placental changes associated with fetal outcome in the plasmodium
TRANSCRIPT

158
Am. J. Trop. Med. Hyg., 63(3, 4), 2000, pp. 158–173Copyright� 2000 by The American Society of Tropical Medicine and Hygiene
PLACENTAL CHANGES ASSOCIATED WITH FETAL OUTCOME IN THEPLASMODIUM COATNEYI/RHESUS MONKEY MODEL OF MALARIA IN PREGNANCY
BILLIE B. DAVISON, FRANK B. COGSWELL, GARY B. BASKIN, KATHRINE P. FALKENSTEIN,ELIZABETH W. HENSON,AND DONALD J. KROGSTAD
Departments of Pathology, Parasitology, and Veterinary Science, Tulane Regional Primate Research Center, Covington,Louisiana; Tulane School of Public Health and Tropical Medicine, Tulane University Medical Center, New Orleans, Louisiana
Abstract. Term placentas collected surgically from sevenPlasmodium coatneyi–infected rhesus monkeys, oneabortion, and five controls were evaluated histopathologically. The placentas fromPlasmodium-infected dams hadmore significant pathologic changes than those from controls for six parameters (P � 0.05) and higher numbers ofactivated (LN5� Zymed) macrophages in the intervillous space (IVS) (P � 0.0173). Total parasite load (TPL) wasdefined as the sum of all weekly peripheral infected red blood cell counts for each trimester and for the entirepregnancy. High first trimester PLs were more likely to result in fetal demise (P � 0.0476) or increased placentaldamage in surviving infants. As trimester 2–3 TPL increased, so did the number of activated macrophages (P �0.05) and the total malaria pigment scores (P � 0.05). Low birth weight (LBW) and intrauterine growth retardation(IUGR) were associated with high pigment scores and high numbers of activated macrophages in the IVS. Highplacental damage scores were not associated with IUGR, LBW, or early infant mortality.
INTRODUCTION
Infection of pregnant women withPlasmodium falcipa-rum has been associated with abortion, intrauterine growthretardation (IUGR), low birth weight (LBW), neonatal mor-tality, and congenital infection.1–5 We have recently estab-lished that the pregnant rhesus monkey inoculated withP.coatneyi is an excellent animal model of this disease process,demonstrating these same adverse fetal/infant outcomes.6
Examination of the placenta is essential to explaining ab-normal fetal and neonatal outcome. However, only a few re-ports have included a histopathologic examination of the pla-centa fromPlasmodium-infected women.7–14 Studies that havereported placental histopathologic findings were primarily de-scriptive and did not relate their findings to maternal morbid-ity and fetal/infant outcome.8,15–19 Malaria induced placentalchanges identified in these studies included focal syncytiotro-phoblast necrosis, irregular thickening of the trophoblast base-ment membrane, the presence of parasitized erythrocytes andaggregates of leukocytes (including macrophages) within theintervillous space, macrophages containing malaria pigment,malaria pigment deposited in fibrin and fibrinoid, fibrinoiddeposits, and fibrinoid necrosis of villi.
Most animal studies of malaria during pregnancy havebeen conducted in rodents withP. berghei.20–25 Unfortunate-ly, the reproductive systems of rodents do not represent thehuman reproductive system very well. In contrast, the non-human primate, unlike rodents, has a villous hemochorialplacenta26 and fetal organogenesis comparable to the hu-man.27 Rodents have also not proven to be useful models ofcongenital malaria.28
To fully establish theP. coatneyi-infected pregnant rhesusmonkey as a comprehensive model of malaria during preg-nancy, the results of the pathologic evaluation of the placen-tal tissues surgically collected from seven term pregnanciesand one first trimester intrauterine death are reported here.In addition, these results are correlated with maternal para-sitemia, morbidity, and fetal/infant outcome.
MATERIALS AND METHODS
General study design. Animals. The clinical details ofthis study have been published.6 Briefly, 10 pregnant, ma-
laria-naive, Indian-origin rhesus monkeys (Macaca mulatta)were inoculated intravenously withP. coatneyi during thefirst trimester between gestational days (GDs) 30 and 54.These 10 females had been born at the Tulane Regional Pri-mate Research Center (TRPRC) and therefore had no pre-vious exposure to malaria parasites. Three dams were pri-migravidas (M181, L226, and L422), one was a secundi-gravida (L414), and six were multigravidas (D673, E412,J653, L412, L434, and M488). In a subsequent study, an-other malaria-naive primigravid monkey (N833) was inoc-ulated withP. coatneyi on GD 61. Although the results ofthis animal’s inoculation were not previously published, theplacenta from this animal was included in this report.
Rhesus monkey gestation averages 165 days (� 10 days).Thus, the first trimester in the rhesus monkey is from GD 0to 55, equivalent to GD 0–90 in the human; the second tri-mester is from GD 56 to 110, equivalent to GD 91–180 inthe human; and the third trimester is from GD 111 to 165,equivalent to GD 181–270 in the human.
Inoculation and delivery scheme. Initially, fresh bloodfrom a P. coatneyi-infected rhesus monkey with a parasit-emia of 7% (106 infected red blood cells [IRBCs]) was in-oculated into four monkeys (M181, M488, L412, and E412)during their first trimester (� GD 55). Because three of fourinoculations resulted in two abortions and surgical removalof one dead fetus at 7–10 days post-infection (PI) (M181,M488, and L412), the size of the effective inoculum wasreduced. Frozen blood from the same donor (105 IRBCs) wasused to inoculate six additional monkeys (D673, J653, L226,L414, L422, and L434). All six became parasitemic within14 days PI, and along with monkey E412, carried their in-fants to term. Monkey N833 was inoculated with a similarfrozen inoculum (105 IRBCs) from a different donor and alsodelivered a viable-term infant.
Seven infants were delivered by elective cesarean sectionbefore the onset of labor (GD 155) to ensure collection ofthe placenta, which is otherwise consumed by the dam. De-livery prior to labor also ensured that the placentas did notcontain lesions associated with labor and delivery29 (DavisonBB, unpublished data). Dam D673 delivered spontaneouslyand her infant’s placenta was therefore eliminated from the

159PLACENTAL MALARIA IN MONKEYS
FIGURE 1. Schematic of a full-thickness section through a placental cotyledon. A main stem villus containing fetal vessels extends fromthe chorionic plate (fetal side) to anchor on the maternal side at the basal plate. The villous ‘‘ tree’’ extends into ramifications of intermediateand then terminal villi, which are seen as cross sections in the figures that follow. These villi contain fetal vessels and are surrounded by theintervillus space, which contains maternal blood. The syncytiotrophoblast layer, which surrounds each villus, is the definitive barrier betweenthe fetal and maternal blood supply.
TABLE 1Scoring criteria for some placental parameters studied (full definitions presented in the Methods section)
Parameter
Severity grades
0 1 2 3 4
Fibrinoid necrosis of villi* none 1 per 100 2 per 100 3 per 100 � 4 per 100Distinct fibrinoid lesion (DFL)† none 30 affected villi 60 affected villi 90 affected villi � 90 affected villi
Infarcts‡Marginal none � 4 mm � 4 mm, � 6 mm � 6 mm, � 8 mm � 8 mmBasal none � 2 mm � 2 mm, � 4 mm � 4 mm, � 6 mm � 6 mm
Chorionic plate thrombosis(CPT)§
none � 20% � 20%, � 40% � 40%, � 60% � 60%
Syncytiotrophoblast disruption(SD)¶
none � 10% � 10%, � 20% � 20%, � 30% � 30%
Chorionic plate SD none � 15% � 15%, � 30% � 30%, � 50% � 50%Malaria pigment (hemozoin)# none rarely seen or small
amounts in 1–2fields
small amounts inmore than 2 fields
multiple fields withspecific focal are-as
abundant pigmentthroughout area
Pigment in macrophages**Decidua none 1–4 5–10 11–30 � 30Basal plate none 1–3 4–8 9–20 � 20Intervillous space (IVS) none � 0.5/field � 1/field � 1.5–2/field � 3/field
Parasites in sections†† no IRBC’s rare 1 IRBC/1–2 HPF � 2 IRBC’s/HPF numerous IRBC’s
* Number of affected villi per 100 villi examined in randomly chosen microscopic fields.† Number of fetal villi affected in one area.‡ Greatest diameter.§ Percentage of chorionic plate affected.¶ SD � percentage of villi affected per 40X field. Chorionic plate SD � percentage of plate affected when entire syncytrophoblast layer examined.# Amount of pigment found in examined fields.** Highest number of pigmented macrophages found in any 40X single field or the mean number of pigmented macrophages per field.†† Number of infected RBC’s (IRBC) per high power field (HPF).
study. Infants were weighed, measured, and returned to theirdams within 24 hr of delivery. Infants rejected by theirmothers were nursery-reared. Surgery was performed onmonkey L412 to collect the fetoplacental tissues when fetaldemise was diagnosed in utero at 37 days gestation. Five
monkeys treated in the same manner, with the exception ofPlasmodium inoculation, served as controls.
Pathologic examination of placentas. Tissue collection.Placental tissues were collected at the time of elective ce-sarean section, weighed, and examined grossly. A cross-sec-

