phylogenetic evaluation of the tropical camptotypus genus-group (hymenoptera: ichneumonidae), with a...
TRANSCRIPT

This article was downloaded by: [University of Saskatchewan Library]On: 05 October 2012, At: 19:27Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Phylogenetic evaluation of thetropical Camptotypus genus-group(Hymenoptera: Ichneumonidae), witha key to the world generaIlari E. Sääksjärvi a , Ian D. Gauld b & Jukka Salo aa The Zoological Museum, Centre for Biodiversity, Departmentof Biology, FIN-20014, University of Turku, Finland E-mail:b The Natural History Museum of London, Cromwell Road,London SW7 5BD, UK E-mail:
Version of record first published: 19 May 2010.
To cite this article: Ilari E. Sääksjärvi, Ian D. Gauld & Jukka Salo (2004): Phylogeneticevaluation of the tropical Camptotypus genus-group (Hymenoptera: Ichneumonidae), with a keyto the world genera , Journal of Natural History, 38:21, 2759-2778
To link to this article: http://dx.doi.org/10.1080/0022293032000140949
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make anyrepresentation that the contents will be complete or accurate or up to date. Theaccuracy of any instructions, formulae, and drug doses should be independentlyverified with primary sources. The publisher shall not be liable for any loss, actions,claims, proceedings, demand, or costs or damages whatsoever or howsoever causedarising directly or indirectly in connection with or arising out of the use of thismaterial.

Phylogenetic evaluation of the tropical Camptotypus genus-group(Hymenoptera: Ichneumonidae), with a key to the world genera
ILARI E. SAAKSJARVI{, IAN D. GAULD{ and JUKKA SALO{
{The Zoological Museum, Centre for Biodiversity,
Department of Biology, FIN-20014, University of Turku, Finland;
e-mail: [email protected]
{The Natural History Museum of London, Cromwell Road,
London SW7 5BD, UK; e-mail: [email protected]
(Accepted 10 May 2003)
The phylogeny of the disjunct pantropical Camptotypus genus-group (Hymenoptera,Ichneumonidae, Pimplinae) was reconstructed using 40 morphological charac-ters.The monophyly of many established genera (Parvipimpla, Clydonium,Odontopimpla) was found to be supported, but Hemipimpla was found to nestwithin Camptotypus and has thus been treated as a junior synonym of it (syn. n.). Agroup of undescribed Neotropical species were found to form a separatemonophyletic group, herein referred to as Genus A. Species of the Neotropicalgenus Zonopimpla formed a basal paraphyletic grade which could not satisfactorilybe resolved into discrete monophyletic units. Rather than subsuming all the morederived genera within one large group, or attempting to erect numerous new andprobably ephemeral genera (for isolated species of Zonopimpla), we propose theretention of this grade-group as a separate genus at present. The host utilizationpatterns of the genus-group, and the biogeography of the genus-group are discussed.The group is hypothesized as having originated and radiated in the Neotropics, asectoparasitoids of weakly concealed lepidopterous hosts. One lineage, Clydonium,has become specialized at attacking hosts in woody galls where it utilizescoleopterous as well as lepidopterous hosts. The Old World genera (Camptotypusand Parvipimpla) comprise a monophyletic group and are suggested to haveoriginated from a single dispersal event across the widening Atlantic. No evidencefor either a southern or northern circum-polar dispersal was observed in thisentirely tropical genus-group. One lineage in the Afrotropical region has switchedfrom attacking lepidopterous hosts, to attacking pre-pupal and pupal polistinevespids.
KEYWORDS: Biogeography, classification, parasitoid, host utilization, pantropical.
Introduction
The Camptotypus genus-group is one of the five generic groups currently
recognized in the tribe Ephialtini within the ichneumonid subfamily Pimplinae
Journal of Natural HistoryISSN 0022-2933 print/ISSN 1464-5262 online # 2004 Taylor & Francis Ltd
http://www.tandf.co.uk/journalsDOI: 10.1080/0022293032000140949
JOURNAL OF NATURAL HISTORY, 2004, 38, 2759–2778
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

(Gauld, 1991; Gauld et al., 2002). The species of this group are usually brightly
coloured and often strikingly patterned ichneumonids, and they are found
throughout all tropical regions of the world, but not elsewhere. Most are
encountered in lowland wet forests, where they can be found flying in sunlit
clearings, often in the company of similarly coloured and generally much more
numerous braconids. As many of these braconids might be distasteful (Buckingham
and Sharkey, 1988) and models for mimicry complexes (Quicke, 1986; Gauld,
1991), we suspect that these pimplines are Batesian mimics of them. Other species
apparently mimic aggressive vespids, such as the Costa Rican Clydonium cabrerai
Gauld that with black body and white-tipped black wings closely resembles
Parachartergus species (Gauld, 1991).The monophyly of the Camptotypus genus-group has long been recognized
(Townes and Townes, 1960) and is attested by the marked reduction of the occipital
carina, which at the most is only present on the lower part of the head. Current
phylogenetic investigation places the Camptotypus genus-group very close to the
Sericopimpla and Ephialtes genus-groups, which together form a derived group with
respect to the Alophosternum and Pseudopimpla genus-groups (Gauld et al., 2002)
thus: Pseudopimpla g-g z (Alophosternum g-g z (Camptotypus g-g z Ephialtes g-g
z Sericopimpla g-g)). As currently defined, the Camptotypus genus-group
comprises five genera: Camptotypus, Parvipimpla, Clydonium, Odontopimpla and
Zonopimpla (including Cenodontis), whilst a sixth taxon, Hemipimpla, has
sometimes been recognized as a separate genus (e.g. by Gupta and Tikar, 1978),
and at other times treated as a subgenus of Camptotypus (e.g. by Baltazar, 1961;
Townes, 1969). Biogeographically, the Camptotypus genus-group is of particular
interest because all species are entirely tropical. Camptotypus, Hemipimpla and
Parvipimpla occur in the Old World and northern Australia, and the remaining
three genera are restricted to tropical America. This is a particularly unusual
distribution pattern for the Ephialtini, as the other genus-groups are well-
represented in temperate habitats, and in cases where they are species-rich in the
tropics, highly derived and separate lineages occur in the Old and New World
(Gauld et al., 2002). For example, in the Ephialtes genus-group, Anastelgis is
restricted to the New World, whilst the structurally similar Xanthophenax only
occurs in the Old World, where both apparently parasitize concealed lepidopterous
borers (Townes, 1969). Similarly, within the Polysphincta group of genera,
Acrotaphus and Hymenoepimecis are endemic New World genera, whereas a
quite separate lineage, Eriostethus, fills a similar niche in the Old World, attacking
orb-web spiders (Townes, 1969; Gauld, 1984b, 1991).
In recent years, as exhaustive collecting has been undertaken in tropical
America, it has become apparent that a plethora of undescribed species of the
Camptotypus genus-group occur in the region. Some of these cannot be placed to
genera, and many other superficially dissimilar species seem to belong in
Zonopimpla, making the monophyly of the genus as it is presently delineated,
improbable. The aims of the present study are to resolve the phylogeny of the
species-groups within the Camptotypus genus-group so as to provide a basis for
more rigorously defining the genera, and to discover the phylogenetic relationships
between Old and New World taxa.
2760 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

