, Morocco, Reunion, and China. Large numbers of species are hybrids (Lis-Balchin 1995). Gera-nium oil contains: 3-hexenol, menthone, isomenthone, linalool, �-bourbonene, citronellyl formate, �-caryophyllene, geranyl formate, citronel-lol, geranyl acetate, citronellyl butyrate, geraniol and 2-phenyl-ethyl-alcohol (Duke 2004). Certain Pelargonium species are used for producing commer-cial rose-scented geranium oil (Kulkarni et al. 1998).

18
Neem is the common name for Azadirachta indica Adrs. Juss. (syn. Ante-laea azadirachta (L.) Adelb, Melia azadirachta L.) belonging to the Meli-aceae family. The neem tree is fast growing, may reach up to 25 m and originates from southern and southeastern Asia, though today it grows in tropical and subtropical areas of Africa, Australia, and America (Schmutterer 1990). The neem tree is quite tolerant of severe drought and poor soil. The essential oil is obtained from the seeds of the neem tree and is known for its insecticidal, antifeedant, and repellent properties (Mulla and Su 1999). Its main active constituent is azadirachtin, which constitutes approximately 40% of the oil in neem seeds (Isman et al. 1990). Other active compounds in neem seed kernels are salannin, salannol, salannol acetate, 3-deacetylsalannin, azadiradione, 14-epoxyazaradion, gedunin, nimbinen and deacetylnimbinen (Schmutterer 1990).
Methyl jasmonate (MJ): This is a volatile derivative of jasmonic acid,
which is involved in plant–plant communication and plant defence against herbivore attack (Bruin and Dicke 2001). It has been described as a compo-nent of the essential oil of Jasminum grandiforum L. and Rosmarinus offici-nalis L. (Duke 2004). MJ occurs naturally in most higher plant species and it is proposed that MJ plays a role in plant wound and pathogen responses (Creelman et al. 1992). Biosynthesis of MJ starts with linolenic acid and proceeds through a number of stages involving lipoxidation, cyclisation and �-oxidation (Vick and Zimmerman 1984).
Hyptis suaveolens (L.) Poit belongs to the family Lamiaceae and is a na-
tive of a tropical America, but is now found throughout the tropics and sub-tropics. H. suaveolens is normally restricted to disturbed sites and may be considered as a ruderal species (Wulff 1973). Annual or perennial, broad-leaved, erect, strongly aromatic herbs, up to 3 m high. Hyptis is frequently used for the treatment of gastrointestinal infections, pain, cramps, and in the treatment of skin infections (Azevedo et al. 2002). The essential oil from the aerial parts of H. suaveolens has been analyzed by GC-flame ionization detector and GC-MS (Rivas et al. 2002); the most abundant constituents of the oil were found to be: 1,8-cineole, fenchone and �-pinene. However, the oils from H. suaveolens vary in composition according to the geographic origin of the plants (Azevedo et al. 2001).
Pistacia atlantica Desf. (family: Anacardiaceae), is a decidous tree, 10-20
m in height, which grows wild in Mediterranean regions. The leaves are ovate or oblong in outline and flowers are very short pedicelled. This plant has been reputed to possess medicinal properties and is traditionally used in Libya as a treatment for gastrointestinal infections and abdominal discomfort (El-Gadi and Bshana 1997). Various constituents have been reported: for instance, �-pinene in the essential oil (Delazar et al. 2004), as well as

19
monoterpenes and oxygenated sesquiterpenes such as terpinen-4-ol and elemol (Barrero et al. 2005).
Juniperus phoenicea L. belongs to the family Cupressaceae is an 8 m
high tree, native to the Mediterranean. The branches are erect; leaves are needle-like and fruits are reddish brown. The young twigs are widely rec-ommended by herbalists for stomach inflammation, and for treatment of urinary tract disorders (El-Gadi and El- Mogrebi 2003). The dry leaves are used to treat mild skin inflammations of babies in Egypt (Batanouny et al. 1999), and previous generations in Libya smouldered this plant to repel in-sects. The essential oil of J. phoenicea has been analyzed by GC-MS and the major compounds were found to be: �-pinene, �-pinene, �-3-carene, sa-binene, myrcene, �-phellandrene, limonene, and D-germacrene (Angioni et al. 2003).
Salvadora persica Wall (family: Salvadoraceae) is an evergreen shrub or
small tree, up to 6-7 m in height; the main trunk is erect or trailing and is abundantly branched. The leaf of S. persica is known for its antisyphilitic effects and the root is used for cleaning the teeth and gums (Batanouny et al. 1999). An extract of the leaves has a considerable anti-bacterial effect on several different oral aerobic bacteria (Alali and Al-Lafi 2003). The stem contains N-benzyl-2-phenylacetamide which has been revealed to have a moderate anti-bacterial activity against Escherichia coli and a significant inhibitory effect on human collagen (Khalil 2006). The volatile oil extracted from S. persica leaves was analyzed by GC-MS and found to contain: ben-zyl nitrile, eugenol, thymol, isothymol, eucalyptol, isoterpinolene and �-caryophyllene (Alali and Al-Lafi 2003).

20
Aims The specific aims of this thesis were to:
1. Evaluate the potential of selected plant products as sources for effective and cheap tick-repellent chemicals to protect people.
2. Evaluate in field experiments the effectiveness of selected compounds at different concentrations. 3. Identify compounds that may be used against the tick I. ricinus. 4. Investigate oils extracted from the leaves of plants from Guinea Bissau and Laos that have known repellency against mosquito, and investigate oils from Libyan plants that are considered to be of medicinal use and might also repel I. ricinus nymphs. 5. Study the infra-specific variability in the chemical constituents of oils from individual plants, for example plant grown in different localities and to identify the oil constituents which vary.

21
Materials and Methods
Study areas The field studies and tick samplings (for laboratory tests) have been con-ducted in a woodland area (Alsike, Uppland) where the density of I. ricinus is known to be high. Alsike (59o47' N, 17o40'E), is situated about 10 km south of Uppsala, east-central Sweden. Field work was performed between 2004 and 2006, generally from May/June to August/September. Addition-ally, one field study was conducted at Lidingö, Stockholm in September 2004.
The vegetation at the locality of Alsike is mainly mixed coniferous forest (spruce, Picea) and broad-leave/ deciduous (Betula) forest with trees, i.e., Pinus, Populus, Alnus, Salix, Querqus and Corylus. The ground and field vegetation consists mainly of lichens, bryophytes, ferns (Dryopteris), grasses (Poaceae), blueberry and lingonberry (Vaccinium spp.), meadow-sweet (Filipendula) and raspberry (Rubus). Lidingö is a mixed spruce (Picea sp.) – broad-leave (Betula sp.) woodland area with a ground layer similar to the one at Alsike. Tick hosts in Sweden are mainly small, medium-sized and large mammals and ground-feeding birds (Jaenson et al. 1994).
Sampling method The ticks were collected by dragging a 1 m2 cotton flannel cloth over the vegetation (Mejlon and Jaenson 1993). The cloths were inspected every 10 m to collect all nymphs found. Collections and field experiments were al-ways performed during dry weather in daylight hours between 9 a.m. and 7 p.m. Sampling was not conducted shortly after or during rainfall. Rainy weather would increase the rate of loss of repellent activity of the test sub-stances. Temperature and humidity were recorded each sampling day before and after collection and field experiments.
All ticks brought back to the laboratory were put in tubes with moistened tissue paper and kept at 4 °C and 80-95% relative humidity in complete darkness until the ticks were used in laboratory experiments. Ticks used in the bioassays had been in captivity for between 1 week and 7 months and were discarded after one test. Prior to testing, they were adapted to the test environment for 24 h (� 22°C and a photoperiod of 12: 12 h).

22
Plant oils and arthropod repellents There are plant oils and other natural ingredients in some formulations of commercial insect repellents tested on I. ricinus in a separate study of this thesis. In study I, lemon-scented eucalyptus (C. citriodora) oil (30%) was tested in the laboratory and in the field. C. citriodora is present in some commer-cial anti-mosquito products in Europe and North America. MyggA Natural®, a commercially available repellent against blood-feeding arthropods in Swe-den, contains 30% of C. citriodora oil with a minimum of 50% p-menthane-3, 8-diol (PMD), an active ingredient of many mosquito- repellent products. This product was tested in the laboratory and in the field. The essential oils (1% and 30%) of lavender (L. angustifolia) and geranium (P. graveolens) were tested separately in the laboratory. These oils also occur in MyggA Natural® in small amounts (1%).
In study II, MyggA Natural® spray (new formulation containing 217.9 g/l of PMD) diluted in two concentrations: 3.2 g/m2 and 4.2 g/m2 and 3 g/m2 of RB86 were tested in the field. RB86 contains about 70% neem oil, about 30% rape seed oil and the active ingredients are azadirachtin (0.1%) and citronella (<1%). RB86 is marketed in Sweden as a repellent to be used on horses against ticks and blood-feeding insects.
In study III the effect of the compound methyl jasmonate MJ (Aldrich, 95% purity) was investigated in different concentrations: 0.075, 0.15, 0.30 and 0.75 mg/cm2 (diluted in 1, 2-propanediol) in the laboratory and 0.05, 0.1 and 0.2 mg/cm2 (diluted in acetone) in the field.
In study IV, the essential oils of H. suaveolens (10% in acetone) and the sesquiterpene oxides and sulphides (10% diluted in hexane) which are found in the oils were tested in the laboratory.
In study V, the hydrodistilled essential oils (20% diluted in acetone) of S. persica, P. atlantica and J. phoenicea, were tested against I. ricinus nymphs in the laboratory.
Plant materials (IV and V) The plants were collected where they naturally grow. H. suaveolens was brought from Guinea Bissau and Laos and cultivated in a greenhouse at The Botanical Garden, Uppsala University. In addition H. suaveolens has been grown and distilled in Laos. The leaves of three plant species: S. persica, P. atlantica and J. phoenicea were collected from Al-Jabal Al-Ahder, north-eastern Libya. The plant samples were stored in plastic bags in a freezer at -20°C for a few months and then distilled and the constituents identified in Sweden.

23
Laboratory bioassays
Repellency test on I. ricinus nymphs
A- Tube and palm bioassay (I, III, IV, and V) The experiments were conducted, as described (Jaenson et al. 2003), using one or five unfed I. ricinus nymphs in a FalconTM
vial in each replicate. The Falcon vial is a 50 ml centrifugal tube, 116 x 29 mm, made of transparent plastic. Its wall was perforated with small holes to prevent saturation of air with odor of the experimenter, test substance or control. In each replicate, one previously unfed nymph was first tested with the control for 5 min, and then with the test substance for 5 min in each of 40-100 replicates. The test substances were diluted in 1, 2-propanediol also known as propylene glycol, (study I and III) or in acetone (study IV and V), applied with pipettes to cot-ton cloths and left to dry before each test. The standard application was 50 �l (study I) or 100 �l (study III, IV and V) or 150 �l of substances (study IV). Each cloth was attached with a rubber band to the open upper end (660 mm²) of a Falcon vial. To simulate host stimuli to attract the nymphs the same observer held his right (in the blank control) or left (for the test substance) palm tight to the outside surface of the cloth for each 5 min period. The rea-son for not randomizing the use of left and right palm between controls and tests is the danger of contamination of the control palm with test substance. The numbers of nymphs which attached at the interior surface of the cloth during this period were recorded in the protocol as “attracted” while ticks that did not were recorded as “repelled”. For a tick to be regarded as “at-tracted” to the cloth it had to detach all its legs from the vial’s surface. The position of the nymph, i.e. its location above the tube's bottom was recorded every minute. However, only the results after 2 and 5 min were analyzed. The proportion of nymphs that were below the 30 mm level of the 115 mm tall test tube was presented as strongly repelled by the test substance (study I).
B- Wrapped finger bioassay (V) An additional investigation was conducted during study V to evaluate the re-pellency of P. atlantica, J. phoenicea and S. persica. This investigation em-ployed a modification of the the human bioassay described by Schreck et al. (1995), in which questing nymphs are exposed to a finger wrapped in cloth treated, as follows: 6 × 6.5 cm of cotton cloth was treated with acetone (con-trol) or with 10 μl/ cm2 of 20% oil in acetone using a 100-μl micropipette. After 10 min the cloth had dried and was wrapped around the index finger from the first to the third joints. To keep the cloth wrapped around the finger, three non-smelling dabs of glue were smeared on the upper surface of the in-ner layer of cloth where they overlapped. The control trials with untreated cloth wrapped around the index finger were performed in a similar manner.

24
A Petri dish was attached with glue to the bottom of a larger glass Petri dish which was then filled with water to a depth of 1 cm to prevent ticks from escaping. Ten ticks were collected from the transport vials before each test trial and placed in a Petri dish for 10 min. The finger was tilted to the vertical postion with the tip pointing down within a Petri dish. The locations of the ticks were recorded at 10 min. Any nymph climbing onto repellent-treated cloth was considered as attracted. If the nymph reversed its direction or dropped off before reaching the 3rd joint, the tick was considered as re-pelled. Nymphs dropping off the finger were collected with forceps and placed in another tube. Repellent treatments were tested hourly (with 10 new unfed nymphs) until failure.
Steam distillation (IV and V)
To obtain essential oils of the plants, steam distillation of leaves of H. suaveolens, S. persica, P. atlantica and J. phoenicea was carried out. The leaves of each species were placed in a glass vessel and steam produced in a separate container at elevated pressure was introduced into the vessel. The steam from the vessel containing the leaf/water mixture was cooled in a Lie-big cooler and the plant oil/water mixture was collected in glass vials. This mixture was then extracted with hexane (study IV) or with 95% n-pentane (Kebo) and dried with Mg SO4 (study V).The mixture was filtered and con-centrated under reduced pressure.
Collection of volatile substances from the leaves and oils (IV and V)
The volatiles emitted from both dried and fresh leaves of H. suaveolens were collected with solid phase microextraction (SPME). Leaves (0.5 g) were placed in a glass vial (50 ml), which was sealed with aluminium foil to release the volatiles for 15 min. An SPME syringe, with the fibre coated with 65 μm polydimethylsiloxane–divinylbenzene (Supelco, Bellefont, PA, U.S.A.), was placed in the vial, and the volatiles were sampled for 1 min (study IV).
Volatiles emitted from oils of H. suaveolens, S. persica, P. atlantica and J. phoenicea were collected with a SPME; 100 μl of the oil diluted in ace-tone (concentration = 1/100) was added to a small vial (1 ml), which was placed in a glass beaker (50 ml) after the evaporation of the solvent. An SPME fibre was placed close to the small vial in the beaker, which was sealed with aluminium foil. After 5 min the volatiles emitted from the oil were sampled for 1 min.

