physiology and behavior at adult ecdysis · physiology and behavior at adult ecdysis 3039 recorded....
TRANSCRIPT

Insects undergo a periodic molting of their exoskeleton toaccommodate growth and changes in morphology. The moltbegins with apolysis, the detachment of the epidermis from theold cuticle, progresses to the secretion of the new cuticle andends with ecdysis, the series of events that bring about theshedding of the old cuticle and expansion of the new one. Theoverall progression of the events of the molt is controlled bythe titre of the steroid molting hormones ecdysone and itsactive metabolite 20-hydroxyecdysone (Riddiford, 1993). Atthe end of the molt, peptide hormones are released in seriesand act on the steroid-primed tissues to regulate the precisetiming of ecdysis.
The hormonal regulation of ecdysis is best understood in thelarval stages of the tobacco hornworm Manduca sexta(Truman, 1992). In M. sexta, ecdysis is under the control of atleast three peptide hormones, eclosion hormone (EH) (Truman,1992), ecdysis-triggering hormone (ETH) (Zitnan et al., 1996)and crustacean cardioactive peptide (CCAP) (Gammie and
Truman, 1997). EH and ETH act reciprocally as releasinghormones for each other (Ewer et al., 1997; Kingan et al., 1997;Gammie and Truman, 1999), and each is capable of elicitingpremature ecdysis when injected into animals that are nearingthe end of the molting period. In addition, ETH causes pre-ecdysis behavior, while EH facilitates CCAP release bycausing an increase in levels of the second messengerguanosine 3′,5′-cyclic monophosphate (cGMP) in the CCAPcells. CCAP, in turn, directly triggers ecdysis-related motoroutput (Gammie and Truman, 1997, 1999).
Although EH appears to play a central role in thecoordination of ecydsis in M. sexta, the evidence for a similarrole in ecdysis control in D. melanogaster is weaker.Immunocytochemical studies show that EH is released priorto ecdysis of the third-instar larva and prior to adult eclosion,suggesting a function during at least these two ecdyses(Horodyski et al., 1993). The selective killing of the EH cellsin early larval stages by ectopic expression of cell death genes
3037The Journal of Experimental Biology 202, 3037–3048 (1999)Printed in Great Britain © The Company of Biologists Limited 1999JEB2144
In insects, ecdysis is thought to be controlled by theinteraction between peptide hormones; in particularbetween ecdysis-triggering hormone (ETH) from theperiphery and eclosion hormone (EH) and crustaceancardioactive peptide (CCAP) from the central nervoussystem. We examined the behavioral and physiologicalfunctions of the first two of these peptides in Drosophilamelanogaster using wild-type flies and knockout flies thatlacked EH neurons. We used ETH from Manduca sexta(MasETH) to induce premature ecdysis and compared theresponses of the two types of flies.
The final release of EH normally occurs approximately40 min before ecdysis. It is correlated with cyclic guanosinemonophosphate (cGMP) production in selected neuronsand tracheae, by an elevation in the heart rate and by thefilling of the new tracheae with air. Injection of developingflies with MasETH causes all these events to occurprematurely. In EH cell knockouts, none of these changesoccurs in response to MasETH, and these flies show apermanent failure in tracheal filling. This failure can be
overcome in the knockouts by injecting them withmembrane-permeant analogs of cGMP, the secondmessenger for EH.
The basis for the 40 min delay between EH release andthe onset of ecdysis was examined by decapitating flies atvarious times relative to EH release. In flies that hadalready released EH, decapitation was always followedwithin 1 min by the start of ecdysis. Immediate ecdysis wasnever observed when the EH cell knockout flies weredecapitated. We propose that EH activates both ventralcentral nervous system elements necessary for ecdysis(possibly the CCAP cells) and descending inhibitoryneurons from the head. This descending inhibitionestablishes a delay in the onset of ecdysis that allows thecompletion of EH-activated physiological processes such astracheal filling. A waning in the inhibition eventually allowsecdysis to begin 30–40 min later.
Key words: eclosion hormone, ecdysis-triggering hormone, trachea,eclosion, peptide, hormone, Drosophila melanogaster.
Summary
Introduction
THE HORMONAL COORDINATION OF BEHAVIOR AND PHYSIOLOGY AT ADULTECDYSIS IN DROSOPHILA MELANOGASTER
JAMES D. BAKER*, SUSAN L. MCNABB AND JAMES W. TRUMANDepartment of Zoology, University of Washington, Box 351800, Seattle, WA 98195, USA
*e-mail: [email protected]
Accepted 16 August; published on WWW 13 October 1999

3038
results in the death of 50 % of the progeny during the larvalstages (McNabb et al., 1997), but many of these flies (EH cellknockouts, hereafter called ‘knockouts’) are able to completeadult eclosion successfully. These flies have defects inmultiple aspects of eclosion and post-eclosion behavior(McNabb et al., 1997). They are not responsive to a ‘lightson’ signal that induces a burst of eclosion in wild-type flies.The intensity of the eclosion response in the knockouts isweaker, and they require twice as long to eclose once theprocess has begun. In addition, 76 % of eclosing knockoutsfail to show wing expansion, the last event of the ecdysissequence. The present study uses wild-type and knockout D.melanogaster to explore the relationship between the EH cells,ETH and the key physiological changes important for thecompletion of eclosion. Using these tools, we found that thefilling of the tracheal system with air is under the control ofthe products of the EH cells, acting via an increase in cGMPlevels. Our results are consistent with the concept that therelease of a product from the EH cells, probably EH, initiatesboth an ecdysis program within the ventral central nervoussystem and a brain-based inhibition of that program. Thissimultaneous activation and inhibition would allow for thecompletion of EH-cell-triggered peripheral changes, such asthose in the tracheae and cuticle, before the initiation ofeclosion itself.
Materials and methodsStocks
Drosophila melanogaster Meig. were raised on a standardcornmeal, molasses medium. We used three genotypes in thisstudy. Canton-S was our wild-type stock. EH cell knockouts(McNabb et al., 1997) were produced by the ectopic expressionof the cell death gene reaper (rpr) (Nordstrom et al., 1996) inthe EH cells using the GAL4-UAS system (Brand andPerrimon, 1993). To generate the knockouts, transgenic fliescontaining an EH upstream sequence fused to GAL4 (EHups-GAL4) were crossed with flies of a strain that contained a UAS-rpr construct. The control flies were the progeny of a crossbetween the w1118 line used in preparing the EHups-GAL4 fliesand the UAS-rpr stock.
Staging
Two methods were used to stage D. melanogaster. Animalswere collected at the white puparium stage and maintainedthrough adult development in a humid chamber at 25 °C. Thesewere used in experiments requiring large numbers of pharateadults. When necessary, developing flies were restaged usingmorphological criteria (Bainbridge and Bownes, 1981).
