physiology - booksca.ca
TRANSCRIPT


PhysiologySEVENTHEDITION

PhysiologySEVENTHEDITION
LindaS.Costanzo,Ph.D.ProfessorofPhysiologyandBiophysicsSchoolofMedicineVirginiaCommonwealthUniversityRichmond,Virginia

AcquisitionsEditor:CrystalTaylorDevelopmentEditor:AndreaVosburghEditorialCoordinator:TimRinehartMarketingManager:MichaelMcMahonProductionProjectManager:MarianBellusDesignCoordinator:HollyMcLaughlinManufacturingCoordinator:MargieOrzechPrepressVendor:SPiGlobal
Seventhedition
Copyright©2019WoltersKluwer
Copyright©2015,2011,2007,2003,1998,1995WoltersKluwerHealth.Allrightsreserved.Thisbookisprotectedbycopyright.Nopartofthisbookmaybereproducedortransmittedinanyformorbyanymeans,includingasphotocopiesorscanned-inorotherelectroniccopies,orutilizedbyanyinformationstorageandretrievalsystemwithoutwrittenpermissionfromthecopyrightowner,exceptforbriefquotationsembodiedincriticalarticlesandreviews.MaterialsappearinginthisbookpreparedbyindividualsaspartoftheirofficialdutiesasU.S.governmentemployeesarenotcoveredbytheabove-mentionedcopyright.Torequestpermission,pleasecontactWoltersKluweratTwoCommerceSquare,2001MarketStreet,Philadelphia,PA19103,[email protected],orviaourwebsiteatlww.com(productsandservices).
987654321
PrintedinChina
LibraryofCongressCataloging-in-PublicationDataNames:Costanzo,LindaS.,1947-author.Title:Physiology/LindaS.Costanzo.Othertitles:Boardreviewseries.Description:Seventhedition.|Philadelphia:WoltersKluwer,[2019]|Series:BRS|Includesindex.Identifiers:LCCN2017054852|ISBN9781496367617Subjects:|MESH:PhysiologicalPhenomena|ExaminationQuestionsClassification:LCCQP40|NLMQT18.2|DDC612.0076—dc23LCrecordavailableathttps://lccn.loc.gov/2017054852
Thisworkisprovided“asis,”andthepublisherdisclaimsanyandallwarranties,expressorimplied,includinganywarrantiesastoaccuracy,comprehensiveness,orcurrencyofthecontentofthiswork.
Thisworkisnosubstituteforindividualpatientassessmentbaseduponhealthcareprofessionals’examinationofeachpatientandconsiderationof,amongotherthings,age,weight,gender,currentorpriormedicalconditions,medicationhistory,laboratorydataandotherfactorsuniquetothepatient.Thepublisherdoesnotprovidemedicaladviceorguidanceandthisworkismerelyareferencetool.Healthcareprofessionals,andnotthepublisher,aresolelyresponsiblefortheuseofthisworkincludingallmedicaljudgmentsandforanyresultingdiagnosisandtreatments.
Givencontinuous,rapidadvancesinmedicalscienceandhealthinformation,independentprofessionalverificationofmedicaldiagnoses,indications,appropriatepharmaceuticalselectionsanddosages,andtreatmentoptionsshouldbemadeandhealthcareprofessionalsshouldconsultavarietyofsources.Whenprescribingmedication,healthcareprofessionalsareadvisedtoconsulttheproductinformationsheet(themanufacturer’spackageinsert)accompanyingeachdrugtoverify,amongotherthings,conditionsofuse,warningsandsideeffectsandidentifyanychangesindosagescheduleorcontraindications,particularlyif
themedicationtobeadministeredisnew,infrequentlyusedorhasanarrowtherapeuticrange.Tothemaximumextentpermittedunderapplicablelaw,noresponsibilityisassumedbythepublisherforanyinjuryand/ordamagetopersonsorproperty,asamatterofproductsliability,negligencelaworotherwise,orfromanyreferencetoorusebyanypersonofthiswork.
LWW.com

ForRichardAnd
forDan,Rebecca,andSheilaAnd
forEliseandMax

Preface
Thesubjectmatterofphysiologyis thefoundationof thepracticeofmedicine,and a firm grasp of its principles is essential for the physician. This book isintended to aid the student preparing for theUnited StatesMedical LicensingExamination (USMLE) Step 1. It is a concise review of key physiologicprinciples and is intended to help the student recallmaterial taught during thefirst and second years of medical school. It is not intended to substitute forcomprehensivetextbooksorforcoursesyllabi,althoughthestudentmayfinditausefuladjuncttophysiologyandpathophysiologycourses.
The material is organized by organ system into seven chapters. The firstchapter reviews general principles of cellular physiology. The remaining sixchapters review the major organ systems—neurophysiology, cardiovascular,respiratory,renalandacid–base,gastrointestinal,andendocrinephysiology.
Difficult concepts are explained stepwise, concisely, and clearly, withappropriate illustrative examples and sample problems. Numerous clinicalcorrelations are included so that the student can understand physiology inrelation to medicine. An integrative approach is used, when possible, todemonstrate how the organ systems work together to maintain homeostasis.Morethan130full-colorillustrationsandflowdiagramsandmorethan50tableshelp the student visualize thematerial quickly and aid in long-term retention.Appendices contain “Key Physiology Topics for USMLE Step 1,” “KeyPhysiologyEquationsforUSMLEStep1,”and“NormalBloodValues.”
QuestionsreflectingthecontentandformatofUSMLEStep1areincludedattheendofeachchapterandinaComprehensiveExaminationattheendofthebook. These questions, many with clinical relevance, require problem-solvingskills rather than straight recall. Clear, concise explanations accompany thequestions and guide the student through the correct steps of reasoning. Thequestionscanbeusedasapretesttoidentifyareasofweaknessorasaposttesttodetermine mastery. Special attention should be given to the ComprehensiveExamination, because its questions integrate several areas of physiology andrelatedconceptsofpathophysiologyandpharmacology.
Newtothisedition:

Additionofnewfull-colorfiguresUpdatedorganizationandtextExpanded coverage of neurophysiology, and respiratory, renal,gastrointestinal,andendocrinephysiologyAdditionofnewmulti-stepquestions
BestofluckinyourpreparationforUSMLEStep1!LindaS.Costanzo,Ph.D.

Acknowledgments
IthasbeenapleasuretobeapartoftheBoardReviewSeriesandtoworkwiththestaffatWoltersKluwer.CrystalTaylorandAndreaVosburghprovidedexperteditorialassistance.
My sincere thanks to students in the School of Medicine at VirginiaCommonwealthUniversity/MedicalCollegeofVirginia,whohaveprovidedsomanyhelpfulsuggestionsforBRSPhysiology.Thanksalsotothemanystudentsfromothermedicalschoolswhohavetakenthetimetowritetomeabouttheirexperienceswiththisbook.
LindaS.Costanzo,Ph.D.