25
Identification of compounds (IV and V)
Components were identified by gas chromatography-mass spectrometry (GC–MS) analysis, using a Varian 3400 GC connected to a Finnigan SSQ 7000 instrument with electronic ionization and the ion source at 150°C. A DB-1 (J&W Scientific) capillary column (internal diameter 0.25 mm, film thickness 0.25 μm and length 30 m) was used for the separation of the vola-tiles. The temperature programming was 40°C (1 min) followed by 5°C min�1 to 220°C (12 min). The injector temperature was kept at 215°C and helium was used as carrier gas with a pressure of 67 kPa. The compounds were identified by means of their retention times (GC–MS) and/or mass spectral data (MS) from authentic samples or with reference to the National Institute of Standards and Technology (NIST) library.
Synthesis of test compounds (IV) The syntheses of caryophyllene and humulene episulfides have been re-ported earlier (Ashitani and Nagahama 1999). However, a slightly modified synthesis is described in study IV. To study the relationship between mo-lecular structures of compounds in the leaves and tick repellency (study IV) the hydrocarbons were separated from other natural sources, i.e Humulus lupulus and a fraction of orange oil. Structurally related oxides and sulfides were also synthesized to investigate repellency–activity vs. structure among the sesquiterpenoids with 8, 9 and 10 carbon member rings to compare the changes in activity when substituting oxygen with sulphur.

26
Field experiments
Repellency experiment on blankets (I, II and III) Field trials were conducted using the cotton flannel cloth dragging method: a 1 m2 cloth was attached to a 1 m long wooden pole at adjacent corners. The substances were diluted in acetone (study I, III) and sprayed on one side of the blankets and diluted in acetone or in water (study II) and applied by a paintbrush until the brush was nearly dry (study II). During the test the cloth was dragged with the treated side downwards over the vegetation. Addition-ally, one identical control blanket onto which 100 ml acetone was applied was used in study I; acetone was used in this case instead of 1, 2-propanediol because the latter is more viscous and cannot be sprayed easily. In studies II and III, an untreated blanket was used as control.
Study I was conducted on 4 days during a 6-day period in early June 2004; unsuitable weather prevented us from carrying out the experiment on day 4 and 5. In the early morning of day one 100 ml of the test substance was evenly applied to one side of each one of two light-colored cotton flan-nel blankets (1 m2 each). Test substances used were 20 ml of MyggA Natu-ral® in 80 ml acetone on one blanket and 20 ml 30% C. citriodora oil in 80 ml acetone on the other blanket. In study II we tested all substances during 16 days in late July and early August 2005. The potential anti-tick repellent activity of two concentrations of MyggA Natural® spray (4.2g /m2, 3.2 g /m2) and one of RB86 with 70% neem oil (3 g/m2) were compared with the con-trol. In the early morning of day 0 three light-colored blankets (1m2 each) were impregnated using a paintbrush with 10 ml (MyggA Natural®) or 12 ml (RB86) of the test substance to cover the whole downward surface of the test cloth. Substances used were 4.6 ml (4.2 g) MyggA Natural® spray (contain-ing 1000 mg PMD) in 5.4 ml acetone on one blanket, 3.5 ml (3.2 g) MyggA
Natural® spray (containing 750 mg PMD) in 6.5 ml acetone on one blanket and 3 ml (3 g) RB86 with 70% neem oil containing 3 mg azadirachtin in 9 ml water on one blanket. The cloths were washed and re-impregnated after 2-4 days when the test substance had lost most of its activity, i.e., when the number of nymphs recorded on the test blankets were similar to that of the control blanket. If there was heavy rain on the test days the blankets lost their effect. The cloth in study III was sprayed with 10 ml of the test sub-stance MJ to cover the whole downward surface. The final concentrations of MJ on the cloths were 0.05 mg/cm2, 0.1 mg/cm2 and 0.2 mg/cm2 (diluted in acetone). Nymphs that attached were counted and collected every 10 m2. Each tick was removed and put into marked vial. This procedure was re-peated 10 times for each replicate. The number of nymphs recorded on the treated blanket was compared with the number of nymphs recorded on the control blanket, which was dragged parallel to and about 1 m from the test blankets (by another person or tester) in the same way over the same vegeta-

27
tion. Treated and untreated (control) blankets were allocated by a randomiza-tion procedure to the testers but in such a way that each tester pulled each blanket an equal number of drags each day.
The blankets when not in use were kept separately in air-proof plastic bags to avoid contamination and to avoid loss of activity of substances.
Repellency experiment on trousers An additional field study was conducted (study I), to evaluate the repellency of MyggA Natural® by one person during two days in September 2004 at Lidingö, Stockholm. Two rings, each one about 2 cm broad, of MyggA Natural® "roll on" were applied to the legs on light-coloured trousers (one at 30 cm above ground level, i.e., just below the knee and one at the lower edge, at 0 cm above ground level) . The order in which the treated trousers or the untreated control trousers were worn was randomized. In each “test” the person slowly walked 100 m through the vegetation. Nymphs that attached were counted and collected after every 10 m.
Statistical analyses In the laboratory experiment (all studies), percentage of repellency was cal-culated as [(no. of ticks recorded as "attracted" in the control vial - no. of ticks recorded as "attracted" in the test vial)/ no. of ticks recorded as "at-tracted" in the control vial] x 100. The McNemar chi-square test for the sig-nificance of change in frequencies between control and test substance was used (study I, III and IV).
In the field experiments the statistical procedures generally performed are those adapted for distribution-free or non-parametric data. Kruskal-Wallis one-way analysis, using Stat Soft Statistica 7.0 was performed to determine the effect of each substance, person and day (study I and III). �2- test was used to test if repellent activity of substances declined from day 1 to day 6 (Study II), and Mann-Whitney U-test was used to compare the mean number of ticks/10 m2 between treated trousers and control trousers (study I).
In study II, a type III generalized linear model with a log link function and a Poisson distribution in the advanced linear/non-linear model, using Stat Soft Statistica 7.0 was performed to determine the effect of each treat-ment and person. The percentage repellency of a field experiments (study I, II and III) was calculated using the following formula: % repellency = [(no. of nymphs on control blanket or leg - no. of nymphs on test blanket or leg)/no. of nymphs on control blanket or leg] x 100.