160 DAVISON AND OTHERS
FIGURE 2. (a) Matrix-type fibrinoid (F-arrows) replacing villousstroma termed, ‘‘fi brinoid necrosis of villi’’ (H&E, 100X). (b) Per-ivillous fibrinoid (arrow) replacing the syncytiotrophoblast layerwhich surrounds a villus.
FIGURE 3. (a) A ‘‘ distinct fibrinoid lesion’’ (DFL) with fibrinoidnecrosis of �120 villi (grade 4). Fibrinoid is deposited within andbetween villi, obscuring the villous outlines. There are inflammatorycells, macrophages, malaria pigment, and increased numbers of cy-totrophoblast cells. A clear demarcation exists between affected villi(left), and unaffected villi (V) (right) (H&E, 10X). (b) Higher mag-nification (40X) of the DFL in Figure 3a. with fibrinoid (F), depositsof malaria pigment (arrowheads), and inflammatory cells (arrows).
tion was removed during surgery and blotted onto five slidesto make thick and thin placental blood smears. Immediatelyfollowing surgery, sections 3–4 mm in width were takenrandomly from five different cotyledons by starting at thematernal decidua and cutting through the full thickness ofthe cotyledon,30 including the fetal chorionic plate (Figure1), and placed into tissue cassettes. In addition, five sections

161PLACENTAL MALARIA IN MONKEYS
of the umbilical cord and one strip of chorioamniotic mem-brane, rolled with cut end down,30 were placed into cassettes.Tissues were fixed in 10% neutral-buffered formalin for 24hr for histopathologic analysis or in Streck Tissue Fixative�
(STF) (Streck Laboratories Inc., Omaha, NE) for immuno-histochemical analysis. Tissue blocks were sectioned at 5�m and stained with hematoxylin and eosin, periodic acid–Schiff, and Giemsa.
Scoring of lesions. The scoring was completed by onepathologist without knowing the identity of the animals. Allsections were histologically evaluated using a list of param-eters graded from 0 to 4, with zero representing ‘‘ none’’ ,one being minimal, and 4 the most severe. For each case,the parameter score was the sum of the scores given for eachsection. For example, if the score for chorionic plate throm-bosis was 4 for each of five cotyledon sections examined,the severity score would be 20 for this parameter. Only dis-tinct fibrinoid necrosis lesions and infarcts were scored as asum of the scores for all individual lesions rather than sec-tions. The criteria are described below and summarized inTable 1.
Description of parameters evaluated. Fibrinoid deposits(FD). Fibrinoid is an acellular, intensely eosinophilic mate-rial of which there are two types located in placental tissue.These are 1) fibrin-type fibrinoid, a blood clotting productthat is free of extravillous trophoblast cells and usually incontact with the intervillous space (IVS); and 2) matrix-typefibrinoid, in which are embedded varying numbers of ex-travillous trophoblast cells and which itself is a secretoryproduct of these cells.31 When matrix-type fibrinoid replacesvillous stroma, leaving an intact trophoblastic surface, it isreferred to as fibrinoid necrosis of villi (Figure 2a). Perivil-lous fibrinoid is a matrix-type fibrinoid which replaces thetrophoblastic cover of the villus (Figure 2b).
Distinct fibrinoid lesion (DFL). This is a term coined bythe principle investigator. This parameter is qualitatively andquantitatively different from the lesion fibrinoid necrosis ofvilli as described by other investigators16–19 (see fibrinoid de-posits). The DFL is an area of fibrinoid necrosis involving� 30 fetal villi viewed in cross section. The affected areahas a complete loss of normal villous architecture. Fibrin-type fibrinoid is deposited within villi and matrix-type fibri-noid is deposited between villi, obscuring the villous outlines(Figures 3a and b). Adjacent tissues such as the basal plate,marginal sinus, and the chorionic plate are often, but notalways, involved. The DFLs may include infiltrates of in-flammatory cells and macrophages, deposits of malaria pig-ment, and often contain increased cytotrophoblast cells.
Infarcts. Marginal infarcts were defined as areas of ische-mic necrosis of villi adjacent to the marginal sinus of a cot-yledon, while basal infarcts originated adjacent to the basalplate and extended toward the amnion. Infarcts were gradedaccording to location and size, measured across their greatestdiameter using a reticle (Table 1).
Chorionic plate thrombosis (CPT). This refers to thrombiwithin fetal vessels of the chorionic plate often leading tothromboses (Figure 4a 1 and 2, 4b 1 and 2), which extendinto the major ramifications of fetal vessels within stem villi.The lesion resulted in an infarction like lesion in the chori-onic plate itself, but was not considered to be a true infarc-
tion, which only occurs when the maternal blood supply isdisrupted.31
Syncytiotrophoblast disruption (SD). The syncytiotropho-blasts form a continuous cell layer which covers the villi andchorionic plate. The syncytiotrophoblast layer was examinedfor areas of disruption (Figure 5).
Inflammation. The severity of polymorphonuclear andmononuclear infiltrates was evaluated within the IVS, villi,umbilical cord, chorioamniotic membrane, DFL (Figure 3b),and surrounding vessels in the decidua (perivascular cuff-ing). The pathologist used previous control studies to estab-lish guidelines for levels of inflammatory cells consideredas acceptable numbers seen in preterm delivered placentas.
Placental damage score. This is the sum of all scores forDFL, infarcts, CPT, fibrinoid deposits, syncytiotrophoblastdisruption, and inflammation in all sections examined for acase.
Malaria pigment (hemozoin). This is an extracellular,brown to black, iron-negative, birefringent byproduct of themetabolism of hemoglobin by Plasmodium. It was evaluatedwithin the decidua, basal plate, villi (Figure 5), intervillousspace, maternal and fetal RBCs, trophoblasts, fibrinoid de-posits, and chorionic plate.
Macrophages containing malaria pigment. These (Figure6) were evaluated in the decidua, basal plate, IVS, villi,DFL, and fetal vessels. The entire decidua or basal plate wasscanned at 40� and the highest number of pigmented mac-rophages visualized in a single field was used to determinethe score (Table 1).
Total pigment score. This is the sum of the scores forpigment within fibrin and pigment within macrophages in allsections examined for a case.
Placental parasitemia. A cross-section of placental coty-ledon was removed during surgery and blotted onto fiveslides to make thick and thin placental blood smears thatwere stained with Giemsa. The IBRC were counted in 10microscopic fields each containing approximately 200 RBCs(1,000� magnification) and recorded as a percent of RBCsparasitized.
Parasites in sections. The IRBCs were identified by lightmicroscopy in Giemsa-stained placental sections by focusingup and down on RBCs within the IVS, fetal vessels (Figure7), and umbilical cord at 100� under oil immersion optics.
Peripheral parasitemia/parasite load. To better facilitatean evaluation of the impact of circulating parasites on pla-cental lesions and fetal outcome, coupled with the gestation-al time when they occurred, the percent of maternal IRBCwas determined weekly. The percent of maternal IRBCs wasdetermined by counting 10 microscopic fields (1,000� mag-nification) each of which contained approximately 200RBCs. The RBC count on the same day was used to cal-culate the number of IRBCs/mm3 of blood. No attempt wasmade to count individual parasites within RBC. The totalparasite load (TPL) was determined to be the sum of allweekly IRBC counts during pregnancy. This sum was alsocalculated for each trimester of pregnancy with T1, T2, andT3 representing the first, second, and third trimesters, re-spectively. These figures appear in Table 2.
Activated macrophages (AM). These were identified byimmunohistochemical staining with monoclonal antibodiesto LN-5 (Zymed) (Figure 8). For each case, the macrophages

162 DAVISON AND OTHERS
FIGURE 4a. (1) A full thickness section of a cotyledon from a Plasmodium-infected monkey (E412). Chorionic plate thrombosis is severein 2 areas (arrows). Computerized image (H&E, 2X). (2) A higher magnification of the area under the arrow located outside the section inFigure 4b.1. Chorionic plate thrombosis (CPT) resulted in ischemia and necrosis which developed into an infarction-like lesion. Three fetalvessels (FV) are shown on the surface of the chorionic plate and the fetal villi (V) are seen in cross section below. Computerized image (H&E,10X).