Materials and terminology
The specimens examined in this study were borrowed from or deposited in the
following collections: The Natural History Museum, London, UK (BMNH);
Department of Primary Industries, Indooroopilly, Queensland, Australia; Instituto
Nacional de Biodiversidad, Santo Domingo, Costa Rica; The Zoological Museum,
University of Turku, Turku, Finland. The Peruvian specimens of Genus A (see
below), currently deposited at the University of Turku, will be deposited in the
Museum of Natural History of San Marcos, Lima, Peru. The morphological
terminology is mostly that of Gauld (1991).
Taxa
For the purpose of this study a representative of the basal pimpline lineage,
Delomerista mandibularis (Gravenhorst) [collected in Finland] and relatively
unspecialized member of the Ephialtini, Iseropus stercorator (Fabricius) [England]
were used as out-groups (see Wahl and Gauld, 1998 and Gauld et al., 2002 for
preliminary analysis of Pimplinae). Representatives of all described genera in the
Camptotypus genus-group have been included. Clydonium was represented by three
species: C. analuisae Gauld, Ugalde Gomez and Hanson [Costa Rica], C. cabrerai
Gauld [Costa Rica] and C. sp. 1 [Peru] (in BMNH) and the small genus
Odontopimpla by two: O. fasciata (Brulle) [Peru] and O. maxima (Morley) [Peru].
The very large and morphologically heterogeneous genus Zonopimpla was
represented by 12 species that encompass the morphological variation present in
the taxon: Z. aquilari Gauld [Costa Rica], Z. atriceps (Cresson) [El Salvador], Z.
aurae Gauld [Costa Rica], Z. bermudezi Gauld [Costa Rica], Z. carolinae Gauld
[Costa Rica], Z. fasciata (Townes) (the type-species of Cenodontis) [Brazil], Z.
mirandai Gauld [Costa Rica], Z. snortumi Gauld, Ugalde Gomez and Hanson
[Costa Rica], Z. tenorioi Gauld, Ugalde Gomez and Hanson [Costa Rica], Z. sp. 2
[Peru] (in BMNH), Z. sp. 4 [Peru] (in BMNH) and Z. sp. 10 [Peru] (in BMNH).
The Indo-Australian genus Camptotypus was represented by four species: C.
olynthius (Cameron) [India], C. pseudostigmaticus Gupta and Tikar [Solomon Isles],
C. rugosus (DeGeer) [Sarawak] and C. sellatus Kriechbaumer [Australia], and the
small Australian genus, Parvipimpla, was represented by the only described species,
P. petita Gauld [Australia]. Hemipimpla, an Afrotropical group of species, was
represented by three species: H. flavicaput Morley [Uganda], H. sp. 2 [Angola] (in
BMNH) and H. superba (Szepligeti) [Malawi]. In addition we included five
Neotropical species, Genus A 1–5 [Brazil, Colombia, Guyana and Peru] that could
not be placed into any of the described genera.
Characters
The following set of 40 characters was used in the analysis. The presumed
plesiomorphic condition is denoted by a ‘0’ and derived states by other integers.
The data matrix for character distributions is shown in table 1.
(1) Occipital carina: (0) complete; (1) dorsally absent, ventrally present joining
hypostomal carina; (2) completely absent. We treat this as being a
transformation series, 1.0p1.1p1.2.
(2) Mediodorsal notch of occiput: (0) present, more or less in the form of an
inverted V; (1) absent entirely; (2) with lateral swellings present but broadly
separated.
Phylogenetic evaluation of the tropical Camptotypus 2761
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

(3) Anterior margin of pronotum, dorsal view: (0) centrally reflexed; (1)
transverse.
(4) Anterior part of pronotum, lateral view: (0) anteriorly horizontal, shelf-like,
forming a more or less right angle with the hind margin (figures 1, 2); (1)
more or less vertical in entirety (figure 3).
(5) Epomia: (0) complete; (1) present centrally as small, sharp remnant; (2)
absent or its position indicated by an obscure, low rounded swelling.
(6) Mesoscutal sculpture: (0) uniformly closely punctate, the punctures
separated from each other by about their own diameters; (1) uniformly
sparsely punctate, the punctures separated by about three or more times
their own diameters; (2) smooth and without punctures over most of surface
(7) Notauli: (0) distinct on anterior 0.5 of mesoscutum; (1) shallow, present only
anteriorly as a vestige.
(8) Lateral margin of mesoscutum in front of scutellum: (0) narrowly carinate; (1)
rounded, without a carina or ridge.
(9) Epicnemial carina: (0) dorsal end reaching far above level of lower corner
Table 1. Data matrix.
1111111111 2222222222 3333333333 4123456789 0123456789 0123456789 0123456789 0
Delomerista mandibularis 000000000 0000200000 0010000000 0000100000 0Iseropus stercorator 000000000 0000001000 0011103010 0000000000 1Odontopimpla maxima 211022011 1022202110 0011103031 2000100300 0Odontopimpla fasciata 211022011 1021202111 0011120031 2000100300 0Clydonium sp. 1 110022011 0021202110 0001123031 2001020222 0Clydonium analuisae 100022012 0031202100 0101103032 0001000222 0Clydonium cabrerai 100022011 0011202110 0001123030 2001020222 0Genus A sp. 1 110022011 2112202100 0101123031 0000021100 0Genus A sp. 2 110022011 0132202100 0111121031 0000021100 0Genus A sp. 3 110022011 0102202100 0001103031 0000020100 0Genus A sp. 4 110022012 0101202100 0001113031 0000020110 0Genus A sp. 5 110012011 0131202100 0001113031 0000010100 0Zonopimpla tenorioi 200012010 3002202100 0111120031 0000000000 0Zonopimpla snortumi 200012110 0010202100 0111113032 1000000000 0Zonopimpla aquilari 200012010 0010202100 0011103010 0000000100 0Zonopimpla aurae 200012011 0000202100 0001103030 0000010100 0Zonopimpla atriceps 200012011 0002202100 0001123032 0000000000 0Zonopimpla bermudezi 200011010 0000202100 0011102032 0000000000 0Zonopimpla carolinae 200011010 0000202100 0011103032 0000000000 0Zonopimpla mirandai 200012011 0000202100 0001122030 1000000001 0Zonopimpla sp. 2 200011010 0010202100 0111113032 0000000000 0Zonopimpla sp. 4 200012010 0000202100 0001103032 0000000000 0Zonopimpla sp. 10 200012011 0010202100 0001103032 0000000100 0Camptotypus rugosus 111112110 0042112100 0011120111 0110000000 2Camptotypus pseudostigmatus 111112010 0042102100 1011120010 0010000000 1Camptotypus olynthius 111122010 0042112100 0011121121 0010000000 1Camptotypus sellatus 111122010 0042102100 0111120021 0010000000 1Hemipimpla flavicaput 120122010 0003002100 0011100010 0010000000 1Hemipimpla sp. 2 120122010 0003002100 0001100010 0010000000 1Hemipimpla superba 120122010 0003002100 0011100010 0020000000 1Cenodontis fasciatus 200012012 0030202100 0011113031 0000000001 0Parvivipimpla petita 211112012 0032102100 0211113031 0020000000 1
2762 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