28
Results and Discussion
Laboratory bioassays
Repellency test on I. ricinus nymphs
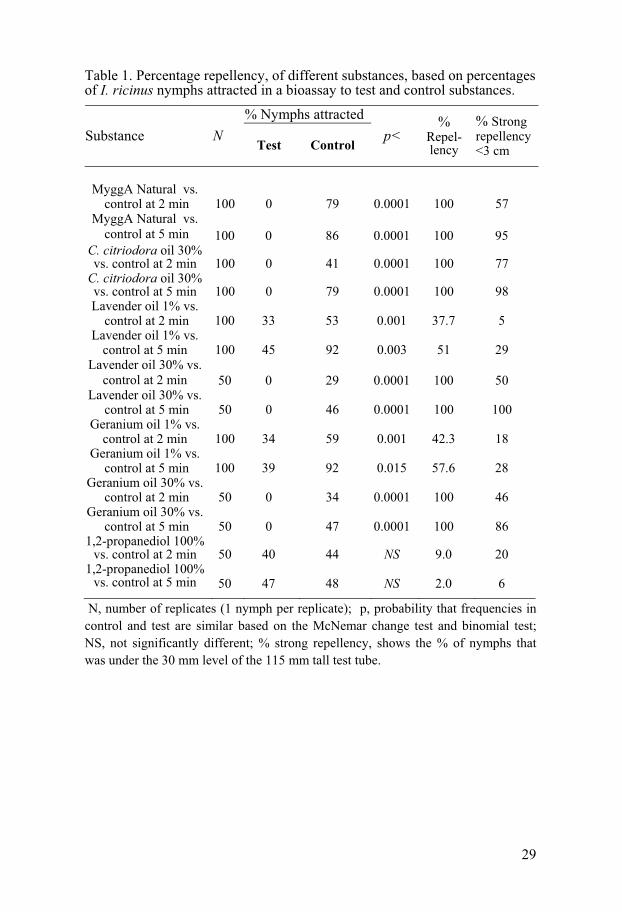
A- Tube and palm bioassay (I, III, IV and V) The results of the laboratory bioassay to test the potential repellency on I. ricinus nymphs of MyggA Natural®, C. citriodora (lemon eucalyptus) oil, L. angustifolium (lavender) oil, P. graveolens (geranium) oil or 1,2-propanediol (study I) after 5 min are shown in Table 1. Generally, the differences be-tween MyggA Natural and the substances which occur in MyggA Natural were significant compared to the control (by McNemar test). 30% C. citrio-dora (containing a minimum of 50% PMD) or 30% lavender or geranium oils showed 100% repellency, whereas 1% lavender or geranium oils gave medium repellency of about 51% and 58%, respectively.
Several trials have been done on PMD in different parts of the world. One laboratory study showed that 30% PMD was almost as effective as DEET the most widely used synthetic repellent (Trigg 1996). PMD seems to be extremely effective in the laboratory as well as in the field. Currently PMD is found in many insect repellents available commercially in many countries in Europe and in the U.S.A.
In laboratory studies oils of geranium, lavender and other plants have been tested. The duration of activity of the substances was short and the ef-fect was lost within 4 h (Thorsell et al. 2006). Essential oils of a large number of plants have been found to have repellent properties against different hematophagous arthropods, and some have formed the basis of commercial repellent formulations (Curtis et al. 1991), an example being MyggA Natural in Sweden .
In study III, methyl jasmonate (MJ) with concentrations of 0.075, 0.15, 0.30 and 0.75 mg/cm2 gave repellencies of 57%, 71%, 92% and 99%, re-spectively. All concentrations of MJ were effective for up to 30 minutes. The McNemar test shows significant differences in repellency among all concentrations of MJ and the controls (Table 2).
MJ has been mainly studied in relation to phytophagous pest control (Birkett et al. 2000, Ibrahim et al. 2005, James 2005). Here we demonstrate the effect of MJ on I. ricinus nymphs in the laboratory.
29
Table 1. Percentage repellency, of different substances, based on percentages of I. ricinus nymphs attracted in a bioassay to test and control substances.
N, number of replicates (1 nymph per replicate); p, probability that frequencies in control and test are similar based on the McNemar change test and binomial test; NS, not significantly different; % strong repellency, shows the % of nymphs that was under the 30 mm level of the 115 mm tall test tube.
% Nymphs attracted Substance
N Test Control
p< %
Repel-lency
% Strong repellency <3 cm
MyggA Natural vs.
control at 2 min 100 0 79 0.0001 100 57 MyggA Natural vs.
control at 5 min 100 0 86 0.0001 100 95 C. citriodora oil 30% vs. control at 2 min 100 0 41 0.0001 100 77
C. citriodora oil 30% vs. control at 5 min 100 0 79 0.0001 100 98 Lavender oil 1% vs.
control at 2 min 100 33 53 0.001 37.7 5 Lavender oil 1% vs.
control at 5 min 100 45 92 0.003 51 29 Lavender oil 30% vs.
control at 2 min 50 0 29 0.0001 100 50 Lavender oil 30% vs.
control at 5 min 50 0 46 0.0001 100 100 Geranium oil 1% vs.
control at 2 min 100 34 59 0.001 42.3 18 Geranium oil 1% vs.
control at 5 min 100 39 92 0.015 57.6 28 Geranium oil 30% vs.
control at 2 min 50 0 34 0.0001 100 46 Geranium oil 30% vs.
control at 5 min 50 0 47 0.0001 100 86 1,2-propanediol 100%
vs. control at 2 min 50 40 44 NS 9.0 20 1,2-propanediol 100%
vs. control at 5 min
50
47
48
NS
2.0
6
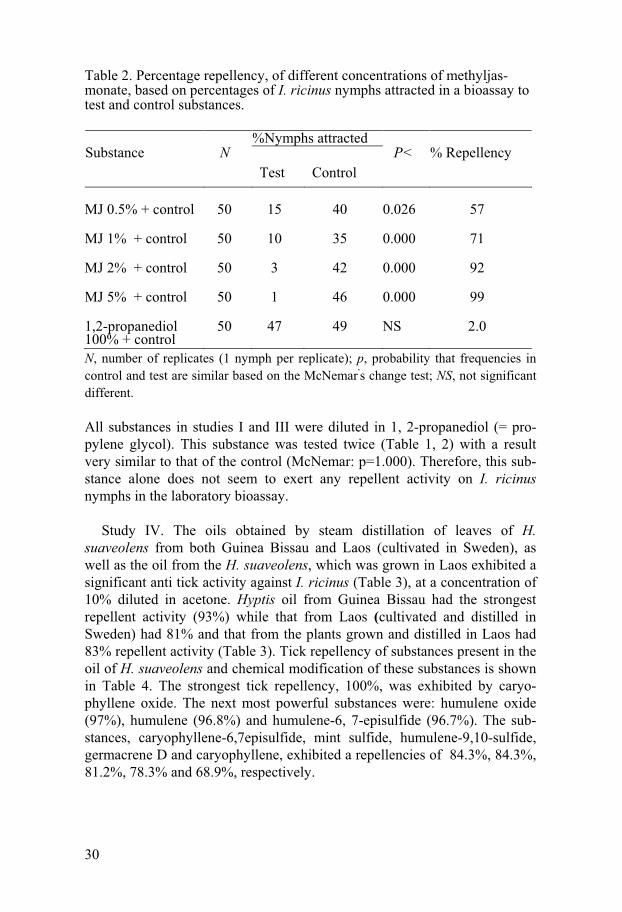
30
Table 2. Percentage repellency, of different concentrations of methyljas-monate, based on percentages of I. ricinus nymphs attracted in a bioassay to test and control substances.
N, number of replicates (1 nymph per replicate); p, probability that frequencies in control and test are similar based on the McNemar
,s change test; NS, not significant
different.
All substances in studies I and III were diluted in 1, 2-propanediol (= pro-pylene glycol). This substance was tested twice (Table 1, 2) with a result very similar to that of the control (McNemar: p=1.000). Therefore, this sub-stance alone does not seem to exert any repellent activity on I. ricinus nymphs in the laboratory bioassay.
Study IV. The oils obtained by steam distillation of leaves of H.
suaveolens from both Guinea Bissau and Laos (cultivated in Sweden), as well as the oil from the H. suaveolens, which was grown in Laos exhibited a significant anti tick activity against I. ricinus (Table 3), at a concentration of 10% diluted in acetone. Hyptis oil from Guinea Bissau had the strongest repellent activity (93%) while that from Laos (cultivated and distilled in Sweden) had 81% and that from the plants grown and distilled in Laos had 83% repellent activity (Table 3). Tick repellency of substances present in the oil of H. suaveolens and chemical modification of these substances is shown in Table 4. The strongest tick repellency, 100%, was exhibited by caryo-phyllene oxide. The next most powerful substances were: humulene oxide (97%), humulene (96.8%) and humulene-6, 7-episulfide (96.7%). The sub-stances, caryophyllene-6,7episulfide, mint sulfide, humulene-9,10-sulfide, germacrene D and caryophyllene, exhibited a repellencies of 84.3%, 84.3%, 81.2%, 78.3% and 68.9%, respectively.
%Nymphs attracted
Substance
N Test Control
P< % Repellency
MJ 0.5% + control
50
15
40
0.026
57
MJ 1% + control
50 10 35 0.000 71
MJ 2% + control
50 3 42 0.000 92
MJ 5% + control
50 1 46 0.000 99
1,2-propanediol 100% + control
50 47 49 NS 2.0
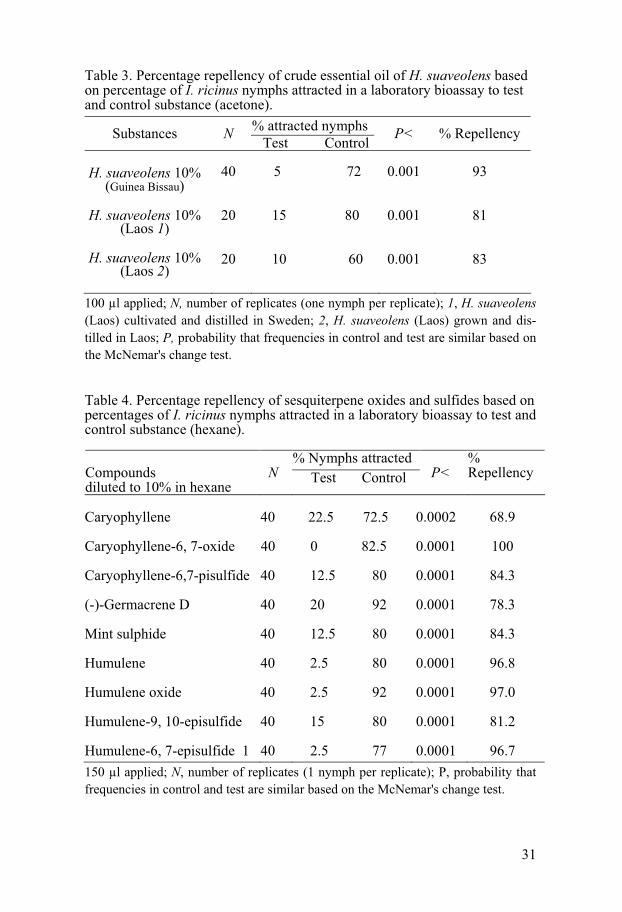
31
Table 3. Percentage repellency of crude essential oil of H. suaveolens based on percentage of I. ricinus nymphs attracted in a laboratory bioassay to test and control substance (acetone).
Substances N % attracted nymphs Test Control
P< % Repellency
H. suaveolens 10%
(Guinea Bissau) 40
5 72
0.001
93
H. suaveolens 10% (Laos 1)
H. suaveolens 10%
(Laos 2)
20
20
15 80
10 60
0.001
0.001
81
83
100 μl applied; N, number of replicates (one nymph per replicate); 1, H. suaveolens (Laos) cultivated and distilled in Sweden; 2, H. suaveolens (Laos) grown and dis-tilled in Laos; P, probability that frequencies in control and test are similar based on the McNemar's change test.
Table 4. Percentage repellency of sesquiterpene oxides and sulfides based on percentages of I. ricinus nymphs attracted in a laboratory bioassay to test and control substance (hexane).
150 μl applied; N, number of replicates (1 nymph per replicate); P, probability that frequencies in control and test are similar based on the McNemar's change test.
% Nymphs attracted Compounds diluted to 10% in hexane
N Test Control P< %Repellency
Caryophyllene
40 22.5 72.5
0.0002
68.9
Caryophyllene-6, 7-oxide 40 0 82.5
0.0001 100
Caryophyllene-6,7-pisulfide
40 12.5 80 0.0001 84.3
(-)-Germacrene D
40 20 92 0.0001 78.3
Mint sulphide
40 12.5 80 0.0001 84.3
Humulene
40 2.5 80 0.0001 96.8
Humulene oxide
40 2.5 92 0.0001 97.0
Humulene-9, 10-episulfide
40 15 80 0.0001 81.2
Humulene-6, 7-episulfide 1 40 2.5 77 0.0001 96.7
32
H. suaveolens is distributed in the tropics and subtropics. Hyptis is well known for its aromatic smell and its essential oil varies in composition according to the geographic origin of the plant (Azevedo et al. 2001, Azevedo et al. 2002). The aromatic plant is traditionally used in Guinea-Bissau, West Africa to repel mosquitoes (Palsson and Jaenson 1999a) and exhibits significant mosquito repellent activity in the laboratory and in the field (Palsson and Jaenson 1999b, Jaenson et al. 2006).
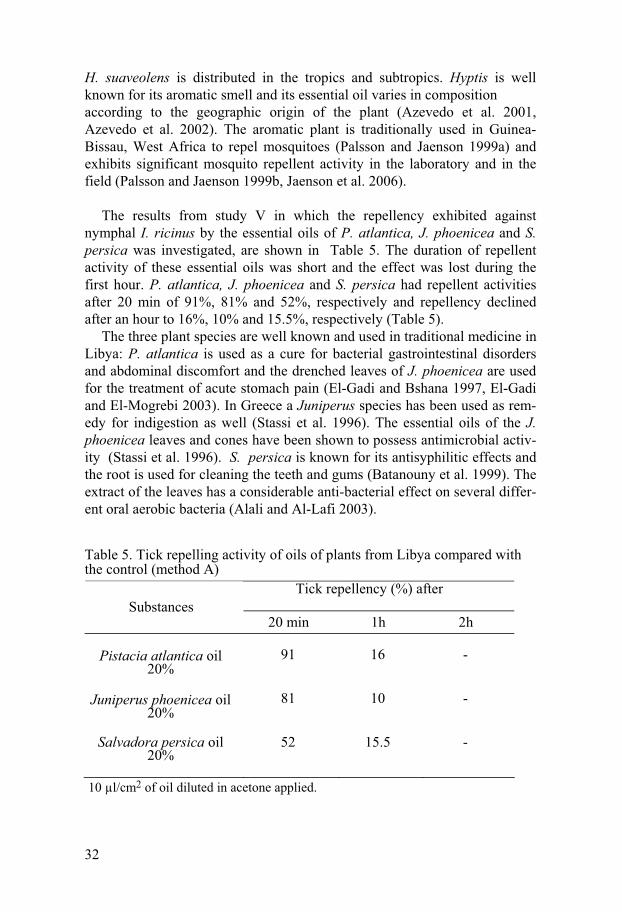
The results from study V in which the repellency exhibited against
nymphal I. ricinus by the essential oils of P. atlantica, J. phoenicea and S. persica was investigated, are shown in Table 5. The duration of repellent activity of these essential oils was short and the effect was lost during the first hour. P. atlantica, J. phoenicea and S. persica had repellent activities after 20 min of 91%, 81% and 52%, respectively and repellency declined after an hour to 16%, 10% and 15.5%, respectively (Table 5).
The three plant species are well known and used in traditional medicine in Libya: P. atlantica is used as a cure for bacterial gastrointestinal disorders and abdominal discomfort and the drenched leaves of J. phoenicea are used for the treatment of acute stomach pain (El-Gadi and Bshana 1997, El-Gadi and El-Mogrebi 2003). In Greece a Juniperus species has been used as rem-edy for indigestion as well (Stassi et al. 1996). The essential oils of the J. phoenicea leaves and cones have been shown to possess antimicrobial activ-ity (Stassi et al. 1996). S. persica is known for its antisyphilitic effects and the root is used for cleaning the teeth and gums (Batanouny et al. 1999). The extract of the leaves has a considerable anti-bacterial effect on several differ-ent oral aerobic bacteria (Alali and Al-Lafi 2003).
Table 5. Tick repelling activity of oils of plants from Libya compared with the control (method A)
Tick repellency (%) after Substances
20 min 1h 2h
Pistacia atlantica oil 20%
91
16
-
Juniperus phoenicea oil 20%
81
10
-
Salvadora persica oil 20%
52 15.5 -
10 μl/cm2 of oil diluted in acetone applied.

33
Our result indicates that the extract and/or oils have a significant repellent activity against I. ricinus nymphs. We are not aware of any previous study concerning the effect of these species on ticks or as repellents of other blood-feeding insects.
B-Wrapped finger bioassay (V) An additional procedure was conducted, to evaluate the tick repellency of plants from Libya. P. atlantica, J. phoenicea and S. persica were tested on I. ricinus in the laboratory for two hours. Repellencies of P. atlantica, J. phoenicea and S. persica declined significantly from 100%, 90.9% and 61% after 20 min; to 0%, 20% and 28% respectively, after an hour (Table 6). The procedure employed, wrapped finger bioassay, allows the ticks to hostseek, so they take a longer time to find the host stimuli than during the tube and palm bioassay. However, both procedures gave almost the same results. The results from the wrapped finger bioassay confirmed that long-lasting protec-tion against ticks may be difficult to accomplish with the use of P. atlantica, J. phoenicea and S. persica.
Table 6. Tick repelling activity of oils of plants from Libya compared with the control (method B)
Tick repellency (%) after
Substance
20min 1h 2h P. atlantica oil 20%
100
0
-
J. phoenicea oil 20%
90.9
20
-
S. persica oil 20%
61 28 14
10 μl/cm2 of oil diluted in acetone applied.
Volatile Compounds from the oils and leaves (IV and V) The main volatile compounds detected by SPME from fresh and dried leaves of H. suaveolens from Guinea-Bissau and Laos (cultivated and distilled in Sweden) in study IV were: �-caryophyllene, limonene, sabinene, 1, 8-cineol, and �-phellandrene and minor amounts of caryophyllene oxide (Table 7). The proportions of caryophyllene oxide are greater in dried leaves than in fresh ones. The naturally occurring sesquiterpenes caryophyllene, humulene and caryophyllene oxide in fresh and dried H. suaveolens leaves were found to be highly tick repellent (Table 4).
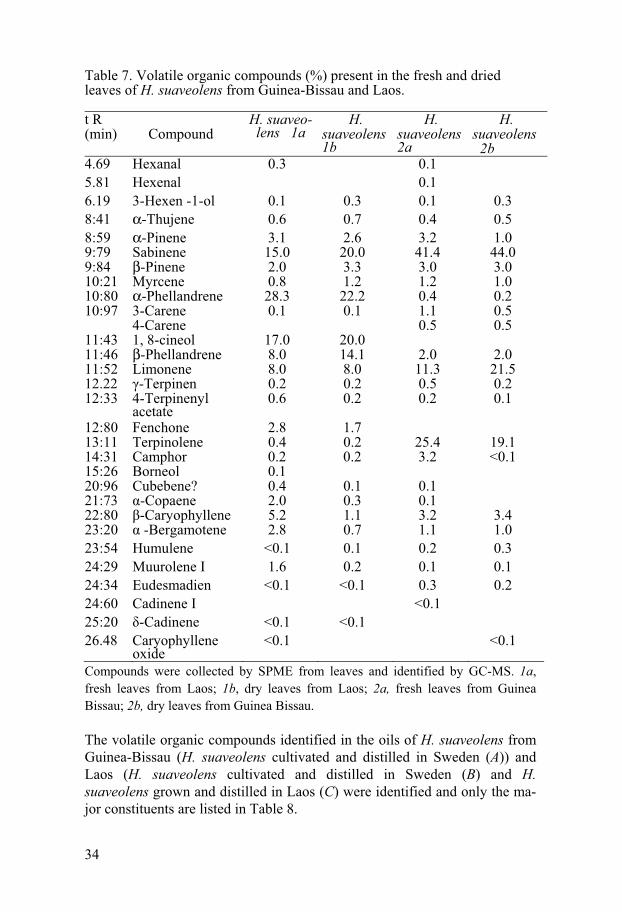
34
Table 7. Volatile organic compounds (%) present in the fresh and dried leaves of H. suaveolens from Guinea-Bissau and Laos.
Compounds were collected by SPME from leaves and identified by GC-MS. 1a, fresh leaves from Laos; 1b, dry leaves from Laos; 2a, fresh leaves from Guinea Bissau; 2b, dry leaves from Guinea Bissau.
The volatile organic compounds identified in the oils of H. suaveolens from Guinea-Bissau (H. suaveolens cultivated and distilled in Sweden (A)) and Laos (H. suaveolens cultivated and distilled in Sweden (B) and H. suaveolens grown and distilled in Laos (C) were identified and only the ma-jor constituents are listed in Table 8.
t R (min)
Compound
H. suaveo-lens 1a
H.suaveolens 1b
H.suaveolens 2a
H. suaveolens 2b
4.69 Hexanal 0.3 0.1 5.81 Hexenal 0.1 6.19 3-Hexen -1-ol 0.1 0.3 0.1 0.3 8:41 α-Thujene 0.6 0.7 0.4 0.5 8:59 α-Pinene 3.1 2.6 3.2 1.0 9:79 Sabinene 15.0 20.0 41.4 44.0 9:84 β-Pinene 2.0 3.3 3.0 3.0 10:21 Myrcene 0.8 1.2 1.2 1.0 10:80 α-Phellandrene 28.3 22.2 0.4 0.2 10:97 3-Carene 0.1 0.1 1.1 0.5 4-Carene 0.5 0.5 11:43 1, 8-cineol 17.0 20.0 11:46 β-Phellandrene 8.0 14.1 2.0 2.0 11:52 Limonene 8.0 8.0 11.3 21.5 12.22 -Terpinen 0.2 0.2 0.5 0.2 12:33 4-Terpinenyl
acetate 0.6
0.2 0.2 0.1
12:80 Fenchone 2.8 1.7 13:11 Terpinolene 0.4 0.2 25.4 19.1 14:31 Camphor 0.2 0.2 3.2 <0.1 15:26 Borneol 0.1 20:96 Cubebene? 0.4 0.1 0.1 21:73 �-Copaene 2.0 0.3 0.1 22:80 �-Caryophyllene 5.2 1.1 3.2 3.4 23:20 � -Bergamotene 2.8 0.7 1.1 1.0 23:54 Humulene <0.1 0.1 0.2 0.3 24:29 Muurolene I 1.6 0.2 0.1 0.1 24:34 Eudesmadien <0.1 <0.1 0.3 0.2 24:60 Cadinene I <0.1 25:20 �-Cadinene <0.1 <0.1 26.48 Caryophyllene
oxide <0.1 <0.1
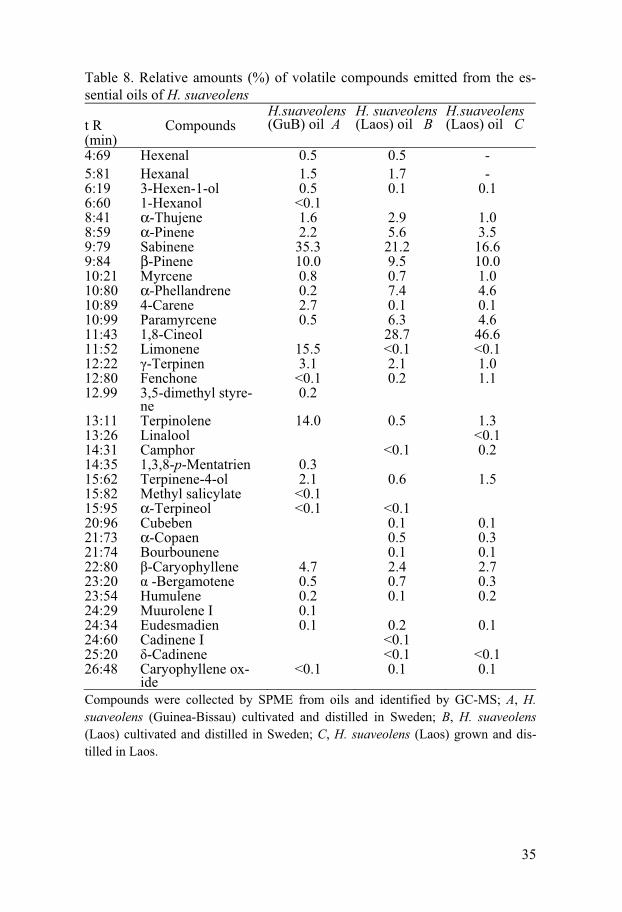
35
Table 8. Relative amounts (%) of volatile compounds emitted from the es-sential oils of H. suaveolens t R (min)
Compounds
H.suaveolens (GuB) oil A
H. suaveolens (Laos) oil B
H.suaveolens (Laos) oil C
4:69 Hexenal 0.5 0.5 - 5:81 Hexanal 1.5 1.7 - 6:19 3-Hexen-1-ol 0.5 0.1 0.1 6:60 1-Hexanol <0.1 8:41 α-Thujene 1.6 2.9 1.0 8:59 α-Pinene 2.2 5.6 3.5 9:79 Sabinene 35.3 21.2 16.6 9:84 β-Pinene 10.0 9.5 10.0 10:21 Myrcene 0.8 0.7 1.0 10:80 α-Phellandrene 0.2 7.4 4.6 10:89 4-Carene 2.7 0.1 0.1 10:99 Paramyrcene 0.5 6.3 4.6 11:43 1,8-Cineol 28.7 46.6 11:52 Limonene 15.5 <0.1 <0.1 12:22 -Terpinen 3.1 2.1 1.0 12:80 Fenchone <0.1 0.2 1.1 12.99 3,5-dimethyl styre-
ne 0.2
13:11 Terpinolene 14.0 0.5 1.3 13:26 Linalool <0.1 14:31 Camphor <0.1 0.2 14:35 1,3,8-p-Mentatrien 0.3 15:62 Terpinene-4-ol 2.1 0.6 1.5 15:82 Methyl salicylate <0.1 15:95 α-Terpineol <0.1 <0.1 20:96 Cubeben 0.1 0.1 21:73 α-Copaen 0.5 0.3 21:74 Bourbounene 0.1 0.1 22:80 �-Caryophyllene 4.7 2.4 2.7 23:20 � -Bergamotene 0.5 0.7 0.3 23:54 Humulene 0.2 0.1 0.2 24:29 Muurolene I 0.1 24:34 Eudesmadien 0.1 0.2 0.1 24:60 Cadinene I <0.1 25:20 �-Cadinene <0.1 <0.1 26:48 Caryophyllene ox-
ide <0.1 0.1 0.1
Compounds were collected by SPME from oils and identified by GC-MS; A, H. suaveolens (Guinea-Bissau) cultivated and distilled in Sweden; B, H. suaveolens (Laos) cultivated and distilled in Sweden; C, H. suaveolens (Laos) grown and dis-tilled in Laos.