The second method for staging utilized the timetable givenby Kimura and Truman (1990) for the events at the endof adult development. These stages are based onmorphologically recognizable changes, which correlatedirectly with changes in the physiology. For example, thetransition from smooth to smooth/grainy is due to thebeginning of resorption of the molting fluid. The transition
from grainy to grainy/white is due to the inflation of the largetracheal sacs in the head. In the knockouts, the head tracheaedo not fill before the molting fluid is resorbed and, therefore,they have no grainy/white stage. For convenience, stages havebeen converted to times in the text.
Measurement of heart rate
Pupae of various stages were stuck to a microscope slideusing double-sided sticky tape. When the insect was observedthrough a dissecting microscope, the heart was clearly visiblethrough the puparium, pupal and adult cuticles. Heart rate wasmeasured for 1 min intervals using a handheld counter and atimer.
Immunocytochemistry
Nervous systems were dissected from pupae of differentstages submerged in cold saline (Ephrussi and Beadle, 1936)and were subsequently fixed in 4 % formaldehyde inphosphate-buffered saline (PBS) for 4 h at room temperature(23–26 °C) or overnight at 4 °C. For cGMP immunostaining,it was necessary to dissect insects in 4 % formaldehyde inPBS. After fixation, the nervous systems were washed threetimes in PBS containing 0.3 % Triton X-100 (PBS-TX) for1–3 h. Primary antibodies were diluted in PBS-TX containing1 % normal donkey serum (NDS) (Jackson ImmunoresearchLaboratories) and 0.005 % sodium azide. Rabbit anti-EHantiserum was added at a dilution of 1:100. Mouse anti-SCP(small cardioactive peptide) was used at a dilution of 1:40.Rabbit anti-PDH (pigment-dispersing hormone) (Dircksen etal., 1987) was added at a dilution of 1:1000, and rabbit anti-CCAP (Dircksen and Keller, 1988) was added at a dilutionof 1:5000. Sheep anti-cGMP was added at a dilution of1:10 000. The tissue was incubated in the primary antibodyovernight at 4 °C. Following washes in PBS-TX, the tissueswere incubated in a donkey secondary antibody conjugatedto fluorescein isothiocyanate, Texas Red or CY5 (JacksonImmunoresearch Laboratories). Secondary antibodies wereused at a dilution of 1:1000 in PBS-TX with 1 % NDS and0.005 % sodium azide for 8 h at room temperature orovernight at 4 °C. The nervous systems were then washedthree times in PBS-TX for 1–3 h. For double and triple labels,the tissues were incubated in all primary antibodiessimultaneously.
Stained tissues were mounted on poly-lysine-coatedcoverslips, dehydrated through an ethanol series and cleared inxylene. The specimens were mounted in DPX (FlukaBiochemika). Images were collected using a BioRAD MRC-600 confocal microscope.
Eclosion hormone measurements
z-series were collected to include all stained regions of theEH neurons. The z-series were imported into NIH-Image v. 1.6and projected in the z-axis accumulating to the brightest point.The images were then imported into Photoshop v. 3.0 (Adobe),where the axons of the EH-immunopositive cells were outlinedusing the selection tools, and the number of pixels was
J. D. BAKER, S. L. MCNABB AND J. W. TRUMAN

3039Physiology and behavior at adult ecdysis
recorded. The pixel number was converted to an absolute areaon the basis of the magnification.
Video analysis of pre-eclosion behavior
The puparial case was dissected 9–15 h before eclosion toexpose the upper thorax and head of the developing adults.The ventral part of the puparial case was then secured to acoverslip and mounted in a humid recording chamber, whichconsisted of a plastic Petri dish with an approximately 1 cmhole cut in its upper surface and a glass coverslip glued to thedish using epoxy resin to cover it. Saliva was used to preventcondensation onto the glass. Care was taken to avoidincreases in temperature by monitoring the temperature nextto the recording chamber, and the light intensity was adjustedaccordingly. Time-lapse video recordings spanning the9–15 h were recorded on a Panasonic time-lapse S-videotape recorder at a 24-fold compression (approximately1 frame s−1). We chose this level of compression afterobserving that, on average, eclosion movements lasted atleast 1 s. Films were scored for movements of the abdomenor head, and the timestamp was used to record the time ofeach movement.
Injection of MasETH and cGMP
Synthetic Manduca sexta ETH (MasETH) (Zitnan et al.,1996) was diluted into saline from a 1 mmol l−1 stock solution.Staged developing adults, sorted by sex, were stuck tocoverslips using double-sided sticky tape, and the operculumand anterior dorsal portion of the puparium were removed. Thedeveloping adults were then maintained in a humid chamberuntil they reached the desired stage (Kimura and Truman,1990). Pharate adults were injected in the lateral thorax usingglass micropipettes and a Picospritzer II (General ValveCorporation). The volume of injectate was estimated at thestart and end of the injection series by measuring the diameterof a drop injected into immersion oil. As controls, equalvolumes of saline were injected into animals at the samedevelopmental stage. Injection volume ranged from 80 to300 nl. Pupae were returned to a humid chamber and monitoredat 5 min intervals. When the role of cGMP was being tested,injections were performed as described above using 2 mmol l−1
8-Br-cGMP.
Ligations
Pupae were mounted on double-sided sticky tape so that thehead extended past the edge of a glass microscope slide. Thepuparial case was dissected away, leaving the head and neckexposed. A blood-tight ligature was placed around the neckusing very fine monofilament nylon line, and the head wasquickly cut open and the brain removed to eliminate thepossible effect of incomplete ligations. Animals were returnedto a humid chamber and monitored periodically for eclosionbehavior. In animals younger than the grainy stage, an openingwas torn in the anteriormost portion of the pupal cuticle priorto ligation, permitting the loosened pupal cuticle to seal aroundthe neck during the ligation, thereby preventing desiccation.
Observations of tracheal filling
The extent of filling of the tracheae of the central nervoussystem (CNS) with gas was observed by dissecting animalsin 4 % formaldehyde at room temperature. Maintaining thetissues at room temperature prevented the air trapped in thetracheal system from dissolving, as it will if the tissue ischilled. The central nervous systems were fixed for aminimum of 2 h, rinsed for 20–30 min in PBS and mountedon polylysine-coated coverslips in 100 % glycerol. Thenervous systems were then allowed to equilibrate with theglycerol overnight before scoring. This treatment permittedus to visualize the extent of filling of the tracheae using lightmicroscopy and differential interference contrast optics.Images were collected from a Sony CCD camera connectedto a Nikon Optiphot microscope. On average, 30 focal planesof each region of the ventral nervous system were collected,and these were collapsed into a single image using thelayering palette in Adobe Photoshop 3.0. Different regionswere then assembled into a single montage. This methodallowed us to produce single images containing informationabout the state of tracheal inflation through the entire ventralnervous system.