Contents
PrefaceAcknowledgments
1.CELLPHYSIOLOGYI.CellMembranesII.TransportAcrossCellMembranesIII.OsmosisIV.DiffusionPotential,RestingMembranePotential,andActionPotentialV.NeuromuscularandSynapticTransmissionVI.SkeletalMuscleVII.SmoothMuscleVIII.ComparisonofSkeletalMuscle,SmoothMuscle,andCardiacMuscleReviewTest
2.NEUROPHYSIOLOGYI.AutonomicNervousSystem(ANS)II.OrganizationoftheNervousSystemIII.SensorySystemsIV.MotorSystemsV.HigherFunctionsoftheCerebralCortexVI.Blood–BrainBarrierandCerebrospinalFluid(CSF)VII.TemperatureRegulationReviewTest
3.CARDIOVASCULARPHYSIOLOGYI.CircuitryoftheCardiovascularSystemII.HemodynamicsIII.CardiacElectrophysiologyIV.CardiacMuscleandCardiacOutputV.CardiacCycle

VI.RegulationofArterialPressureVII.MicrocirculationandLymphVIII.SpecialCirculationsIX.IntegrativeFunctionsoftheCardiovascularSystem:Gravity,Exercise,andHemorrhageReviewTest
4.RESPIRATORYPHYSIOLOGYI.LungVolumesandCapacitiesII.MechanicsofBreathingIII.GasExchangeIV.OxygenTransportV.CO2TransportVI.PulmonaryCirculationVII.V/QDefectsVIII.ControlofBreathingIX.IntegratedResponsesoftheRespiratorySystemReviewTest
5.RENALANDACID–BASEPHYSIOLOGYI.BodyFluidsII. RenalClearance,RenalBloodFlow (RBF), andGlomerularFiltrationRate(GFR)III.ReabsorptionandSecretionIV.NaClRegulationV.K+RegulationVI.RenalRegulationofUrea,Phosphate,Calcium,andMagnesiumVII.ConcentrationandDilutionofUrineVIII.RenalHormonesIX.Acid–BaseBalanceX.DiureticsXI.IntegrativeExamplesReviewTest
6.GASTROINTESTINALPHYSIOLOGYI.StructureandInnervationoftheGastrointestinalTract

II.RegulatorySubstancesintheGastrointestinalTractIII.GastrointestinalMotilityIV.GastrointestinalSecretionV.DigestionandAbsorptionVI.LiverPhysiologyReviewTest
7.ENDOCRINEPHYSIOLOGYI.OverviewofHormonesII.CellMechanismsandSecondMessengersIII.PituitaryGland(Hypophysis)IV.ThyroidGlandV.AdrenalCortexandAdrenalMedullaVI.EndocrinePancreas–GlucagonandInsulinVII.CalciumMetabolism(ParathyroidHormone,VitaminD,Calcitonin)VIII.SexualDifferentiationIX.MaleReproductionX.FemaleReproductionReviewTest
ComprehensiveExamination
APPENDIX A.KEY PHYSIOLOGY TOPICS FOR USMLESTEP1
APPENDIXB.KEYPHYSIOLOGYEQUATIONSFORUSMLESTEP1
APPENDIXC.NORMALBLOODVALUES
Index

Chapter 1 CellPhysiology
Foradditionalancillarymaterialsrelatedtothischapter,pleasevisitthePoint.
I.CELLMEMBRANESarecomposedprimarilyofphospholipidsandproteins.
A.Lipidbilayer1.Phospholipidshaveaglycerolbackbone,whichisthehydrophilic(watersoluble) head, and two fatty acid tails, which are hydrophobic (waterinsoluble).Thehydrophobictailsfaceeachotherandformabilayer.
2. Lipid-soluble substances (e.g., O2, CO2, steroid hormones) cross cellmembranesbecausetheycandissolveinthehydrophobiclipidbilayer.
3.Water-solublesubstances(e.g.,Na+,Cl−,glucose,H2O)cannotdissolveinthelipidofthemembrane,butmaycrossthroughwater-filledchannels,orpores,ormaybetransportedbycarriers.
B.Proteins1.Integralproteins
are anchored to, and imbedded in, the cell membrane throughhydrophobicinteractions.mayspanthecellmembrane.include ion channels, transport proteins, receptors, andguanosine5′-triphosphate(GTP)–bindingproteins(Gproteins).
2.Peripheralproteinsarenotimbeddedinthecellmembrane.arenotcovalentlyboundtomembranecomponents.arelooselyattachedtothecellmembranebyelectrostaticinteractions.

C.Intercellularconnections1.Tightjunctions(zonulaoccludens)
aretheattachmentsbetweencells(oftenepithelialcells).may be an intercellular pathway for solutes, depending on the size,charge,andcharacteristicsofthetightjunction.maybe“tight”(impermeable),asintherenaldistaltubule,or“leaky”(permeable),asintherenalproximaltubuleandgallbladder.
2.Gapjunctionsare the attachments between cells that permit intercellularcommunication.for example, permit current flow and electrical coupling betweenmyocardialcells.
II. TRANSPORT ACROSS CELLMEMBRANES(TABLE1.1)
table1.1CharacteristicsofDifferentTypesofTransport
*Oneormoresolutesaretransporteduphill;Na+istransporteddownhill.
A.Simplediffusion1.Characteristicsofsimplediffusion
istheonlyformoftransportthatisnotcarriermediated.

occursdownanelectrochemicalgradient(“downhill”).doesnotrequiremetabolicenergyandthereforeispassive.
2.Diffusioncanbemeasuredusingthefollowingequation:J=−PA(C1−C2)where:J=flux(flow)(mmol/sec)P=permeability(cm/sec)A=area(cm2)C1=concentration1(mmol/L)C2=concentration2(mmol/L)
3.SamplecalculationfordiffusionThe urea concentration of blood is 10 mg/100 mL. The ureaconcentration of proximal tubular fluid is 20 mg/100 mL. If thepermeability to urea is 1 × 10−5 cm/sec and the surface area is 100cm2,whatarethemagnitudeanddirectionoftheureaflux?
Note:Theminus signpreceding thediffusionequation indicates thatthedirectionofflux,orflow,isfromhightolowconcentration.Itcanbe ignored if the higher concentration is called C1 and the lowerconcentrationiscalledC2.Alsonote:1mL=1cm3.
4.PermeabilityisthePintheequationfordiffusion.describestheeasewithwhichasolutediffusesthroughamembrane.dependsonthecharacteristicsofthesoluteandthemembrane.
a.Factorsthatincreasepermeability:↑Oil/waterpartitioncoefficientof thesolute increasessolubility inthelipidofthemembrane.