36
Important differences in the amounts of the major constituents were found. The Highest percentages of limonene, terpinolene, sabinene and �-caryophyllene were found in the oil of H. suaveolens from Guinea-Bissau (A) than in the other oils (B and C).
The oil of H. suaveolens from Laos B and C contained large amount of 1, 8-cineol and �- phellandrene. Limonene was only found in trace amount from H. suaveolens oil from Laos (B and C) and 1, 8-cineol was not found in H. suaveolens oil from Guinea-Bissau (A), whereas a Limonene was present (15.5%) in the H. suaveolens oil from Guinea-Bissau and 1, 8-carene was present (28.7% and 46.6%) in the H. suaveolens oil from Laos B and C (Ta-ble 8). The oils distilled from H. suaveolens cultivated in Sweden (A and B) contained hexenal and hexanal which were absent from the oil (C) from H. suaveolens grown in Laos. Geographic variations seem to be a factor influ-encing the oil content (Azevedo et al. 2002). Interestingly, the chemical compounds mentioned above have previously been described as pesticidal, insecticidal and/ or insect repellent according to Duke (2004) and Ndungu et al. (1995a).
Study V. The main volatile compounds detected by SPME from oils of P.
atlantica, J. phoenicea and S. persica are shown in Table 9. The emissions from the J. phoenicea oil obtained here were composed of large amounts of �-pinene, �-pinene, �-myrcene and �-phellandrene. The steam distilled oils of J. phoenicea from Sardinia have been found to contain mainly �-pinene, �-pinene, �-3-carene, sabinene, myrcene, �-phellandrene, limonene, and D-germacrene (Angioni et al. 2003). The main components of the oil of J. phoenicea from Corsica are reported to be �-pinene, �-phellandrene, �-terpinyl acetate, � -3-carene, myrcene and �-phellandrene (Rezzi et al. 2001). We did not detect �-3-carene in the oil examined.
The main volatiles detected from P. atlantica oil were �-pinene, sabinene, �-pinene, �-myrcene, �-phellandrene, 2-carene, terpinolene, terpinen-4-ol and -terpinene (Table 9). All of these compounds are known to be repellent and/or toxic to arthropods (Table 9). The essential oil from aerial parts (from male and female plants) of P. atlantica have been analyzed by GC- MS and GC (Tzakou et al. 2007) and qualitative and quantitative differences among the samples and different organs have been observed. Various compounds were characterized: for instance �-pinene and �-pinene as major components together with sesquiterpenes in the essential oil of Algerian P. atlantica has recently been reported (Mecherara-Idjeri et al. 2008). In another study, oxy-genated monoterpenes and sesquiterpenes such as terpinen-4-ol or elemol have been reported (Barrero et al. 2005). Our findings are in general agree-ment with these previous data.
Volatile compounds from S. persica or Miswak oil include high amounts of 2(E)-hexenal, eucalyptol, �-pinene, �-pinene and caryophllenoxide (Table 9) in combination with smaller amounts of other known insect repellent,
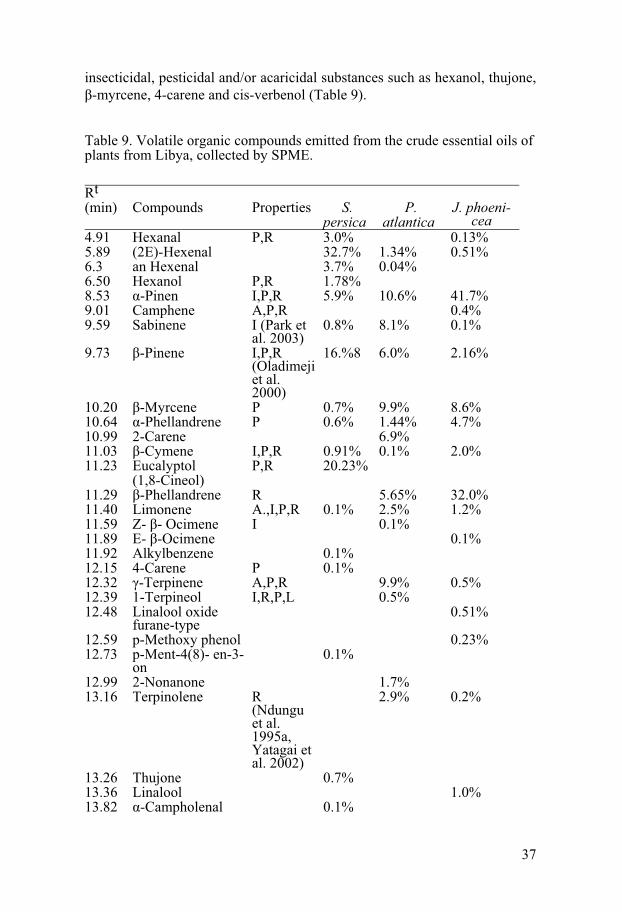
37
insecticidal, pesticidal and/or acaricidal substances such as hexanol, thujone, �-myrcene, 4-carene and cis-verbenol (Table 9).
Table 9. Volatile organic compounds emitted from the crude essential oils of plants from Libya, collected by SPME.
Rt (min)
Compounds
Properties
S.
persica
P.
atlantica
J. phoeni-
cea 4.91 Hexanal P,R 3.0% 0.13% 5.89 (2E)-Hexenal 32.7% 1.34% 0.51% 6.3 an Hexenal 3.7% 0.04% 6.50 Hexanol P,R 1.78% 8.53 �-Pinen I,P,R 5.9% 10.6% 41.7% 9.01 Camphene A,P,R 0.4% 9.59 Sabinene I (Park et
al. 2003) 0.8% 8.1% 0.1%
9.73 �-Pinene I,P,R (Oladimeji et al. 2000)
16.%8 6.0% 2.16%
10.20 �-Myrcene P 0.7% 9.9% 8.6% 10.64 10.99
�-Phellandrene 2-Carene
P
0.6% 1.44% 6.9%
4.7%
11.03 �-Cymene I,P,R 0.91% 0.1% 2.0% 11.23 Eucalyptol
(1,8-Cineol) P,R 20.23%
11.29 �-Phellandrene R 5.65% 32.0% 11.40 Limonene A.,I,P,R 0.1% 2.5% 1.2% 11.59 Z- �- Ocimene I 0.1% 11.89 E- �-Ocimene 0.1% 11.92 Alkylbenzene 0.1% 12.15 4-Carene P 0.1% 12.32 -Terpinene A,P,R 9.9% 0.5% 12.39 1-Terpineol I,R,P,L 0.5% 12.48 Linalool oxide
furane-type 0.51%
12.59 p-Methoxy phenol 0.23% 12.73 p-Ment-4(8)- en-3-
on 0.1%
12.99 2-Nonanone 1.7% 13.16 Terpinolene R
(Ndungu et al. 1995a, Yatagai et al. 2002)
2.9% 0.2%
13.26 Thujone 0.7% 13.36 Linalool 1.0% 13.82 �-Campholenal 0.1%

38
13.90 Pinone 1.2% 14.02 2-Cyclohexen-1-
ol,1-methyl-4-(1-methylethyl-cis)1-terpineol
0.53%
14.29 Camphor 0.4% 14.34 2(10) Pinen-3- ol 1.97% 14.58 cis-Verbenol R 0.2% 14.82 Bicyclo(2,2,1)
hepton-3- one 6,6-methyl-2-methylene
2.36%
15.19 3-Pinanone 0.1% 15.81 6,6-Dimethyl-2-
norpinene-2-carboxaldehyde
2.0%
15.82 Methyl salicylate 0.5% 15.84 Terpinen-4-ol
P ,R (Ndungu et al. 1995a)
0.65% 28.4% 0.28%
16.11
p-Menth-1-en-8-ol(�-Terpineol)
1.5%
1.7%
16.12 2-Pinene-10-ol (+ verbenon)
1.97%
0.2%
16.26 16.52
p-Menth-3-ol cis and trans
0.1%
17.11 Citronellol 0.12% 17.50 Cyclohexen-1-
one-2-isopropyl-5-methyl
0.63%
18.68 Bornyl acetate 0.1% 18.9 2-Undecanone 0.2% 20.50 p-Menthe-1-en-8-
ol, acetate �-Terpinyl acetate
0.08% 0.61%
20.56 Eugenol A,P 0.1% 21.78 �-Bourbonene 0.1% 0.1% 22.67 Caryophyllene P,R,I 0.8% 24.26 Germacrene D P 0.9% 25.2 �-Cadinene 0.1 26.32 Caryophyllenoxide
I,P,R 7.1%
Compounds were identified by MS. A, acaricidal; I, insecticidal; P, pesticidal; R, insect repellent; L, larvicide (according to Duke (2004) or other source(s) as referred to in this table.
Chemical identification of the volatiles in leaves of S. persica from various origins has been done previously (Alali and Al-Lafi 2003, Khalil 2006). The volatile oil extracted from S. persica leaves was analyzed by GC-MS (Alali

39
and Al-Lafi 2003) and identified benzyl nitrile, eugenol, thymol, isothymol, eucalyptol, isoterpinolene, and �-caryophyllene. The main components of the root oil of S. persica from Jordan were benzyl isothiocyanate, eucalyptol, caryophyllene, �-pinene, �-pinene, and limonene (Bader et al. 2002). We did not find benzyl nitrile or thymol in the oil examined (Table 9).
Synthesis of test compounds (IV) The chemical structures of caryophyllene, caryophyllene oxide, caryo-phyllene-6, 7-episulfide, germacrene D, mint sulfide, humulene, humulene oxide, humulene-9, 10-episulfide and humulene-6, 7-episulfide are shown in Table 10. Substitution of oxygen with sulfur, in the same position, in caryo-phyllene oxide, resulted only in an insignificant change in repellency.
Table 10. The chemical structures of the sesquiterpenes and corresponding oxides and sulfides of tested for repellency.
Compounds
Structure
Caryophyllene
Caryophyllene oxide
Caryophyllene-6,7-episulfide
Humulene
Humulene oxide

40
Humulene-9,10-episulfide
Humulene-6,7-episulfide
Humulene oxide
Germacrene D
Mint sulfide
Humulene-6, 7-sulphide had similar repellency, in the same range, as the corresponding oxide. However, humulene-6, 7-episulfide was a stronger tick repellent than humulene-9, 10-episulfide with the sulfur attached to carbons 9 and 10 position (Table 4). Sulfur in this position is less “exposed” since it is covered by two methyl groups compared to humulene6, 7-episulfide with only one methyl group on the adjacent carbon. Mint sulfide which has azule-noide structure and a transannular bonds; it showed less activity compared to both the 9 and 11 membered ring structures of caryophyllene sulfide and humulene sulfides. The high flexibility of the humulene derivatives might be important in the interactions with the receptor site.