ResultsPhysiological and behavioral change during the end of adult
development in wild-type D. melanogaster and EH cellknockouts
Although developing flies are confined within a rigidpuparial case, they exhibit spontaneous movements that arevisible through the puparium. These movements includeperistaltic contractions of the abdomen and expansion of thefront of the head. Given the role of EH in initiating ecdysisbehavior in M. sexta, we hypothesized that the release of EHprior to eclosion might be accompanied by a change inbehavior of the pharate adults. We monitored the activity ofpupae using time-lapse video over the last 9–15 h ofdevelopment (Fig. 1). We scored only abdominalcontractions that resembled peristalsis and all expansions ofthe head. While there was variation from animal to animal,there was a consistent change in the frequency of theseactivities over time. Abdominal and head movementsoccurred frequently until approximately 4 h before ecdysis,when the insects became quiescent. Quiescence was abruptlyterminated by the resumption of activity 30–60 min beforeeclosion. Activity then decreased prior to the onset ofeclosion. Eclosion behavior itself typically lasted only a fewseconds.
Although the activity of the developing adult showed aquiescent phase between 1 and 5 h before adult eclosion, theheart rate showed no such reduction (Fig. 2, filled squares).During the last 10 h of adult development, the heart rateincreased in an approximately linear fashion, peaking atapproximately 1 h before ecdysis at the grainy/white stage. Theheart rate then fell by approximately 10–15 % as the insectentered the extended ptilinum stage, approximately 35 min

3040
before eclosion. We did not determine the heart rate at theactual time of ecdysis since the effect of restraining flies wouldhave made the observation difficult to interpret. The rise inheart rate of knockouts is similar to that seen in wild-type flies(Fig. 2, open circles), differing significantly only during thesmooth/grainy stage (P<0.002) and grainy stage (P<0.01; two-tailed t-test).
The properties of the new cuticle change rapidly duringthe period preceding ecdysis. At least part of the moltingfluid is thought to be resorbed across the new cuticle (Wolfe,1954), but prior to ecdysis the cuticle must be made
waterproof to prevent dessication of the newly ecdysedinsect. We assessed the timing of waterproofing of thecuticle simply by severing the wings of staged developingadults, removing them from the pupal cuticle and floatingthem on distilled water. During the grainy stage, the wingsrapidly absorbed water and expanded. After the entry intothe white stage, the severed wings no longer showed thisresponse. We have interpreted this transition as the result ofwaterproofing of the cuticle.
The filling of the adult tracheal system coincidesapproximately with the waterproofing of the external cuticle.The filling of the tracheae of the head is visible through theadult cuticle and characterizes the entry into the grainy/whitestage at approximately 0.9 h before eclosion (denoted −0.9 h).In dissected preparations, tracheal filling is especiallyobvious within the ventral CNS (Fig. 3). Prior to thegrainy/white stage, only scattered large tracheal trunks werefilled with air. In contrast, developing adults that had enteredthe grainy/white stage consistently (six of six) had airextending into the fine tracheolar branches that ramifiedthroughout the CNS (Fig. 3B). This extensive tracheal fillingof the CNS was then maintained into the adult stage(Fig. 3E). The filling of the trachea within the ventral nervoussystem was abnormal in knockout flies. As seen in Fig. 3F–J,the timing of tracheal filling was delayed, and the extent offilling was largely limited to the main tracheal trunks in theventral CNS.
The EH neurons show an extensive depletion of stored EHprior to adult ecdysis (Horodyski et al., 1993). The timing ofthis EH release was estimated by staining the ventral CNS withan antibody to EH and quantifying the amount of EHimmunoreactivity (EH-IR) present at successive stages. Wefound two times late in development that were characterizedby a reduction in EH-IR (Fig. 4A). The first period of EHdepletion occurred at approximately 6 h before eclosion(around the time of the smooth/grainy stage) and was evidentas a reduction in the amount of EH staining in the thoracic
J. D. BAKER, S. L. MCNABB AND J. W. TRUMAN
Ec
Num
ber
of m
ovem
ents
Time before eclosion (h)14 1312 11 9 7 6 5 4 3 2 1810 14 1312 11 9 7 6 5 4 3 2 1810
20
10
0
20
10
0
20
10
0
20
10
020
10
020
10
0
A
EcTime before eclosion (h)
B
Fig. 1. Analysis of time-lapse videorecordings of Drosophila melanogastermonitoring their activity over the last hoursof adult development. (A) Frequency ofabdominal peristaltic movements for threeanimals tabulated in 5 min bins. (B) Thefrequency of head expansion movements forthe same individuals. Ec, eclosion.
Time before adult eclosion (h)
11 10 9 7 5 4 3 2 1 0
200
250
150
100
50
0
WEP
G/W
GS/GS
Hea
rt r
ate
(bea
tsm
in-1
)
8 6
Fig. 2. Changes in heart rate during the last 11 h of adultdevelopment for wild-type (filled squares, N=20 per stage) andknockout (open circles, N=10 per stage) Drosophila melanogaster.Symbols show means ± S.E.M. Arrows along the x-axis delineate theduration of the various stages. S, smooth; S/G, smooth/grainy; G,grainy; G/W, grainy/white; W, white; EP, extended ptilinum.

3041Physiology and behavior at adult ecdysis
portion of the EH axons (Fig. 4C). The level of EH-IR thenremained unchanged through the grainy stage. EH staining wasfurther decreased at the grainy/white stage (−0.9 h) and wasnearly undetectable by ecdysis (Fig. 4A,F).
In M. sexta, the release of EH is accompanied by anincrease in immunoreactivity for cGMP in a set of centralneurons (Ewer and Truman, 1997). In D. melanogaster, weoccasionally observed the presence of high levels of cGMP
Fig. 3. The timing of air filling of thetracheae in the ventral nervous system ofdeveloping wild-type (A–E) and knockout(F–J) Drosophila melanogaster. (A) In thegrainy-stage animals (−2.9 h), the tracheaeare mostly unfilled, with the exception of thelargest tracheal elements. (B) At thegrainy/white stage (−0.9 h), the tracheae arefilled down to the finest tracheoles, and theyremain air-filled through the white (C),extended ptilinum (D) and post-eclosion (E)stages. (F–J) Knockouts fail to fill theirtracheoles completely at any stage examined:(F) grainy stage, (G) white stage, (H,I)extended ptilinum, (J) newly eclosed. Asindicated by the arrows, the −0.90 time pointis missing for the knockouts because theylack a grainy/white stage. Two examples thatspan the range of filling that was observed arepresented for the extended ptilinum timepoint. The nervous system from the post-eclosion knockouts (J) is the sample with thegreatest extent of filling. The lower range wassimilar to that of H. A minimum of sixnervous systems was observed for each stageand genotype. Scale bar, 50 µm.