↓Radius (size) of the solute increases the diffusion coefficient andspeedofdiffusion.↓Membranethicknessdecreasesthediffusiondistance.
b. Small hydrophobic solutes (e.g., O2, CO2) have the highestpermeabilitiesinlipidmembranes.
c.Hydrophilicsolutes(e.g.,Na+,K+)mustcrosscellmembranesthroughwater-filled channels, or pores, or via transporters. If the solute is anion (is charged), then its flux will depend on both the concentrationdifferenceandthepotentialdifferenceacrossthemembrane.
B.Carrier-mediatedtransportincludes facilitated diffusion and primary and secondary activetransport.Thecharacteristicsofcarrier-mediatedtransportare
1. Stereospecificity. For example, D-glucose (the natural isomer) istransported by facilitated diffusion, but the L-isomer is not. Simplediffusion, in contrast, would not distinguish between the two isomersbecauseitdoesnotinvolveacarrier.
2.Saturation.Thetransportrateincreasesastheconcentrationofthesoluteincreases,until thecarriersaresaturated.Thetransportmaximum(Tm)isanalogoustothemaximumvelocity(Vmax)inenzymekinetics.
3.Competition. Structurally related solutes compete for transport sites oncarrier molecules. For example, galactose is a competitive inhibitor ofglucosetransportinthesmallintestine.
C.Facilitateddiffusion1.Characteristicsoffacilitateddiffusion
occurs down an electrochemical gradient (“downhill”), similar tosimplediffusion.doesnotrequiremetabolicenergyandthereforeispassive.ismorerapidthansimplediffusion.iscarriermediatedandthereforeexhibitsstereospecificity,saturation,andcompetition.
2.ExampleoffacilitateddiffusionGlucosetransportinmuscleandadiposecellsis“downhill,”iscarriermediated,andisinhibitedbysugarssuchasgalactose;therefore,itis

categorized as facilitated diffusion. In diabetes mellitus, glucoseuptakebymuscleandadiposecellsisimpairedbecausethecarriersforfacilitateddiffusionofglucoserequireinsulin.
D.Primaryactivetransport1.Characteristicsofprimaryactivetransport
occursagainstanelectrochemicalgradient(“uphill”).requiresdirect input ofmetabolic energy in the form of adenosinetriphosphate(ATP)andthereforeisactive.iscarriermediatedandthereforeexhibitsstereospecificity,saturation,andcompetition.
2.Examplesofprimaryactivetransporta.Na+,K+-ATPase(orNa+–K+pump)incellmembranestransportsNa+
from intracellular to extracellular fluid and K+ from extracellular tointracellular fluid; it maintains low intracellular [Na+] and highintracellular[K+].BothNa+ and K+ are transported against their electrochemicalgradients.EnergyisprovidedfromtheterminalphosphatebondofATP.Theusualstoichiometryis3Na+/2K+.SpecificinhibitorsofNa+,K+-ATPasearethecardiacglycosidedrugsouabainanddigitalis.
b.Ca2+-ATPase(orCa2+pump) in the sarcoplasmic reticulum (SR)orcellmembranestransportsCa2+againstanelectrochemicalgradient.Sarcoplasmic and endoplasmic reticulum Ca2+-ATPase is calledSERCA.
c.H+,K+-ATPase(orprotonpump)ingastricparietalcellsandrenalα-intercalatedcellstransportsH+intothelumen(ofthestomachorrenaltubule)againstitselectrochemicalgradient.Itisinhibitedbyprotonpumpinhibitors,suchasomeprazole.
E.Secondaryactivetransport1.Characteristicsofsecondaryactivetransporta.Thetransportoftwoormoresolutesiscoupled.b.Oneofthesolutes(usuallyNa+)istransported“downhill”andprovides