41
Field experiments
Repellency experiment on blankets Studies on the repellency of MyggA Natural® and C. citriodora oil in study I, two concentrations of a new formulation of MyggA Natural® spray and RB 86® in study II and three concentrations of MJ in study III were per-formed on I. ricinus during 2004-2006. Generally, all substances showed some repellency but do not seem to provide 100% protection against the ticks. To evaluate the anti-tick activity of MyggA Natural® two test methods, one on blankets and one on trousers were used. Tick repellency of MyggA Natural®, and C. citriodora oil was tested on blankets for 4 days during a 6-day period. The mean number of I. ricinus nymphs on blankets treated with MyggA Natural® or C. citriodora oil was significantly different from that on the control blanket [H (2, N=320) =51.83, p<0.001] (Fig.1). Numbers of nymphs on the test blanket impreg-nated with MyggA Natural® were not significantly different from that on the test blanket impregnated with C. citriodora oil (Mann-Whitney: p=0.55) (Fig.1). Repellency of MyggA Natural® and C. citriodora oil on treated blankets declined from day 1 to day 6 from 74% to 45% and from 85% to 42%, respectively. The repellencies, 73.8% and 85.2%, on day 1 are similar (p=0.39 and p= 0.23) to that (89.3%) of blankets treated in the same manner with 19% DEET (Jaenson et al. 2003).
Repellency experiment on trousers A field test with MyggA Natural® applied to the legs of white cotton trousers showed that with two rings of MyggA Natural on each leg of the treated trousers, 29 nymphs in total were caught compared with 50 nymphs on the untreated legs. The difference is significant (P < 0.05; n = 120). This corresponds to an overall repellency of 42.0%. With five rings of MyggA Natural on each leg of the treated trousers, 13 nymphs in total were caught compared with 32 nymphs on the control legs. The difference is significant (P < 0.05; n = 60) and corresponds to a total repellency of 59.3%. With two rings and five rings of MyggA Natural roll on each leg of the treated trousers, the overall repellencies were 42 and 59%, respectively, which are similar (52%) to that of trousers treated in a similar way with MyggA containing 19% deet (Jaenson et al. 2003). These products are commonly used in Sweden as mosquito repellents and our data demonstrate that they can also be used as tick repellents. One previous study showed that another commercial product, Mosiguard Natural, based on an extract from lemon eucalyptus “may become a useful means in reducing the number of tick bites and thereby tick-borne infections,
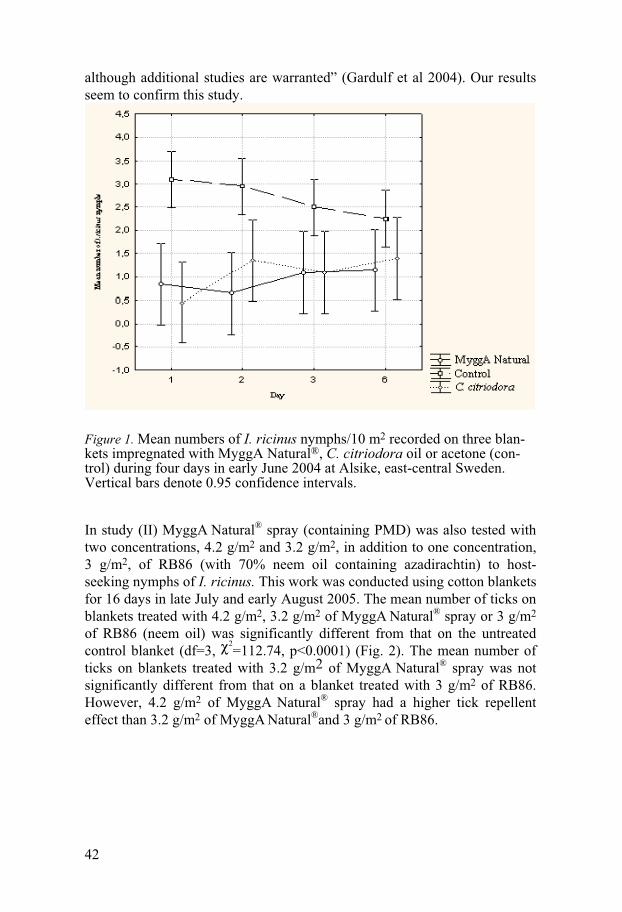
42
although additional studies are warranted” (Gardulf et al 2004). Our results seem to confirm this study.
Figure 1. Mean numbers of I. ricinus nymphs/10 m2 recorded on three blan-kets impregnated with MyggA Natural®, C. citriodora oil or acetone (con-trol) during four days in early June 2004 at Alsike, east-central Sweden. Vertical bars denote 0.95 confidence intervals.
In study (II) MyggA Natural® spray (containing PMD) was also tested with two concentrations, 4.2 g/m2 and 3.2 g/m2, in addition to one concentration, 3 g/m2, of RB86 (with 70% neem oil containing azadirachtin) to host-seeking nymphs of I. ricinus. This work was conducted using cotton blankets for 16 days in late July and early August 2005. The mean number of ticks on blankets treated with 4.2 g/m2, 3.2 g/m2 of MyggA Natural® spray or 3 g/m2
of RB86 (neem oil) was significantly different from that on the untreated control blanket (df=3, χ2
=112.74, p<0.0001) (Fig. 2). The mean number of ticks on blankets treated with 3.2 g/m2 of MyggA Natural® spray was not significantly different from that on a blanket treated with 3 g/m2 of RB86. However, 4.2 g/m2 of MyggA Natural® spray had a higher tick repellent effect than 3.2 g/m2 of MyggA Natural®and 3 g/m2 of RB86.
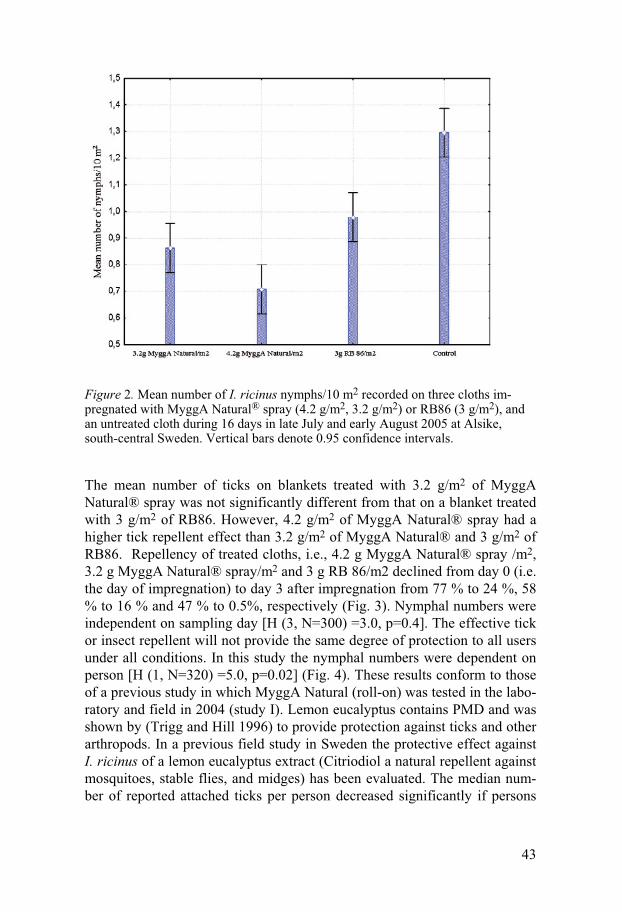
43
Figure 2. Mean number of I. ricinus nymphs/10 m2 recorded on three cloths im-pregnated with MyggA Natural® spray (4.2 g/m2, 3.2 g/m2) or RB86 (3 g/m2), and an untreated cloth during 16 days in late July and early August 2005 at Alsike, south-central Sweden. Vertical bars denote 0.95 confidence intervals.
The mean number of ticks on blankets treated with 3.2 g/m2 of MyggA Natural® spray was not significantly different from that on a blanket treated with 3 g/m2 of RB86. However, 4.2 g/m2 of MyggA Natural® spray had a higher tick repellent effect than 3.2 g/m2 of MyggA Natural® and 3 g/m2 of RB86. Repellency of treated cloths, i.e., 4.2 g MyggA Natural® spray /m2, 3.2 g MyggA Natural® spray/m2 and 3 g RB 86/m2 declined from day 0 (i.e. the day of impregnation) to day 3 after impregnation from 77 % to 24 %, 58 % to 16 % and 47 % to 0.5%, respectively (Fig. 3). Nymphal numbers were independent on sampling day [H (3, N=300) =3.0, p=0.4]. The effective tick or insect repellent will not provide the same degree of protection to all users under all conditions. In this study the nymphal numbers were dependent on person [H (1, N=320) =5.0, p=0.02] (Fig. 4). These results conform to those of a previous study in which MyggA Natural (roll-on) was tested in the labo-ratory and field in 2004 (study I). Lemon eucalyptus contains PMD and was shown by (Trigg and Hill 1996) to provide protection against ticks and other arthropods. In a previous field study in Sweden the protective effect against I. ricinus of a lemon eucalyptus extract (Citriodiol a natural repellent against mosquitoes, stable flies, and midges) has been evaluated. The median num-ber of reported attached ticks per person decreased significantly if persons
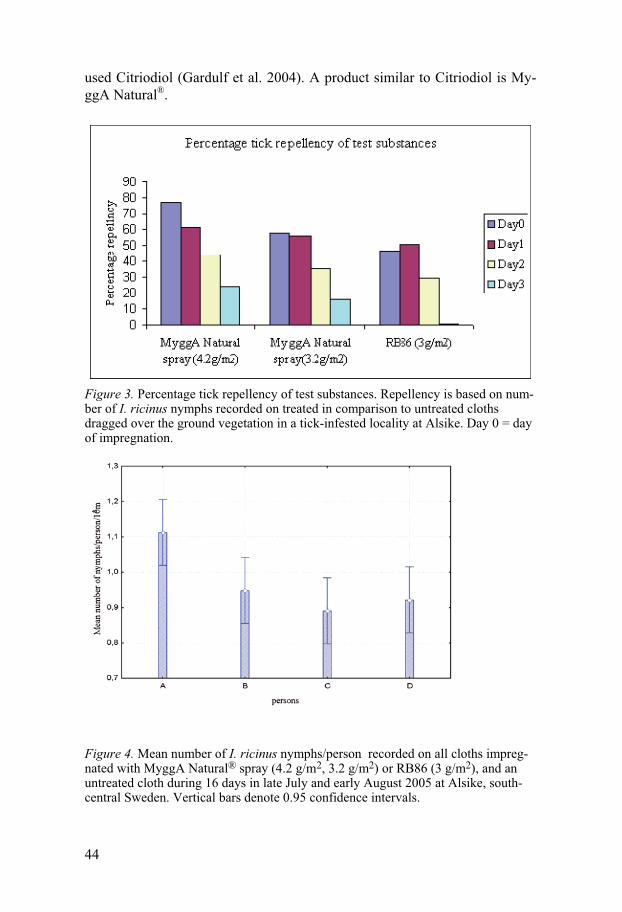
44
used Citriodiol (Gardulf et al. 2004). A product similar to Citriodiol is My-ggA Natural®.
Figure 3. Percentage tick repellency of test substances. Repellency is based on num-ber of I. ricinus nymphs recorded on treated in comparison to untreated cloths dragged over the ground vegetation in a tick-infested locality at Alsike. Day 0 = day of impregnation.
Figure 4. Mean number of I. ricinus nymphs/person recorded on all cloths impreg-nated with MyggA Natural® spray (4.2 g/m2, 3.2 g/m2) or RB86 (3 g/m2), and an untreated cloth during 16 days in late July and early August 2005 at Alsike, south-central Sweden. Vertical bars denote 0.95 confidence intervals.
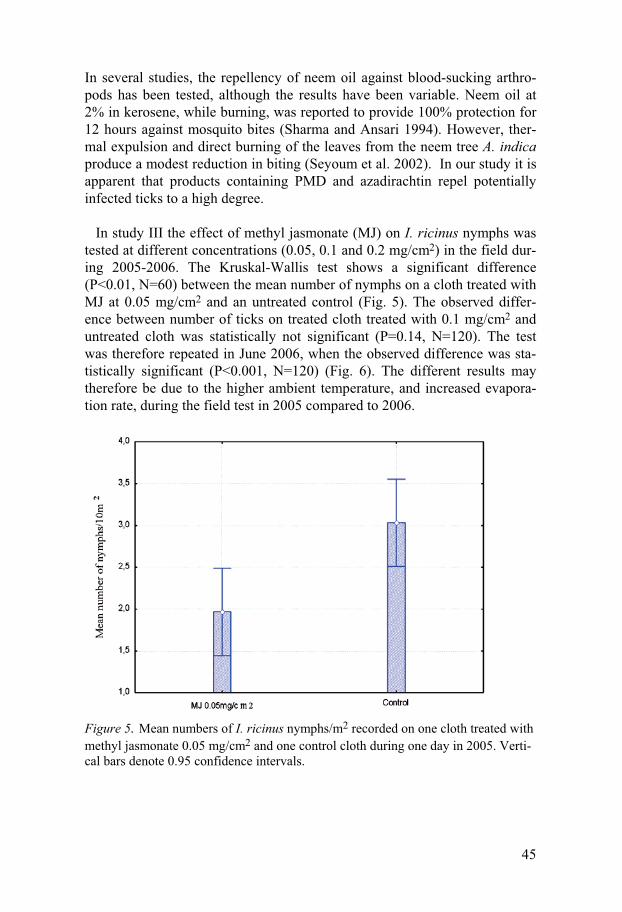
45
In several studies, the repellency of neem oil against blood-sucking arthro-pods has been tested, although the results have been variable. Neem oil at 2% in kerosene, while burning, was reported to provide 100% protection for 12 hours against mosquito bites (Sharma and Ansari 1994). However, ther-mal expulsion and direct burning of the leaves from the neem tree A. indica produce a modest reduction in biting (Seyoum et al. 2002). In our study it is apparent that products containing PMD and azadirachtin repel potentially infected ticks to a high degree.
In study III the effect of methyl jasmonate (MJ) on I. ricinus nymphs was tested at different concentrations (0.05, 0.1 and 0.2 mg/cm2) in the field dur-ing 2005-2006. The Kruskal-Wallis test shows a significant difference (P<0.01, N=60) between the mean number of nymphs on a cloth treated with MJ at 0.05 mg/cm2 and an untreated control (Fig. 5). The observed differ-ence between number of ticks on treated cloth treated with 0.1 mg/cm2 and untreated cloth was statistically not significant (P=0.14, N=120). The test was therefore repeated in June 2006, when the observed difference was sta-tistically significant (P<0.001, N=120) (Fig. 6). The different results may therefore be due to the higher ambient temperature, and increased evapora-tion rate, during the field test in 2005 compared to 2006.
Figure 5. Mean numbers of I. ricinus nymphs/m2 recorded on one cloth treated with methyl jasmonate 0.05 mg/cm2 and one control cloth during one day in 2005. Verti-cal bars denote 0.95 confidence intervals.
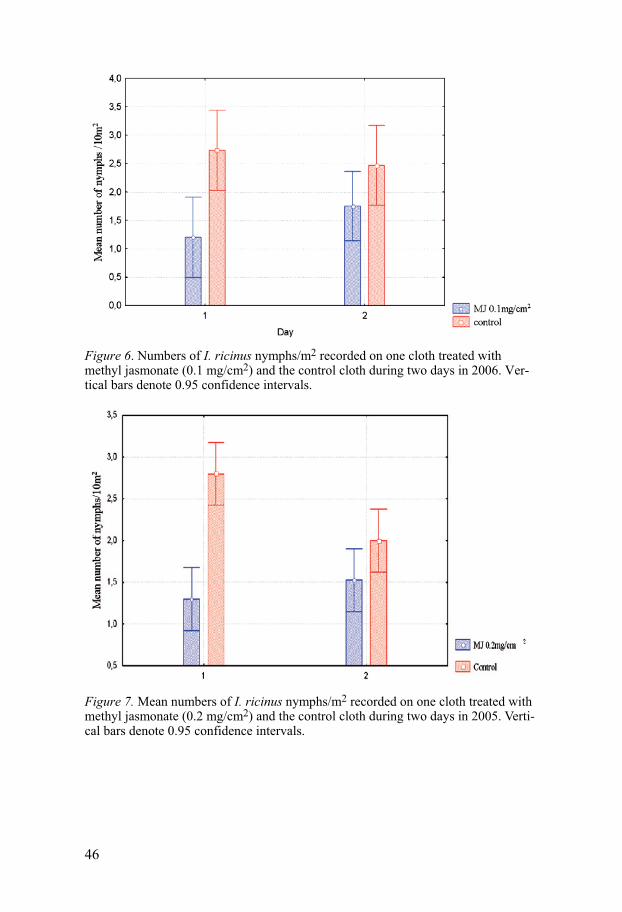
46
Figure 6. Numbers of I. ricinus nymphs/m2 recorded on one cloth treated with methyl jasmonate (0.1 mg/cm2) and the control cloth during two days in 2006. Ver-tical bars denote 0.95 confidence intervals.
Figure 7. Mean numbers of I. ricinus nymphs/m2 recorded on one cloth treated with methyl jasmonate (0.2 mg/cm2) and the control cloth during two days in 2005. Verti-cal bars denote 0.95 confidence intervals.