163PLACENTAL MALARIA IN MONKEYS
FIGURE 4b. (1) A full thickness section of a cotyledon from a control monkey illustrating a normal chorionic plate (arrowheads) with afetal vessel (arrow) on the surface. Cross sections of fetal villi lie beneath the chorionic plate. Compare with Figure 4a.1 (H&E, 2X). (2) Ahigher magnification of the area of normal chorionic plate indicated by the large arrow in Figure 4b.1. There is a large fetal vessel (FV) abovethe chorionic plate (CP). Normal cross sections of fetal villi lie beneath the plate. Compare with Figure 4a.2 (10X H&E).
(stained red with amino ethyl carbazole (AEC) were countedin 10 randomly selected high-power fields (HPF) (100� oilimmersion) within the IVS and this total was divided by 10to establish the mean. The actual mean was used as the scorefor each section. Only HPFs containing substantial amountsof blood within the IVS were evaluated.
Fetal macrophages (Hofbaur cells). These were deter-mined by the same method by counting immunostained mac-
rophages in ten randomly selected terminal villi. Each villusexamined was roughly 0.06 � 0.06 mm in diameter.
Immunohistochemistry. Placental tissues were fixed inStreck Tissue Fixative (STF; Streck Laboratories, Inc. Oma-ha, NE) and activated macrophages were identified usingLN-5 (Zymed Laboratories, South San Francisco, CA) usingroutine immunohistochemistry methods.
Statistical analysis. The resulting median scores from the

164 DAVISON AND OTHERS
FIGURE 5. A fetal villus containing fibrinoid (F) and deposits of malaria pigment (arrowheads). A macrophage (small arrow) containingmalaria pigment lies adjacent to an area of syncytiotrophoblast disruption (large arrow). The syncytiotrophoblast cell layer is indicated (ST)(100X H&E).
FIGURE 6. Macrophages containing malaria pigment (arrows) in the intervillous space. Villi (V) and the intervillous space (IVS) areindicated (H&E, 100X).

165PLACENTAL MALARIA IN MONKEYS
FIGURE 7. Plasmodium coatneyi ring-stage within fetal red blood cells (RBC) (arrow) within a fetal vessel in a villus from the placenta ofmonkey L414. This infant became congenitally infected with malaria at 80 days of age (Giemsa, 100X).
TABLE 2Parasite load during each trimester coupled with maternal red blood cell count/anemia, fetal outcome and placental lesion scores
Mom/baby Gravid p1 p2 p3 TP wt g/l LRBC A IUGR pwt bwt status PD PIG INF MAC
L412M181M488N833/A109E412/V443
31317
4.8537.43
0.0033.36
4.11
59.629.65
3.901.20
63.0244.21
(�)650(�)700
3.183.813.453.493.04
nrr
m(�)
88.97113.48
4.14
abortabortabortdeadalive
154201
7132
5234
9.46.9
L422/V522L434/V485L414/V435J653/V783L226/V595
15231
0.000.000.001.200.00
2.792.58
15.572.42
12.89
1.882.832.581.385.90
4.675.41
18.155.01
18.79
(�)100(�)100(�)400(�)400(�)300
3.383.213.303.223.00
nrnrrnrnr
(�)(�)s(�)s
114.48111.38111.27107.6
82.68
4434.243.34
deaddeadalive Cdeaddead
10992
175220139
23286940
115
2112303131
2.21.59.968.5
p1–p3 � parasite loads during each trimester � 10,000, pwt � placental weight in grams (control mean � 123.9); TP � total parasite load during pregnancy � 10,000; bwt � birth weightin grams � 100 (control mean � 4.56); LRBC � lowest red blood cell count � 100,000; status � status of infant; abort � fetal demise; dead � early infant mortality (3–33 days); and alive� still alive at this time; C � congenitally infected with malaria; A � anemia with r � recovery of normal RBC count (�4.5) and nr � no recovery (�4.5) during gestation; wt g/l � weightgain or loss during pregnancy (control mean � [�]1706.25); IUGR � intrauterine growth retardation; m � mild during late gestation; s � severe; PD � placental damage score, PIG � totalpigment score, INF � inflammation score; shaded background � highest scores; MAC � average number of activated macrophages/HPF. Scores reflect total for all sections examined.There was a significant association between parasitemia (high or low) occurring during T-1 and high placental damage scores (rs � 0.7275, P � 0.05). As total pigment increased, infant
weight decreased (rs � �0.8524, P � 0.05). As macrophages increased, infant weight decreased (rs � �0.8524, P � 0.05). As parasite load during T-2 and T-3 increased, so did the numberof activated macrophages in the intervillous space (rs � 0.7500, P � 0.05).
evaluation of the parameters for Plasmodium-infected pla-centas (MedM) were compared to control placentas (MedC)using the Mann-Whitney test. Means and standard deviations(SD) were calculated for the number of activated maternalmacrophages and Hofbaur cells and placental weights. Dif-ferences between means were tested using the student t-testfor independent samples. When presented in the results, theSD follow the mean with a � sign. Correlations betweenmaternal clinical parameters, placental parameter scores andinfant outcomes were compared using the Spearman rankcorrelation. Comparisons made between selected parametersand combinations of the following variables were made us-
ing the Fisher Exact test: IUGR (presence or absence), fetaldemise (alive or dead), early infant mortality (yes or no),parasite-infected fetal RBC (presence or absence). All testswere two-tailed with the level of significance set at P � 0.05.
RESULTS
Placental weight. Placental weights in the P. coatneyi-infected group were lower than in non-infected controls (t� 2.5548, degrees of freedom [df] � 15, P � 0.0220). Theplacental weight range was 83–115 grams with a mean �SD of 104.28 � 12.80 grams. The average placental weight