of pronotum; (1) dorsal end more or less level with lower corner of
pronotum; (2) laterally entirely absent.
(10) Medioventral part of epicnemial carina: (0) not raised into a conspicuous
flange-like structure; (1) expanded into a flange-like structure: (2) swollen,
tooth-like; (3) forming a pair of flatten lobes.
(11) Epicnemium: (0) without a distinct secondary carina, at most with a minute
tooth present on upper edge; (1) with a distinct secondary carina present
near lower corner of pronotum (figure 13).
(12) Submetapleural carina: (0) complete, anteriorly very sharp and posteriorly
bifurcate with an inner and outer branch; (1) outer branch absent; (2)
present anteriorly, but both branches absent; (3) entirely absent; (4) inner
branch absent.
(13) Pleural carina: (0) complete; (1) represented by sharp groove; (2) more or
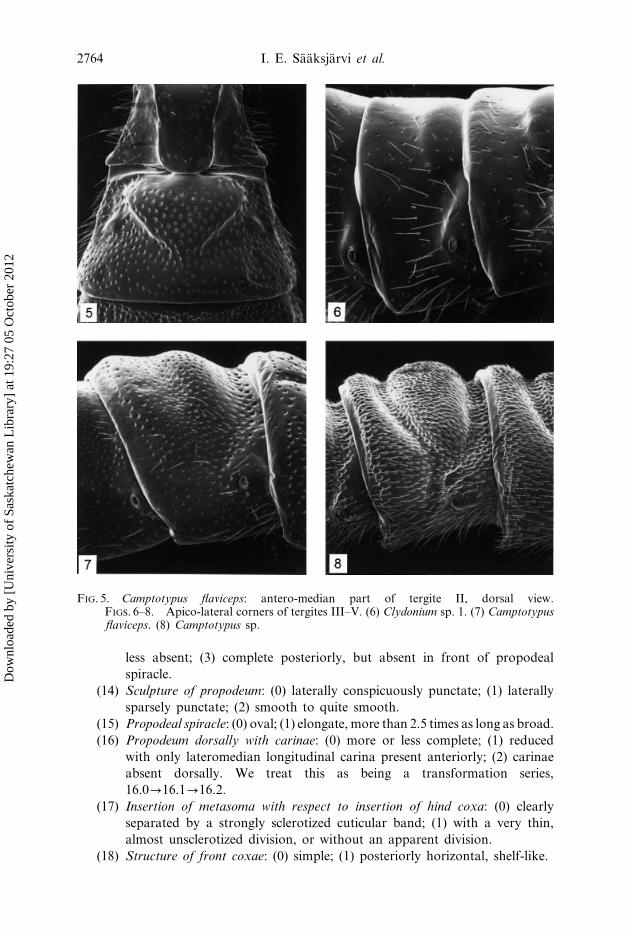
FIGS. 1–3. Anterior part of pronotum, lateral view. (1) Zonopimpla atriceps. (2) Clydoniumsp. 1. (3) Camptotypus flaviceps. FIG. 4. Clydonium sp. 1: antero-median part oftergite II, dorsal view.
Phylogenetic evaluation of the tropical Camptotypus 2763
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2
less absent; (3) complete posteriorly, but absent in front of propodeal
spiracle.
(14) Sculpture of propodeum: (0) laterally conspicuously punctate; (1) laterally
sparsely punctate; (2) smooth to quite smooth.
(15) Propodeal spiracle: (0) oval; (1) elongate, more than 2.5 times as long as broad.
(16) Propodeum dorsally with carinae: (0) more or less complete; (1) reduced
with only lateromedian longitudinal carina present anteriorly; (2) carinae
absent dorsally. We treat this as being a transformation series,
16.0p16.1p16.2.
(17) Insertion of metasoma with respect to insertion of hind coxa: (0) clearly
separated by a strongly sclerotized cuticular band; (1) with a very thin,
almost unsclerotized division, or without an apparent division.
(18) Structure of front coxae: (0) simple; (1) posteriorly horizontal, shelf-like.
FIG. 5. Camptotypus flaviceps: antero-median part of tergite II, dorsal view.FIGS. 6–8. Apico-lateral corners of tergites III–V. (6) Clydonium sp. 1. (7) Camptotypusflaviceps. (8) Camptotypus sp.
2764 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

(19) Hind femur: (0) simple; (1) with a ventral tooth.(20) Pterostigma: (0) normal; (1) conspicuously enlarged.(21) Fore wing with areolet: (0) subrhombic; (1) subtriangular; (2) absent, i.e.
with vein 3rs-m missing.(22) Hind wing with cu-a: (0) short, about 0.3 or less of length of abscissa of Cu1
between M and cu-a ; (1) longer than 0.5 times length of abscissa of Cu1
between M and cu-a.(23) Hind tarsal claw of female with base: (0) simple; (1) lobate.(24) Hind tarsal claw of female: (0) with a small internal membranous vesicle;
(1) without a vesicle.(25) Tergite I with lateromedian carinae: (0) weak, only reaching back to level of
spiracle; (1) absent; (2) very strong, reaching to or past centre of tergite.(26) Tergite I with lateral longitudinal carinae: (0) strong, reaching to or past
centre of tergite, above spiracle; (1) strong, reaching to or past centre of tergite,
below spiracle; (2) weak, reaching back to level of spiracle; (3) absent.
FIGS. 9–12. Apex of lower valve of ovipositor. (9) Zonopimpla atriceps. (10) Odonto-pimpla fasciata. (11) Clydonium sp. 1. (12) Camptotypus flaviceps.
Phylogenetic evaluation of the tropical Camptotypus 2765
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

(27) Sternite II: (0) without prominent flap-like fold; (1) with prominent flap-
like fold.
(28) Tergite II with raised part: (0) coriaceous; (1) closely regularly punctate; (2)
with few scattered punctures centrally; (3) impunctate.
(29) Tergite II with impressed part: (0) closely regularly punctate; (1) with few
scattered punctures centrally; (2) impunctate.
(30) Antero-median part of tergite II: (0) simple (figure 5); (1) with a broad,
shallow indentation; (2) with a strong, deep indentation (figure 4).
(31) Tergites II–V: (0) without a longitudinal carina; (1) with a discontinuous
median longitudinal carina.
(32) Apico-lateral corners of tergites III–V: (0) slightly rounded (figure 6); (1) a
sharp right-angle (figure 7); (2) with a small acute tooth (figure 8). We treat
this as being a transformation series, 32.0p32.1p32.2.
FIG. 13. Genus A sp. 1: epicnemium. FIGS. 14, 15. Apex of lower valve of ovipositor.(14) Genus A sp. 1. (15) Genus A sp. 2.
2766 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