47
The number of nymphs recorded from the cloth treated with 0.2 mg/cm2 and the control cloth was significantly different (P<0.0001, N=120; Fig. 7).
From the above results, it was demonstrated that MJ has a repellent effect against blood-seeking I. ricinus nymphs in the field. Environmental factors such as temperature can affect the degree of protection. Many studies have demonstrated that MJ has a highly significant effect in the control of phyto-phagous pests (Birkett et al. 2000, Slesak et al. 2001, James 2005).

48
Concluding remarks
The data, which are presented in this thesis, suggest that certain natural product-based repellents and essential oils are able to repel Ixodes ricinus ticks at relatively low concentrations.
Products containing PMD or neem oil and products based on C. citrio-dora oil repel I. ricinus nymphs to a high degree.
Methyl jasmonate (MJ) which is a naturally occurring growth and devel-opment regulator in plants could also repel the tick I. ricinus.
The oils of L. angustifolia, P. graveolens and H. suayeolens from Laos and Guinea Bissau exhibited significant tick repellent activity on nymphal stage of I. ricinus. The essential oils from H. suaveolens leaves differ in composition which might be due to geographic origin and chemotype of the plants. Chemical modifications by oxidation and sulfidation of the main sesquiterpene hydrocarbons of H. suaveolens oil revealed strong tick repel-lency.
Medicinal plants such as S. persica, P. atlantica and J. phoenicea contain substances which have significant tick repellent activities for short duration. Several of these volatiles are listed as insect repellents, insecticides, acaricides, and/or pesticides.
However, these products or oils do not seem to provide 100% protection
against I. ricinus nymphs and will not provide the same degree of protection to all users against the ticks. Many factors can affect the protection afforded by the repellent such as environmental factors including a temperature.
There is a need for increased research into and development of natural repel-lents further so that they will provide a higher degree of protection and be more long-lasting. Hopefully, in the future plant-based repellents will pro-vide the public with natural alternatives to synthetic repellents, which can be applied to the skin, clothing, or both.

49
Sammanfattning (Swedish summary)
Växtbaserade Kemisha Ämnen som Fästingrepellenter Fästingar är globalt sett, näst efter stickmyggor, de viktigaste vektorerna för överföring av sjukdomar till människan. Varje fästingart som kan fungera som vektor har sina specifika miljökrav och sin specifika utbredning, som i sin tur avgör var riskområdena för olika fästingburna sjukdomar kommer att vara belägna. I tempererade områden på det norra halvklotet är den viktigas-te vektorburna sjukdomen borrelios (Lyme disease, LD). Den orsakas av bakterien Borrelia burgdorferi (Johnson) s.l. och överförs av fästingarter inom släktet Ixodes.
Personligt skydd genom användning av olika repellenter är en viktig strategi för att förebygga infektioner med vektorburna sjukdomar. Arbetet med att utveckla alternativ till de syntetiska repellenter som finns, inkluderar utvinning och utvärdering av växtprodukter för detta ändamål. Det är väl känt att växter producerar en mängd olika föreningar som fungerar som för-svar mot olika växtätare. Att utveckla och använda lokalt tillgängliga växt-produkter som fungerar som repellenter kan därför vara en alternativ strategi även för att förebygga sjukdomar som sprids via fästingar. Flera studier har genomförts för att utvärdera den repellerande effekten av ämnen från olika delar av växter. Vissa har visat sig ge ett bra skydd mot många blodsugande leddjur (Davis 1985, Gupta & Rutledge 1989, Malonza et al. 1992, Ansari & Razdan 1994, Barnard 1999, Barnard 2000, Jaenson et al. 2003). Produkter baserade på sådana ämnen kan ge ett billigt och effektivt skydd mot smittbä-rande leddjur, speciellt när andra åtgärder inte är genomförbara.
I vårt sökande efter växtprodukter med repellerande effekt mot fäs-tingar som ska kunna användas som alternativ till existerande produkter, har vi jämfört effekten av eteriska oljor från olika växter med det välkända med-let MyggA Natural (Bioglan, Lund, Sverige). Detta är en kommersiellt till-gänglig repellent mot blodsugande leddjur, som innehåller 30% olja från citroneukalyptus, Corymbia citriodora (Hook.) K.D.Hill & L.A.S. Johnson (Myrtaceae), en olja som består av minst 50% p-menthane-3, 8-diol (PMD). MyggA Natural innehåller också små mängder av eteriska oljor från laven-del, Lavandula angustifolia Mill. (Lamiaceae) och rosengeranium, Pelargo-nium graveolensL’Her. (Geraniaceae). Vid laboratorieundersökningar kun-de 100% repellens mot nymfer av Ixodes ricinus (L.) (Acari: Ixodidae) påvi-sas hos MyggA Natural och olja från C. citriodora. Lavendel och geranium-

50
olja i 1% lösning i 1,2-propandiol hade svag effekt mot nymfer av I. ricinus, men i 30% lösning var den repellerande effekten 100%. Enbart 1,2-propandiol (100%) hade ingen signifikant repellerande effekt. I fältstudier i fästingområden i Mellansverige jämfördes fästingrepellens av MyggA Natu-ral och C. citriodora-olja genom att preparerade filtar drogs genom naturlig vegetation upprepade gånger under en 6-dagarsperiod. Repellensen för dag 1 (74 och 85%) var av samma storleksordning (89%) som påvisats för 19% dietyl-metyl-bensamid vid ett tidigare försök. Effekten visade sig minska betydligt från dag 1 till dag 6 (från 74 till 45% repellens för MyggA Natural och från 85 till 42% repellens för C. citriodora-olja).
I områden i södra och mellersta Sverige testade vi genom en randomi-serad och standardiserad metod potentiell effekt mot fästingar av två kon-centrationer av MyggA Natural spray (innehållande PMD, 4,2 g/m2 och 3,2 g/m2) och en av RB86 (med 70% Neem-olja innehållande azadirachtin, 3 g/m2). Ämnena applicerades var för sig på 1 m2 stora dukar av bomullsfla-nell. De preparerade dukarna drogs genom vegetationen och antalet fästing-nymfer registrerades var 10:e meter. Antalet registrerade nymfer skilde sig kraftigt mellan de preparerade dukarna och en obehandlad kontroll (d.f. = 3, �2 = 112,74, p <0,0001). Antalet skilde sig också avsevärt mellan olika per-soner som samlade in data (d.f. = 3, �2 = 15,80, p <0,001). Repellens hos preparerade dukar, dvs. 4,2 g eller 3,2 g Mygga Natural spray/m2 och 3 g RB86/m2 visade sig minska från dag 0 (dvs dagen för impregnering) till dag 3 efter impregnering från respektive 77% till 24%, 58% till 16% och 47% till 0.5%. Denna studie tyder på att alla tre behandlingarna har betydande repel-lerande effekt mot nymfer av I. ricinus.
Vi undersökte effekten av den hos växter välkända signalsubstansen metyljasmonat (MJ) med hjälp av fästingnymfer av I. ricinus. I laboratorie-tester visade sig tygstycken preparerade med MJ i fyra olika koncentrationer, 0,075, 0,15, 0,30 och 0,75 mg/cm2, ge ökad repellens mot nymferna: respek-tive 57%, 71%, 92% och 99% av nymferna klamrade sig inte fast vid tyg-stycket. Repellens av MJ undersöktes också i fästingområden i Mellansveri-ge. Tygstycken av bomullsflanell, preparerade med 0.05, 0.1 eller 0.2 mg/cm2 MJ upplöst i aceton, drogs över marken genom vegetationen. Anta-let nymfer på de preparerade tygstyckena var avsevärt lägre än på en obe-handlad kontroll. Således har MJ i de testade koncentrationerna en betydan-de repellens mot nymfer av I. ricinus.
Beståndsdelar i färska och torkade blad och i eterisk olja från Hyptis suaveolens (L.) Poit (Lamiaceae) från Laos och Guinea-Bissau identifierades genom SPME-GC-MS. Två huvudsakliga sesquiterpen-kolväten i oljan var �-caryophyllen och humulen. De testades och modifierades genom oxidation och sulfidering för att få fram fästingrepellerande föreningar med lägre flyk-tighet. De oljor som framställts genom ångdestillation av blad av H. suaveo-lens från både Guinea-Bissau och Laos som odlats i Sverige, liksom från plantor naturligt förekommande H. suaveolns i Laos, visade sig ha en avse-

51
värd repellens mot I. ricinus. Vid en koncentration av 10% utspädd i aceton hade Hyptis-olja från Guinea-Bissau den starkaste repellerande effekten (93%), medan effekten hos olja i Hyptis från Laos, odlad och destillerad i Sverige, var 81%, och från Hyptis, naturligt förekommande och destillerad i Laos, var 83%. När olika substanser av olja från H. suaveolens jämfördes hade caryophyllenoxid den starkaste repellensen. De näst mest effektiva substanserna visade sig vara humulenoxid, 97%, humulen, 96,8%, och hu-mulen-6,7-episulfid 96,7% repellens. Ämnena caryophyllensulfid, menthol-sulfid, humulen-9,10-episulfid, germacren D och caryophyllen uppvisade repellens på 84,3%, 84,3%, 81,2%, 78,3 % and 68,9%, respektive.
En signifikant repellens mot nymfer av I. ricinus påvisades också hos eteriska oljor från Pistacia atlantica (Desf.), Juniperus phoenicia (L.) och Salvadora persica (Wall). Effekten hos dessa oljor var dock kortvarig. Dessa tre arter hade efter 20 minuter en repellens på 91, 81 respektive 52%, men efter en timme hade den sjunkit till 16, 10, respektive 15,5%. Dessa tre arter är välkända och mycket använda inom traditionell medicin i Libyen. De viktigaste monoterpena kolväten som kunde identifieras genom SPME-GC-MS i oljor från dessa tre växtarter var, i olika proportioner, �-pinen, sabinen, �-pinen, myrcen, �-phellandren, 4-karen, �-phellandren och -terpinen. De olika enantiomererna av varje kiral monoterpen särskildes ej i denna studie. Flera av dessa flyktiga ämnen är kända som insektsrepellenter, insekticider, acaricider och/eller andra bekämpningsmedel (Duke 2004).
Slutsatser De resultat som presenteras i denna avhandling visar följande:
Vissa naturprodukter och eteriska oljor kan repellera fästingar av arten Ix-odes ricinus även vid relativt låga koncentrationer.
Många potentiella naturliga repellenter fungerar genom att avskräcka fästingar från att klamra sig fast eller tränga in i huden. Dessa produkter eller oljor ger inte 100% skydd mot nymfer av I. ricinus.
Det finns behov av ökad forskning kring och utveckling av nya repelle-rande substanser.
Förhoppningsvis kommer i framtiden växtbaserade repellenter, som kan
appliceras på huden och/eller kläderna, att finnas tillgängliga för allmänhe-ten som ett alternativ till syntetiska repellenter.