166 DAVISON AND OTHERS
FIGURE 8. Activated macrophages (LN5+) appear as dark black cells (arrowheads) within the intervillous space (IVS) adjacent to fetal villi(V) in the placenta from a Plasmodium-infected monkey (AEC with H&E counterstain, 100X).
in controls was 123.9 � 20.90 grams. The placenta weighing83 grams belonged to the infant with the most severe growthretardation and a birth weight of 334 grams (control mean� 466 grams). This placenta and one other were mildly cir-cumvallate (Figure 9); however, the significance of this find-ing is unknown. All other gross parameters in the seven termplacentas appeared normal.
Placental parasitemia and parasites in sections com-pared to peripheral parasitemia. In all seven cases, thepercentage of IRBC in the placental blood smears was thesame as that of the dam’s peripheral blood on the day ofdelivery. Although IRBCs could be identified within the IVSof placental tissue sections in all seven cases, there was noincreased accumulation of IRBC within the IVS nor werethere notable IRBC adhering to the surface of the syncytio-trophoblast layer of the villi.
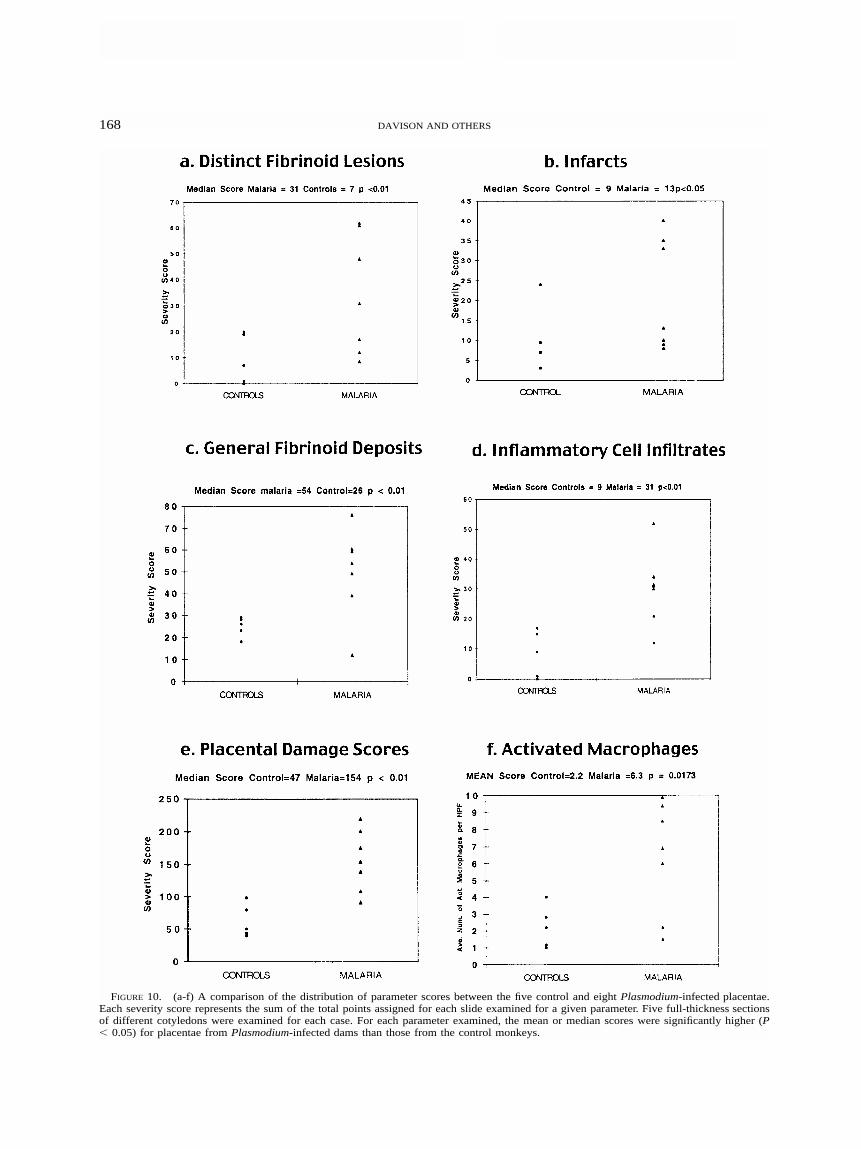
Histopathology of cesarean-delivered term placentasfrom infected versus non-infected dams. The placental his-topathology from P. coatneyi-infected monkeys (Figures 2–8) resembled that seen in P. falciparum-infected hu-mans.8,16–19,32 The seven placentas from P. coatneyi-infecteddams had more significant pathologic changes than did theplacentas from 5 control animals when the median scoreswere compared for each parameter using the Mann-Whitneytest (Figure 10 a–f). The distinct fibrinoid lesions (Figure 3aand b) were more numerous and severe when the medianscores for malaria-infected (MedM � 31) were comparedwith that of the controls (MedC � 7) (P � 0.01) (Figure10a). When the medians were compared for marginal, basal,and central infarct scores (Figure 10b), they were more se-vere in the malaria placentas (MedM � 13) than in the control
placentas (MedC � 9) (P � 0.05). Fibrinoid deposits (Figure2a and b) were more severe than those seen in controls(MedM � 54 versus MedC � 26) (P � 0.01) (Figure 10c).One of the most significant findings was the increased infil-trates of inflammatory cells (Figure 3b) in P. coatneyi-in-fected placentas (MedM � 31) compared with controls (MedC
� 9) (P � 0.01) (Figure 10d). Malaria placentas had signif-icantly increased scores for disruption of the syncytiotro-phoblast layers of both villi (Figure 5) and the chorionicplate (MedM � 18 versus MedC � 11) (P � 0.01). The pla-cental damage scores for malaria cases (MedM � 154) weresignificantly higher than that of the controls (MedC � 47) (P� 0.01) (Figure 10e). The mean number of activated mac-rophages (LN-5�) in the IVS of malaria placentas (6.3 �3.359) (Figure 8) was significantly higher than the mean forcontrols (2.2 � 1.272) (t � 2.849, df � 10, P � 0.0173)(Figure 10f). Activated Hofbaur cells (fetal macrophages)were significantly increased in malaria placentas (mean �3.7 � 1.521) over controls (mean � 1.7 � 0.303) (t � 4.353,df � 10, P � 0.0014).
Histopathology from Plasmodium-induced intrauterinedeath. Upon ultrasonographic determination of fetal deathat seven days PI, 37 days gestation, the placenta and fetuswere surgically recovered from monkey L412. This monkeyhad quickly developed active malaria with a parasitemia of21,915 parasites/mm3 of RBCs. The placenta was evaluated,but did not receive parameter scores. The etiology of fetaldeath appeared to be related to severe acute placental in-farction. Within the attached uterine tissue (endometrium)there was infarction with inflammatory cell infiltrates andsequestration of parasitized erythrocytes in endometrial cap-

167PLACENTAL MALARIA IN MONKEYS
FIGURE 9. The primary disc of this bidiscoidal rhesus monkey placenta (L226) is circumvallate. The membranes of the chorion laeva donot insert at the edge of the placental disc but at an inward distance from the margin (arrowheads). The significance of this finding is unknown.
illaries. The number of capillaries with sequestration in-creased towards the placental tissue. There was infarction ofthe basal plate with mild neutrophilic infiltrates. The inter-villous space contained deposits of fibrinoid material. Thesedeposits contained parasitized erythrocytes and malaria pig-ment. Macrophages associated with these areas containedphagocytized P. coatneyi and malaria pigment. These areaswere associated with fibrinoid necrosis of villi.
Statistical analysis of parameters studied versus preg-nancy outcome in Plasmodium-infected dams. Associa-tions between placental parameter scores, the sums of pa-rameter scores, total parasite load during pregnancy, parasiteload during individual trimesters, and various fetal outcomeswere compared by the Spearman rank correlation (rs). Onlysignificant results or those showing a high correlation arereported (Table 2).
Monkeys M488, L412, E412, and J653 were inoculatedduring the first trimester (T-1) and all four experienced par-asitemic episodes during T-1. Three had high T-1 parasiteloads of greater than 100,000 IRBCs/ml of blood (M488,L412, and E412), while monkey J653 had low levels(12,000). Two of the three mothers with high T-1 parasiteloads (M488 and L412) aborted. The placental lesions frommonkey L412 are described above. The placentas from theviable births of monkeys E412 and J653 were the only twoof seven term placentas with significant CPT (Figure 4a).Parasites in T-1 were significantly associated with CPT (rs
� 0.8321, P � 0.05) (Figure 11). These two placentas alsohad the highest overall placental damage scores. There wasa significant association between parasitemia (high or low)occurring during T-1 and high damage scores (rs � 0.7572,P � 0.05). Chorionic plate thrombosis scores were positively
associated with DFL scores (Figure 3a and b) (rs � 0.6875),but this did not reach significance. The third abortion in thisstudy (M181) was inoculated late in T-1 (GD 51) resultingin a high parasite load (65,000) early in the 2nd trimester(GD 58–60). Parasite inoculations during T-1, resulting inhigh parasite load during early gestation, were more likelyto result in fetal demise (P � 0.0476).
Low birth weight and IUGR were associated with highpigment scores and high numbers of activated macrophagesin the IVS. As the sum of the parasite load during T-2 andT-3 increased, so did the number of activated maternal mac-rophages (rs � 0.7500, P � 0.05) (Figure 8) and the totalpigment scores (rs � 0.7500, P � 0.05) (Figure 12). Therelationships between total pigment scores, activated mater-nal macrophages, and infant weight were highly significant.As total pigment increased, infant weight decreased (rs ��0.8524, P � 0.05). This relationship became even strongerwhen pigment in fibrin (rs � �0.9266, P � 0.05) (Figure 5)and pigment in macrophages (rs � �0.9266, P � 0.05) (Fig-ure 6) were examined independently. As macrophages in-creased, infant weight decreased (rs � �0.8524, P � 0.05).
Monkeys N833, L414, and L226 had the highest levels ofactivated macrophages and the highest total pigment scores.Monkeys N833 and L226 gave birth to infants that died atthree and five days of age. The infant born to L226 hadsevere IUGR, LBW, and the lowest placental weight. Mon-key L414 delivered a LBW infant with IUGR that survived,and congenital malaria was diagnosed at three months ofage. This infant’s placenta contained the highest level of ac-tivated macrophages in our study (Table 2).
As total parasite load increased, the scores for total in-
168 DAVISON AND OTHERS
FIGURE 10. (a-f) A comparison of the distribution of parameter scores between the five control and eight Plasmodium-infected placentae.Each severity score represents the sum of the total points assigned for each slide examined for a given parameter. Five full-thickness sectionsof different cotyledons were examined for each case. For each parameter examined, the mean or median scores were significantly higher (P� 0.05) for placentae from Plasmodium-infected dams than those from the control monkeys.