Fig. 4. Developmental timing of depletion of eclosionhormone (EH) from the ventral central nervous system(CNS) of Drosophila melanogaster. (A) Measurement ofthe area of EH staining in EH cell axons of the ventralCNS at various stages using confocal microscopy andimaging techniques. (B–F), Examples of ventral centralnervous systems immunostained for EH at differentstages before eclosion. Values in A are means based on6–10 ventral central nervous systems for each stage, andthe error bars represent the S.E.M. By analysis ofvariance, af, bf, P<0.001; ab, P<0.005; ac, cf, P<0.01;difference between other means are not significant. Scalebar, 20 µm. S, smooth; S/G, smooth/grainy; G, grainy;G/W, grainy/white; W, white; EP, extended ptilinum; Ec,eclosion. Ec
Ec

3042
in neurons and tracheae. In grainy-stage pharate adults (−2.9to −1 h), just prior to the second reduction in EH staining, weobserved elevated levels of cGMP in both the neurons andthe tracheae (Fig. 5A–C) in five of 15 nervous systems. Thisincrease in cGMP level was transient and was rarely observedin other pre-ecdysial stages, although it was evident at thetime of adult eclosion. The most prominent cGMP-immunoreactive neurons we observed were a group of fourlarge cells in the abdominal CNS, which appear to be thosedescribed by Ewer and Truman (1996) at eclosion. Wetypically saw labeling in 1–4 of these neurons, at least one ofwhich projected out from the abdominal nerve trunk. Theywere not immunoreactive for CCAP, for pigment-dispersinghormone or for small cardioactive peptide (data not shown,five ventral nervous systems for each), and their identityremains uncertain.
The tracheae were also cGMP-immunoreactive, withstaining typically confined to the larger tracheal trunks.Staining was often restricted to the area around the nuclei ofthe tracheal cells. In one case, we observed tracheal cGMPstaining only in proximity to the EH cell axons (Fig. 5C). Inthis micrograph, a projection of five sections taken at 1 µmintervals, the EH axons can be seen passing directly over thecGMP-positive trachea.
Endocrine induction of ecdysis-related physiology in wild-type and EH cell knockout flies
Injecting EH from M. sexta into D. melanogaster wasunsuccessful in evoking precocious ecdysis behavior (J. D.Baker, S. L. McNabb and J. W. Truman, unpublished data).This may be due to the failure of EH to cross the blood–brainbarrier (Gammie and Truman, 1997) or to an inability of D.melanogaster receptors to bind the moth peptide. In contrast,MasETH is very effective in causing precocious eclosion in D.melanogaster (McNabb et al., 1997). When injected into wild-type animals at the grainy stage (approximately 3 h beforeecdysis), a dose of 0.02 pmol or more of MasETH resulted in
the onset of ecdysis within approximately 50 min (Fig. 6). Asreported earlier, the knockout flies are able to eclose, but theyshow no behavioral response to injection of MasETH (McNabbet al., 1997).
Assessment of the physiological actions of MasETH in M.sexta is complicated by its ability to cause release ofendogenous stores of EH (Ewer et al., 1997). The samecomplication also appears to exist in D. melanogaster. Fig. 7shows the changes in EH immunoreactivity in the ventralCNS following injection of MasETH into flies at the grainystage. At 10 min after injection, EH staining was similar topre-injection levels (Fig. 7A,C), but a major decrease thenoccurred between 20 and 30 min post injection (Fig. 7A,D,E).Saline injections had no effect on the levels of EH-IR overthis period (N=14). To discriminate between direct responsesto MasETH and those that were mediated through EH release,we examined responses in both wild-type and EH cellknockouts. Injections of 0.05–0.15 pmol of MasETHaccelerated physiological events associated with ecdyses. Inwild-type flies injected at the grainy stage, heart rate waselevated by 85 beats min−1 within 10 min of MasETHinjection (Fig. 8). At this stage, the knockouts had a heart rateslightly higher than that of controls (Fig. 8, see also Fig. 2),but MasETH injection had no effect on their heart rate(Fig. 8).
Injection of MasETH induced precocious tracheal filling inthe ventral CNS within 15 min in six of seven individuals.Unlike wild-type flies, injection of MasETH into knockouts atthe grainy stage was ineffective in inducing premature trachealfilling around the ventral CNS (see Fig. 11A) or in the trachealsacs overlying the brain (data not shown, N>50).
In M. sexta, EH acts on selected central neurons via anincrease in intracellular cGMP (Gammie and Truman, 1999).
J. D. BAKER, S. L. MCNABB AND J. W. TRUMAN
Fig. 5. Staining for cGMP in the ventral central nervous system(CNS) of Drosophila melanogaster at the grainy stage. (A) cGMPimmunoreactivity (cGMP-IR) in the tracheae (arrowhead) or neurons(arrows). The inset is a detail of one tracheal element. (B) A singleneuron. (C) The eclosion hormone (EH) axons are visible (arrow)and pass directly over the cGMP-positive tracheal element(arrowhead). In C, the CNS was double-stained for both EH andcGMP. The diffuse speckled staining is background consistentlypresent at these stages.
500
250
200
150
100
50
0
Lat
ency
(m
in)
Amount of MasETH injected (pmol)0.001 0.01 0.1 1 10
Fig. 6. Dose–response relationship showing the sensitivity ofDrosophila melanogaster to MasETH. Pharate adults at the grainystage were injected with varying concentrations of MasETH or withsaline. Open circles are for saline-injected flies, filled circles are forMasETH-injected flies. The line is fitted by eye through the meanvalues for the MasETH-injected animals. Injection of 0.02 pmol ofMasETH was sufficient to induce premature ecdysis.