(33) Apical part of ovipositor: (0) subcylindrical; (1) strongly laterally
compressed.(34) Form of upper valve of ovipositor tip: (0) slender; (1) stout and apically
abruptly rounded.(35) Apex of upper valve of ovipositor: (0) simple (figures 9, 10); (1) with low
serrations (figure 11); (2) with clearly discernible teeth (figures 14, 15).(36) Extreme apex of upper valve of ovipositor: (0) simple; (1) with a small apical
swelling (figures 14, 15).(37) Relationship of upper and lower valves of ovipositor: (0) lower valve not
enclosing the upper, and with upper part of the central teeth of lower valve
exposed (figures 9, 12); (1) lower valve with clear lobe partially enclosing
upper valve; (2) lower valve apically more or less entirely enclosing upper
valve (figure 11); (3) upper valve partly enclosing the lower, so dorsal
margins of central teeth of lower valve are obscured (figure 10).(38) Lower valve of ovipositor tip just basal to apex: (0) without scabrous area(s);
(1) with a single long scabrous area proximal to basal tooth; (2) with two
scabrous areas.(39) Apex of lower valve of ovipositor: (0) with distinctly oblique, moderately
interspaced teeth (figures 9, 10); (1) without discernible teeth; (2) with quite
closely interspaced, vertical teeth (figure 11).(40) Surface of ovipositor sheath: (0) with fine short hairs; (1) with long coarse
hairs; (2) with exceptionally dense thick pubescence. We treat this as being
a transformation series, 40.0p40.1p40.2.
Analytical technique
Data were manipulated using Winclada Ver. 0.9.99 (Nixon, 2000). Data were
analysed searching for minimum-length trees under equal character weights using
the NONA program (version 1.6) of Goloboff (1997). NONA was run with all
multistate characters treated as non-additive, except for #1, 16, 32 and 40, all of
which are obviously transformation series. The heuristic search command mult*
(random addition sequence with tree bisection and reconnection) was implemented
with 80 replications, with 12 trees held during each replication (hold/); the resulting
trees were then subjected to branch swapping using tree bisection/reconnection
(max*). The concave function selected was 8. NONA’s successive weighting option
(swt.run) was used to reduce the number of trees.
Results of analysis
Analysis of the data set yielded 92 cladograms with a length of 136 steps, a
consistency index of 0.50, and a retention index of 0.73. Successive weighting
reduced this to three very similar trees differing only in the relationships of the
species of Zonopimpla. One of these is illustrated in figure 16. In the alternative trees
either Z carolinae z Z. bermudezi forms an unresolved trichotomy with Z. snortumi
z Z. sp. 2 and Z. sp. 4z all higher taxa, or Z. carolinae and Z. bermudezi form an
unresolved tetrachotomy with the other taxa.
It can be seen that the Old World species form a strongly supported
monophyletic group (figure 16) which is defined by the following synapomorphies:
anterior part of pronotum in lateral view centrally subvertical (character 4-1);
propodeum laterally sparsely punctate (character 14-1); apico-lateral corners of
tergites III–V angulate (character 32-1). Within this group Parvipimpla is
Phylogenetic evaluation of the tropical Camptotypus 2767
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2
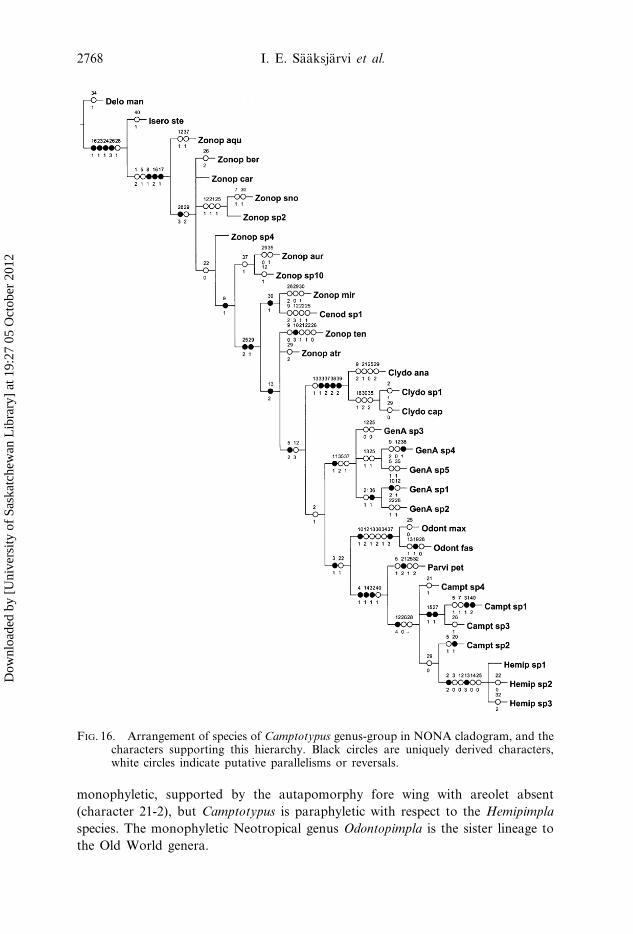
monophyletic, supported by the autapomorphy fore wing with areolet absent
(character 21-2), but Camptotypus is paraphyletic with respect to the Hemipimpla
species. The monophyletic Neotropical genus Odontopimpla is the sister lineage to
the Old World genera.
FIG. 16. Arrangement of species of Camptotypus genus-group in NONA cladogram, and thecharacters supporting this hierarchy. Black circles are uniquely derived characters,white circles indicate putative parallelisms or reversals.
2768 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

The unplaced group of species from South America form a distinct
monophyletic group, which is supported by the presence of a distinct secondary
carina on the epicnemium near lower corner of pronotum (character 11-1). This
group may be the sister lineage to Odontopimpla z Old World genera. The species
of Clydonium form a strongly supported monophyletic group defined by the
following synapomorphies: ovipositor subapically strongly compressed (character
33-1); lower valve of ovipositor apically entirely enclosing upper valve (character
37-2); lower valve of ovipositor tip just basal to apex with two scabrous areas
(character 38-2); apex of lower valve of ovipositor with closely interspaced vertical
teeth (character 39-2). It is the sister lineage to the unplaced group of species from
South America z Odontopimpla z Old World genera.
The major problem remaining concerns the species of Zonopimpla which
together comprise a paraphyletic grade showing little robust structure. Type-species
of Cenodontis (Zonopimpla fasciata) nests amongst the other Zonopimpla species,
supporting the synonymy advocated by Gauld (1991).
Classification of the Camptotypus genus-group
The results of this analysis present a problem which is quite commonly
encountered within the Ichneumonidae, and indeed the Hymenoptera as a whole
(Belshaw et al., 2001). That is, within a clearly definable higher group a number of
discrete monophyletic groups of species have been accorded generic status, whilst
one genus remains paraphyletic. This is known to be the case within the Pimplini
(Ichneumonidae, Pimplinae) where Theronia is a basal paraphyletic assemblage, in
the Glyptini (Ichneumonidae, Banchinae) where Glypta is paraphyletic, and within
the Atrophini (Ichneumonidae, Banchinae) where Lissonota is paraphyletic (Gauld,
1991, 2002). A similar situation almost certainly exists in most other groups of
ichneumonids, and probably most families of Hymenoptera.
Establishing a formal classification for groups such as the Camptotypus genus-
group presents a real problem. Although we accept that strictly monophyletic
groups are desirable, the only way this could be achieved in the present case is
either to include the entire group of species into a single genus, or to erect
numerous new genera for various species of Zonopimpla. The disadvantages of
either approach are obvious. First, creating numerous new genera for species of
Zonopimpla at present is unlikely to produce nomenclatural stability, because the
complex includes a large number of rarely encountered species that are difficult to
group in any meaningful way. Obvious species-groups have not been found and
any proposal of new genera would almost certainly have to be revised repeatedly
because numerous additional species are being discovered all the time as the
fauna of the Andean–Amazonian interface becomes better known. Many of
these species are likely to comprise additional separate lineages that ultimately
would require new genera or blur the distinction between newly postulated ones.
Second, amalgamating all the currently recognized genera would result in a loss
of biological meaning. For example, the very distinctive form of the ovipositor of
the Clydonium species is an adaptation for penetrating tough plant tissue to oviposit
on to hosts living deeply concealed inside the plant (Gauld, 1991). Placing
Clydonium into the same genus together with other species of the Camptotypus
genus-group would make the use and prediction of biological information more
difficult.
We propose a compromise classification. Hemipimpla is here treated as a
Phylogenetic evaluation of the tropical Camptotypus 2769
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

