phycocyanin 645 - journal of biological chemistry
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 258, No. 23, Issue of December 10, pp. 14327-14329,1983 Prrnted in U.S.A.
Phycocyanin 645 THE CHROMOPHORE ASSAY OF PHYCOCYANIN 645 FROM THE CRYPTOMONAD PROTOZOA CHROOMONAS SPECIES*
(Received for publication, May 20, 1983)
Robert MacColl and Deborah Guard-Friar From the Center for Laboratories and Research, New York State Department of Health, Albany, New York 12201
Phycocyanin 645 was isolated and purified from the cryptomonad Chroomonas species. Its chromophore content was obtained from absorption spectra in acidic 8.0 M urea €or both whole protein and the separated subunits. The principal method used to separate the (Y
and /i’ subunits was gel filtration through a Sephacryl S-200 column in acidic urea. The subunits were shown to be completely separated during this procedure by sodium dodecyl sulfate-gel electrophoresis. Spectra were analyzed by three component Beer’s law equa- tions. The whole protein was found to consist of four phycocyanobilins (A,,, at 662 nm), two cryptoviolins (A,,, at 590 nm), and two unnamed bilins with an A,,, at 697 nm. The separated subunits were analyzed, and the /i’ subunit was shown to have two phycocyanobilins for each cryptoviolin and (Y was composed of the 697- nm bilin exclusively. A comparison of the total amounts of (Y and B from the Sephacryl columns showed that the molar ratios of phycocyanobilin on /i’ to the 697-nm bilin on (Y was 2: 1, and the ratio of cryptoviolin on /i’ to 697-nm bilin on (Y was 1:l. W e therefore propose that, assuming a symmetrical distribution, each /i’ subunit on the crZ& protein has two phycocyanobilins and one cryptoviolin and each (Y subunit has one 697-nm bilin.
This chromophore distribution differs from one pre- viously reported in which the subunits were separated on a BioRex 70 cation exchange resin in 12% formic acid via a 4-10 M urea gradient.
Tetrapyrroles are the chromophore moieties of the bilipro- teins, photosynthetic light-harvesting pigments (for bilipro- tein reviews, See Refs. 2, 3, 6, and 7). When all of the types of biliprotein are denatured in acidic urea, a total of five distinct visible absorption bands are observed and each is believed to be produced by a chemically unique tetrapyrrole: two major types, phycocyanobilin and phycoerythrobilin, and three minor types, cryptoviolin, phycourobilin and an unusual bilin with a 697-nm absorption maximum. The analysis of the covalent structures of these less frequently observed bilins has yet to be performed, and it is possible that some of these bilins are quite closely related to each other. Except for phycocyanin 645, all biliproteins have either one type of bilin (e.g. C-phycocyanin, allophycocyanins, C-phycoerythrin, b- phycoerythrin) or two (e.g. R-phycocyanin, R-phycoerythrin, B-phycoerythrin, phycoerythrocyanin, phycoerythrin 545,
*This research was supported by National Institutes of Health Grant GM 26050 awarded by the National Institute of General Medical Science, United States Public Health Service/Department of Health and Human Services. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
phycocyanin 612). Spectral studies (4) on the separated a and p subunits of phycocyanin 645 indicates that it is composed of three bilins: phycocyanobilin, cryptoviolin (also named phycobiliviolin), and a unique bilin with an absorption max- imum at 697 nm (1, 5). We have studied the number, type, and distribution of the chromophores on phycocyanin 645 using both whole protein and separated subunits. The results establish the chromophore content of this protein.
EXPERIMENTAL PROCEDURES AND RESULTS’
DISCUSSION
When the spectrum of four phycocyanobilins is subtracted from the spectrum of phycocyanin 645 in acidic 8.0 M urea, the residue has bands a t 590 and 697 nm (Fig. 1). These bands correspond very closely to those of cryptoviolin (A,,, at 590 nm) and the unusual bilin with a 697-nm absorption maxi- mum. Using Equations 1-3, we demonstrated that there are four phycocyanobilins, two cryptoviolins, and two 697-nm bilins/a& structure.