energyforthe“uphill”transportoftheothersolute(s).c.Metabolic energy isnotprovideddirectlybut indirectly from theNa+
gradientthatismaintainedacrosscellmembranes.Thus,inhibitionofNa+,K+-ATPasewilldecreasetransportofNa+outofthecell,decreasethe transmembrane Na+ gradient, and eventually inhibit secondaryactivetransport.
d.Ifthesolutesmoveinthesamedirectionacrossthecellmembrane,itiscalledcotransportorsymport.ExamplesareNa+-glucosecotransportinthesmallintestineandrenalearlyproximaltubuleandNa+–K+–2Cl–cotransportintherenalthickascendinglimb.
e.Ifthesolutesmoveinoppositedirectionsacrossthecellmembranes,itiscalledcountertransport,exchange,orantiport.ExamplesareNa+-Ca2+exchangeandNa+–H+exchange.
2.ExampleofNa+–glucosecotransport(Figure1.1)a. The carrier for Na+–glucose cotransport is located in the luminal
membraneofintestinalmucosalandrenalproximaltubulecells.b.Glucoseistransported“uphill”;Na+istransported“downhill.”c.Energyisderivedfromthe“downhill”movementofNa+.Theinwardly
directed Na+ gradient is maintained by the Na+–K+ pump on thebasolateral (blood side) membrane. Poisoning the Na+–K+ pumpdecreases the transmembrane Na+ gradient and consequently inhibitsNa+–glucosecotransport.
3.ExampleofNa+–Ca2+countertransportorexchange(Figure1.2)a. Many cell membranes contain a Na+–Ca2+ exchanger that transports
Ca2+“uphill”fromlowintracellular[Ca2+]tohighextracellular[Ca2+].Ca2+andNa+moveinoppositedirectionsacrossthecellmembrane.
b.Theenergyisderivedfromthe“downhill”movementofNa+.Aswithcotransport, the inwardly directed Na+ gradient is maintained by theNa+–K+ pump. Poisoning the Na+–K+ pump therefore inhibits Na+–Ca2+exchange.
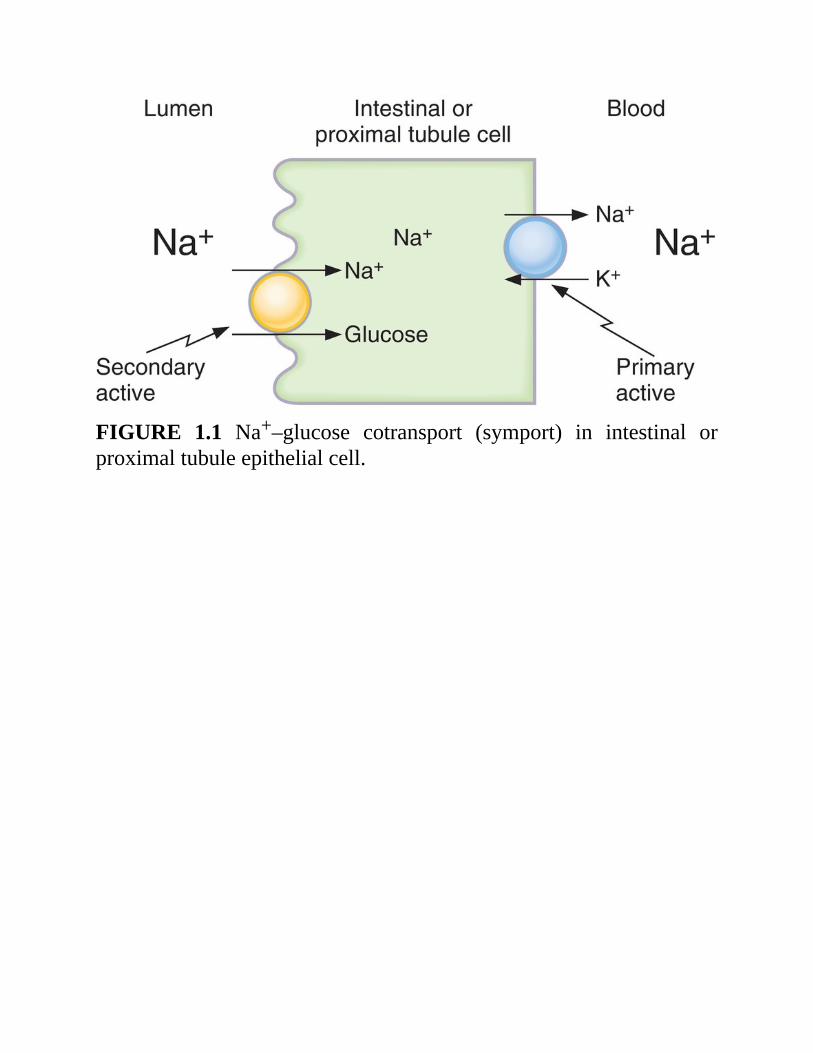
FIGURE 1.1 Na+–glucose cotransport (symport) in intestinal orproximaltubuleepithelialcell.
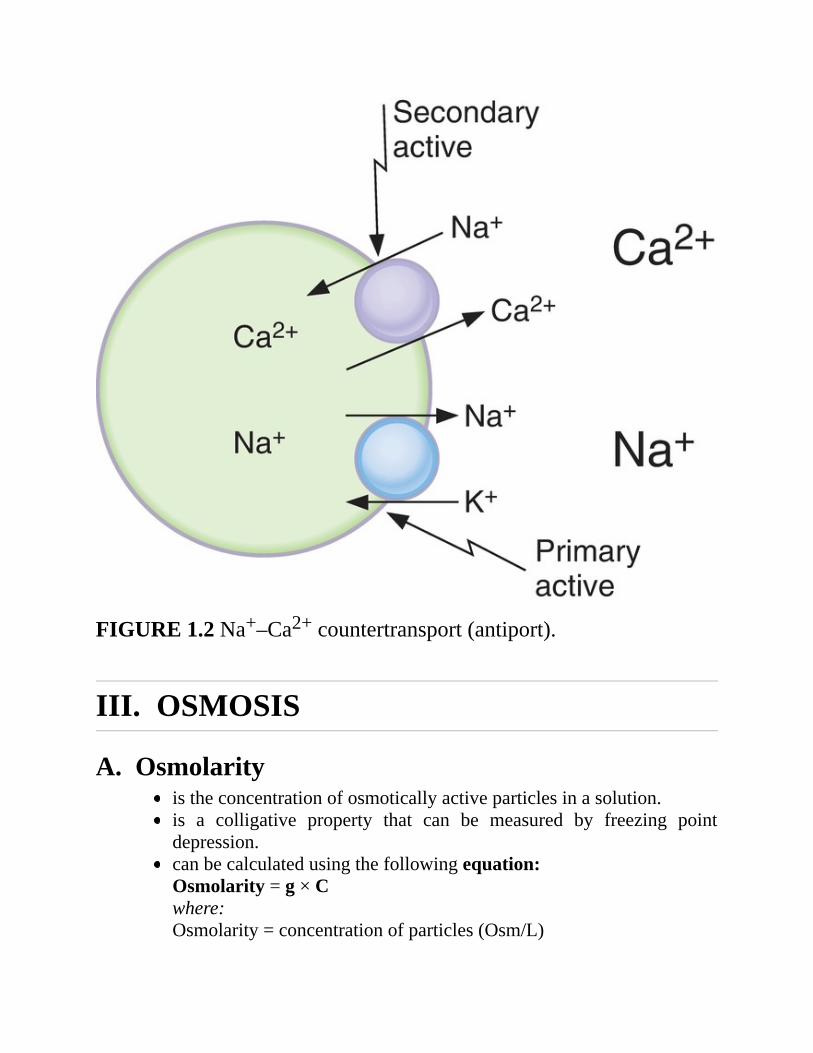
FIGURE1.2Na+–Ca2+countertransport(antiport).
III.OSMOSIS
A.Osmolarityistheconcentrationofosmoticallyactiveparticlesinasolution.is a colligative property that can be measured by freezing pointdepression.canbecalculatedusingthefollowingequation:Osmolarity=g×Cwhere:Osmolarity=concentrationofparticles(Osm/L)

g=numberofparticlesinsolution(Osm/mol)[e.g.,gNaCl=2;gglucose=1]C=concentration(mol/L)Twosolutionsthathavethesamecalculatedosmolarityareisosmotic.If two solutions have different calculated osmolarities, the solutionwiththehigherosmolarityishyperosmoticandthesolutionwiththelowerosmolarityishyposmotic.Samplecalculation:Whatistheosmolarityofa1MNaClsolution?
B.OsmosisandosmoticpressureOsmosisistheflowofwateracrossasemipermeablemembranefromasolutionwithlowsoluteconcentrationtoasolutionwithhighsoluteconcentration.
1.Exampleofosmosis(Figure1.3)a. Solutions 1 and 2 are separated by a semipermeable membrane.
Solution 1 contains a solute that is too large to cross themembrane.Solution 2 is pure water. The presence of the solute in solution 1producesanosmoticpressure.
b.Theosmoticpressuredifferenceacross themembranecauseswater toflow from solution 2 (which has no solute and the lower osmoticpressure) to solution 1 (which has the solute and the higher osmoticpressure).
c. With time, the volume of solution 1 increases and the volume ofsolution2decreases.
2.Calculatingosmoticpressure(van’tHoff’slaw)a.Theosmoticpressureofsolution1(seeFigure1.3)canbecalculated
byvan’tHoff’slaw,whichstatesthatosmoticpressuredependsontheconcentration of osmotically active particles. The concentration ofparticlesisconvertedtopressureaccordingtothefollowingequation:π=g×C×RTwhere:π=osmoticpressure(mmHgoratm)g=numberofparticlesinsolution(osm/mol)C=concentration(mol/L)