169PLACENTAL MALARIA IN MONKEYS
FIGURE 11. First trimester parasitemia, regardless of level, was significantly associated with high scores for chorionic plate thrombosis(CPT). See Figure 4a (P � 0.05) and total placental damage (P � 0.05). This finding is illustrated by the results observed with monkeys E412and J653. Monkey E412 had a very high parasite load during T-1 and monkey J653 had a low parasite load during T-1. Other T-1 parasitemiasresulted in abortion (P � 0.0476) (see Table 1).
FIGURE 12. High parasite loads during the latter two-thirds of pregnancy were significantly associated with high total-pigment scores (P� 0.05) and increased numbers of activated macrophages in the intervillous space (P � 0.05) (see Figure 8). As pigment increased, infantbirth weight decreased (P � 0.05) and as the number of activated macrophages increased, infant birth weight decreased (P � 0.05). Very low-birth-weight infants were born to L226 and L414 (see Table 1).
flammatory cell infiltrates increased (rs � 0.7768, P � 0.05)(Figure 3b). As scores for total inflammatory cell infiltratesincreased, so did numbers of activated Hofbaur cells (rs �0.9375, P � 0.05) and DFL scores (rs � 0.7946, P � 0.05).The correlation between inflammation and total placental
damage scores was also positive, but did not reach signifi-cance (rs � 0.6161, P � 0.05). Unexpectedly, high total par-asite loads for the entire pregnancy were not significantlyrank correlated with CPT, DFL, or the highest placental dam-age scores. High DFL scores and high placental damage

170 DAVISON AND OTHERS
scores were not associated with IUGR (P � 0.4286), LBW(rs � �0.2594), or early infant mortality (P � 1.000).
As total parasite load increased, the scores for IRBCs ob-served within the IVS also increased (rs � 0.8214, P � 0.05).The number of fetal IRBCs in vessels of placental villishowed a positive correlation with total parasite load, but thisdid not reach significance (rs � 0.6736). Fetal IRBCs wereidentified in vessels of placental villi (Figure 7) in the fourmonkeys with the highest total parasite loads (� 150,000 par-asites/mm3 of RBCs), but not in the three with low total par-asite load (� 55,000 total). This was a significant finding (P� 0.0286). Interestingly, as activated maternal macrophagesincreased within the IVS, the number of fetal IRBCs identifiedin vessels of placental villi increased (r s � 0.7769, P � 0.05).
The two monkeys with the highest parasite loads during thethird trimester, N833 and L226, had the lowest placentalweights (88.97 and 82.68 grams, respectively). Although therewas a positive association, (rs � �0.6786), it did not reachsignificance. These infants died at three and five days of age.
DISCUSSION
We used the Plasmodium coatneyi-infected pregnant rhe-sus monkey model to evaluate the role of the placenta in thepathogenesis of the adverse effects of malaria on pregnancyoutcome. We followed maternal parasite load, maternal RBCcounts, and fetal development throughout the course of in-fection. When combined with the results of thorough pla-cental histopathology (from cesarean section–delivered pla-centas that were free from the effects of parturition) andpregnancy outcome, important insights were derived.
Malaria during pregnancy in non-immune women is high-ly associated with abortion and stillbirth. For instance, in onestudy 13 of 14 women presenting with malaria in T-1 abort-ed, as opposed to eight of 19 presenting in T-2.33 In ourmodel, all three abortions occurred in dams with high par-asitemia in T-1, as did the two most severely damaged pla-centas. Thus, the degree and timing of parasitemia may beimportant factors in inducing abortion. Placental infarctionappears to be one pathologic mechanism that leads to abor-tion. Infarction may be secondary to obstruction of endo-metrial vessels with parasitized RBCs. While worse preg-nancy outcomes have been reported in primigravidas,34,35
parasitemia in T-1 appears to outweigh the effect of parityin monkeys and humans.33 Abortion as a result of infectiousdisease during early gestation is not unique to malaria.36,37
While high parasitemia in T-1 is associated with abortion,high parasitemia in T-2/3 is associated with IUGR, LBW,and congenital infection. In our study, these outcomes cor-related with high levels of activated macrophages within theintervillous space and heavy deposits of malaria pigment.
The mechanism by which placental malaria contributes toIUGR and LBW is not fully understood. In response to Plas-modium, activated placental macrophages could induce pla-cental damage, both mechanical and functional, through therelease of cytokines such as interferon- and tumor necrosisfactor-. A strong local (placental) Th1 response to Plas-modium such as that observed in a primary infection of anaive monkey may compromise pregnancy in a manner suchas we and others have postulated.32,38 In our study, areas ofsyncytiotrophoblast disruption were often associated with in-
creased aggregates of macrophages (Figure 5) and inflam-matory cells, as has been observed in human studies.8,16
While increased malaria pigment may have a primary rolein contributing to poor fetal outcome, it is more likely thatit serves as a marker for high parasitemia late in gestation.
Because it is not feasible to examine blood smears daily,levels of placental hemazoin may provide the investigatorwith a marker of severe parasitemia sometime during thelater half of pregnancy. This can easily be missed clinically.High levels of pigment in the placenta should alert the phy-sician that the infant may be at high risk for malaria inducedcomplications. However, low pigment scores will not allowthe clinician to rule out severe parasitemia during early preg-nancy. High pigment scores would not be expected to beassociated with parasitemia during the first trimester becausefibrin, where most malaria pigment is deposited, is not amajor component of a first trimester placenta. As the pla-centa matures, fibrin becomes a more prominent feature.31
Another factor affecting hemazoin levels might be the lengthof time it remains in placental tissue. Although it is thoughtto remain for many months,19 it may not last the entire ges-tation. Some women with documented peripheral blood par-asitemia during the first half of gestation had no placentalmalaria pigment at term delivery.39 This is illustrated bymonkey E412 who had one of the highest total parasite loadsin this study, but a low total pigment score. Seventy-fivepercent of the parasites in this monkey occurred during T-1.Although this placenta had high placental damage scores,the infant had no complications. In order to use hemazoinas a diagnostic tool, several placental sections must be ex-amined. Another method, which we have not yet exploredin the monkey model, would be to quantitate the hemazoinlevels in placental tissue by spectrophotometric or fluoro-metric assays.40
Intuitively, one would expect high placental damagescores to be associated with the worst infant outcome. Thiswas not the case in this study. The two placentas with thehighest placental damage scores (J653 and E412) were notassociated with IUGR, LBW, or congenital infection. Onecommon factor, unlike the other six term pregnancies, wasthat both monkeys had parasitemias during the first trimester.Monkey E412 had a high parasite load during the first tri-mester and monkey J653 had a low parasite load during T-1. Thus, it appears that any parasites occurring during T-1can result in extreme placental damage. In monkeys E412and J653, the resulting damage was manifested as throm-bosis of fetal vessels in the chorionic plate and stem villiwith extensive DFLs that appeared to result from the vas-cular occlusion. Thrombosis of chorionic vessels is knownto occur in placental toxoplasmosis41 and in infections withcytomegalovirus as well.42,43
Fibrinoid necrosis of villi was a prominent finding in Plas-modium-infected placentas; however, it was not a hallmarkof poor infant outcome. Fibrinoid necrosis and distinct fi-brinoid lesions of the placenta were defined in a mannerwhich is qualitatively different than that reported in the hu-man placental malaria literature.8,15–19 Fibrinoid necrosis isknown to be a normal component of term placental villi;31,44
however, extensive fibrinoid necrosis is abnormal.31 In aneffort to differentiate normal fibrinoid from a ‘‘ distinct fi-brinoid lesion’’ , we only considered large areas of necrosis