52
Acknowledgements
Writing a thesis is supposedly a lonely pursuit. This makes me realise how fortunate I have been to be surrounded by people who have helped me by sharing my happiness when things have gone well and by offering advice and unending support when things have not. Even though I will not be able to thank all of those who deserve it in the space allotted to me here, please know that you all have my heartfelt thanks.
First and foremost, I would like to thank my supervisor, Professor Thomas Jaenson, who has patiently read and re-read various versions of my text and checked the language and for his encouragement and invaluable support. I would also like to thank my assistant supervisor, Dr. Katinka Påls-son for constantly trying very hard to help me, her friendship and support and for answering all my questions. My sincere thanks go to professor Mats Thulin for reading the summary in Swedish and for correcteing and checking the language. Thanks also go to Professor Anna-Karin Borg-Karlson for fruitful collaboration. I would like to thank Janet Ahlberg for checking the English in my thesis
,s summary.
Many thanks go to the Libyan Government, represented by the Higher Education Section, for the scholarships. Thanks also to Dr. Omar Elsodany for having given me the opportunity to continue the postgraduate studies. I would like to thank the People’s Bureau of Libya for their always welcome to help.
I am also very grateful to my friends, Fawzia El-Mahalli has been a great office mate and a true friend, and always there to share good and bad, and any biscuits she had. Ibtesam El-Kalaf, Amal Elfaitouri, Hanan Sharif, Najat Ratabe, Shaima Mahjoubi, Wafa, Abeer, Eman for your company has made me feel less homesick and without your friendship, my time as a PhD student would not have been as interesting and stimulating as it was. Many thanks go to Asma El-Shebani for introducing me to the key people in order to obtain academic acceptance and for helping me to know every thing in Sweden. Thanks to all my friends at Arab Medical Unversity.
To my present (and former) fellow students and post-docs at the de-partment of Systematic Biology: Andreas, Anders, Anja, Anneleen, Catarina, Karolina, Cajsa, Christina, Carlota, Hugo, Donatha, Ding, Chanda, II-Chan, Henrik, Johanna, Mattias, Sanja, Sunniva, Gemma, Annika, Björn-Axel, Bozo, Elisabeth, Frida, Hege, Maria, Per E., Per K., Rikke, Sylvain, Yonas,

53
Åsa, John, Pravech and Vichith thanks a lot for your interesting personali-ties, comments, discussions and smiles.
Afsaneh and Nahida you have been of great help, thank you for your sup-port and asking about me and my children. The administrative staff members at the department,Ulla Hedenquist and Elisabeth Bergström have been a great help to me during my years here, and help regarding computers I got from Stefan and Håkan; thanks. I would also like to thank Emma Jaenson and David Jaenson for their assistance in field work and for sending cards from the countries they visited.
On a more personal note, I would like to thank my wonderful mother Fathia, to whom this dissertation is dedicated, without her help, support, and never-ending faith in me this dissertation would never have been completed, or even begun. I would also like to thank my borthers Ibrahim, Ahmed, Os-sama, and Mohamed, and my sisters Eman and Kamila for support, encour-agement and unfailing love. For always being there to help.
I have been privileged to be surrounded by my husband’s family whose encouragement has always been forthcoming: My parents in law Mussa and Fatma, my brothers in law Jawud, Salem, especially Mohamed and Ali who helped me to collect the plants from Libya, my sisters in law Mahmola who-has been a great support to me, Saaida, Fathia, Antesar, Abrika, Fawzia, Samira, Asma and their respective families.
Last, but most, I want to thank my husband Jumaa for keeping my feet on the ground, my nervousness at bay and my future looking bright. This disser-tation would not have come into being without his assistance. To my fabu-lous children, Maram, Rouaa and Moussa deserve the highest praise for their patience during many moments of absence and for their support in various ways.
Above all, I thank my God Allah for allowing me to successfully com-plete my study.
Samira, 2008

54
References
Abdel-Shafy, S., and A. A. Zayed. 2002. In vitro acaricidal effect of plant extract of neem seed oil (Azadirachta indica) on egg, immature, and adult stages of Hyalomma anatolicum excavatum (Ixodoidea: Ixodi-dae). Vet. Parasitol. 106: 89-96.
Alali, F., and T. Al-Lafi. 2003. GC-MS analysis and bioactivity testing of the volatile oil from the leaves of the toothbrush tree Salvadora per-sica L. Nat. Prod. Res. 17: 189-94.
Angioni, A., A. Barra, M. T. Russo, V. Coroneo, S. Dessi, and P. Cabras. 2003. Chemical composition of the essential oils of Juniperus from ripe and unripe berries and leaves and their antimicrobial activity. J. Agric. Food Chem. 51: 3073-8.
Ansari, M. A., and R. K. Razdan. 1994. Repellent action of Cymbopogon martinii martinii Stapf var. sofia oil against mosquitoes. Indian J. Malariol. 31: 95-102.
Ashitani, T., and S. Nagahama. 1999. Direct episulfidation of caryo-phyllene and humulene. Nat. Prod. Letter 13: 163-167.
Assenga, S. P., M. You, C. H. Shy, J. Yamagishi, T. Sakaguchi, J. Zhou, M. K. Kibe, X. Xuan, and K. Fujisaki. 2006. The use of a recom-binant baculovirus expressing a chitinase from the hard tick Hae-maphysalis longicornis and its potential application as a bioacaricide for tick control. Parasitol. Res. 98: 111-8.
Azevedo, N. R., I. F. Campos, H. D. Ferreira, T. A. Portes, S. C. Santos, J. C. Seraphin, J. R. Paula, and P. H. Ferri. 2001. Chemical vari-ability in the essential oil of Hyptis suaveolens. Phytochemistry 57: 733-6.
Azevedo, N. R., I. F. P. Campos, H. D. Ferreira, T. A. Portes, J. C. Seraphin, J. Realino de Paula, S. C. Santos, and P. H. Ferri. 2002. Essential oil chemotypes in Hyptis suaveolens from Brazilian Cerrado. Biochem. Syst. Ecol. 30: 205–216.
Bader, A., G. Flamini, P. L. Cioni, and I. Morelli. 2002. The composition of the root oil of Salvadora persica L. J. Essent. Oil Res. 14: 128-129.
Badolo, A., E. Ilboudo-Sanogo, A. P. Ouedraogo, and C. Costantini. 2004. Evaluation of the sensitivity of Aedes aegypti and Anopheles gambiae complex mosquitoes to two insect repellents: DEET and KBR 3023. Trop. Med. Int. Health 9: 330-4.
Balandrin, M. F., J. A. Klocke, E. S. Wurtele, and W. H. Bollinger. 1985. Natural plant chemicals: sources of industrial and medicinal materi-als. Science 228: 1154-60.

55
Baranton, G., D. Postic, I. Saint Girons, P. Boerlin, J. C. Piffaretti, M. Assous, and P. A. Grimont. 1992. Delineation of Borrelia burgdor-feri sensu stricto, Borrelia garinii sp. nov., and group VS 461 asso-ciated with Lyme borreliosis. Int. J. Syst. Bacteriol. 42: 378-83.
Barnard, D. R. 1999. Repellency of essential oils to mosquitoes (Diptera: Culicidae). J. Med. Entomol. 36: 625-9.
Barnard, D. R. 2000. Repellents and toxicants for personal protection. WHO/CDS/WHOPES/GCDPP. 48 pp.
Barrero, A. F., M. M. Herrador, J. F. Arteaga, M. Akssira, F. Mellouki, A. Belgarrabe, and M. A. Blazquez. 2005. Chemical composition of the essential oils of Pistacia atlantica Desf. J. Essent. Oil Res 17: 52-54.
Batanouny, K. H., S. Abou Tabl, M. Shabana, and F. Soliman. 1999. Wild medicinal plants in Egypt. An inventory to support conserva-tion and sustainable use. With Support of the Swiss Development Co-operation (SDC). Academy of Scientific Research and Technol-ogy. Egypt International Union for Conservation (IUCN).
Benjamin, M. A., E. Zhioua, and R. S. Ostfeld. 2002. Laboratory and field evaluation of the entomopathogenic fungus Metarhizium anisopliae (Deuteromycetes) for controlling questing adult Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 39: 723-8.
Birkett, M. A., C. A. Campbell, K. Chamberlain, E. Guerrieri, A. J. Hick, J. L. Martin, M. Matthes, J. A. Napier, J. Pettersson, J. A. Pickett, G. M. Poppy, E. M. Pow, B. J. Pye, L. E. Smart, G. H. Wadhams, L. J. Wadhams, and C. M. Woodcock. 2000. New roles for cis-jasmone as an insect semiochemical and in plant de-fense. Proc. Natl. Acad. Sci. U. S. A. 97: 9329-34.
Browne, L. B. 1977. Host related responses and their suppression some bahavioural considerations. In chemical control of insect behaviour. John Wiley & Sons,New York: 117-127.
Bruin, J., and M. Dicke. 2001. Chemical information transfer between wounded and unwounded plants: backing up the future. Biochem. Syst. Ecol. 29: 1103-1113.
Carroll, J. F. 1995. Laboratory evaluation of predatory capabilities of a common wolf spider (Araneae: Lycosidae) against two species of ticks (Acari: Ixodidae). Proc. Entomol.soc. 97: 746-49.
Carroll, J. F., V. B. Solberg, J. A. Klun, M. Kramer, and M. Debboun. 2004. Comparative activity of deet and AI3-37220 repellents against the ticks Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) in laboratory bioassays. J. Med. Entomol. 41: 249-54.
Cavanagh, H. M., and J. M. Wilkinson. 2002. Biological activities of lav-ender essential oil. Phytother Res. 16: 301-8.
Cole, M. M. 1965. Biological control of ticks by the use of hymenopterous parasites a review. World Health Org. Publ. WHO/ebl/43 65:1-12.
Creelman, R., M. Tierney, and J. Mullet. 1992. Jasmonic acid/methyl jasmonate accumulate in wounded Soybean Hypocotyls and Modu-late wound gene expression. Nat. Acad. Sci. 89: 4938-4941.

56
Crippa, M., O. Rais, and L. Gern. 2002. Investigations on the mode and dynamics of transmission and infectivity of Borrelia burgdorferi sensu stricto and Borrelia afzelii in Ixodes ricinus ticks. Vector Borne Zoonotic Dis. 2: 3-9.
Curtis, C. F., J. D. Lines, B. Lu, and A. Renz. 1991. Natural and synthetic repellents 75-92. C. F. Curtis: Control of disease vectors in the community. Wolfe, London.
Curtis, C. F., J. D. Lines, J. Ijumba, A. Callaghan, N. Hill, and M. A. Karimzad. 1987. The relative efficacy of repellents against mos-quito vectors of disease. Med. Vet. Entomol. 1: 109-19.
Davis, E. E. 1985. Insect repellents: concepts of their mode of action rela-tive to potential sensory mechanisms in mosquitoes (Diptera: Culi-cidae). J. Med. Entomol. 22: 237-43.
Delazar, A., R. G. Reid, and S. D. Sarker. 2004. GC-MS analysis of the essential oil from the oleoresin of Pistacia atlantica var. Mutica. Chem. Nat. Compd. 40: 24-27.
Duke, J. A. 2004b. Dr. Duke's Phytochemical and Ethnobotanical Database. Agricultural Research Service: http://www.ars-grin.gov/duke/ (ac-cessed Sept. 2008).
El-Gadi, A., and S. Bshana. 1997. Usages of some plants in Libyan folk-medicine. Publication. Zaouia, Libya (Arabic document) Vol 1: 121pp.
El-Gadi, A., and M. El-Mogrebi. 2003. Usages of some plants in Libyan folk-medicine. Publication. Zaouia, Libya (Arabic document) Vol 3: 104pp.
Fleetwood, S. C., P. D. Teel, and G. Thompson. 1984. Impact of imported fire ants on lone star tick mortality in open and canopied pasture habitats of east central Texas. South-west Entomol. 9: 158-162.
Francisco, M. C., C. cruz-vázquez, M. F. Ruvalcaba, J. Molina-Torres, J. S. Cruz, and M. R. Parra. 2003. Repellence of Boophilus mi-croplus larvae in Stylosanthes humilis and Stylosanthes hamata plants. Parasitol. Latinoam. 58: 118-121.
Gardulf, A., I. Wohlfart, and R. Gustafson. 2004. A prospective cross-over field trial shows protection of lemon eucalyptus extract against tick bites. J. Med. Entomol. 41: 1064-7.
Gern, L., and R. C. Falco. 2000. Lyme disease. Rev. Sci. Tech. 19: 121-35. Gray, J. S. 1991. The development and seasonal activity of the tick Ixodes
ricinus: a vector of Lyme borreliosis. Rev. Med. Vet. Entomol.: 323-39.
Gupta, R. K., and L. C. Rutledge. 1989. Laboratory evaluation of con-trolled-release repellent formulations on human volunteers under three climatic regimens. J. Am. Mosq. Control Assoc. 5: 52-55.
Gupta, R. K., and L. C. Rutledge. 1994. Role of repellents in vector con-trol and disease prevention. Am. J. Trop. Med. Hyg. 50: 82-86.
Honzakova, E., J. Olejnicek, V. Cerny, M. Daniel, and F. Dusbabek. 1975. Relationship between numbers of eggs deposited and body

57
weight of engorged Ixodes ricinus female. Folia. Parasitol. Prague. 22: 37-43.
Ibrahim, M. A., A. Nissinen, and J. K. Holopainen. 2005. Response of Plutella xylostella and its parasitoid Cotesia plutellae to volatile compounds. J. Chem. Ecol. 31: 1969-84.
ILADS. 2006. Basic Information about Lyme disease. International Lyme and Associated Diseases Society: http://www.ilads.org/basic.html (accessed 2008).
Isman, M. B., O. Koul, A. Luczynski, and J. Kaminski. 1990. Insecticidal and antifeedant bioactive of neem oils and their relationship to azadirachtin content. J. Agric. Food Chem. 38: 1406-1411.
Jaenson, T. G. 1991. The epidemiology of Lyme borreliosis. Parasitol. To-day. 7: 39-45.
Jaenson, T. G., K. Palsson, and A. K. Borg-Karlson. 2005a. Evaluation of extracts and oils of tick-repellent plants from Sweden. Med. Vet. En-tomol. 19: 345-52.
Jaenson, T. G., K. Palsson, and A. K. Borg-Karlson. 2006. Evaluation of extracts and oils of mosquito (Diptera: Culicidae) repellent plants from Sweden and Guinea-Bissau. J. Med. Entomol. 43: 113-9.
Jaenson, T. G., L. Talleklint, L. Lundqvist, B. Olsen, J. Chirico, and H. Mejlon. 1994. Geographical distribution, host associations, and vec-tor roles of ticks (Acari: Ixodidae, Argasidae) in Sweden. J. Med. Entomol. 31: 240-56.
Jaenson, T. G. T. 1999. Fästing en Ixodes ricinus som sjukdomsöverförare i Skandinavien (the tick Ixodes ricinus as a vector of diseases in Scandinavia). Växtskyddsnotiser 63: 53-59.
Jaenson, T. G. T., A. Lindstrom, and K. Palsson. 2003. Repellency of the mosquito repellent MyggA (N, N-diethyl-3-methyl-benzamide) to the common ticks Ixodes ricinus (L.) (Acari: Ixodidae) in the labora-tory and field. Entomologisk Tidskrift: 245-51.
Jaenson, T. G. T., S. Garboui, and K. Palsson. 2005b. Repellency of lemon eucalyptus (Corymbia citriodora) oil and the mosquito repel-lent MyggA Natural® to the common tick Ixodes ricinus (L.) (Acari: Ixodidae) in the laboratory and field. J. Med. Entomol. 43: 731-736.
James, D. G. 2005. Further field evaluation of synthetic herbivore-induced plant volatiles as attractants for beneficial insects. J. Chem. Ecol. 31: 481-95.
Kalsbeek, V., F. Frandsen, and T. Steenberg. 1995. Entomopathogenic fungi associated with Ixodes ricinus ticks. Exp. Appl. Acarol. 19: 45-51.
Kayaa, G. P. 2000. The potential for anti-tick plants as components of an integrated tick control strategy. Ann. New York Acad. Sci. 916: 576-582.
Kettle, D. S. 1995. Medical and veterinary entomology. CAB International, UK. The University Press, Cambridge.
Khalil, A. T. 2006. Benzylamides from Salvadora persica. Arch. Pharm. Res. 29: 952-6.