R=gasconstant(0.082L—atm/mol—K)T=absolutetemperature(K)
b. The osmotic pressure increases when the solute concentrationincreases.Asolutionof1MCaCl2hasahigherosmoticpressurethandoesasolutionof1MKClbecause,foragivenvolume,thenumberofosmoticallyactiveparticlesishigher.
c.Thehighertheosmoticpressureofasolution,thegreaterthewaterflowintoit.
d.Twosolutionshavingthesameeffectiveosmoticpressureare isotonicbecausenowater flowsacrossa semipermeablemembraneseparatingthem. If two solutions separated by a semipermeablemembranehavedifferent effective osmotic pressures, the solution with the highereffective osmotic pressure is hypertonic and the solution with thelower effective osmotic pressure ishypotonic.Water flows from thehypotonictothehypertonicsolution.
e.Colloidosmoticpressure,oroncoticpressure,istheosmoticpressurecreatedbyproteins(e.g.,plasmaproteins).
3.Reflectioncoefficient(σ)isanumberbetweenzeroandonethatdescribestheeasewithwhichasolutepermeatesamembrane.
a. If the reflection coefficient is one, the solute is impermeable.Therefore, it is retained in the original solution, it creates anosmoticpressure,anditcauseswaterflow.Serumalbumin(alargesolute)hasareflectioncoefficientofnearlyone.
b.Ifthereflectioncoefficientiszero,thesoluteiscompletelypermeable.Therefore, it will not exert any osmotic effect, and it will not causewaterflow.Urea(asmallsolute)usuallyhasareflectioncoefficientofclosetozeroanditis,therefore,anineffectiveosmole.
4.CalculatingeffectiveosmoticpressureEffectiveosmoticpressureistheosmoticpressure(calculatedbyvan’tHoff’slaw)multipliedbythereflectioncoefficient.If the reflection coefficient is one, the solute will exert maximaleffective osmotic pressure. If the reflection coefficient is zero, thesolutewillexertnoosmoticpressure.

FIGURE1.3OsmosisofH2Oacrossasemipermeablemembrane.
IV. DIFFUSION POTENTIAL, RESTINGMEMBRANE POTENTIAL, AND ACTIONPOTENTIAL
A.Ionchannelsareintegralproteinsthatspanthemembraneand,whenopen,permitthepassageofcertainions.
1.Ionchannelsareselective;theypermitthepassageofsomeions,butnotothers.Selectivityisbasedonthesizeofthechannelandthedistributionofchargesthatlineit.
For example, a small channel lined with negatively charged groupswill be selective for small cations and exclude large solutes andanions. Conversely, a small channel lined with positively chargedgroupswillbeselectiveforsmallanionsandexcludelargesolutesandcations.
2. Ion channels may be open or closed. When the channel is open, theion(s) for which it is selective can flow through. When the channel isclosed,ionscannotflowthrough.
3. The conductance of a channel depends on the probability that thechannel is open. The higher the probability that a channel is open, thehighertheconductance,orpermeability.Openingandclosingofchannelsarecontrolledbygates.

a.Voltage-gatedchannelsareopenedorclosedbychangesinmembranepotential.The activation gate of the Na+ channel in nerve is opened bydepolarization.The inactivation gate of the Na+ channel in nerve is closed bydepolarization.Whenboth theactivationand inactivationgatesonNa+ channels areopen, the channels are open and permeable to Na+ (e.g., during theupstrokeofthenerveactionpotential).If either the activation or inactivation gate on the Na+ channel isclosed,thechannelisclosedandimpermeabletoNa+.Forexample,atthe restingpotential, theactivationgatesareclosedand thus theNa+channelsareclosed.
b.Ligand-gated channels are opened or closed by hormones, secondmessengers,orneurotransmitters.For example, the nicotinic receptor for acetylcholine (ACh) at themotor end plate is an ion channel that openswhenACh binds to it.Whenopen,itispermeabletoNa+andK+,causingthemotorendplatetodepolarize.
B.DiffusionandequilibriumpotentialsAdiffusion potential is the potential difference generated across amembranebecauseofaconcentrationdifferenceofanion.A diffusion potential can be generated only if the membrane ispermeabletotheion.The size of the diffusion potential depends on the size of theconcentrationgradient.Thesignofthediffusionpotentialdependsonwhetherthediffusingionispositivelyornegativelycharged.Diffusionpotentialsarecreatedbythediffusionofveryfewionsand,therefore, do not result in changes in concentration of the diffusingions.The equilibrium potential is the potential difference that wouldexactly balance (oppose) the tendency for diffusion down aconcentration difference. At electrochemical equilibrium, thechemicalandelectricaldrivingforcesthatactonanionareequalandopposite,andnofurthernetdiffusionoftheionoccurs.

1.ExampleofaNa+diffusionpotential(Figure1.4)a.TwosolutionsofNaClareseparatedbyamembranethatispermeable
toNa+butnot toCl−.TheNaClconcentrationof solution1 ishigherthanthatofsolution2.
b. Because the membrane is permeable to Na+, Na+ will diffuse fromsolution 1 to solution 2 down its concentration gradient. Cl− isimpermeableandthereforewillnotaccompanyNa+.
c. As a result, adiffusion potential will develop at themembrane andsolution1willbecomenegativewithrespecttosolution2.
d.Eventually,thepotentialdifferencewillbecomelargeenoughtoopposefurther net diffusion of Na+. The potential difference that exactlycounterbalancesthediffusionofNa+downitsconcentrationgradientistheNa+ equilibrium potential. At electrochemical equilibrium, thechemical andelectricaldriving forcesonNa+ are equal andopposite,andthereisnonetdiffusionofNa+.
2.ExampleofaCl-diffusionpotential(Figure1.5)a.TwosolutionsidenticaltothoseshowninFigure1.4arenowseparated
byamembranethatispermeabletoCl−ratherthantoNa+.b.Cl−will diffuse from solution 1 to solution 2 down its concentration
gradient.Na+isimpermeableandthereforewillnotaccompanyCl−.c. Adiffusion potential will be established at the membrane such that
solution1willbecomepositivewithrespecttosolution2.Thepotentialdifference that exactly counterbalances the diffusion of Cl− down itsconcentration gradient is the Cl- equilibrium potential. Atelectrochemicalequilibrium,thechemicalandelectricaldrivingforcesonCl−areequalandopposite,andthereisnonetdiffusionofCl−.
3.UsingtheNernstequationtocalculateequilibriumpotentialsa.TheNernstequationisusedtocalculatetheequilibriumpotentialata
given concentration difference of a permeable ion across a cellmembrane. It tells us what potential would exactly balance thetendencyfordiffusiondowntheconcentrationgradient;inotherwords,atwhatpotentialwouldtheionbeatelectrochemicalequilibrium?
where:

E=equilibriumpotential(mV) z=chargeontheion(+1forNa+,+2forCa2+,−1forCl−)Ci=intracellularconcentration(mM)Ce=extracellularconcentration(mM)
b.SamplecalculationwiththeNernstequationIftheintracellular[Na+] is15mMandtheextracellular[Na+] is150
mM,whatistheequilibriumpotentialforNa+? Note:Youneednotrememberwhichconcentrationgoesinthenumerator.Becauseitisa logfunction,performthecalculationeitherway toget theabsolutevalue of 60 mV. Then use an “intuitive approach” to determine thecorrect sign. (Intuitive approach:The [Na+] is higher in extracellularfluid than in intracellular fluid, so Na+ ions will diffuse fromextracellulartointracellular,makingtheinsideofthecellpositive[i.e.,+60mVatequilibrium].)
c.Approximatevaluesforequilibriumpotentialsinnerveandskeletalmuscle
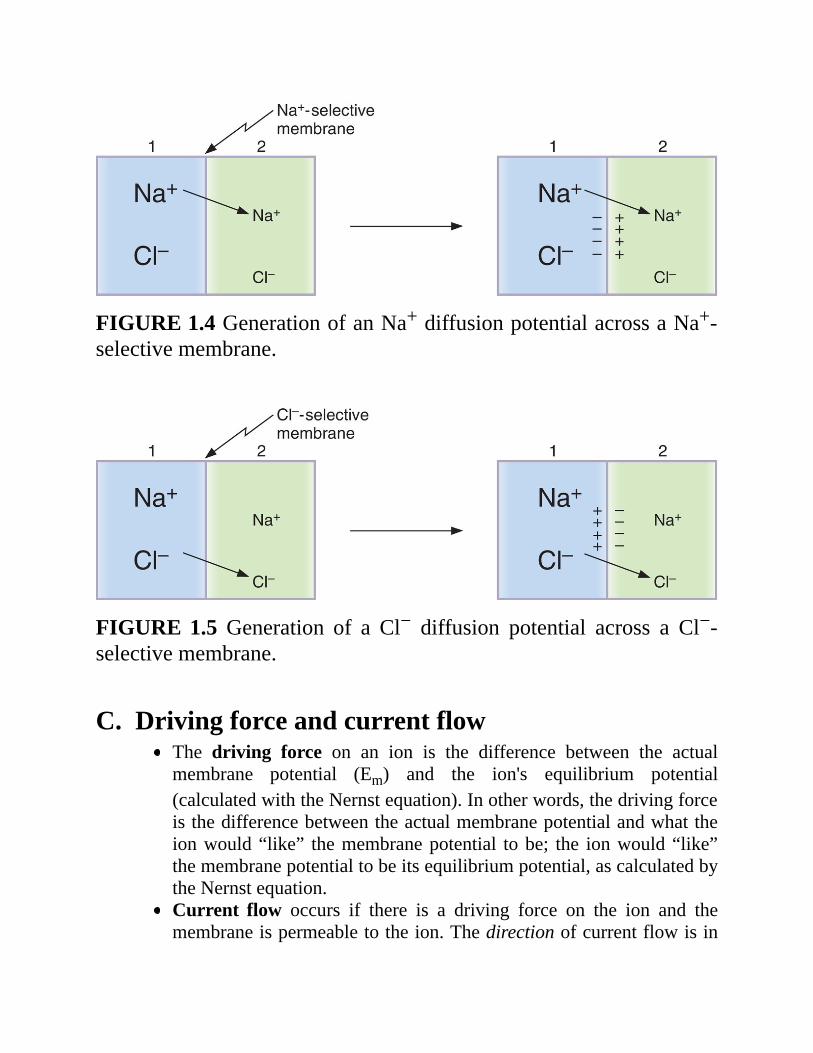
FIGURE1.4GenerationofanNa+diffusionpotentialacrossaNa+-selectivemembrane.
FIGURE1.5 Generation of a Cl− diffusion potential across a Cl−-selectivemembrane.
C.DrivingforceandcurrentflowThe driving force on an ion is the difference between the actualmembrane potential (Em) and the ion's equilibrium potential(calculatedwiththeNernstequation).Inotherwords,thedrivingforceisthedifferencebetweentheactualmembranepotentialandwhattheionwould“like” themembranepotential tobe; the ionwould“like”themembranepotentialtobeitsequilibriumpotential,ascalculatedbytheNernstequation.Current flow occurs if there is a driving force on the ion and themembraneispermeabletotheion.Thedirectionofcurrentflowisin

thesamedirectionasthedrivingforce.Themagnitudeofcurrentflowisdeterminedbythesizeofthedrivingforceandthepermeability(orconductance) of the ion. If there is no driving force on the ion, nocurrentflowcanoccur.Ifthemembraneisimpermeabletotheion,nocurrentflowcanoccur.
D.Restingmembranepotentialis expressed as the measured potential difference across the cellmembraneinmillivolts(mV).is,byconvention,expressedastheintracellularpotentialrelativetotheextracellularpotential.Thus,arestingmembranepotentialof−70mVmeans70mV,cellnegative.
1.Therestingmembranepotentialisestablishedbydiffusionpotentialsthatresultfromconcentrationdifferencesofpermeantions.
2.Eachpermeableionattemptstodrivethemembranepotentialtowardits equilibrium potential. Ions with the highest permeabilities, orconductances, will make the greatest contributions to the restingmembrane potential, and thosewith the lowest permeabilitieswillmakelittleornocontribution.
3.Forexample, therestingmembranepotentialofnerveis−70mV,whichisclosetothecalculatedK+equilibriumpotentialof−85mV,butfarfromthe calculatedNa+ equilibrium potential of +65mV.At rest, the nervemembraneisfarmorepermeabletoK+thantoNa+.
4.TheNa+–K+pumpcontributesonlyindirectlytotherestingmembranepotential by maintaining, across the cell membrane, the Na+ and K+
concentrationgradients that thenproducediffusionpotentials.Thedirectelectrogeniccontributionof thepump(3Na+pumpedoutof thecell forevery2K+pumpedintothecell)issmall.
E.Actionpotentials1.Definitionsa.Depolarizationmakes themembranepotential lessnegative (the cell
interiorbecomeslessnegative).b.Hyperpolarizationmakesthemembranepotentialmorenegative (the
cellinteriorbecomesmorenegative).c. Inward current is the flow of positive charge into the cell. Inward

currentdepolarizesthemembranepotential.d. Outward current is the flow of positive charge out of the cell.
Outwardcurrenthyperpolarizesthemembranepotential.e.Actionpotentialisapropertyofexcitablecells(i.e.,nerve,muscle)that
consists of a rapid depolarization, or upstroke, followed byrepolarization of the membrane potential. Action potentials havestereotypicalsizeandshape,arepropagating,andareall-or-none.
f.Threshold is themembrane potential at which the action potential isinevitable. At threshold potential, net inward current becomes largerthan net outward current. The resulting depolarization becomes self-sustainingandgivesrise to theupstrokeof theactionpotential. Ifnetinwardcurrentislessthannetoutwardcurrent,noactionpotentialwilloccur(i.e.,all-or-noneresponse).
2.Ionicbasisofthenerveactionpotential(Figure1.6)a.Restingmembranepotential
isapproximately−70mV,cellnegative.istheresultofthehighrestingconductancetoK+,whichdrivesthemembranepotentialtowardtheK+equilibriumpotential.At rest, although the inactivation gates on Na+ channels are open(having been opened by repolarization from the preceding actionpotential),theactivationgatesonNa+channelsareclosedandthustheNa+channelsareclosedandNa+conductanceislow.
b.Upstrokeoftheactionpotential1.Inwardcurrentdepolarizesthemembranepotentialtothreshold.2.DepolarizationcausesrapidopeningoftheactivationgatesoftheNa+channels. Now, both activation and inactivation gates are open and theNa+conductanceofthemembranepromptlyincreases.
3.TheNa+ conductancebecomeshigher than theK+ conductance, and themembrane potential is driven toward (but does not quite reach) theNa+equilibriumpotentialof+65mV.Thus,therapiddepolarizationduringtheupstrokeiscausedbyaninwardNa+current.
4.Theovershootisthebriefportionatthepeakoftheactionpotentialwhenthemembranepotentialispositive.
5. Tetrodotoxin (TTX) and lidocaine block these voltage-sensitive Na+channelsandabolishactionpotentials.