171PLACENTAL MALARIA IN MONKEYS
involving 30 or more villi. (Figure 2). There could be manyreasons why fibrinoid necrosis was not associated with poorinfant outcome. Although these lesions were extensive insome cases, they were focal and the majority of fetal villiremained unaffected. Five sections do not necessarily rep-resent the scope of all placental lesions or indicate the extentof unaffected tissue. We have observed relatively unaffectedsections in placentas that have extensive lesions elsewhere.The placenta is a large organ and compensatory responsesby the remaining healthy tissue could be adequate to supporta healthy fetus. Many investigators believe that fibrinoid isimmunologically protective,45–47 possibly masking fetal an-tigens from the maternal immune system.45 Fibrinoid is alsothought to serve as an immunoprotective sponge. It may ex-press target antigens that bind circulating maternal antibod-ies, and the resulting immune complexes are then thought tocontribute to the deposition of fibrinoid.48 The bound anti-bodies are internalized and degraded within 4–6 hr.49,50 TheP. coatneyi-infected placentas showed a 3.4-fold increase ininflammatory infiltrates and a 2.2-fold increase in Hofbaurcells compared with control placentas. As inflammatory in-filtrates and Hofbaur cells increased in P. coatneyi-infectedplacentas, so did DFL and placental damage scores. The in-crease in these activated fetal macrophages may indicate thatthe fetus is exposed in utero to Plasmodium and or Plas-modium antigen. In humans, prenatal immune priming tomalarial antigens occurs in fetuses whose mothers had ma-laria during pregnancy.51 Perhaps in placentas exposed tocirculating Plasmodium-infected erythrocytes, fibrinoid pro-tects the placenta/fetus against Plasmodium-sensitized mac-rophages and lymphocytes and absorbs maternal antibodiesto Plasmodium. In one study, LBW was linked to increasedIVS inflammation but not to fibrin deposits or placental par-asitemia.32 It will be interesting to explore the implicationsof activated Hofbaur cells on fibrin deposition, fetal/infantprotection against congenital infection, and subsequent pro-tection from exposure after birth. Increases in Hofbaur cellshave been observed in humans.19
Sequestration of Plasmodium-infected erythrocytes was notobserved in the placentas in this study. Neither cytoadherenceof IRBCs to syncytiotrophoblasts nor increased concentrationof IRBCs in the IVS was seen. If sequestration does occur inthe monkey model, our failure to observe it may be the resultof small sample size. Sequestration may also be associatedwith labor and vaginal delivery while the placentas in thisstudy were cesarean derived. Sequestration may be a terminalevent. As uterine contractions begin, disruption of blood flowto the placenta could lead to a more stagnant flow in circu-lation with IRBC accumulation in the IVS. Very high levelsof IRBC occur in only about 25% of placentas from malaria-infected women, especially primigravidae.52 After a criticalreview of the literature, it is our opinion that the term ‘‘ se-questration’’ has been used differently in reference to the pla-cental intervillous space than is commonly used in referenceto the microvasculature of the brain. Originally, ‘‘ placentalsequestration’’ referred to the observation that higher levelsof parasitemia occurred within blood collected from the in-tervillous space or in IVS blood in histopathologic sectionsthan in the peripheral blood of the mother at the time of de-livery.8,17 Clear evidence has been presented by many inves-tigators demonstrating an increased accumulation of IRBC
within the IVS in vivo.8,15–17 The general term sequestration isoften used by pathologists in this manner. On the other hand,malariologists have a more specific definition for sequestra-tion, which describes a receptor/ligand driven phenomenonwhereby IRBCs adhere to the vascular endothelium. An ex-ample is cerebral malaria. The distinction between these twophenomena has now been lost in that placental sequestrationhas been interpreted by some investigators to describe anevent whereby IRBCs preferentially adhere to the syncytio-trophoblast cell layer. Binding of IRBCs to syncytiotropho-blast has rarely been observed in vivo. This type of placentalsequestration phenomenon was first explored by Bray andSinden in 1979.15 Based on the literature concerning seques-tration as an endothelial cell event, Bray and Sinden exploredthe possibility that this same type of phenomenon occurred inthe placenta by histopathology and electron microscopy. Theirstudies did not reveal any significant number of IRBCs liningthe syncytiotrophoblast. This type of in vivo sequestration hasnot been described by others.16–19,53,54 However, there are manyreports that describe specific receptor driven placental se-questration in vitro, and the mechanism by which IRBC ac-cumulate within the IVS is now being advanced as a receptordriven event.38,54–59
In an ex vivo model, Fried and Duffy55 used human pla-cental sections and demonstrated that IRBCs from pregnantwomen bound exclusively to chondroitin sulfate A (CSA), areceptor that occurs on the surface membrane of placentalsyncytiotrophoblasts,57,60 while no IRBCs from nonpregnantwomen were bound. Maternal IRBCs from the peripheral cir-culation had a CD-36 binding phenotype, a CSA-binding phe-notype, or a mixed phenotype. There was no adhesion of pla-cental IRBC to CD36, thrombospondin (TSP), or intercellularadhesion molecule-1 (ICAM-1) in vitro.55 The role of variantantigens on the surface of infected erythrocytes in cytoadhe-sion to purified CSA, CD36, and ICAM-1 was explored usingIRBCs from placentas, maternal peripheral blood, and bloodfrom malaria-infected children.54 In contrast to Fried and Duf-fy, Beeson and others found that the phenotype of IRBC thatbind to CSA may not be exclusively localized to the placen-ta.54 The results indicated that 80% of midpregnancy periph-eral blood isolates adhere to CSA as did 78% of placentalisolates and 13.6% of child isolates. Four placental isolatesdid not bind to CSA. Adhesion to CD36 occurred in 65% ofchild isolates, 47.1% of peripheral maternal isolates, and17.6% of placental isolates. There was an absence of signif-icant adhesion of both placental and maternal peripheral bloodisolates to ICAM-1 whereas 84.2% of child isolates bind tothis receptor. The investigators suggested not only a role forCSA as a factor for sequestration, but indicated that low pla-cental blood flow, reduced cell deformability of IRBC, andimpaired T-cell responses to infection are also likely to beinvolved. In a comprehensive review of maternal malaria andparasite adhesion, Fried and Duffy explain the lack of in vivoattachment of IRBCs to villi as resulting from mechanicaldislodgment during vaginal delivery.61 The rhesus monkeymalaria-infected placentas were delivered by cesarean sectionand IRBCs were not observed adhering to the surface of villi.The monkey model will make a valuable contribution towardtesting this and many other hypotheses. Whether in vitro stud-ies of cytoadhesion are representative of mechanisms in vivoremains to be seen.