A chromatographic procedure using Sephacryl ,9200 is
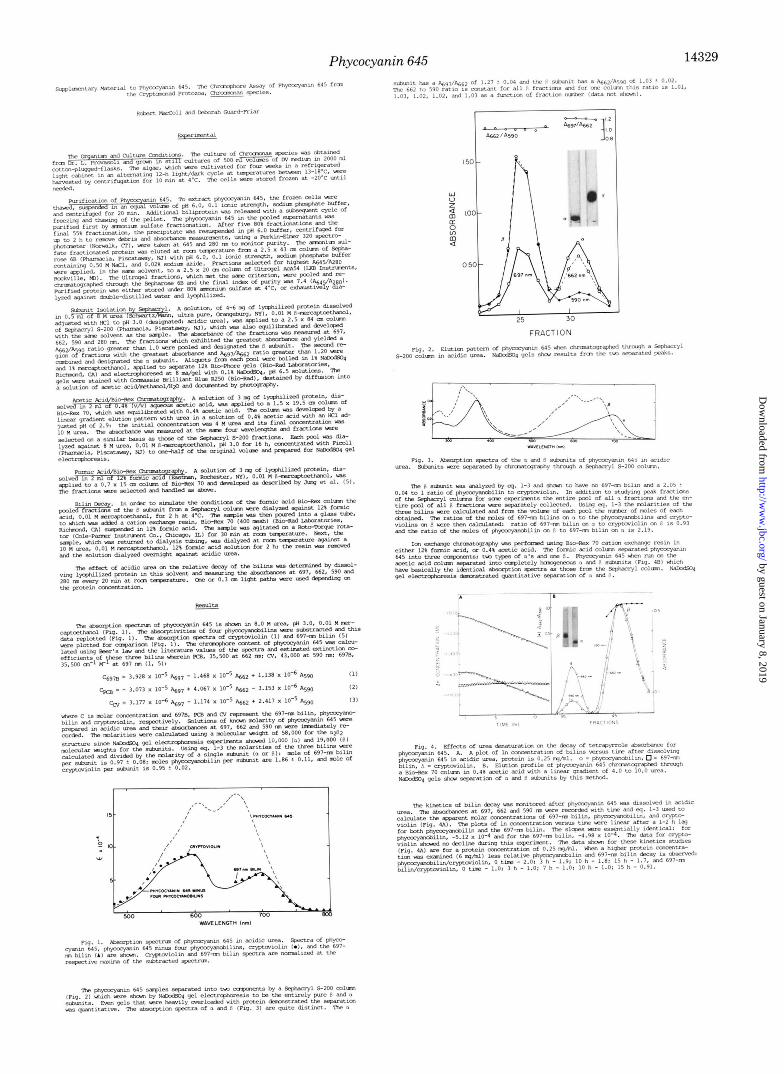
shown by sodium dodecyl sulfate-gel electrophoresis to be entirely effective in separating the a and p subunits of phy- cocyanin 645 (Fig. 2). The solvent selected for this protocol is acidic urea, the identical solvent used for chromophore identification and quantitation. Therefore, the fractions from this column may be directly studied for chromophore content by Equations 1-3. Two peaks are obtained from this column; the faster eluting band is pure p and the slower is pure a (Fig. 3). The (Y subunit is seen to have a spectrum identical with the 697-nm bilin; the Beer’s law analysis shows p is composed of two phycocyanobilins/one cryptoviolin and no 697-nm bilin. Analysis of total pools from these columns demonstrates that the ratio of phycocyanobilin on p to the 697-nm bilin on a is 2:1, and the ratio of cryptoviolin on p to 697-nm bilin on a is 1:l. There is, then, more than sufficient data to construct an unequivocal arrangement of the eight chromophores on the a& structure, assuming symmetry: two phycocyanobilins and one cryptoviolin on each /3 and one 657-nm bilin on each
Jung et al. (5) determined from their spectra of the sepa- rated subunits a saliently different chromophore arrange-
’ Portions of this paper (including “Experimental Procedures,” “Results,” Figs. 1-4, and Equations 1-3) are presented in miniprint at the end of this paper. The abbreviation used is: NaDodSor, sodium dodecyl sulfate. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 83M-1389, cite the authors, and include a check or money order for $4.40 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
a.
14327
by guest on January 8, 2019http://w
ww
.jbc.org/D
ownloaded from

14328 Phycocyanin 645
TABLE I Comparison of the two proposals for the chromophore composition of
phycocyanin 645 with calculations of Equations I 3 using the experimental results from dissolving whole protein in acidic urea
Chromophore ratios
Property Proposed Experimental Proposed model data ~~f~f~~;okm of Jung et al. (5)
Phycocyanobilin/ 2 1.5 2.00 f 0.10
Phycocyanobilin/ 2 3 1.91 -+ 0.25
697-nm bilin/ 1 0.5 1.05 f 0.10
cryptoviolin
697-nm bilin
cryptoviolin
ment. They reported spectra consistent with each p having one phycocyanobilin and one cryptoviolin and one CY having a phycocyanobilin and the second a 697-nm bilin. Using our data in a different way, the two models can be readily com- pared. Instead of directly calculating the actual molar amounts of each bilin, we can simply obtain their relative molar ratios using Equations 1-3. This simplification elimi- nates several sources of experimental error from the calcula- tions: weighing the protein, measuring the volumes, quanti- tative solubility of the protein, and inaccuracy in the molec- ular weight. Therefore, this relative measurement should give a more accurate and precise criterion of the chromophore distribution. Table I shows the bilin ratio from our model, the previous model, and experimental data.