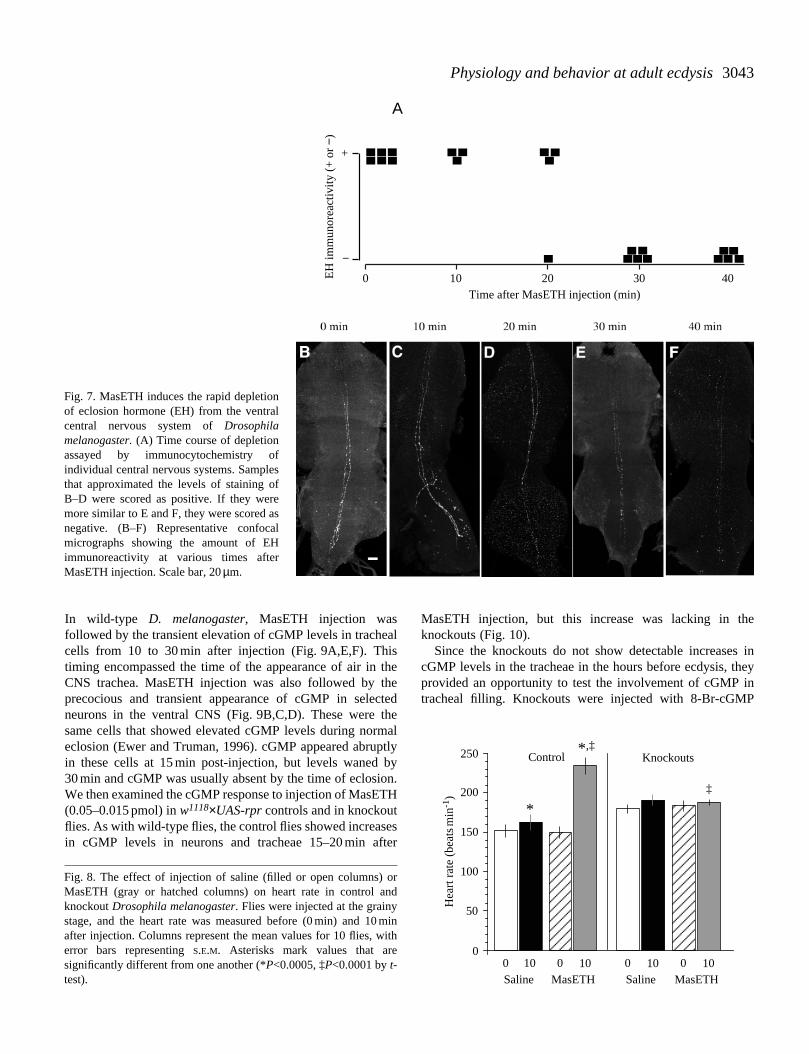
3043Physiology and behavior at adult ecdysis
In wild-type D. melanogaster, MasETH injection wasfollowed by the transient elevation of cGMP levels in trachealcells from 10 to 30 min after injection (Fig. 9A,E,F). Thistiming encompassed the time of the appearance of air in theCNS trachea. MasETH injection was also followed by theprecocious and transient appearance of cGMP in selectedneurons in the ventral CNS (Fig. 9B,C,D). These were thesame cells that showed elevated cGMP levels during normaleclosion (Ewer and Truman, 1996). cGMP appeared abruptlyin these cells at 15 min post-injection, but levels waned by30 min and cGMP was usually absent by the time of eclosion.We then examined the cGMP response to injection of MasETH(0.05–0.015 pmol) in w1118×UAS-rpr controls and in knockoutflies. As with wild-type flies, the control flies showed increasesin cGMP levels in neurons and tracheae 15–20 min after
MasETH injection, but this increase was lacking in theknockouts (Fig. 10).
Since the knockouts do not show detectable increases incGMP levels in the tracheae in the hours before ecdysis, theyprovided an opportunity to test the involvement of cGMP intracheal filling. Knockouts were injected with 8-Br-cGMP
0 10 20 40Time after MasETH injection (min)
EH
imm
unor
eact
ivity
(+
or
−)
+
−
A
30
Fig. 7. MasETH induces the rapid depletionof eclosion hormone (EH) from the ventralcentral nervous system of Drosophilamelanogaster. (A) Time course of depletionassayed by immunocytochemistry ofindividual central nervous systems. Samplesthat approximated the levels of staining ofB–D were scored as positive. If they weremore similar to E and F, they were scored asnegative. (B–F) Representative confocalmicrographs showing the amount of EHimmunoreactivity at various times afterMasETH injection. Scale bar, 20 µm.
250
200
150
100
50
0
Hea
rt r
ate
(bea
tsm
in-1
)
0 10
Control Knockouts *,‡
*‡
MasETH0 10Saline
0 10MasETH
0 10Saline
Fig. 8. The effect of injection of saline (filled or open columns) orMasETH (gray or hatched columns) on heart rate in control andknockout Drosophila melanogaster. Flies were injected at the grainystage, and the heart rate was measured before (0 min) and 10 minafter injection. Columns represent the mean values for 10 flies, witherror bars representing S.E.M. Asterisks mark values that aresignificantly different from one another (*P<0.0005, ‡P<0.0001 by t-test).

3044
(160–600 pmol), a membrane-permeant analog. When 8-Br-cGMP was injected into knockout flies at the grainy stage,three of six animals responded by filling their tracheoles withair (Fig. 11B,C). When injected with 8-Br-cGMP at the white
stage, all six animals responded with tracheal filling afterinjection with 8Br-cGMP (Fig. 11D).
Development of the ecdysis response system
The development of the ecdysis control system was assessedin two ways: (1) by testing the ability of developing animalsto respond to exogenous ETH and (2) by using neck ligationto remove the source of cephalic signals at various times indevelopment.
To assay the developmental sensitivity of D. melanogasterto MasETH, we injected developing wild-type adults of avariety of stages (Fig. 12). Flies first became responsive toMasETH at 6 h prior to eclosion (−6 h). These animalsresponded with strong and persistent bouts of ecdysis behavior,which propelled them partially out of the puparium, but theycould not successfully escape from it. The premature ecdysisbouts began before the animals had resorbed their molting fluidor inflated their head tracheae. Their inability to escape fromthe puparium may have been due to the unresorbed moltingfluid that remained between the adult and pupal cuticle. Mostanimals injected at −4 h or later were able to eclosesuccessfully. Flies injected at or after −0.9 h were unaffectedby MasETH injection, presumably because they had alreadyreleased their own EH (see Fig. 4A).
To assess the role of head-derived factors in controllingeclosion behavior, we neck-ligated pharate adults at varioustimes prior to eclosion (Fig. 13A). Ligation at the mid-smooth/grainy stage inhibited eclosion in all but twoindividuals. Developing adults ligated at the grainy stage(−2.9 h before ecdysis) invariably eclosed, although eclosionwas delayed by between 20 min and almost 5 h. Developingflies ligated at the grainy/white or white stages typicallyecdysed on schedule or earlier. Interestingly, at the extendedptilinum stage, flies ecdysed immediately (<1 min) after neckligation.
Similar ligation experiments were carried out on developing
J. D. BAKER, S. L. MCNABB AND J. W. TRUMAN
Tracheae
A
B
Time after MasETH injection (min)
cGM
P im
mun
orea
ctiv
ity (
+ o
r −)
−
+
−
+
0 10 15 20 30 40
Neurons
0 10 15 20 30 40
Fig. 9. The time course of elevation of cGMP levels in the tracheae(A) and neurons (B) of the ventral central nervous system (ventralCNS) following MasETH injection in Drosophila melanogaster.Any staining in a tracheal cell or neuron was scored as a positive.(C–F) Confocal images showing examples of cGMP expression inventral CNS cells following MasETH injections. (C) 10 min afterinjection of MasETH, both tracheae (arrowhead) and neurons(arrow) are immunopositive for cGMP. The inset shows thearborization of the neuron. (D) After 15 min, 1–4 neurons in theabdominal ganglion frequently show elevated levels of cGMP. Thetracheae are typically cGMP-positive 15 min (E) and 20 min (F) afterinjection. In E, the arrowheads show staining for cGMP that isconfined to the region around the nucleus of the tracheal cell. Scalebar, 20 µm.
Tracheae Neurons
0
20
40
60
80
100 16
12
14
16
12
14Ner
vous
sys
tem
s w
ithcG
MP-
posi
tive
cells
(%
)
Fig. 10. Immunoreactivity for cGMP of wild-type (hatchedcolumns), controls (w1118×UAS-rpr) (open columns) and knockout(filled columns) Drosophila melanogaster 15–20 min after injectionof MasETH. The percentage of central nervous systems with cGMP-immunoreactive tracheae or neurons is plotted, with the number offlies shown above each column.