synonym of Camptotypus eliminating one incidence of paraphyly and recognizing a
single distinctive genus for most Old World species. Parvipimpla, Odontopimpla and
Clydonium are treated as distinct genera, as is the group of species (Genus A sp. 1–
sp. 5) from South America. This new genus has been lately described formally in
another paper that also delineates all the species (Saaksjarvi et al., 2003). We
continue to recognize Zonopimpla as a plesiomorphic basal genus, at least until such
time as the overwhelming majority of species are described, when a more exhaustive
analysis might reveal clear groups of species that could be accorded separate generic
status. Details of the classification proposed, together with a key to genera and
diagnoses of genera is given in Appendices 1 and 2.
The biology and patterns of host-utilization by the Camptotypus genus-group
Little is known about the biology of the Camptotypus genus-group. From
information available (Sonan, 1930; Seyrig, 1932; Beeson and Chatterjee, 1935;
Costa Lima, 1945; LePelley, 1954; Keeping and Crewe, 1983; Brooks and Wahl,
1987; Gauld, 1991) it is apparent that the species are idiobiont ectoparasitoids of
the pupae and prepupae of holometabolous insects. Because many of the more
basal taxa in closely related groups utilize hosts concealed in plant tissue—including
Scambus species in the Ephialtes genus-group (Fitton et al., 1988), Pseudopimpla in
the Pseudopimpla genus-group (Bruzzese, 1982; Matsumoto and Saigusa, 1999),
Alophosternum in the Alophosternum genus-group (Cushman, 1933) and Perithous
in the Delomeristini (Danks, 1971)—we suspect that the group may have originated
attacking similar hosts. Although it is a less parsimonious assumption, the
alternative scenario that the ancestors of the group arose attacking cocooned hosts,
cannot be entirely discounted. This is because some basal taxa in related groups—
including Delomerista in the Delomeristini (Furniss and Dowden, 1941) and
Acropimpla and Iseropus in the Sericopimpla genus-group (Fitton et al., 1988)—only
attack such hosts. Both habits occur widely in the Camptotypus genus-group,
although species of one genus, Clydonium, are apparently specialists in attacking
hosts in rather woody galls (Costa Lima, 1945; Gauld, 1991).
The majority of host records for the group are from immature stages of
Lepidoptera. Species of Zonopimpla, Clydonium and Camptotypus all are known to
parasitize such hosts, an observation that suggests Lepidoptera may be the host
group that the Camptotypus lineage diversified using. However, as with many other
idiobiont ectoparasitoids, ordinal-level host switching has occurred. We speculate
that some species of Clydonium have switched from attacking lepidopterous larvae
in galls to attacking gall-forming curculionid larvae (Costa Lima, 1945), and a few
species of Afrotropical Camptotypus have probably switched from Lepidoptera to
attacking the prepupae or pupae of species of polistine Vespidae, such as
Belonogaster and Rhopalidia, in paper nests (Keeping and Crewe, 1983; Brooks and
Wahl, 1987).
The biogeography of the Camptotypus genus-groupWhen trying to understand the biogeographic relationships between Old and
New World species-groups of the Camptotypus genus-group, several things must be
taken into account:
(1) Without exception, all basal lineages (figure 16) are present only in the
Neotropical region and the generic and species richness of the group is
2770 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

highest in New World tropics, where four genera are present (Genus A,
Clydonium, Odontopimpla and ‘Zonopimpla’), which together probably
include more than 100 species.(2) The entire modern Camptotypus genus-group is tropical. There is no
evidence any species has ever spread into adjacent subtropical areas, and
unlike many other predominantly tropical American Pimplinae (such as
Acrotaphus and Anastelgis), none are present in southern USA (vide Townes
and Townes, 1960).(3) The group has a disjunct (non-contiguous) distribution. Four genera are
restricted to the New World tropics, while two genera occur in the Old
World (Camptotypus and Parvipimpla), where they probably comprise fewer
than 50 species.(4) The Old World species form a strongly monophyletic group, and are derived
with respect to the Neotropical fauna (figure 16).
These data suggest that the group originated and primarily radiated in what is
today tropical America. Because the more basal groups of Pimplinae are
predominantly northern (Gauld et al., 2002), we speculate that the ancestor of
the Camptotypus group may have reached South America from North America
during the late Mesozoic or early Paleogenic, by dispersal across the putative
archipelagic connection (Coney, 1982). This Neotropical origin scenario poses the
question of how to explain the disjunct distribution of the group.
Although there is evidence that the family Ichneumonidae existed at the time of
the break-up of Gondwanaland (see discussion in Gauld and Wahl, 2002), we
doubt that the disjunct distribution pattern of the Camptotypus genus-group
resulted from a vicariance event, the separation of Africa from South America. If
such were the case, one might expect to find several lineages of the genus-group
present in both the Old World and the New—just as one finds many lineages of the
Labeninae (a putative southern vicariant group) present in both Australia and
South America (Gauld and Wahl, 2000). The most plausible and parsimonious
explanation of the phylogenetic pattern observed is that the Old World fauna
results from a single dispersal event from tropical America. But how was such
dispersal accomplished?
As nothing has been published on the phylogenetic differentiation rates of
Ichneumonidae, it is difficult to formally link the projected dispersal event of
the genus-group with the New World discrete faunal units (Marshall and Sempere,
1993) and the suggested dispersion routes. We consider the southern Paleogenic
dispersal (sensu Axelrod and Raven, 1978) of the Camptotypus genus-group to
be improbable because the group is absent from temperate South America as
well as from temperate Australia. The groups of ichneumonids that are believed
to have originated in and/or dispersed across Gondwanaland, such as the
Labeninae (Gauld and Wahl, 2000) and Eucerotinae (Gauld and Wahl, 2002),
have basal species present today in the temperate parts of South America and
Australasia. Furthermore, in the Australasian region, such taxa usually show a
drastic reduction in species richness westwards across Wallacea (Whitmore, 1981),
or are even restricted to the Australian tectonic plate (Gauld, 1984a; see also
Crisci et al., 1991 for many other examples of southern distributions). This is
not the case in Camptotypus. All basal taxa are tropical American species
(figure 16), and the genus-group is well-represented in the continental Old
Phylogenetic evaluation of the tropical Camptotypus 2771
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