c.Repolarizationoftheactionpotential1.Depolarizationalso closes the inactivationgatesof theNa+channels(but more slowly than it opens the activation gates). Closure of theinactivation gates results in closure of the Na+ channels, and the Na+conductancereturnstowardzero.
2. Depolarization slowly opens K+ channels and increases K+
conductance to even higher levels than at rest. Tetraethylammonium(TEA)blocksthesevoltage-gatedK+channels.
3.ThecombinedeffectofclosingtheNa+channelsandgreateropeningofthe K+ channels makes the K+ conductance higher than the Na+conductance, and the membrane potential is repolarized. Thus,repolarizationiscausedbyanoutwardK+current.d.Undershoot(hyperpolarizingafterpotential)
TheK+ conductance remains higher than at rest for some time afterclosure of the Na+ channels. During this period, the membranepotentialisdrivenveryclosetotheK+equilibriumpotential.
3.Refractoryperiods(seeFigure1.6)a.Absoluterefractoryperiod
istheperiodduringwhichanotheractionpotentialcannotbeelicited,nomatterhowlargethestimulus.coincideswithalmosttheentiredurationoftheactionpotential.Explanation:RecallthattheinactivationgatesoftheNa+channelsareclosed when the membrane potential is depolarized. They remaincloseduntil repolarizationoccurs.Noactionpotentialcanoccuruntiltheinactivationgatesopen.
b.Relativerefractoryperiodbeginsattheendoftheabsoluterefractoryperiodandcontinuesuntilthemembranepotentialreturnstotherestinglevel.Anactionpotentialcanbeelicitedduring thisperiodonly ifa largerthanusualinwardcurrentisprovided.Explanation: The K+ conductance is higher than at rest, and themembrane potential is closer to the K+ equilibrium potential and,therefore, farther from threshold;more inward current is required tobringthemembranetothreshold.
c.Accommodation

occurswhenthecellmembraneisheldatadepolarizedlevelsuchthatthethresholdpotentialispassedwithoutfiringanactionpotential.occurs because depolarization closes inactivation gates on the Na+channels.is demonstrated in hyperkalemia, in which skeletal musclemembranes are depolarized by the high serum K+ concentration.Although the membrane potential is closer to threshold, actionpotentialsdonotoccurbecauseinactivationgatesonNa+channelsareclosedbydepolarization,causingmuscleweakness.
4.Propagationofactionpotentials(Figure1.7)occursbythespreadoflocalcurrentstoadjacentareasofmembrane,whicharethendepolarizedtothresholdandgenerateactionpotentials.Conductionvelocityisincreasedby:
a.↑fibersize.Increasingthediameterofanervefiberresultsindecreasedinternalresistance;thus,conductionvelocitydownthenerveisfaster.
b.Myelination. Myelin acts as an insulator around nerve axons andincreases conduction velocity. Myelinated nerves exhibit saltatoryconduction because action potentials can be generated only at thenodesofRanvier,where there are gaps in themyelin sheath (Figure1.8).
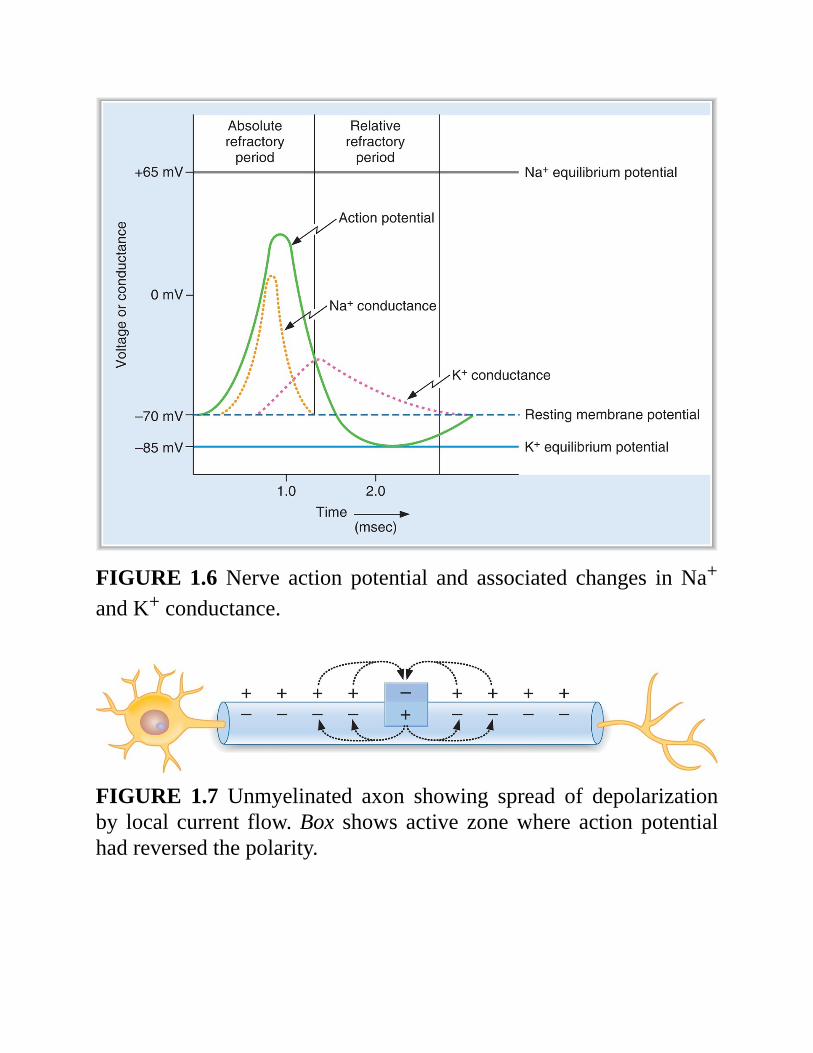
FIGURE1.6Nerve action potential and associated changes inNa+
andK+conductance.
FIGURE 1.7 Unmyelinated axon showing spread of depolarizationby local current flow.Box shows active zonewhere action potentialhadreversedthepolarity.
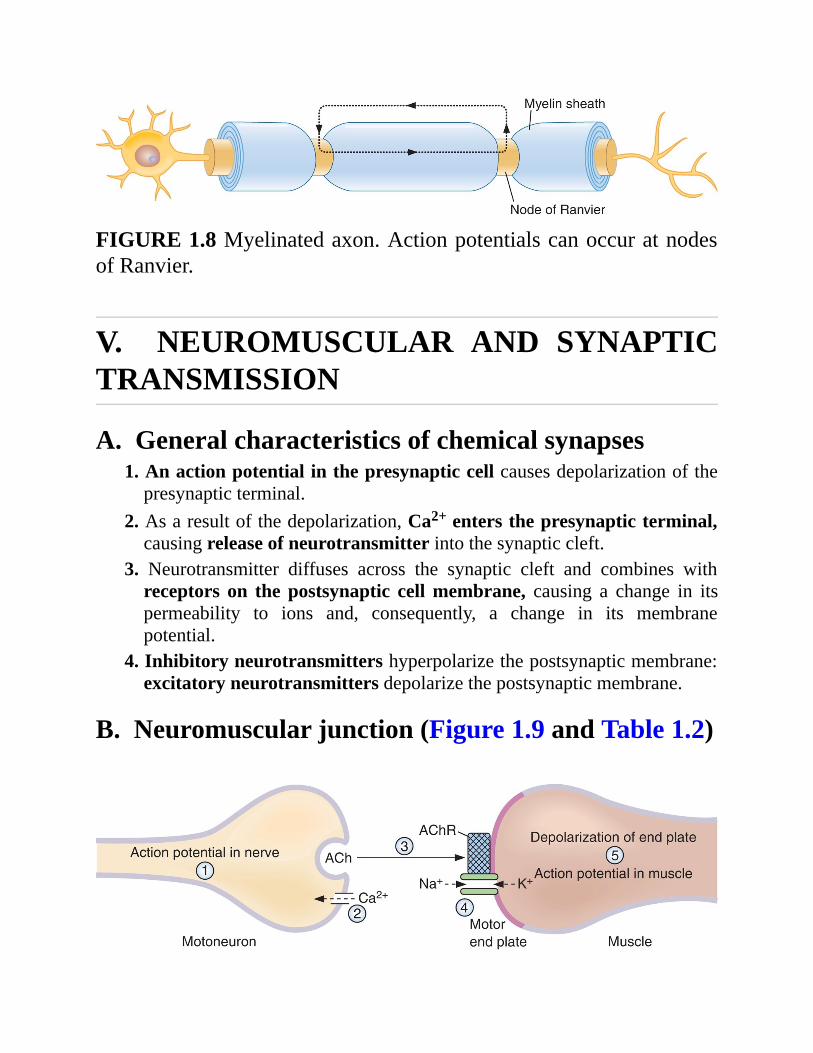
FIGURE1.8Myelinatedaxon.ActionpotentialscanoccuratnodesofRanvier.
V. NEUROMUSCULAR AND SYNAPTICTRANSMISSION
A.Generalcharacteristicsofchemicalsynapses1.Anactionpotentialinthepresynapticcellcausesdepolarizationofthepresynapticterminal.
2.Asaresultofthedepolarization,Ca2+entersthepresynapticterminal,causingreleaseofneurotransmitterintothesynapticcleft.
3. Neurotransmitter diffuses across the synaptic cleft and combines withreceptors on the postsynaptic cellmembrane, causing a change in itspermeability to ions and, consequently, a change in its membranepotential.
4.Inhibitoryneurotransmittershyperpolarizethepostsynapticmembrane:excitatoryneurotransmittersdepolarizethepostsynapticmembrane.
B.Neuromuscularjunction(Figure1.9andTable1.2)