58
Klun, J. A., D. Strickman, E. Rowton, J. Williams, M. Kramer, D. Rob-erts, and M. Debboun. 2004. Comparative resistance of Anopheles albimanus and Aedes aegypti to N, N-Diethyl-3-methylbenzamide (Deet) and 2-Methylpiperidinyl-3-cyclohexen-1-carboxamide (AI3-37220) in laboratory human-volunteer repellent assays. J. Med. En-tomol.: 418–422.
Kulkarni , R. N., G. R. Mallavarapu, K. Baskaran, S. Ramesh, and S. Kumar. 1998. Composition of the essential oils of two isomen-thone-rich variants of geranium (Pelargonium sp.). Flavour Fragr. J. 13: 389-392.
Lefcort, H., and L. A. Durden. 1996. The effect of infection with Lyme disease spirochetes (Borrelia burgdorferi) on the phototaxis, activ-ity, and questing height of the tick vector Ixodes scapularis. Parasi-tology. 113: 97-103.
Lindgren, E., L. Talleklint, and T. Polfeldt. 2000. Impact of climatic change on the northern latitude limit and population density of the disease-transmitting European tick Ixodes ricinus. Environ. Health Perspect. 108: 119-23.
Lis-Balchin, M. 1995. The chemistry and bioactivity of essential oils. Roch-ester, UK: Amber Wood Publishing Lted.
Lwande, W., A. J. Ndakala, A. Hassanali, L. Moreka, E. Nyandat, M. Ndungu, H. Amiani, P. M. Gitu, M. M. Malonza, and D. K. Punyua. 1998. Gynandropsis gynandra essential oil and it constitu-ents as tick (Rhipicephalus appendiculatus) repellents. Phytochemis-try: 401–405.
Malonza, M. M., O. O. Dipeolu, A. O. Amoo, and S. M. Hassan. 1992. Laboratory and field observations on anti-tick properties of the plant Gynandropsis gynandra (L.) Briq. Vet. Parasitol. 42: 123-36.
Mecherara-Idjeri, S., A. Hassani, V. Castola, and J. Casanova. 2008. Composition of leaf, fruit and gall essential oils of Algerian Pistacia atlantica. Desf. J. Essent. Oil Res. 20: 215-219.
Mejlon, H. A., and T. G. Jaenson. 1993. Seasonal prevalence of Borrelia burgdorferi in Ixodes ricinus in different vegetation types in Swe-den. Scand. J. Infect. Dis. 25: 449-56.
Mulla, M. S., and T. Su. 1999. Activity and biological effects of neem products against arthropods of medical and veterinary importance. J. Am. Mosq. Control Assoc. 15: 133-52.
Mwangi, E., O. Dipeolu, R. Newson, G. Kaaya and S. Hassan. 1991a. Predators, parasitoids and pathogens of ticks: A review. Bio-control Sci. Technol. 1: 147-156.
Mwangi, E. N., R. M. Newson, and G. P. Kaaya. 1991b. Predation of free-living engorged female Rhipicephalus appendiculatus. Exp. Appl. Acarol. 12: 153-62.
Ndumu, P. A., J. B. George, and M. K. Choudhury. 1999. Toxicity of neem seed oil (Azadiracta indica) against the larvae of Amblyomma variegatum a three-host tick in cattle. Phytother Res. 13: 532-4.

59
Ndungu, M., W. Lwande, A. Hassanali, L. Moreka, and S. C. Chhabra. 1995b. Cleome monophylla essential oil and its constituents as tick (Rhipicephalus appendiculatus) and maize weevil (Sitophilus zea-mais) repellents. Entomol. Exp. Appl. 76: 217-22.
Nentwig, G., J. Boeckh, F. P. Hoever, B. W. Krüger, and K. Röder. 2002. Bayrepel® (KBR 3023), a new mosquito repellent: from labo-ratory synthesis to a worldwide commercial product. Proceedings II European Conference on Travel Medicine, Florence: p. 85.
Nilsson, K., O. Lindquist, A. J. Liu, T. G. Jaenson, G. Friman, and C. Pahlson. 1999. Rickettsia helvetica in Ixodes ricinus ticks in Swe-den. J. Clin. Microbiol. 37: 400-3.
Nilsson, K., T. G. Jaenson, I. Uhnoo, O. Lindquist, B. Pettersson, M. Uhlen, G. Friman, and C. Pahlson. 1997. Characterization of a spotted fever group Rickettsia from Ixodes ricinus ticks in Sweden. J. Clin. Microbiol. 35: 243-7.
Oladimeji, F. A., O. O. Orafidiya, T. A. Ogunniyi, and T. A. Adewunmi. 2000. Pediculocidal and scabicidal properties of Lippia multiflora essential oil. J. Ethnopharmacol. 72: 305-11.
Palsson, K., and T. G. T. Jaenson. 1999a. Plant products used as mosquito repellents in Guinea Bissau, West Africa. Acta Tropica. 72: 39-52.
Palsson, K., and T. G. T. Jaenson. 1999b. Comparison of plant products and pyrethroid-treated bed nets for protection against mosquitoes (Diptera: Culicidae) in Guinea Bissau, West Africa. J. Med. Ento-mol. 36: 144-148.
Panella, N. A., J. Karchesy, G. O. Maupin, J. C. Malan, and J. Piesman. 1997. Susceptibility of immature Ixodes scapularis (Acari:Ixodidae) to plant-derived acaricides. J. Med. Entomol. 34: 340-5.
Park, I.-K., S.-G. Lee, D.-H. Choi, J.-D. Park, and Y.-J. Ahn. 2003. In-secticidal activities of constituents identified in the essential oil from leaves of Chamaecyparis obtusa against Callosobruchus chinensis (L.) and Sitophilus oryzae (L). J. Stored Prod. Res. 39: 375–384.
Piesman, J., and L. Gern. 2004. Lyme borreliosis in Europe and North America. Parasitology 129 Suppl: S191-220.
Pretorius, A. M., M. Jensenius, F. Clarke, and S. H. Ringertz. 2003. Re-pellent efficacy of DEET and KBR 3023 against Amblyomma hebraeum (Acari: Ixodidae). J. Med. Entomol. 40: 245-8.
Rao, B. R., P. N. Kaul, K. V. Syamasundar, and S. Ramesh. 2003. Com-parative composition of decanted and recovered essential oils of Eucalyptus citriodora Hook. Flavour Fragr. J. 18: 133 -135.
Raoult, D., and V. Roux. 1997. Rickettsioses as paradigms of new or emerging infectious diseases. Clin. Microbiol. Rev. 10: 694 -719.
Rezzi, S., C. Cavaleiro, A. Bighelli, L. Salgueiro, A. P. da Cunha, and J. Casanova. 2001. Intraspecific chemical variability of the leaf essen-tial oil of Juniperus phoenicea subsp. turbinata from Corsica. Bio-chem. Syst. Ecol. 29: 179-188.
Rivas, R., D. Valera, J. L. Avila, L. Aubert, M. E. Alonso-Amelot, L. B. Rojas, and A. Usubillaga. 2002. The essential oil of Hyptis

60
suaveolens (L.) Poit. and its insect deterrent properties. J. Essent. Oil-Bearing Plants 5: 126–131.
Rutledge, L. C., M. A. Moussa, C. A. Lowe, and R. K. Sofield. 1978. Comparative sensitivity of mosquito species and strains to the repel-lent diethyl toluamide. J. Med. Entomol. 14: 536-41.
Rutledge, L. C., D. M. Collister, V. E. Meixsell, and G. H. Eisenberg. 1983. Comparative sensitivity of representative mosquitoes (Diptera: Culicidae) to repellents. J. Med. Entomol. 20: 506-10.
Samish, M., and J. Rehacek. 1999. Pathogens and predators of ticks and their potential in biological control. Ann. Rev. Entomol. 44: 159-182.
Schmutterer, H. 1990. Properties and potential of natural pesticides from the neem tree, Azadirachta indica. Ann. Rev. Entomol. 35: 271-97.
Schreck, C. E., and B. A. Leonhardt. 1991. Efficacy assessment of Qu-wenling, a mosquito repellent from China. J. Am. Mosq. Control Assoc. 7: 433-6.
Schreck, C. E., D. Fish, and T. P. McGovern. 1995. Activity of repellents applied to skin for protection against Amblyomma americanum and Ixodes scapularis ticks (Acari: Ixodidae). J. Am. Mosq. Control Assoc. 11: 136-40.
Seyoum, A., K. Palsson, S. Kung'a, E. W. Kabiru, W. Lwande, G. F. Killeen, A. Hassanali, and B. G. Knols. 2002. Traditional use of mosquito-repellent plants in western Kenya and their evaluation in semi-field experimental huts against Anopheles gambiae: ethno-botanical studies and application by thermal expulsion and direct burning. Trans. R. Soc. Trop. Med. Hyg. 96: 225-31.
Sharma, V. P., and M. A. Ansari. 1994. Personal protection from mosqui-toes (Diptera: Culicidae) by burning neem oil in kerosene. J. Med. Entomol. 31: 505-7.
SIIDC. 2007. Swedish Institute for Infectious Disease Control/ Smi- ttskyddsinstitutet/SMI), http://www.smittskyddsinstitutet.se/smi (accessed 2008)
Slesak, E., M. Slesak, and B. Gabrys. 2001. Effect of methyl jasmonate on hydroxamic acid content, protease activity, and bird cherry-oat aphid Rhopalosiphum padi (L.) probing behavior. J. Chem. Ecol. 27: 2529-43.
Solberg, V. B., T. A. Klein, K. R. McPherson, B. A. Bradford, J. R. Burge, and R. A. Wirtz. 1995. Field evaluation of deet and a piperidine repellent (AI3-37220) against Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 32: 870-875.
Sonenshine, D. E. 1993. Biology of ticks, vol. 2: Oxford University Press. Oxford University Press 2: 465.
Stafford, K. C., 3rd, A. J. Denicola, and L. A. Magnarelli. 1996. Presence of Ixodiphagus hookeri (Hymenoptera: Encyrtidae) in two Connecti-cut populations of Ixodes scapularis (Acari: Ixodidae). J. Med. En-tomol. 33: 183-8.

61
Stassi, V., A. Verykokidou, and A. Loukis. 1996. The antimicrobial activ-ity of the essential oils of four Juniperus species growing wild in Greece. Flav. Fragr. J. 11: 71–74.
Staub, D., M. Debrunner, L. Amsler, and R. Steffen. 2002. Effectiveness of a repellent containing DEET and EBAAP for preventing tick bites. Wilderness Environ. Med. 13: 12-20.
Sutherst, R., L. J. Wilson, and I. M. Cook. 2000. Predation of the cattle tick, Boophilus microplus (Canestrini) (Acarina: Ixodidae), in three Australian pastures. Austr. J. Entomol. 39: 70-77.
Thavara, U., A. Tawatsin, J. Chompoosri, W. Suwonkerd, U. Chansang, and P. Asavadachanukorn. 2001. Laboratory and field evaluations of the insect repellent 3535 (Ethyl Butylacetylaminopropionate) and Deet against mosquito vectors in Thailand. J. Am. Mosq. Control Assoc. 17: 190-195.
Thompson, K. C., E. J. Roa, and N. T. Romero. 1978. Anti-tick grasses as the basis for developing practical tropical tick control packages. Trop. Anim. Health Prod. 10: 179-82.
Thorsell, W., A. Mikiver, and H. Tunon. 2006. Repelling properties of some plant materials on the tick Ixodes ricinus L. Phytomedicine. 13: 132-4.
Trigg, J. K. 1996. Evaluation of a eucalyptus-based repellent against Anopheles spp. in Tanzania. J. Am. Mosq. Control Assoc. 12: 243-6.
Trigg, J. K., and N. Hill. 1996. Laboratory evalution of an eucalyptus- based repellent against four biting arthropods. Phytother. Res.10: 313-316.
Tzakou, O., I. Bazos, and A. Yannitsaros. 2007. Volatile metabolites of Pistacia atlantica Desf. from Greece. Flavour Fragr. J. 22: 358-362. Vick, B. A., and D. C. Zimmerman. 1984. Biosynthesis of jasmonic acid
by several plant species. Plant Physiol. 75: 458-461. Wulff , R. 1973. Intrapopulational variation in the germination of seeds in
Hyptis suaveolens. Ecology 54: 646–649. Yatagai, M., H. Makihara, and K. Oba. 2002. Volatile components of
Japanese cedar cultivars as repellents related to resistance to Cryp-tomeria bark borer. J. Wood Sci. 48: 51–55.

Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 574
Editor: The Dean of the Faculty of Science and Technology
A doctoral dissertation from the Faculty of Science andTechnology, Uppsala University, is usually a summary of anumber of papers. A few copies of the complete dissertationare kept at major Swedish research libraries, while thesummary alone is distributed internationally through theseries Digital Comprehensive Summaries of UppsalaDissertations from the Faculty of Science and Technology.(Prior to January, 2005, the series was published under thetitle “Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-9368
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2008