3045Physiology and behavior at adult ecdysis
knockout flies. Very few (seven of 43) of the knockoutsattempted to ecdyse after neck ligation, and there was no stage-dependent pattern to these ecdyses (Fig. 13B). Especiallyinteresting were the flies ligated at the extended ptilinum stage.In control genotypes, this always resulted in immediateecdysis, but in the knockouts only two of 11 flies showedecdysis behavior and they eclosed after more than 90 min. Theresponses of ligated animals of the control genotype(w1118×UAS-rpr) were indistinguishable from the wild type.
DiscussionPhysiological and behavioral changes associated with
eclosion
Although eclosion is the most dramatic event associatedwith the end of adult development, it is only one event in asequence of physiological and behavioral changes that beginsa number of hours earlier. Despite being enclosed in a rigidpuparium, developing flies are surprisingly active during the
late phases of adult development. They then enter a period ofquiescence approximately 4–5 h before eclosion, followed bya transient phase of high activity that appears 30–60 min beforeeclosion, just after the final depletion of EH. Eclosion itselfoccurs so rapidly, within a matter of seconds, that it oftenappears inconspicuous on a compressed time-scale such as thatused in Fig. 1.
The behavior shown by developing D. melanogaster maycorrespond to some of the late metamorphic behavioralresponses shown by moths. The transient period of highactivity that immediately precedes eclosion may correspond tothe pre-eclosion behavior shown by giant silkmoths such asHyalophora cecropia (Truman, 1971). As in H. cecropia, thetiming of this activity period in D. melanogaster coincides witha major EH release. In M. sexta, there is an additional bout ofactivity that occurs approximately 10–12 h earlier and is thenfollowed by quiescence until the time of EH release. Thispreparatory behavior is independent of the brain and isregulated by the declining ecdysteroid titer (Truman, 1984).We have not attempted to determine the mechanismscontrolling the early activity period in D. melanogaster, but itsdevelopmental timing is similar to the preparatory behavior ofM. sexta and could be regulated, either directly or indirectly,by the declining ecdysteroid titer.
In flies, adult ecdysis is accompanied and followed bymarked increases in blood pressure. These blood pressureincreases are used in ptilinal expansion during ecdysis itselfand again in the post-ecdysis phase for wing expansion (Zdareket al., 1984). Since these animals have an open circulatorysystem and cannot, therefore, generate significant changes ininternal pressure using the heart, these major increases in bloodpressure are due to contraction of the abdominal body wallmuscles, especially the tergosternal muscles. The heart,however, does show a striking increase in beat rate over the10 h preceding adult eclosion. Compared with wild-type flies,the knockouts showed a similar pattern of increase in heart rateover this period of development, indicating that most of thiselevation of heart rate does not require EH. We did find that,towards the end of adult development, injection with MasETH
Fig. 11. Responses of the tracheae ofknockout Drosophila melanogaster toinjection of MasETH or 8-Br-cGMP. (A) Tracheal filling after injection ofMasETH into grainy-stage knockout flies. Ofthe six flies injected, none showed trachealfilling. (B,C) Injection of 8-Br-cGMP at thegrainy stage. These two ventral nervoussystems span the range of filling afterinjection at this time. Three of the six fliesinjected contained air in the tracheoles. (D) Injection of 8-Br-cGMP at the whitestage (−0.80 h) led to filling in all instances(N=6). Scale bar, 50 µm.
100
80
60
40
20
0
Perc
enta
ge r
espo
ndin
g or
ecl
osin
g
Time before eclosion (h)
10 9 8 7 6 5 4 3 2 1 0
Fig. 12. The onset of sensitivity of developing adults to MasETH.Drosophila melanogaster of varying stages were injected with2–4 pmol of MasETH. Squares show the percentage of animals thatexhibited sustained eclosion behavior. Filled circles show thepercentage of animals that successfully shed the old cuticle. N=10 foreach data point.

3046
induced a 50 % increase in heart rate. However, this MasETH-induced increase appears to be dependent on the EH cells sinceit did not occur in the knockouts. Such a possible dependenceon EH may, however, also be indirect. A number ofcardioactive peptides are known to be released at each ecdysisin M. sexta, and these are thought to act directly on the heart(Tublitz and Truman, 1985a–c). The most prominent of theseis CCAP, which was first identified because of itscardioacceleratory effects (Stangier et al., 1987) and nowappears to be the main neuropeptide responsible for causingecdysis behavior (Gammie and Truman, 1997).
Control over filling of the tracheae during eclosion
While the filling of the tracheae with air at ecdysis has beendescribed for a number of species and molts (for reviews, see
Wigglesworth, 1983; Manning and Krasnow, 1993), the factorsunderlying the regulation of this process are unknown. Severallines of evidence suggest that tracheal filling is caused, eitherdirectly or indirectly, by EH. First, tracheal filling and EHrelease are coincident; they both occur at the grainy/whitestage. Second, at the time that the tracheae are filling, thetracheal cells show an elevation in intracellular cGMP levelsdetectable by immunocytochemistry. In other tissues, such asthe epitracheal glands (Ewer et al., 1997; Kingan et al., 1997)and the transverse nerve (Morton, 1996) of M. sexta, EHinduces an increase in intracellular cGMP levels in target cells.Third, both tracheal filling and the appearance of cGMP intracheal cells can be induced prematurely in response toMasETH injection, but only in flies that possess EH neurons.In the absence of EH cells, tracheal filling is both delayed andincomplete, never fully expanding into the fine tracheoles.Finally, tracheal filling in the knockouts can be induced byinjection of a membrane-permeant analog of cGMP. Together,this evidence supports the idea that EH acts to cause the rapidfilling of the new tracheae by inducing an elevation of cGMPlevels in the tracheal cells. The mechanism of rapid trachealfilling is unknown, but one possibility is that the cGMP acts toincrease the rate of molting fluid transport across the tracheolarepithelium, thereby facilitating the filling of the tracheae andtracheoles with air (Wigglesworth, 1953). At present, the best-described example of a role for cGMP in fluid transport is inthe Malphigian tubules of D. melanogaster (Davies et al.,1995). In that case, a peptide (CAP2b) indirectly stimulates anelevation of cGMP level which, in turn, leads to the rapidsecretion of fluid into the lumen of the tubules. A secondmechanism that might facilitate tracheal filling may relate tothe waterproofing of the external cuticle that occurs at thissame time (Wolfe, 1954). Similar changes occurring within thetracheae to reduce their wetability would also aid the lumen infilling with gas.
One recently described mutation in a presumptivemitochondrial transport protein causes defects in trachealfilling and in wing expansion (Hartenstein et al., 1997). Whilethe relationship between this defect and the hormonallytriggered events we describe here has not been established, thefurther application of molecular genetic analyses to eclosionand post-eclosion behavior and physiology will undoubtedlyhelp clarify the cellular and physical properties that contributeto this process.