World tropics but shows a marked decrease in species richness east of Wallace’s
line (Gupta and Tikar, 1978). Furthermore, the Paleogene–Neogene southern realm
has most probably always remained cold-temperate (Coetzee, 1993), without
evident tropical corridors providing an avenue for the dispersal of the Camptotypus
genus-group.
A Neogene northern dispersal route (via North America and across the Bering
Strait) also appears to be improbable for Camptotypus. Although there is a
considerable body of evidence showing that the climate of the circum-arctic region
was far warmer in the past than it is at present, and that at various times between
the middle Eocene and late Pliocene the region supported a rich plant and mammal
biota (McKenna, 1980; Hsu, 1983; Funder et al., 1985; Woodburne and Swisher,
1995), groups of ichneumonids that have putatively dispersed across this bridge are
mainly cool-tolerant taxa. Even amongst predominantly tropical groups which
putatively dispersed across Beringia (such as the Thyreodon genus-group), one
today finds a few species present as far as 50‡N in Asia and in North America
(Gauld, 1985). There are no modern representatives of the Camptotypus genus-
group in either the Eastern Palaearctic or in North America.
The most plausible scenario seems to be that the group dispersed from tropical
South America to Old World tropics via an archipelagic connection in the
spreading Atlantic. In this, the Camptotypus genus-group may resemble the paper
wasps (Polistinae) that also are absent from southern South America and are
considered to have dispersed via tropical South America–tropical Africa before the
full formation of the Mid-Atlantic Seaway (Carpenter, 1993). It is noteworthy that
Camptotypus is one of the few pimpline genera that is apparently good at dispersing
across water into archipelagos, as today one highly derived species (C.
pseudostigmaticus) is endemic to the Solomon Islands.
Appendix 1. Key to the genera of the Camptotypus genus-group
Using the key below it should be possible to place virtually all specimens of the
Camptotypus genus-group into one or other genus. However, it should be noted
that many of the most reliable characters for separating the genera are found in the
structure of the ovipositor. Whenever possible the identification of the males should
be checked by comparing the specimens to females.
1 Anterior part of pronotum in lateral view centrally vertical, not shelf-like (figure 3);Old World species . . . . . . . . . . . . . . . . . . . . 2
– Anterior part of pronotum in lateral view centrally horizontal, shelf-like (figures 1, 2);New World species . . . . . . . . . . . . . . . . . . . . 3
2 Occipital carina present ventrally, as a short vestige joining hypostomal carina; epicne-mial carina present; fore wing with 3rs-m present . . . . . . . Camptotypus
– Occipital carina entirely absent; epicnemial carina absent; fore wing with 3rs-mabsent . . . . . . . . . . . . . . . . . . . . Parvipimpla
3 Anterior margin of pronotum mediodorsally transverse or slightly reflexed but thenwithout a distinct backwardly direct point present; upper valve of ovipositor unusuallystout, towards the apex with its lower margin partly overhanging lower valve, so thatdorsal margins of central teeth of lower valve are obscured (figure 10) Odontopimpla
– Anterior margin of pronotum mediodorsally reflexed, forming a small backwardlydirected point; upper valve of ovipositor not unusually stout, never partly enclos-ing the lower, so that the dorsal margins of the central teeth of the lower valveare not occluded (figures 9, 11, 12) . . . . . . . . . . . . . . 4
2772 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

4 Epicnemium with a distinct secondary carina (figure 13), and upper valve of ovipo-sitor not distinctly enclosed by lower valve (figures 14, 15) . . . . . Genus A
– Epicnemium without a distinct secondary carina, or if with trace of a secondary carina
then upper valve of ovipositor more or less entirely enclosed by lower valve
(figure 11) . . . . . . . . . . . . . . . . . . . . . . 5
5 Pronotum, in lateral view, with epomia present as small sharp remnant in centre;tergite II mediodorsally simple, without a sharp indentation (cf. figure 5); ovipositorapically subcylindrical, lower valve either not laterally expanded to enclose the uppervalve (figure 9) or with a small lobe partially enclosing upper valve Zonopimpla
– Pronotum, in lateral view, with epomia vestigial or absent; tergite II mediodor-
sally usually with a sharp indentation (figure 4); ovipositor conspicuously com-
pressed, the lower valve of ovipositor more or less entirely enclosing the upper valve
(figure 11) . . . . . . . . . . . . . . . . . . . . Clydonium
Appendix 2. Classification of the Camptotypus genus-group
Genus Camptotypus Kriechbaumer
Camptotypus Kriechbaumer, 1889: 311. Type-species: Camptotypus sellatus
Kriechbaumer, by subsequent designation, Viereck, 1914: 27.
Hemipimpla Saussure, 1892: 13. Type-species: Hemipimpla pulchripennis Saussaure,
by monotypy. Syn. n.
Erythropimpla Ashmead, 1900: 57. Type-species: Erythropimpla abbottii Ashmead
(~Pimpla olynthia Cameron), by original designation.Trichiothecus Cameron, 1903: 136. Type-species: Trichiothecus ruficeps Cameron
(~Ichneumon rugosus DeGeer), by monotypy.
Cosmiopimpla Cameron, 1905: 198. Type-species: Cosmiopimpla ferruginea
Cameron, by monotypy.
Pleurodontoplax Enderlein, 1914: 228. Type-species: Pleurodontoplax congoensis
Enderlein (~Pimpla vipioides Brulle), by original designation.
Phruropimpla Benoit, 1964: 387. Type-species: Hemipimpla flavicaput Morley, by
original designation.
Diagnosis. Camptotypus can be easily separated from the New World genera by
the form of the anterior part of pronotum which is in lateral view centrally more or
less vertical. In this feature it resembles the other Old World genus, Parvipimpla,
but unlike this genus all species of Camptotypus have 3rs-m present in the fore wing.Remarks. Camptotypus is a moderately large genus containing 47 described spe-
cies in the Indo-Australian and Afrotropical regions (Yu and Horstmann, 1997) and
a rather few undescribed species in museum collections. The Indo-Australian species
have been revised by Gupta and Tikar (1978), but there is no modern treatment of
the Afrotropical species. There are few reliable host records for species of this
genus, but what few there are suggests species attack cocooned pupae of Macrolepi-
doptera (e.g. Sonan, 1930; Beeson and Chatterjee, 1935; LePelley, 1954). One puta-
tively monophyletic Afrotropical group attack the prepupae or pupae of polistine
Vespidae (Keeping and Crewe, 1983; Brooks and Wahl, 1987).
Genus Clydonium Townes
Clydonium Townes in Townes and Townes, 1966: 324. Type-species: Hemipimpla
recta Morley, by original designation.
Phylogenetic evaluation of the tropical Camptotypus 2773
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

Diagnosis. Clydonium can be identified by the following set of characters: occi-
pital carina present ventrally, joining the hypostomal carina; epomia vestigial or
absent; ovipositor subapically strongly compressed; upper valve of ovipositor
with subapical serrations; lower valve of ovipositor apically entirely or more or
less enclosing upper valve; lower valve of ovipositor tip just basal to apex with
two scabrous areas; apex of lower valve of ovipositor with closely interspaced
vertical teeth; tergite II usually with a median indentation.Remarks. Clydonium is a rather small Neotropical genus comprising 10
described species (Yu and Horstmann, 1997; Gauld et al., 1998) and a similar
number of undescribed species in museums. Most species are very rarely col-
lected. Recorded hosts are woody gall-forming Thryididae and Curculionidae
(Costa Lima, 1945; Gauld, 1991).
Genus Odontopimpla Cameron
Odontopimpla Cameron, 1886: 272. Type-species: Pimpla pulcherrima Cresson
(~Pimpla fasciata Brulle), by monotypy.
Diagnosis. Odontopimpla is characterized by the following set of characters:
occipital carina entirely absent ventrally; epomia discernible as a vestige; upper
valve of ovipositor not compressed and without serrations; lower valve of ovipo-
sitor apically partly enclosed by the upper valve; lower valve of ovipositor with-
out scabrous areas; apex of lower valve of ovipositor with strong and widely
spaced oblique teeth; tergite II with a weak median indentation.Remarks. Odontopimpla is a very small Neotropical genus containing only two
described species, the type species and O. maxima (Morley). Nothing is known
about the biology of either.
Genus Parvipimpla Gauld
Parvipimpla Gauld, 1984a: 67. Type-species: Parvipimpla minuta Gauld, by original
designation.
Diagnosis. Parvipimpla is easily discernible from other genera of the Campto-
typus genus-group by the absence of the areolet in the fore wing.Remarks. This very small genus is represented only by one described species,
Parvipimpla minuta Gauld, occurring in tropical Australia. Nothing is known
about its biology.
Genus ‘Zonopimpla’ Ashmead
Zonopimpla Ashmead, 1900: 55. Type-species: Zonopimpla albicincta Ashmead, by
original designation.
Cenodontis Townes, 1969: 94. Type-species: Cenodontis fasciata Townes, by original
designation.
Diagnosis. Zonopimpla can be distinguished from other genera of the Campto-
typus genus-group by using the following set of characters: occipital carina
entirely absent ventrally; epomia distinct but short; upper valve of ovipositor
subcylindrical to weakly compressed and usually without serrations; lower valve
of ovipositor apically not or only partly enclosing the upper valve; lower valve
2774 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