FIGURE1.9Neuromuscularjunction.ACh=acetylcholine;AChR=acetylcholinereceptor.
isthesynapsebetweenaxonsofmotoneuronsandskeletalmuscle.Theneurotransmitter released from thepresynaptic terminal isACh,andthepostsynapticmembranecontainsanicotinicreceptor.
1.SynthesisandstorageofAChinthepresynapticterminalCholineacetyltransferasecatalyzestheformationofAChfromacetylcoenzymeA(CoA)andcholineinthepresynapticterminal.AChisstoredinsynapticvesicleswithATPandproteoglycanforlaterrelease.
2.DepolarizationofthepresynapticterminalandCa2+uptakeActionpotentialsareconducteddownthemotoneuron.DepolarizationofthepresynapticterminalopensCa2+channels.WhenCa2+ permeability increases, Ca2+ rushes into the presynapticterminaldownitselectrochemicalgradient.
3.Ca2+uptakecausesreleaseofAChintothesynapticcleftThesynapticvesiclesfusewiththeplasmamembraneandemptytheircontentsintothecleftbyexocytosis.
4. Diffusion of ACh to the postsynapticmembrane (muscle end plate)andbindingofAChtonicotinicreceptors
ThenicotinicAChreceptorisalsoaNa+andK+ionchannel.BindingofAChtoαsubunitsofthereceptorcausesaconformationalchange that opens the central core of the channel and increases itsconductance to Na+ and K+. These are examples of ligand-gatedchannels.
5.Endplatepotential(EPP)inthepostsynapticmembraneBecausethechannelsopenedbyAChconductbothNa+andK+ ions,thepostsynapticmembranepotentialisdepolarizedtoavaluehalfwaybetweentheNa+andK+equilibriumpotentials(approximately0mV).The contents of one synaptic vesicle (one quantum) produce aminiatureendplatepotential(MEPP),thesmallestpossibleEPP.MEPPs summate to produce a full-fledgedEPP.TheEPP is not anactionpotential,butsimplyadepolarizationofthespecializedmuscleendplate.

6.DepolarizationofadjacentmusclemembranetothresholdOnce the end plate region is depolarized, local currents causedepolarization and action potentials in the adjacent muscle tissue.Actionpotentialsinthemusclearefollowedbycontraction.
7.DegradationofAChThe EPP is transient because ACh is degraded to acetyl CoA andcholinebyacetylcholinesterase(AChE)onthemuscleendplate.One-half of the choline is takenback into thepresynaptic endingbyNa+-cholinecotransportandusedtosynthesizenewACh.AChE inhibitors (neostigmine) block the degradation of ACh,prolongitsactionatthemuscleendplate,andincreasethesizeoftheEPP.HemicholiniumblockscholinereuptakeanddepletesthepresynapticendingsofAChstores.
8.Disease—myastheniagravisiscausedbythepresenceofantibodiestotheAChreceptor.ischaracterizedbyskeletalmuscleweaknessandfatigabilityresultingfromareducednumberofAChreceptorsonthemuscleendplate.The size of the EPP is reduced; therefore, it is more difficult todepolarize themusclemembrane to threshold and to produce actionpotentials.Treatment with AChE inhibitors (e.g., neostigmine) prevents thedegradationofAChandprolongstheactionofAChatthemuscleendplate,partiallycompensatingforthereducednumberofreceptors.
table1.2AgentsAffectingNeuromuscularTransmission
ACh=acetylcholine;EPP=endplatepotential.