172 DAVISON AND OTHERS
Although there were many similarities between humanand monkey placental lesions associated with malaria, therewere also differences. It is likely that these differences areattributable to sampling technique. Previous studies of pla-cental pathology in malaria have been primarily descrip-tive,8,15–19 and few have explored the mechanisms of malaria-induced placental lesions or their relationship to fetal out-come. In most human studies, placental pathology has beenperformed on a single 1 cm2 sample of placental tissue takenfrom an area closely associated with the basal plate, ratherthan on full thickness sections. Our study used samplingtechniques as described by the College of American Pathol-ogists, which recommends taking full thickness sectionsfrom the central area of cotyledons by cutting from the fetalto the maternal surface and including both amnion and de-cidua.30 We also examined a minimum of five placental sec-tions taken from five different cotyledons, as well as sectionsof chroioamniotic membrane and umbilical cord. This tech-nique resulted in more extensive qualitative and quantitativedata than has been collected in human studies.
Human studies have often correlated various fetal out-comes with placental malaria. The term ‘‘ placental malaria’’ ,although widely used, has not been well-defined and has,therefore, produced much confusion. In most studies, pla-cental malaria is defined as the presence of Plasmodium orPlasmodium-infected RBCs within the placenta; others haveadded malaria pigment as a criterion.14 Methods used toidentify parasites in the placenta include hematoxylin andeosin- and Giemsa-stained placental tissue sections,11–13 im-pression smears of cut, uncut, or crushed placental tis-sue,10,14,62 smears of blood removed with a pipette or needleand syringe,7–9 smears of blood drippings from cut placentasallowed to sit for a specified time, smears of blood collectedby placental perfusion,63 and blood taken by undescribedtechniques.64 The term placental malaria may or may notdefine the infection status of the placenta on the day of de-livery, is not indicative of past infections which havecleared, or of chronic infections, and should not be confusedwith malaria-related histopathology describing specific pla-cental lesions.
To improve fetal/infant outcome in pregnant mothers withmalaria, the alterations in normal placental morphology andfunction, and the gestational time when these events occurmust be defined. The findings in our study demonstrate thevalidity of the monkey model and begin to define the patho-biology of malaria during pregnancy. In most human studies,therapeutic interventions cannot be avoided and this furthercomplicates the data. Controlled studies in the monkey mod-el can provide information that will be crucial to successfultherapeutic interventions in areas where there are limited re-sources.
Acknowledgments: We are grateful to Renee Grow (Pathology De-partment of the TRPRC) for technical assistance with histology andimmunohistochemistry. We are also grateful to Dr. Peter Gerone(Director of the TRPRC) who made this study possible through hisgenerous support of this project.
Financial support: This work was supported by the NIH grant#5P51-RR00164-31 NIH/NCRR Support for Regional Primate Re-search Centers and NIH/DRR Grant #1K01RR00075 SERCAAWARD.
Authors’ addresses: Billie B. Davison and Gary B. Baskin, Depart-
ment of Pathology, Tulane Regional Primate Research Center, 18703Three Rivers Road, Covington, LA. 70433. Frank B. Cogswell andElizabeth W. Henson, Department of Parasitology, Tulane RegionalPrimate Research Center, 18703 Three Rivers Road, Covington, LA.70433. Kathrine P. Falkenstein, Department of Veterinary Science,Tulane Regional Primate Research Center, 18703 Three RiversRoad, Covington, LA. 70433. Donald J. Krogstad, Tulane School ofPublic Health and Tropical Health and Medicine, Tulane UniversityMedical Center, 1501 Canal Street, New Orleans, LA. 70112.
Reprint requests: Billie B. Davison, Department of Pathology, Tu-lane Regional Primate Research Center, 18703 Three Rivers Road,Covington, LA. 70433.
REFERENCES
1. Desowitz R, 1988. Prenatal immune priming in malaria: anti-gen-specific blastogenesis of cord blood lymphocytes fromneonates born in a setting of holoendemic malaria. Ann TropMed Parasitol 82: 121–125.
2. Menendez C, 1995. Malaria during pregnancy: a priority areaof malaria research and control. Parasitol Today 11: 178–183.
3. Mutabingwa TK, 1994. Malaria and pregnancy: epidemiology,pathophysiology and control options. Acta Trop 57: 239–254.
4. Nathwani D, Currie PF, Douglas JG, Green ST, Smith NC, 1992.Plasmodium falciparum malaria in pregnancy: a review. Br JObstet Gynaecol 99: 118–121.
5. Steketee R, Wirima J, Slutsker L, Heymann D, Breman J, 1996.The problem of malaria and malaria control in pregnancy insub-Sahran Africa. Am J Trop Med Hyg 55 (suppl 1): 2–7.
6. Davison BB, Cogswell FB, Baskin GB, Falkenstein KP, HensonEW, Tarantal AF, Krogstad DJ, 1998. Plasmodium coatneyi inthe rhesus monkey (Macaca mulatta) as a model of malariain pregnancy. Am J Trop Med Hyg 59: 189–201.
7. Jelliffe EFP, 1968. Low birth-weight and maternal infection ofthe placenta. Bull World Health Organ 38: 69–78.
8. Galbraith RM, Fox H, Hsi B, Galbraith MP, Bray RS, Faulk WP,1980. The human materno-foetal relationship in malaria. II.Histological, ultrastructural and immunopathological studiesof the placenta. Trans R Soc Trop Med Hyg 74: 61–72.
9. McGregor IA, Wilson ME, Billewicz WZ, 1983. Malaria infec-tion of the placenta in The Gambia, West Africa: its incidenceand relation to stillbirth, birthweight and placental weight.Trans R Soc Trop Med Hyg 77: 232–244.
10. Desowitz RS, Alpers MP, 1992. Placental Plasmodium falcipa-rum parasitaemia in East Sepik (Papua New Guinea) womenof different parity: the apparent absence of acute effects onmother and foetus. Ann Trop Med Parasitol 86: 95–102.
11. Nyirjesy P, Kavasya T, Axelrod P, Fischer P, 1993. Malaria dur-ing pregnancy: neonatal morbidity and mortality and the ef-ficacy of chloroquine prophylaxis. Clin Infect Dis 16: 127–132.
12. Meuris S, Piko BB, Eerens P, Vanbellinghen A-M, Dramaix M,Hennart P, 1993. Gestational malaria: assessment of its con-sequences on fetal growth. Am J Trop Med Hyg 48: 603–609.
13. Redd SC, Wirma JJ, Steketee RW, 1994. Risk factors for anemiain young children in rural Malawi. Am J Trop Med Hyg 51:170–174.
14. McDermott JM, Wirima JJ, Steketee RW, Breman JG, HeymannDL, 1996. The effect of placental malaria infection on peri-natal mortality in rural Malawi. Am J Trop Med Hyg 55: 61–65.
15. Bray RS, Sinden RE, 1979. The sequestration of Plasmodiumfalciparum infected erythrocytes in the placenta. Trans R SocTrop Med Hyg 73: 716–719.
16. Walter PR, Garin Y, Blot P, 1982. Placental pathologic changesin malaria. Am J Pathol 109: 330–342.
17. Yamada M, Steketee R, Abramowsky C, Kida M, Wirima J,Heymann D, Rabbege J, Breman J, Aikawa M, 1989. Plas-modium falciparum-associated placental pathology: a lightand electron microscopic and immunohistologic study. Am JTrop Med Hyg 41: 161–168.
18. Bulmer JN, Rasheed FN, Francis N, Morrison L, Greenwoods