of ovipositor without scabrous areas; apex of lower valve of ovipositor with
weak; strong and oblique; or sparse and subvertical teeth; or with teeth absent;
tergite II without median indentation.Remarks. Zonopimpla is a large Neotropical genus containing about 30
described species (Yu and Horstmann, 1997; Gauld et al., 1998) and rather large
numbers of undescribed ones. Many of the species are rather infrequently col-
lected, and even in areas that have been intensively sampled, such as Costa
Rica, many new species have been found by additional collecting (compare
Gauld, 1991 and Gauld et al., 1998). We expect many additional species to be
discovered as the fauna of the Amazonian region is better sampled. Such an
incomplete knowledge of the diversity in this genus precludes us from attempting
to delineate monophyletic groups at present.
Very little is known about the biology of species of this group. One or two
species have been reared from cocooned Lepidopterous hosts, and one species is
known to be gregarious within a large Macrolepidopteran host (Gauld, 1991).
Genus A
Diagnosis. Genus A can be distinguished from other genera of the Camptotypus
genus-group by the following set of characters: occipital carina present ventrally as
a vestige, joining hypostomal carina; epicnemium with a distinct secondary carina
near lower corner of pronotum; ovipositor subapically subcylindrical; apex of upper
valve of ovipositor with low serrations or clearly discernible teeth.Remarks. The genus is restricted to the Amazonian region of South America,
and despite very intensive collecting, it has never been found further north, in
Costa Rica. Many new species found during recent studies made in northern
South America belong to this genus, and we now know of species from Brazil,
Ecuador, French Guyana and Peru. Our colleague, Edgard Palacio, has drawn
our attention to species in Colombia. Given the species richness of this genus,
and the fact that all included species were undescribed, the taxon and its compo-
nent species have been lately described formally in another paper (Saaksjarvi
et al., 2003).
Acknowledgements
We are most grateful to Sondra Ward and Reijo Jussila for their assistance in
preparing the figures. Part of the study was financed by the European
Commission’s Large Scale Facility Programme (travel grant to Natural History
Museum of London for I.E.S.) and Graduate School of Biological Interactions,
Ministry of Education, Finland (grant for I.E.S.). Ministry of Agriculture of Peru
provided the collecting and export permits for the Peruvian material. Samuli
Haataja, Andrew Polaszek, Leif Schulman and anonymous referee gave
constructive comments on the manuscript. Finally we would like to thank all
those Costa Rican and Peruvian parataxonomists, biologists and students for their
most valuable help during the study.
References
ASHMEAD, W. H., 1900, Classification of the ichneumon-flies, or the superfamilyIchneumonoidea, Proceedings of the United States National Museum, 23, 1–220.
AXELROD, D. I. and RAVEN, P. H., 1978, Late Cretaceous and Tertiary vegetation history of
Phylogenetic evaluation of the tropical Camptotypus 2775
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

Africa, in M. J. A. Werger (ed.) Biogeography and Ecology of Southern Africa (TheHague: W. Junk), pp. 77–130.
BALTAZAR, C. R., 1961, The Philippine Pimplini, Poemeniini, Rhyssini, and Xoridini(Hymenoptera, Ichneumonidae, Pimplinae), Monographs of the National Institute ofScience and Technology, 7, 1–130.
BEESON, C. F. C. and CHATTERJEE, S. N., 1935, On the biology of the Ichneumonidae(Hymenoptera), Indian Forest Records, 1, 151–168.
BELSHAW, R., LOPEZ-VAAMONDE, C., DEGERLI, N. and QUICKE, D. L. J., 2001, Paraphyletictaxa and taxonomic chaining: evaluating the classification of braconine wasps(Hymenoptera: Braconidae) using 28S D2-3rDNA sequences and morphologicalcharacters, Biological Journal of the Linnean Society, 73, 411–424.
BENOIT, P. L. G., 1964, Ichneumonidae africains du Naturhistorisches Museums de Vienne(Insecta-Hymenoptera), Annalen des Naturhistorischen Museums in Wien, 67, 377–388.
BROOKS, R. W. and WAHL, D. B., 1987, Biology and mature larva of Hemipimplapulchripennis (Saussure), a parasite of Rhopalidia (Hymenoptera: Ichneumonidae,Vespidae), Journal of the New York Entomological Society, 95, 547–552.
BRUZZESE, E., 1982, Observations on the biology of Pseudopimpla pygidiator Seyrig (Hym.,Ichneumonidae), a parasite of the blackberry stem-mining sawfly Hartigiaalbomaculatus (Stein) (Hym., Cephidae), Entomologist’s Monthly Magazine, 118,249–252.
BUCKINGHAM, G. R. and SHARKEY, M. J., 1988, Abdominal exocrine glands in Braconidae(Hymenoptera) in V. K. Gupta (ed.) Advances in Parasitic Hymenoptera Research(Leiden: Brill), pp. 199–242.
CAMERON, P., 1886, Biologia Centrali-Americana. Insecta. Hymenoptera (FamiliesTenthredinidae-Chrysididae), Vol. 1 (London), pp. 265–288.
CAMERON, P., 1903, Description of new genera and species of Hymenoptera taken by MrRobert Shelford at Sarawak, Borneo, Journal of the Straits Branch of the RoyalAsiatic Society, 39, 89–181.
CAMERON, P., 1905, On some new genera and species of Hymenoptera from CapeColony and Transvaal, Transactions of the South African Philosophical Society, 15,195–257.
CARPENTER, J. A., 1993, Biogeographic patterns in the Vespidae (Hymenoptera): two viewsof Africa and South America, in P. Goldblatt (ed.) Biological Relationships BetweenAfrica and South America (New Haven, CT and London: Yale University Press),pp. 139–155.
COETZEE, J. A., 1993, South American paleofloras in P. Goldblatt (ed.) BiologicalRelationships Between Africa and South America (New Haven, CT and London:Yale University Press), pp. 37–61.
CONEY, P. J., 1982, Plate tectonic constraints on the biogeography of Middle America andthe Caribbean region, Annals of the Missouri Botanical Garden, 69, 432–443.
COSTA LIMA, A. DA, 1945, Sobre dois insetos galicolas (Curculionidae—Cryptorhynchinae eIchneumonidae—Pimplinae), Boletim da Sociedade Brasileira de Agronomia, 8,189–192.
CRISCI, J. V., CIGLIANO, M. M., MORRONE, J. J. and ROIG-JUNENT, S., 1991, Historicalbiogeography of southern South America, Systematic Zoology, 40, 152–171.
CUSHMAN, R. A., 1933, Descriptions of new ichneumon-flies with taxonomic notes,Proceedings of the United States National Museum, 82(2955), 1–16.
DANKS, H. V., 1971, Biology of some stem-nesting aculeate Hymenoptera, Proceedings of theRoyal Entomological Society of London, 122, 323–399.
ENDERLEIN, G., 1914, Hymenoptera IV, Ichneumonidae in W. Michaelsen (ed.) Beitrage zurKenntnis der Land-und Susswasserfauna Deutsch-Sudwestafrikas, Band I (Hamburg),452 pp.
FITTON, M. G., SHAW, M. R. and GAULD, I. D., 1988, Pimpline ichneumon-flies Handbooksfor the Identification of British Insects, 7(1), 1–110.
FUNDER, S., ABRAHAMSEN, N., BENNIKE, O. and FEYLING-HANSSEN, R. W., 1985, ForestedArctic: evidence from North Greenland, Geology, 13, 542–546.
FURNISS, R. L. and DOWDEN, P. B., 1941, Western hemlock sawfly, Neodiprion tsugaeMiddleton, and its parasitoids in Oregon, Journal of Economic Entomology, 34, 46–52.
2776 I. E. Saaksjarvi et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