Timing of EH release
In other insects, it is thought that a single release of eclosionhormone occurs at the end of the molt (Reynolds et al., 1979;Ewer et al., 1997). In D. melanogaster, at adult ecdysis, twolines of evidence suggest that, in addition to the release thatimmediately precedes eclosion, a second earlier release mayoccur several hours before adult ecdysis.
One line of evidence involves using animmunocytochemical approach to estimate the relative levelsof EH at various times before eclosion. There appears to be EHdepletion 4–7 h prior to eclosion as well as a release that occurs
J. D. BAKER, S. L. MCNABB AND J. W. TRUMANT
ime
to e
clos
ion
(h)
0
1
2
3
4
5
Tim
e to
ecl
osio
n (h
)
EPWG/WG
S/G
A
B
EPWG/WGS/GTime of ligation
0
1
2
3
4
5
16
>24
>24
Fig. 13. Importance of head-derived factors for eclosion behavior inDrosophila melanogaster. (A) Latency to eclosion following neckligation of wild-type pharate adults. The mean latency for the 16animals at the extended ptilinum time point is 1 min. (B) Latency toeclosion following neck ligation for control flies (w1118×UAS-rpr,filled circles) and knockouts (open triangles). Symbols above the 5 htime level indicate animals that remained alive (determined by arobust heartbeat) but failed to eclose after 1 day. The diagonal linerepresents the mean latency to eclosion for unligated wild-typeanimals of the various stages (from A), and the vertical lines arestandard deviations. Data drawn from Kimura and Truman (1990).Each symbol represents a single individual. S, smooth; S/G,smooth/grainy; G, grainy; G/W, grainy/white; W, white; EP,extended ptilinum.

3047Physiology and behavior at adult ecdysis
less than 1 h before eclosion. The reduction in staining at theearly period was typically confined to the rostral portion of theEH axons. We cannot, however, rule out the possibility thatthe change in staining reflects a change in the axonallocalization of EH due to transport to more caudal release sites,rather than to an actual release.
The second line of evidence involves the neck ligationexperiments. Neck ligation of flies has been used to study thetiming and the cephalic basis of physiological events followingeclosion (Kimura and Truman, 1990) and led to the discoveryof the cuticular tanning hormone bursicon (Cottrell, 1962;Fraenkel and Hsiao, 1962). Our ligation experiments alsosuggest that EH may be released at two times. Early neckligation during the smooth/grainy stage (−5 h) preventssubsequent ecdysis (Fig. 13A). By the grainy stage (−3 h), achange has occurred in the flies that enables them to eclosedespite losing their head. All wild-type flies eclose after neckligation at this time, although the latencies extend up to 5 h.Since the knockouts did not become competent to ecdyse asthey passed from the smooth/grainy to the grainy stage(Fig. 13B), this competence appears to involve the EHneurons. One possibility is that competence is due to an initialrelease of EH. An alternative hypothesis, however, is that theexcitability of the EH neurons may be increasing at this time(as occurs in M. sexta; Hewes and Truman, 1994) and that theisolated axons become capable of releasing enough EH afterligation to cause eventual ecdysis. The latter interpretationwould also be favored by our failure to detect a cGMP responsein neurons around the time of the first putative release.
The response of the knockouts to ligations shows that theability of animals to ecdyse after neck ligation depends heavilyon a product of the EH cells. Even at the extended ptilinumstage, when wild-type flies would ecdyse immediately afterneck ligation, this rapid response to neck ligation was not seenin the knockouts. This suppression is especially strikingconsidering that some of these flies would normally havebegun ecdysis on their own within the next few minutes. Theseobservations on the knockout flies suggest that, in addition tothat provided by the EH neurons, there are other signals fromthe brain that promote ecdysis. When these signals work alone,as in the case of the knockout flies, the ecdysis behavior isweaker and less organized (McNabb et al., 1997).
Control of the expression of ecdysis behavior
Developing adults neck-ligated after the grainy stage wereable to ecdyse and did so as fast or faster than unligaturedcontrols. This was particularly apparent in the insects ligatedat the extended ptilinum stage. These flies ecdysed within1 min of ligation, an advance of up to 40 min (Kimura andTruman, 1990). Similar results have been obtained for adultecdysis in M. sexta (Ewer and Truman, 1997). In the latter,there is normally a 2–3 h delay between EH release andecdysis, but decapitation at any time during that period leadsto immediate ecdysis. These results suggest that EH activatesecdysis-controlling cells in the ventral CNS (e.g. the CCAP-containing neurons), but that there are descending influences
from the head that inhibit this activation. Decapitationinterrupts the inhibition, resulting in the rapid onset of ecdysisbehavior.
We think it is significant that, of the small percentage ofknockout flies that attempted ecdysis after decapitation, noneshowed a rapid ecdysis response that could be interpreted asthe release of descending suppression. Consequently, we thinkthat EH release activates ecdysis-promoting cells in the ventralCNS (possibly the CCAP neurons), but that it also co-activatesdescending neurons that inhibit these cells. Ecdysis wouldfinally occur after the inhibition had waned. This relationshipestablishes a delay between EH release and the onset ofecdysis. Since EH is also involved in peripheral processes suchas tracheal filling and cuticle waterproofing, this delay providesa window for their completion prior to the final emergence ofthe insect.
We gratefully thank Jan De Vente for the anti-cGMPantiserum, K. Ranga Rao for the anti-PDH antiserum, Hans-Jurgen Agricola for the anti-CCAP antiserum, and A. O. D.Willows for the anti-SCP monoclonal antibody. This researchwas funded by NSF grant IBN-9604670 (J.W.T.) and NIHTraining Grant T32 HD07183 (J.D.B.).
ReferencesBainbridge, S. P. and Bownes, M. (1981). Staging the
metamorphosis of Drosophila melanogaster. J. Embryol. Exp.Morph. 66, 57–80.
Brand, A. and Perrimon, N. (1993). Targeting gene expression as ameans of altering cell fates and generating dominant phenotypes.Development 118, 401–415.
Cottrell, C. B. (1962). The imaginal ecdysis of blowflies: detectionof the blood-borne darkening factor and determination of some ofits properties. J. Exp. Biol. 39, 413–430.
Davies, S. A., Huesmann, G. R., Maddrell, S. H. P., O’Donnell,M. J., Skaer, N. J. V., Dow, J. A. T. and Tublitz, N. J. (1995).CAP(2b), a cardioacceleratory peptide, is present in Drosophila andstimulates tubule fluid secretion via cGMP. Am. J. Physiol. 38,R1321–R1326.
Dircksen, H. and Keller, R. (1988). Immunocytochemicallocalization of CCAP, a novel crustacean cardioactive peptide, inthe nervous system of the shore crab, Carcinus maenas L. CellTissue Res. 254, 347–360.