Fig. 4A presents significant information on the use of acidic urea to quantitate bilin ratios. It demonstrates the concept that under these experimental conditions bilins may appar- ently decay a t unequal rates. This, naturally, is most critical when dealing with separated subunits which require a certain amount of time to elute from the chromatographic matrix. This is why we stress the results from whole protein studies where the absorption spectrum can be measured immediately after the lyophilized protein is dissolved in acidic urea. None- theless, the apparent increase of cryptoviolin over both phy- cocyanobilin and the 697-nm bilin is relatively slow at the protein concentrations used on the Sephacryl column (1.6 x 84 cm), and after 6-8 h (the normal run time of our Sephacryl column), cryptoviolin is not sharply enhanced over the other bilins. Thus, our 2:l ratio of phycocyanobilin to cryptoviolin should not need adjustment. Fortunately, the decay of phycocyanobilin and the 697-nm bilin appears to occur a t reasonably similar rates so that their ratios are unaffected. These calculations (Fig. 4A) are for the apparent molar con- centrations of the bilins and do not account for the contri- butions of any reaction products that may arise during the decay. The apparent loss of phycocyanobilin relative to cryp- toviolin appears to be the reason for the lower ratio found by Jung et at. (5) . To test this proposal, a mock column procedure following their formic acid/urea/mercaptoethanol/BioRex 70 protocol was carried out. Homogeneous (3 subunit (from the Sephacryl column) in 12% formic acid is treated first with
BioRex 70 equilibrated with 12% formic acid to which p adheres, then the mixture is dialyzed against 10 M urea in 12% formic acid which liberates it from the resin, and finally dialyzed against acidic urea. After this last dialysis, the ab- sorbances are measured, and the molar ratio of phycocyano- bilin to cryptoviolin dropped about 20% of the value obtained for a control solution. This change is not as large as the true column difference but suggests that the column milieu pro- motes accelerated phycocyanobilin decay. An advantage of the mock protocol is that there is no possibility of chromato- graphic separation being responsible for the altered phycocy- anobilin/cryptoviolin ratio.
The acetic acid column (Fig. 4B) exhibits some heteroge- neity, but the A662/A590 ratio of most of the fractions agrees exactly with the Sephacryl results. These spectral results agree quite nicely with those reported earlier by Glazer and Cohen-Bazire (4), and we have substantiated the significance of this experimental result by presenting sodium dodecyl sulfate gels (Fig. 4B) which prove that this protocol separates the a and p subunits. Utilization of Equations 1-3 with these spectra confirms the 2:l ratio of phycocyanobilin to crypto- violin on p.
A major contribution of the paper by Jung et al. (5) was to demonstrate that there are two distinct CY subunits. Using the formic acid column, we confirm two types of a subunits, but unlike their results we find that both a subunits have the 697-nm bilin. The true differences between a subunits prob- ably reside in their amino acid compositions. The proposal ( 5 ) of one 697-nm bilin and one phycocyanobilin on a would give an &97/&62 ratio of 0.77 when both a subunits are mixed together, while the 697-nm bilin alone should give a ratio of 1.29. Clearly, the Sephacryl experimental value of 1.27 f 0.04 favors the latter. Therefore, data from whole protein and two methods of subunit separation can be resolved into a new chromophore distribution: two phycocyanobilins and one cryptoviolin on each /3 subunit and one 697-nm bilin on each a subunit.
Acknowledgments-We wish to thank William R. Town and Leon H. Wu for their technical contributions.
REFERENCES
1. Bryant, D. A., Glazer, A. N., and Eiserling, F. A. (1976) Arch.
2. Gantt, E. (1981) Annu. Rev. Plant Physiol. 32,327-347 3. Gantt, E. (1979) in Biochemistry and Physiology of Protozoa
(Levandowsky, M., and Hutner, S. A., eds) 2nd Ed., pp. 121- 137, Academic Press, New York
4. Glazer, A. N., and Cohen-Bazire, G. (1975) Arch. Microbiol. 104,
5. Jung, J., Song, P.-S., Paxton, R. J., Edelstein, M. S., Swanson,
6. Scheer, H. (1981) Angew. Chem. Int. Ed. Engl. 20 , 241-261 7. Troxler, R. F. (1977) in Chemistry and Physwlogy of Bile Pigments
(Berk, P. D., and Berlin, N. I., eds) No. 77-1100, pp. 431-454, Department of Health, Education, and Welfare, Washington, D. C.
Microbiol. 110, 61-75
29-32
R., and Hazen, E. E., Jr. (1980) Biochemistry 19, 24-32
by guest on January 8, 2019http://w
ww
.jbc.org/D
ownloaded from
Phycocyanin 645 14329
150 -
w u z 2 100-
2 [L
m 4
050 -
.* 0 -
by guest on January 8, 2019http://w
ww
.jbc.org/D
ownloaded from

R MacColl and D Guard-Friarcryptomonad protozoa Chroomonas species.
Phycocyanin 645. The chromophore assay of phycocyanin 645 from the
1983, 258:14327-14329.J. Biol. Chem.
http://www.jbc.org/content/258/23/14327Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/258/23/14327.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 8, 2019http://w
ww
.jbc.org/D
ownloaded from