173PLACENTAL MALARIA IN MONKEYS
BM, 1993. Placental malaria. I. pathological classification.Histopathology 22: 211–218.
19. Bulmer JN, Rasheed FN, Morrison L, Francis N, GreenwoodsBM, 1993. Placental malaria. II. A semi-quantitative investi-gation of the pathological features. Histopathology 22: 219–225.
20. Odoula A, Holbrook T, Galbraith R, Bank H, Spicer S, 1982.Effects of malaria (Plasmodium berghei) on the maternal-fetalrelationship in mice. J Protozool 29: 77–81.
21. Odoula A, Phillips J, Spicer S, Galbraith R, 1986. Plasmodiumberghei: histology, immunocytochemistry, and ultrastructureof the placenta in rodent malaria. Exp Parasitol 62: 181–193.
22. van Zon AAJC, Eling WMC, Hermsen CC, 1985. Pregnancy-induced recrudescences strengthen malarial immunity in miceinfected with Plasmodium berghei. Parasitol 91: 9–17.
23. Vinayak V, Pathak G, Asnani P, Jain S, Malik A, 1986. Influenceof malarial infection on the maternal-foetal relationship inpregnant mice. Aust J Exp Biol Med Sci 64: 223–227.
24. Desowitz R, Shida K, Pang L, Buchbinder G, 1989. Character-ization of a model of malaria in the pregnant host: Plasmo-dium berghei in the white rat. Am J Trop Med Hyg 41: 630–634.
25. Tegoshi T, Desowitz R, Pirl K, Maeno Y, Aikawa M, 1992.Placental pathology in Plasmodium berghei-infected rats. AmJ Trop Med Hyg 47: 643–651.
26. Martin CB Jr, Ramsey EM, 1970. Gross anatomy of the placentaof rhesus monkeys. Am J Obstet Gynecol 36: 167–177.
27. Gribnau AA, Geijsberts LGM, 1981. Developmental stages inthe rhesus monkey. Adv Anat Embryol Cell Biol 68: 1–83.
28. Fischer P, 1996. Wistar rat-Plasmodium berghei model does notapproximate human congenital malaria. J Parasitol 82: 635–637.
29. Naeye RL, 1992. Disorders of the Placenta, Fetus, and Neo-nate: Diagnosis and Clinical Significance. St. Louis, MO:Mosby-Year Book, Inc.
30. Driscoll S, Langston C, 1991. College of American PathologistsConference XIX on the Examination of the Placenta: reportof the Working Group on methods for placental examination.Arch Pathol Lab Med 115: 704–708.
31. Benirschke K, Kaufmann P, 1995. Pathology of the Human Pla-centa. Third edition. New York: Springer-Verlag.
32. Leopardi O, Naughten W, Salvia L, Colecchia M, Matteelli A,Zucchi A, Shein A, Muchi JA, Carosi G, Ghione M, 1996.Malaric placentas: a quantitative study and clinico-patholog-ical correlations. Pathol Res Pract 192: 892–898.
33. Herd N, Jordon T, 1981. An investigation of malaria duringpregnancy in Zimbabwe. Cent Afr J Med 27: 62–68.
34. Brabin BJ, 1991. The risk and severity of malaria in pregnantwomen. Applied Field Research in Malaria Reports. Geneva:World Health Organization.
35. McGregor IA, 1984. Epidemiology, malaria and pregnancy. JTrop Med Hyg 33: 517–525.
36. Anonymous, 1989. Screening for congenital CMV. Lancet ii:599–600.
37. Desmonts G, Couvreur J, 1974. Congenital toxoplasmosis: aprospective study of 378 pregnancies. N Engl J Med 290:1110–1116.
38. Fried M, Muga RO, Misore AO, Duffy PE, 1998. Malaria elicitstype 1 cytokines in the human placenta: IFN-gamma andTNF-alpha Associated with pregnancy outcomes. J Immunol160: 2523–2530.
39. Watkinson M, Rushton DI, 1983. Plasmodial pigmentation andoutcome of pregnancy in west African mothers. Br Med J287: 251–254.
40. Sullivan A, Meshnick S, 1996. Hemozoin: identification andquantification. Parasitol Today 12: 161–163.
41. Khodr G, Matossian R, 1978. Hydrops fetalis and congenitaltoxoplasmosis: value of direct immunofluorescence test.Obstet Gynecol 51: 74s–77s.
42. Garcia AGP, Fonseca EF, de Marques RL, Lobato Y, 1989. Pla-cental morphology in cytomegalovirus infection. Placenta 10:1–18.
43. Altshuler G, Mc Adams AJ, 1975. The human villitides: a re-
view of chronic intrauterine infection. Curr Top Pathol 60:63–112.
44. Nelson DM, Crouch EC, Curran EM, Farmer DR, 1990. Tro-phoblast interaction with fibrin matrix. Epithelialization ofperivillous fibrin deposits as a mechanism for villous repairin the human placenta. Am J Pathol 136: 855–865.
45. Currie CA, Bagshawe KD, 1967. The masking of antigens ontrophoblasts and cancer cells. Lancet I: 708–710.
46. Kirby DRS, Billington WD, Bradbury S, Goldstein DJ, 1964.Antigen barrier of mouse placenta. Nature 204: 548–549.
47. Chaouat G, Kolb JP, Wegmann TG, 1983. The murine placentaas an immunological barrier between the mother and the fetus.Immunol Rev 75: 31–60.
48. Swinburne LM, 1970. Leucocyte antigens and placental sponge.Lancet ii: 592–593.
49. Raghupathy R, Singh B, Barrington-Leigh J, Wegmann TG,1981. The ontogeny and turnover kinetics of paternal H-2 Kantigenic determinants on the allogenic murine placenta. JImmunol 127: 2074–2079.
50. Raghupathy R, Singh B, Wegmann TG, 1984. Fate of antipa-ternal H-2 antibodies bound to the placenta in vivo. Trans-plantation 37: 296–300.
51. Fievet N, Ringwald P, Bickii J, Dubois B, Maubert B, Le HesranJY, Cot M, Deloron P, 1996. Malaria cellular immune respons-es in neonates from Cameroon. Parasite Immunol 18: 483–490.
52. Maeno Y, Steketee RW, Nagatake T, Tegoshi T, Desowitz RS,Wirima JJ, Aikawa M, 1993. Immunoglobulin complex de-posits in Plasmodium falciparum-infected placentas from Ma-lawi and Papua New Guinea. Am J Trop Med Hyg 49: 574–580.
53. Matteelli A, Caligaris S, Castelli F, Carosi G, 1997. The placentaand malaria. Ann Trop Med Parasitol 91: 803–810.
54. Beeson J, Brown G, Molyneaux M, Mhango C, Dzinjalamala F,Rogerson S, 1999. Plasmodium falciparum isolates from in-fected pregnant women and children are associated with dis-tinct adhesive and antigenic properties. J Infect Dis 180: 464–472.
55. Fried M, Duffy P, 1996. Adherence of Plasmodium falciparumto chondroitin sulfate A in the human placenta. Science 272:1502–1504.
56. Cooke BM, Rogerson SJ, Brown GV, Coppel RL, 1996. Ad-hesion of malaria-infected red blood cells to chondroitin sul-fate A under flow conditions. Blood 88: 4040–4044.
57. Maubert B, Guilbert LJ, Deloron P, 1997. Cytoadherence ofPlasmodium falciparum to intercellular adhesion molecule 1and chondroitin-4-sulfate expressed by the sycytiotrophoblastin the human placenta. Infect Immun 65: 1251–1257.
58. Rogerson SJ, Brown GV, 1997. Chondroitin sulfate A as anadherence receptor for Plasmodium falciparum-infectederythrocytes. Parasitol Today 13: 70–75.
59. Maubert B, Riche D, Deloron P, 1998. An in vitro microassayto assess the ability of Plasmodium falciparum-infected eryth-rocytes to bind to the human syncytiotrophoblast. Am J Re-prod Immunol 40: 401–407.
60. Parmley RT, Takagi M, Denys FR, 1984. Ultrastructural local-ization of glycosaminoglycans in human term placenta. AnatRec 210: 477–484.
61. Fried M, Duffy PE, 1998. Maternal malaria and parasite adhe-sion. J Mol Med 76: 162–171.
62. Archibald HM, 1956. The influence of malarial infection of theplacenta on the incidence of prematurity. Bull World HealthOrgan 15: 842–845.
63. Moore JM, Nahlen B, Ofulla AV, Caba J, Ayisi J, Oloo A,Misore A, Nahmias AJ, Lal AA, Udhayakumar V, 1997. Asimple perfusion technique for isolation of maternal intervil-lous blood mononuclear cells from human placentae. J Im-munol Methods 209: 93–104.
64. Schultz LJ, Steketee RW, Macheso A, Kazembe P, Chitsulo L,Wirima JJ, 1994. The efficacy of antimalarial regimens con-taining sulfadoxine-pyrimethamine and/or chloroquine in pre-venting peripheral and placental Plasmodium falciparum in-fection among pregnant women in Malawi. Am J Trop MedHyg 51: 515–522.