Dircksen, H., Zahnow, C. A., Gaus, G., Keller, R., Rao, K. R. andRiehm, J. P. (1987). The ultrastructure of nerve endings containingpigment-dispersing hormone (PDH) in crustacean sinus glands:Identification by an antiserum against a synthetic PDH. Cell TissueRes. 250, 377–387.
Ephrussi, B. and Beadle, G. W. (1936). A technique oftransplantation for Drosophila. Am. Nat. 70, 218–225.
Ewer, J., Gammie, S. C. and Truman, J. W. (1997). Control ofinsect ecdysis by a positive-feedback endocrine system: roles ofeclosion hormone and ecdysis triggering hormone. J. Exp. Biol.200, 869–881.
Ewer, J. and Truman, J. W. (1996). Increases in cyclic 3′,5′-guanosine monophosphate (cGMP) occur at ecdysis in anevolutionarily conserved crustacean cardioactive peptide-

3048
immunoreactive insect neuronal network. J. Comp. Neurol. 370,330–341.
Ewer, J. and Truman, J. W. (1997). Invariant association of ecdysiswith increases in cyclic 3′,5′-guanosine monophosphateimmunoreactivity in a small network of peptidergic neurons in thehornworm, Manduca sexta. J. Comp. Physiol. A 181, 319–330.
Fraenkel, G. and Hsiao, C. (1962). Hormonal and nervous controlof tanning in the fly. Science 138, 27–29.
Gammie, S. C. and Truman, J. W. (1997). Neuropeptide hierarchiesand the activation of sequential motor behaviors in the hawkmoth,Manduca sexta. J. Neurosci. 17, 4389–4397.
Gammie, S. C. and Truman, J. W. (1999). Eclosion hormoneprovides a link between ecdysis triggering hormone and crustaceancardioactive peptide in the neuroendocrine cascade that controlsecdysis behavior. J. Exp. Biol. 202, 343–352.
Hartenstein, K., Sinha, P., Mishra, A., Schenkel, H., Torok, I.and Mechler, B. M. (1997). The congested-like tracheae geneof Drosophila melanogaster encodes a member of themitochondrial carrier family required for gas-filling of thetracheal system and expansion of the wings after eclosion.Genetics 147, 1755–1768.
Hewes, R. S. and Truman, J. W. (1994). Steroid regulation ofexcitability in identified insect neurosecretory cells. J. Neurosci. 14,1812–1819.
Horodyski, F. M., Ewer, J., Riddiford, L. M. and Truman, J. W.(1993). Isolation, characterization and expression of the eclosionhormone gene of Drosophila melanogaster. Eur. J. Biochem. 215,221–228.
Kimura, K. and Truman, J. W. (1990). Postmetamorphic cell deathin the nervous and muscular systems of Drosophila melanogaster.J. Neurosci. 10, 403–411.
Kingan, T. G., Gray, W., Zitnan, D. and Adams, M. E. (1997).Regulation of ecdysis-triggering hormone release by eclosionhormone. J. Exp. Biol. 200, 3245–3256.
Manning, G. and Krasnow, M. A. (1993). Development of theDrosophila tracheal system. In The Development of Drosophilamelanogaster (ed. M. Bate and A. Martinez-Arias), pp. 609–686.Cold Spring Harbor, NY: Cold Spring Harbor Press.
McNabb, S. L., Baker, J. D., Agapite, J., Steller, H., Riddiford, L.M. and Truman, J. W. (1997). Disruption of a behavioralsequence by targeted death of peptidergic neurons in Drosophila.Neuron 19, 813–823.
Morton, D. B. (1996). Neuropeptide-stimulated cyclic guanosinemonophosphate immunoreactivity in the neurosecretory terminalsof a neurohemal organ. J. Neurobiol. 29, 341–353.
Nordstrom, W., Chen, P., Steller, H. and Abrams, J. M. (1996).Activation of the reaper gene during ectopic cell killing inDrosophila. Dev. Biol. 180, 213–226.
O’Brien, M. A. and Taghert, P. H. (1998). A peritrachealneuropeptide system in insects: release of myomodulin-likepeptides at ecdysis. J. Exp. Biol. 201, 193–209.
Reynolds, S. E., Taghert, P. H. and Truman, J. W. (1979). Eclosionhormone and bursicon titres and the onset of hormonalresponsiveness during the last day of adult development inManduca sexta (L.). J. Exp. Biol. 78, 77–86.
Riddiford, L. M. (1993). Hormone receptors and the regulation ofinsect metamorphosis. Am. Zool. 33, 340–347.
Stangier, J., Hilbich, C., Beyreuther, K. and Keller, R. (1987).Unusual cardioactive peptide (CCAP) from pericardial organs ofthe shore crab Carcinus maenas. Proc. Natl. Acad. Sci. USA 84,575–579.
Truman, J. W. (1971). Physiology of insect ecdysis. I. The eclosionbehavior of saturniid moths and its hormonal release. J. Exp. Biol.54, 805–814.
Truman, J. W. (1984). The preparatory rhythm of the moth Manducasexta: an ecdysteroid-triggered circadian rhythm that isindependent of the brain. J. Comp. Physiol. A 155, 521–528.
Truman, J. W. (1992). The eclosion hormone system of insects.Prog. Brain Res. 92, 361–374.
Truman, J. W. and Riddiford, L. M. (1970). Neuroendocrinecontrol of ecdysis in silkmoths. Science 167, 1624–1626.
Tublitz, N. J. and Truman, J. W. (1985a). Insect cardioactivepeptides. I. Distribution and molecular characteristics of twocardioacceleratory peptides in the tobacco hawkmoth, Manducasexta. J. Exp. Biol. 114, 365–379.
Tublitz, N. J. and Truman, J. W. (1985b). Insect cardioactivepeptides. II. Neurohormonal control of heart activity by twocardioacceleratory peptides in the tobacco hawkmoth, Manducasexta. J. Exp. Biol. 114, 381–395.
Tublitz, N. J. and Truman, J. W. (1985c). Identification of neuronscontaining cardioacceleratory peptides (CAPs) in the ventral nervecord of the tobacco hawkmoth, Manduca sexta. J. Exp. Biol. 116,395–410.
Wigglesworth, V. B. (1953). Surface forces in the tracheal system ofinsects. Q. J. Microsc. Sci. 94, 507–522.
Wigglesworth, V. B. (1983). The physiology of the insect tracheoles.Adv. Insect Physiol. 17, 85–148.
Wolfe, L. S. (1954). Studies of the development of the imaginalcuticle of Calliphora erythrocephala. Q. J. Microsc. Sci. 95,67–78.
Zdarek, J., Zavadilova, J., Su, J. and Fraenkel, G. (1984). Post-eclosion behavior of flies after emergence from the puparium. ActaEnt. Bohemoslov 81, 161.
Zitnan, D., Kingan, T. G., Hermesman, J. L. and Adams, M. E.(1996). Identification of ecdysis-triggering hormone from anepitracheal endocrine system. Science 271, 88–91.
J. D. BAKER, S. L. MCNABB AND J. W. TRUMAN