GAULD, I. D., 1984a, An Introduction to the Ichneumonidae of Australia (London: BritishMuseum (Natural History)), 413 pp.
GAULD, I. D., 1984b, The Pimplinae, Xoridinae, Acaenitinae and Lycorininae of Australia(Hymenoptera: Ichneumonidae), Bulletin of the British Museum (Natural History)(Entomology), 49, 235–339.
GAULD, I. D., 1985, The phylogeny, classification and evolution of parasitic wasps of thesubfamily Ophioninae (Ichneumonidae), Bulletin of the British Museum (NaturalHistory) (Entomology), 51, 61–185.
GAULD, I. D., 1991, The Ichneumonidae of Costa Rica, 1, Memoirs of the AmericanEntomological Institute, 47, 1–589.
GAULD, I. D. (ed.), 2002, The Ichneumonidae of Costa Rica, 4, Memoirs of the AmericanEntomological Institute, 66, 1–768.
GAULD, I. D. and WAHL, D., 2000, The Labeninae (Hymenoptera: Ichneumonidae): a studyin phylogenetic reconstruction and evolutionary biology, Zoological Journal of theLinnean Society, 129, 271–347.
GAULD, I. D. and WAHL, D., 2002, The Eucerotinae: a Gondwanan origin for acosmopolitan group of Ichneumonidae?, Journal of Natural History, 36,2229–2248.
GAULD, I. D., UGALDE GOMEZ, J. A. and HANSON, P., 1998, Guıa de los Pimplinae de CostaRica (Hymenoptera: Ichneumonidae) Revista de Biologia Tropical, 46(1), 1–189.
GAULD, I. D., WAHL, D. and BROAD, G. R., 2002, The suprageneric groups of the Pimplinae(Hymenoptera: Ichneumonidae): a cladistic re-evaluation and evolutionary biologicalstudy, Zoological Journal of the Linnean Society, 136, 421–485.
GOLOBOFF, P., 1997, NONA, version 1.6, MS-DOS program (San Miguel de Tucuman,Argentina: published by the author).
GUPTA, V. K. and TIKAR, D. T., 1978 (1976), Ichneumonologia Orientalis. Part 1. The tribePimplini Oriental Insects Monographs, 1, 1–302.
HSU, J., 1983, Late Cretaceous and Cenozoic vegetation in China, emphasizing theirconnections with North America, Annals of the Missouri Botanic Garden, 70,490–508.
KEEPING, M. G. and CREWE, R. M., 1983, Parasitoids, commensals and colony size in nestsof Belonogaster (Hymenoptera: Vespidae), Journal of the Entomological Society ofSouthern Africa, 46, 309–323.
KRIECHBAUMER, J., 1889, Nova genera et species Pimplidarum, Entomologische Nachrichten,15, 307–312.
LEPELLEY, R. H., 1954, Agricultural Insects of East Africa (Nairobi), 307 pp.MARSHALL, L. G. and SEMPERE, T., 1993, Evolution of the Neotropical Cenozoic land
mammal fauna and its geochronologic, stratigraphic, and tectonic context in P.Goldblatt (ed.) Biological Relationships Between Africa and South America (NewHaven, CT and London: Yale University Press), pp. 329–392.
MATSUMOTO, R. and SAIGUSA, T., 1999, Biology of Pseudopimpla glabripropodeum He andChen (Hymenoptera, Ichneumonidae, Pimplinae) with description of final instar larvaand a redescription of adults Entomological Science, 2, 467–471.
MCKENNA, M. C., 1980, Eocene paleolatitude, climate, and mammals of Ellesmere IslandPalaeogeography, Palaeoclimatology, Palaeoecology, 30, 349–362.
NIXON, K. C., 2000, Winclada, version 0.9.99, documentation and software, http://www.cladistics.com.
QUICKE, D. L. J., 1986, Preliminary notes on homeochromatic associations within andbetween the Afrotropical Braconinae (Hym., Braconidae) and Lamiinae (Col.,Cerambycidae) Entomologist’s Monthly Magazine, 122, 97–109.
SAAKSJARVI, I. E., PALACIO, E., GAULD, I. D., JUSSILA, R. and SALO, J., 2003, A new genusand six new species of the tropical Camptotypus genus-group (Hymenoptera:Ichneumonidae; Pimplinae) from northern South America, Zootaxa, 197, 1–18.
SAUSSURE, H. DE, 1892, Hymenopteres in A. Grandidier (ed.) Histoire Physique Naturelle etPolitique de Madagascar, 20 (Paris), 590 pp.
SEYRIG, A., 1932, Les Ichneumonides de Madagascar, I. Ichneumonidae PimplinaeMemoires de l’Academie Malgache, 11, 1–183.
Phylogenetic evaluation of the tropical Camptotypus 2777
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2

SONAN, J., 1930, A few host known Ichneumonidae found in Japan and Formosa (Hym.),Transactions of the Natural History Society of Formosa, 20, 268–273.
TOWNES, H., 1969, Genera of Ichneumonidae, 1, Memoirs of the American EntomologicalInstitute, 11, 1–300.
TOWNES, H. and TOWNES, M., 1960, Ichneumon-flies of America north of Mexico: 2.Subfamilies Ephialtinae, Xoridinae, Acaenitinae, United States National MuseumBulletin, 216(2), 1–676.
TOWNES, H. and TOWNES, M., 1966, A catalogue and reclassification of NeotropicIchneumonidae, Memoirs of the American Entomological Institute, 8, 1–367.
VIERECK, H. L., 1914, Type species of the genera of Ichneumon flies, Bulletin of the UnitedStates National Museum, 83, 1–186.
WAHL, D. B. and GAULD, I. D., 1998, The cladistics and higher classification of thePimpliformes (Hymenoptera: Ichneumonidae), Systematic Entomology, 23, 265–298.
WHITMORE, T. C. (ed.), 1981, Wallace’s Line and Plate Tectonics (Oxford: Clarendon Press),91 pp.
WOODBURNE, M. O. and SWISHER, C. C., 1995, Land mammal, high resolutiongeochronology, intercontinental overland dispersals, sea-level, climate and vicariancein W. A. Berggren and D. V. Kent (eds) Geochronology, Time Scales and GlobalStratigraphic Correlation (Tulsa, OK: SEPM), pp. 335–364.
YU, D. and HORSTMANN, K., 1997, Catalogue of world Ichneumonidae (Hymenoptera),Memoirs of the American Entomological Institute, 58, 1–1558.
2778 Phylogenetic evaluation of the tropical Camptotypus
Dow
nloa
ded
by [
Uni
vers
ity o
f Sa
skat
chew
an L
ibra
ry]
at 1
9:27
05
Oct
ober
201
2